Human Recombinant Fab Fragment Neutralizes Shiga Toxin Type 2 Cytotoxic Effects in vitro and in vivo

 , , and
, , and
Abstract
:1. Introduction
2. Results
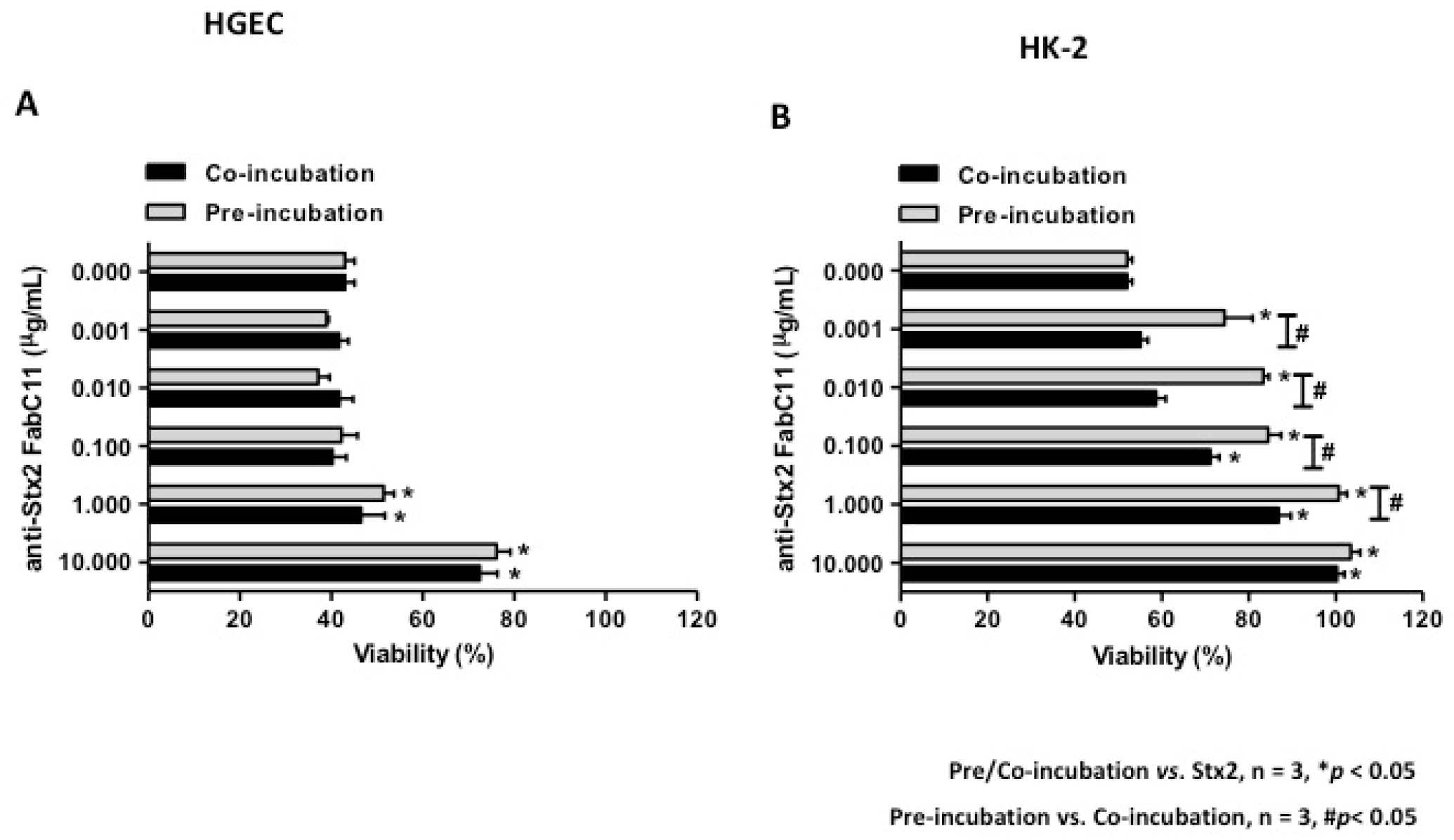
2.1. FabC11:Stx2 Has High Affinity to Stx2 and Protects Human Glomerular Endothelial Cells (HGEC) and Human Kidney (HK-2) Cells from Stx2 Cytotoxicity
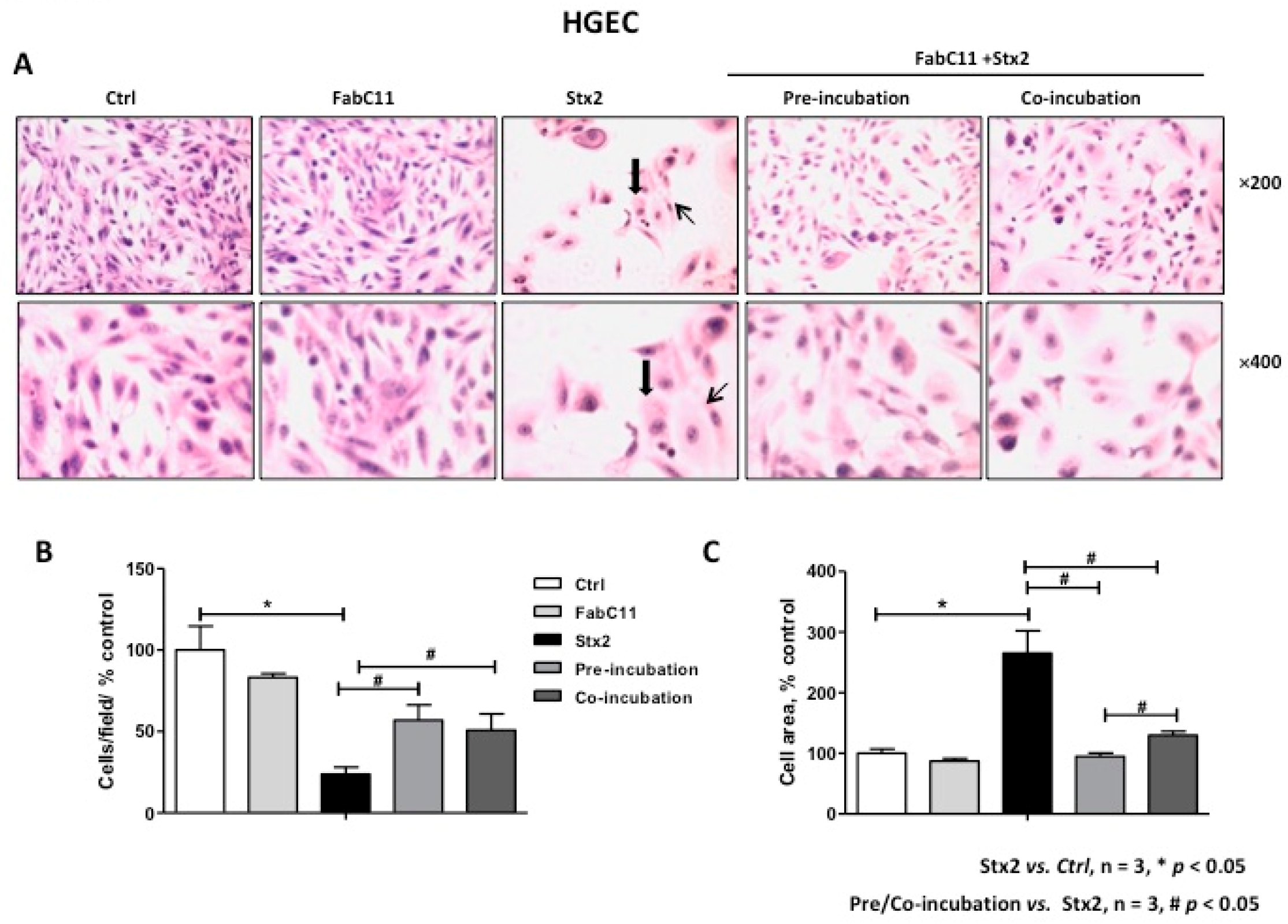
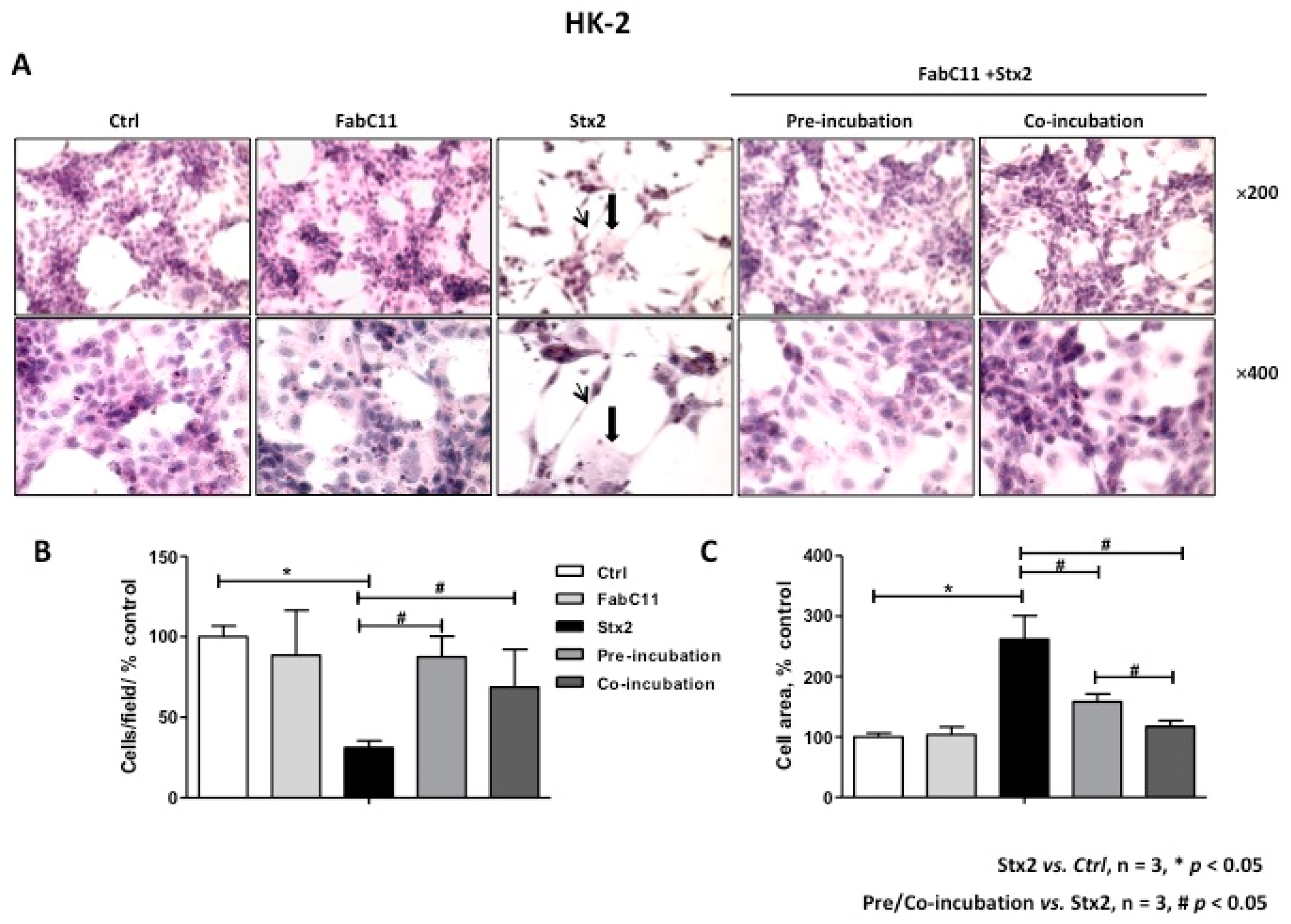
2.2. FabC11:Stx2 Prevents Morphological Alterations Induced by Stx2 in HGEC and HK-2 Cells
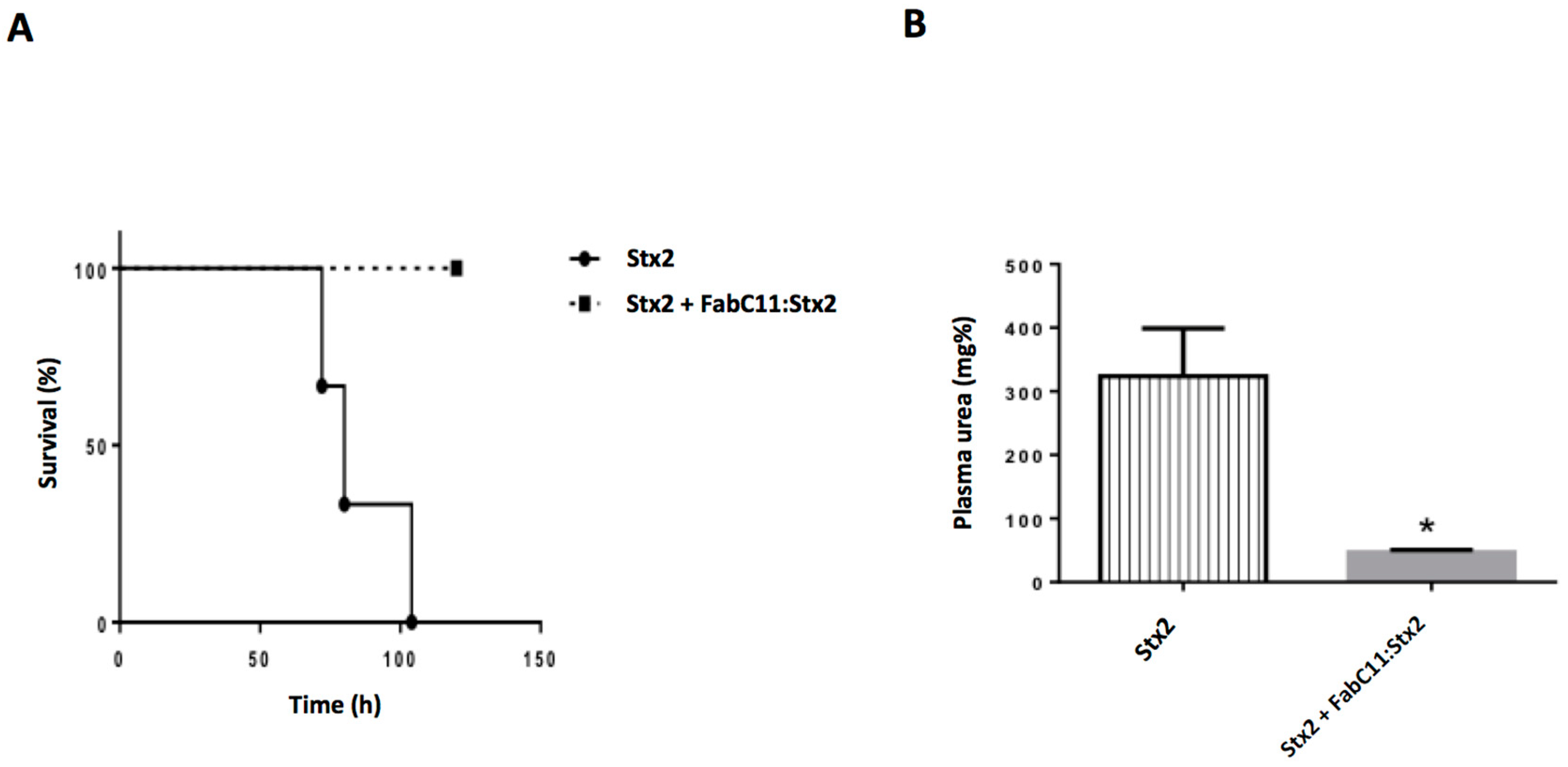
2.3. FabC11:Stx2 Protected Mice Injected with a Lethal Dose of Stx2
3. Discussion
4. Materials and Methods
4.1. Reagents
4.2. FabC11:Stx2 Affinity by Surface Plasmon Ressonance
4.3. Primary Culture
4.4. Cell Line Culture
4.5. Stx2 Neutralization Assay in HGEC and HK-2
4.6. Neutral Red Cytotoxicity Assay
4.7. Cell Morphology Analysis
4.8. In Vivo Stx2-Neutralizing Capacity
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Golshani, M.; Oloomi, M.; Bouzari, S. In silico analysis of Shiga toxins (Stxs) to identify new potential vaccine targets for Shiga toxin-producing Escherichia coli. In Silico Pharmacol. 2017, 5, 2. [Google Scholar] [CrossRef] [PubMed]
- Krüger, A.; Lucchesi, P.M. Shiga toxins and stx phages: Highly diverse entities. Microbiology 2015, 161, 451–462. [Google Scholar] [CrossRef] [PubMed]
- Kavaliauskiene, S.; Lingelem, A.B.D.; Skotland, T.; Sandvig, K. Protection against Shiga Toxins. Toxins 2017, 9, 44. [Google Scholar] [CrossRef] [PubMed]
- Hall, G.; Kurosawa, S.; Stearns-Kurosawa, D.J. Shiga Toxin Therapeutics: Beyond Neutralization. Toxins 2017, 9, 291. [Google Scholar] [CrossRef] [PubMed]
- Karpman, D.; Loos, S.; Tati, R.; Arvidsson, I. Haemolytic uraemic syndrome. J. Intern. Med. 2017, 281, 123–148. [Google Scholar] [CrossRef] [PubMed]
- Spinale, J.M.; Ruebner, R.L.; Copelovitch, L.; Kaplan, B.S. Long-term outcomes of Shiga toxin hemolytic uremic syndrome. Pediatr. Nephrol. 2013, 28, 2097–2105. [Google Scholar] [CrossRef] [PubMed]
- Girard, M.C.; Sacerdoti, F.; Rivera, F.P.; Repetto, H.A.; Ibarra, C.; Amaral, M.M. Prevention of renal damage caused by Shiga toxin type 2: Action of Miglustat on human endothelial and epithelial cells. Toxicon 2015, 105, 27–33. [Google Scholar] [CrossRef] [PubMed]
- Keenswijk, W. Eculizumab is Efficacious but Often Not Accessible. J. Pediatr. Hematol. Oncol. 2018. [Google Scholar] [CrossRef]
- Mejías, M.P.; Hiriart, Y.; Lauché, C.; Fernández-Brando, R.J.; Pardo, R.; Bruballa, A.; Ramos, M.V.; Goldbaum, F.A.; Palermo, M.S.; Zylberman, V. Development of camelid single chain antibodies against Shiga toxin type 2 (Stx2) with therapeutic potential against Hemolytic Uremic Syndrome (HUS). Sci. Rep. 2016, 6, 24913. [Google Scholar] [CrossRef]
- Melton-Celsa, A.R.; O’Brien, A.D. New therapeutic developments against Shiga toxin-producing Escherichia coli. Microbiol. Spectrum 2014, 2, 5. [Google Scholar] [CrossRef]
- Obrig, T.G. Escherichia coli Shiga Toxin Mechanisms of Action in Renal Disease. Toxins 2010, 2, 2769–2794. [Google Scholar] [CrossRef] [PubMed]
- Nilvebrant, J.; Sidhu, S.S. Construction of Synthetic Antibody Phage-Display Libraries. In Phage Display: Methods and Protocols, Methods in Molecular Biology; Hust, M., Lim, T.S., Eds.; Springer: Berlin/Heidelberg, Germany, 2018; Volume 1701, pp. 45–60. ISBN 978-1-4939-7446-7. [Google Scholar]
- Luz, D.; Chen, G.; Maranhão, A.Q.; Rocha, L.B.; Sidhu, S.; Piazza, R.M.F. Development and Characterization of Recombinant Antibody Fragments That Recognize and Neutralize In Vitro Stx2 Toxin from Shiga Toxin-Producing Escherichia coli. PLoS ONE 2015, 10, e0120481. [Google Scholar] [CrossRef] [PubMed]
- Fellouse, F.A.; Li, B.; Compaan, D.M.; Peden, A.A.; Hymowitz, S.G.; Sidhu, S.S. Molecular Recognition by a Binary Code. J. Mol. Biol. 2005, 348, 1153–1162. [Google Scholar] [CrossRef] [PubMed]
- Rocha, L.B.; Luz, D.E.; Moraes, C.T.P.; Caravelli, A.; Fernandes, I.; Guth, B.E.C.; Horton, D.S.P.Q.; Piazza, R.M.F. Interaction between Shiga Toxin and Monoclonal Antibodies: Binding Characteristics and in Vitro Neutralizing Abilities. Toxins 2012, 4, 229–247. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Amaral, M.M.; Sacerdoti, F.; Jancic, C.; Repetto, H.A.; Paton, A.W.; Paton, J.C.; Ibarra, C. Action of Shiga Toxin Type-2 and Subtilase Cytotoxin on Human Microvascular Endothelial Cells. PLoS ONE 2013, 8, e70431. [Google Scholar] [CrossRef] [PubMed]
- Takeda, T.; Yoshino, K.; Adachi, E.; Sato, Y.; Yamagata, K. In Vitro Assessment of a Chemically Synthesized Shiga Toxin Receptor Analog Attached to Chromosorb P (Synsorb Pk) as a Specific Absorbing Agent of Shiga Toxin 1 and 2. Microbiol. Immunol. 1999, 43, 331–337. [Google Scholar] [CrossRef] [PubMed]
- Lopez, E.L.; Contrini, M.M.; Glatstein, E.; González Ayala, S.; Santoro, R.; Allende, D.; Ezcurra, G.; Teplitz, E.; Koyama, T.; Matsumoto, Y.; et al. Safety and Pharmacokinetics of Urtoxazumab, a Humanized Monoclonal Antibody, against Shiga-Like Toxin 2 in Healthy Adults and in Pediatric Patients Infected with Shiga-Like Toxin-Producing Escherichia coli. Antimicrob. Ag. Chemot. 2010, 54, 239–243. [Google Scholar] [CrossRef]
- Bitzan, M.; Poole, R.; Mehran, M.; Sicard, E.; Brockus, C.; Thuning-Roberson, C.; Riviere, M. Safety and Pharmacokinetics of Chimeric Anti-Shiga Toxin 1 and Anti-Shiga Toxin 2 Monoclonal Antibodies in Healthy Volunteers. Antimicrob. Ag. Chemot. 2009, 53, 3081–3087. [Google Scholar] [CrossRef] [Green Version]
- Chen, G.; Sidhu, S.S. Design and Generation of Synthetic Antibody Libraries for Phage Display. In Monoclonal Antibodies: Methods and Protocols, Methods in Molecular Biology; Ossipow, V., Fischer, N., Eds.; Springer: Berlin/Heidelberg, Germany, 2014; Volume 1131, pp. 113–131. [Google Scholar]
- Sidhu, S.S.; Fellouse, F.A. Synthetic therapeutic antibodies. Nat. Chem. Biol. 2006, 2, 682–688. [Google Scholar] [CrossRef]
- Genst, E.D.; Messer, A.; Dobson, C.M. Antibodies and protein misfolding: From structural research tools to therapeutic strategie. Biochim. Biophs. Acta 2014, 1844, 1907–1919. [Google Scholar] [CrossRef]
- Girod, J.; Fenart, L.; Regina, A.; Dehouck, M.P.; Hong, G.; Scherrmann, M.J.; Cecchelli, R.; Roux, F. Transport of Cationized Anti-Tetanus Fab92 Fragments Across an In Vitro Blood–Brain Barrier Model: Involvement of the Transcytosis Pathway. J. Neurochem. 1999, 73, 2002–2008. [Google Scholar] [PubMed]
- Akiyoshi, D.E.; Sheoran, A.S.; Richard, R.L.; Chapman-Bonofiglio, S.; Tzipori, S. Toxin 2 Human Monoclonal Antibody against Shiga and Isotype Variants of a Recombinant Evaluation of Fab and F(ab)2. Infect. Immun. 2010, 78, 1376–1382. [Google Scholar] [CrossRef] [PubMed]
- Ruggiero, A.; Villa, C.H.; Bander, E.; Rey, D.A.; Bergkvist, M.; Batt, C.A.; Manova-Todorova, K.; Deen, W.M.; Scheinberg, D.A.; McDevitt, M.R. Paradoxical glomerular filtration of carbon nanotubes. Proc. Natl. Acad. Sci. USA 2010, 107, 12369–12374. [Google Scholar] [CrossRef] [PubMed]
- Fernandez-Brando, R.J.; Bentancor, L.V.; Mejías, M.P.; Ramos, M.V.; Exeni, A.; Exeni, C.; Laso Mdel, C.; Exeni, R.; Isturiz, M.A.; Palermo, M.S. Antibody Response to Shiga Toxins in Argentinean Children with Enteropathic Hemolytic Uremic Syndrome at Acute and Long-Term Follow-Up Periods. PLoS ONE 2011, 6, e19136. [Google Scholar] [CrossRef] [PubMed]
- Creydt, V.P.; Silberstein, C.; Zotta, E.; Ibarra, C. Cytotoxic effect of Shiga toxin-2 holotoxin and its B subunit on human renal tubular epithelial cells. Microbes Infect. 2006, 8, 410–419. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | ka (1/Ms) | kd (1/s) | KD (M) |
|---|---|---|---|
| Fab:C11 vs. Stx1 | (4.4 ± 0.9) × 105 | (1.5 ± 0.2) × 10−2 | (3.4 ± 0.2) × 10−8 |
| Fab:C11 vs. Stx2 | (4.2 ± 0.2) × 104 | (2.9 ± 0.1) × 10−4 | (7.0 ± 0.1) × 10−9 |
| mAb Stx2 | (1.0 ± 0.2) × 104 | (2.0 ± 0.3) × 10−4 | (2.0 ± 0.1) × 10−8 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Luz, D.; Amaral, M.M.; Sacerdoti, F.; Bernal, A.M.; Quintilio, W.; Moro, A.M.; Palermo, M.S.; Ibarra, C.; Piazza, R.M.F. Human Recombinant Fab Fragment Neutralizes Shiga Toxin Type 2 Cytotoxic Effects in vitro and in vivo. Toxins 2018, 10, 508. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins10120508
Luz D, Amaral MM, Sacerdoti F, Bernal AM, Quintilio W, Moro AM, Palermo MS, Ibarra C, Piazza RMF. Human Recombinant Fab Fragment Neutralizes Shiga Toxin Type 2 Cytotoxic Effects in vitro and in vivo. Toxins. 2018; 10(12):508. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins10120508
Chicago/Turabian StyleLuz, Daniela, Maria Marta Amaral, Flavia Sacerdoti, Alan Mauro Bernal, Wagner Quintilio, Ana Maria Moro, Marina Sandra Palermo, Cristina Ibarra, and Roxane Maria Fontes Piazza. 2018. "Human Recombinant Fab Fragment Neutralizes Shiga Toxin Type 2 Cytotoxic Effects in vitro and in vivo" Toxins 10, no. 12: 508. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins10120508