The Effect of Botulinum Neurotoxin Serotype a Heavy Chain on the Growth Related Proteins and Neurite Outgrowth after Spinal Cord Injury in Rats

Abstract
:1. Introduction
2. Results
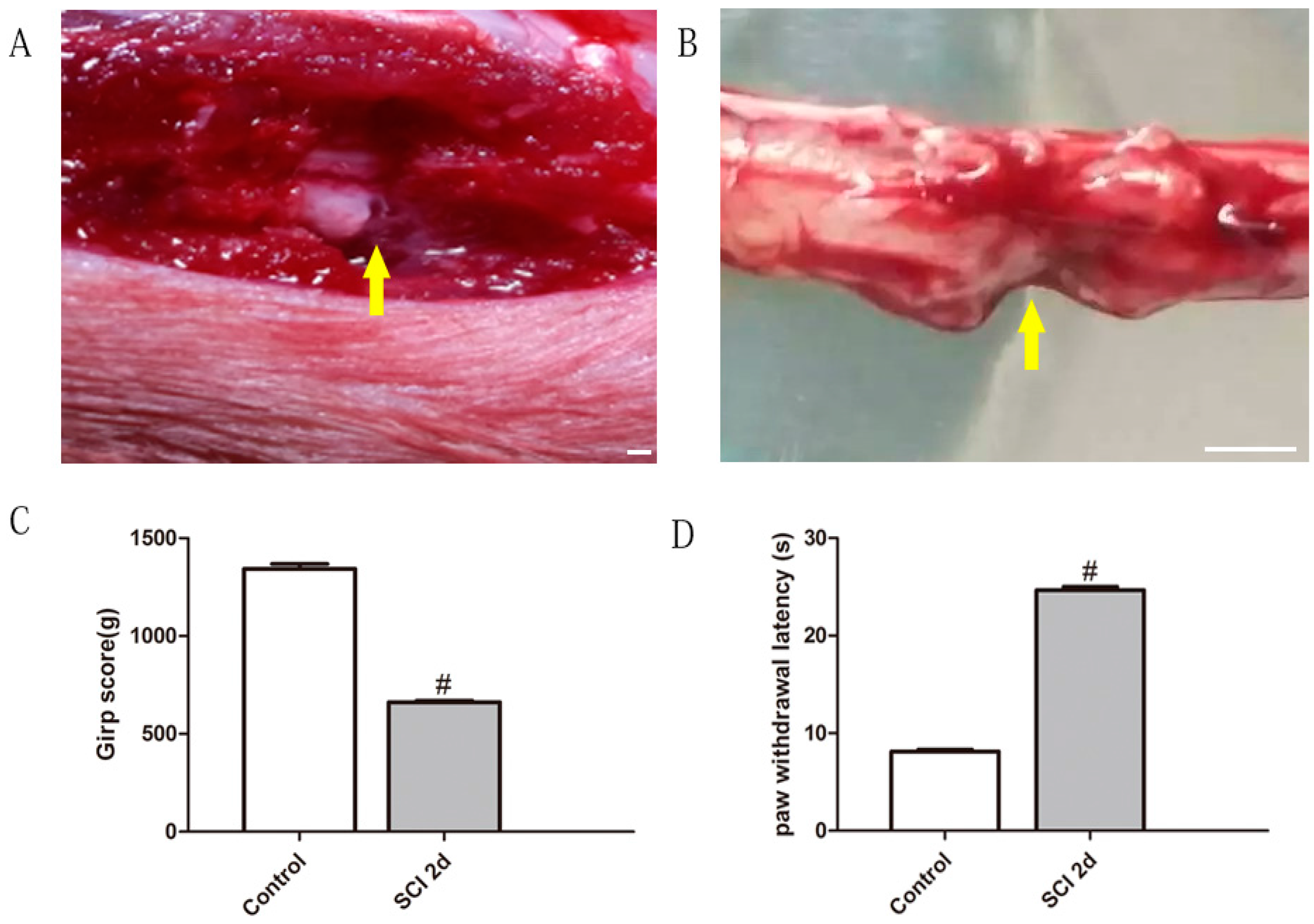
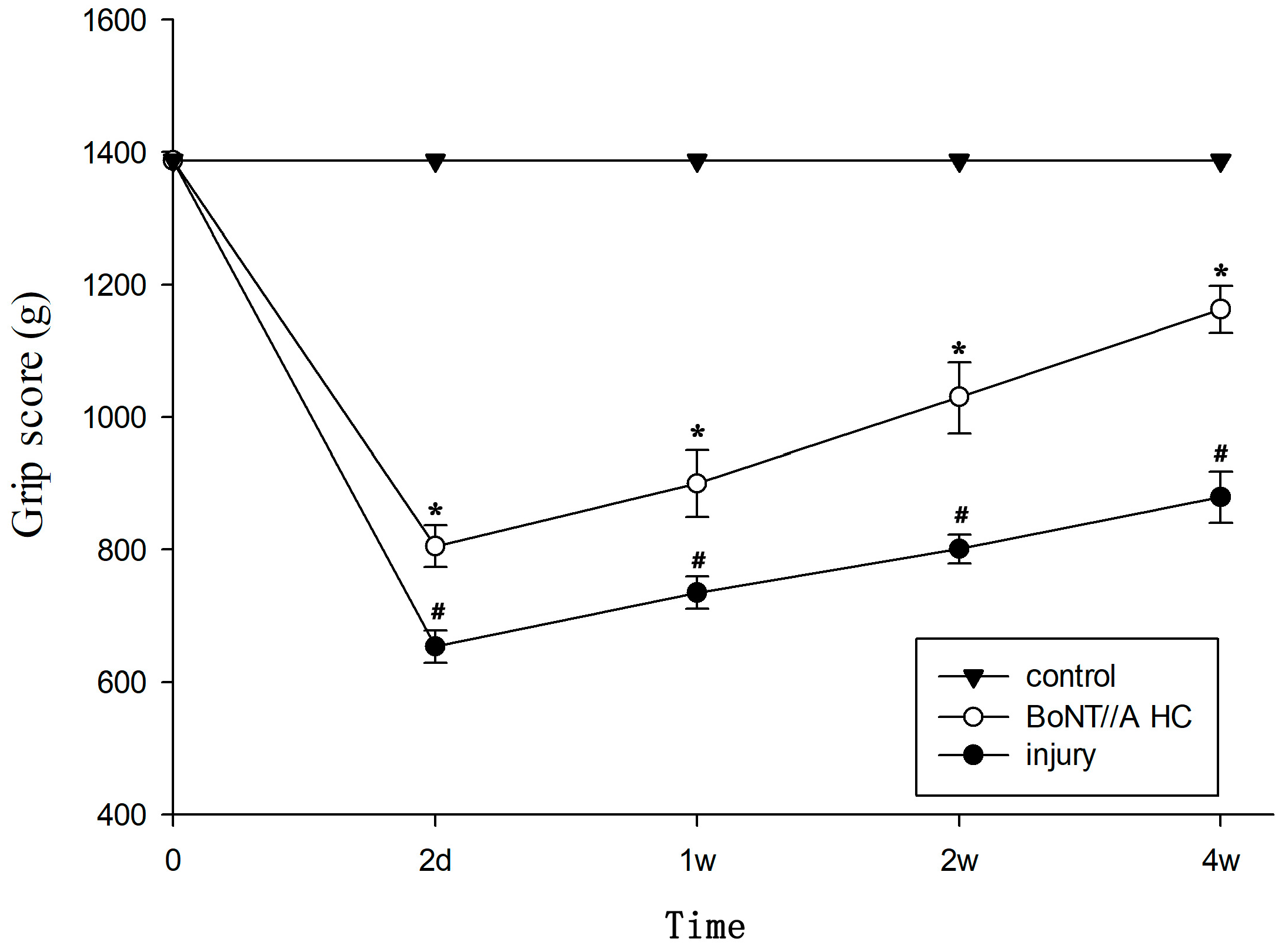
2.1. Unilateral Lumbar SCI Model and Spinal Function Evaluation
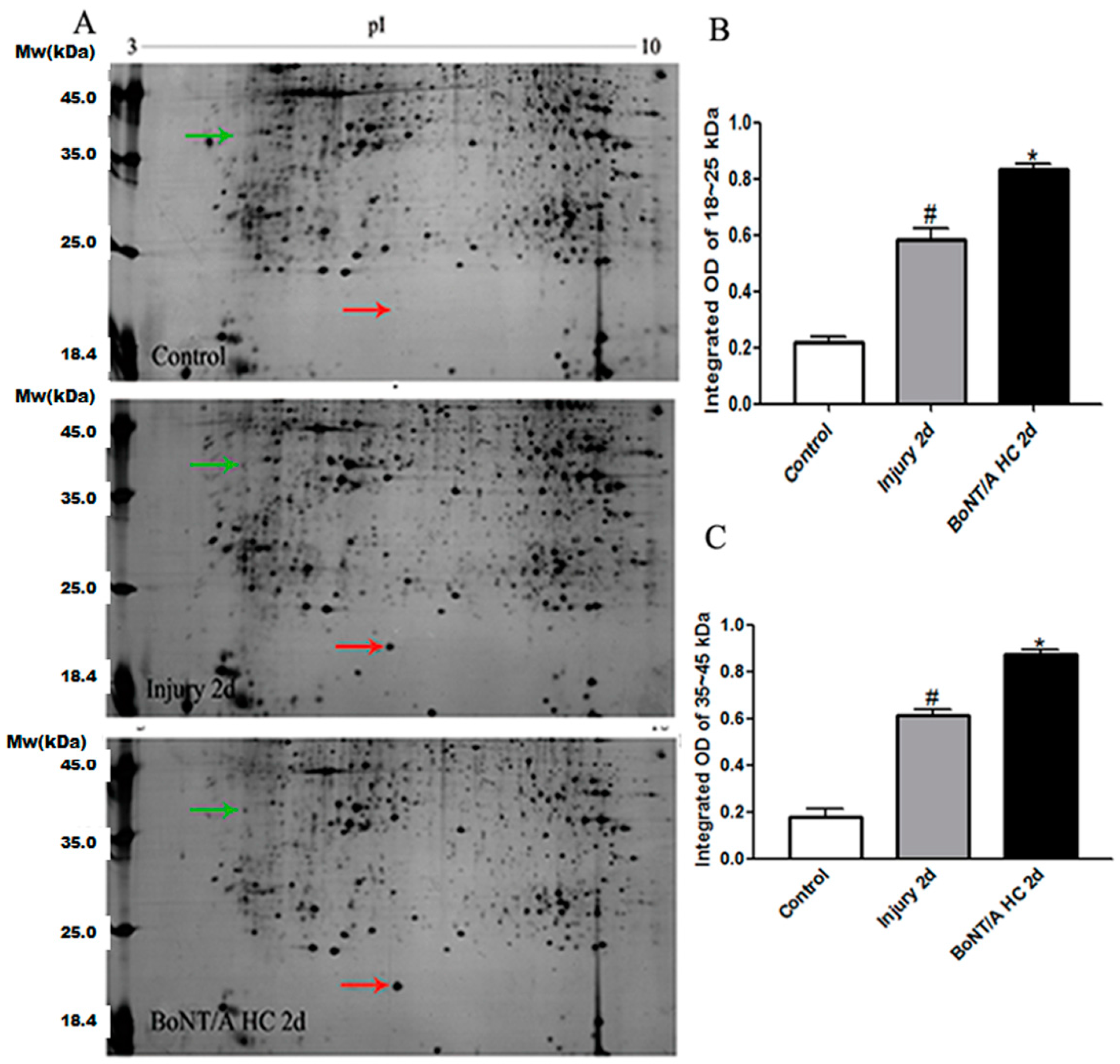
2.2. The Alteration of Local Protein Expression after SCI and the Effect of BoNT/A HC on It
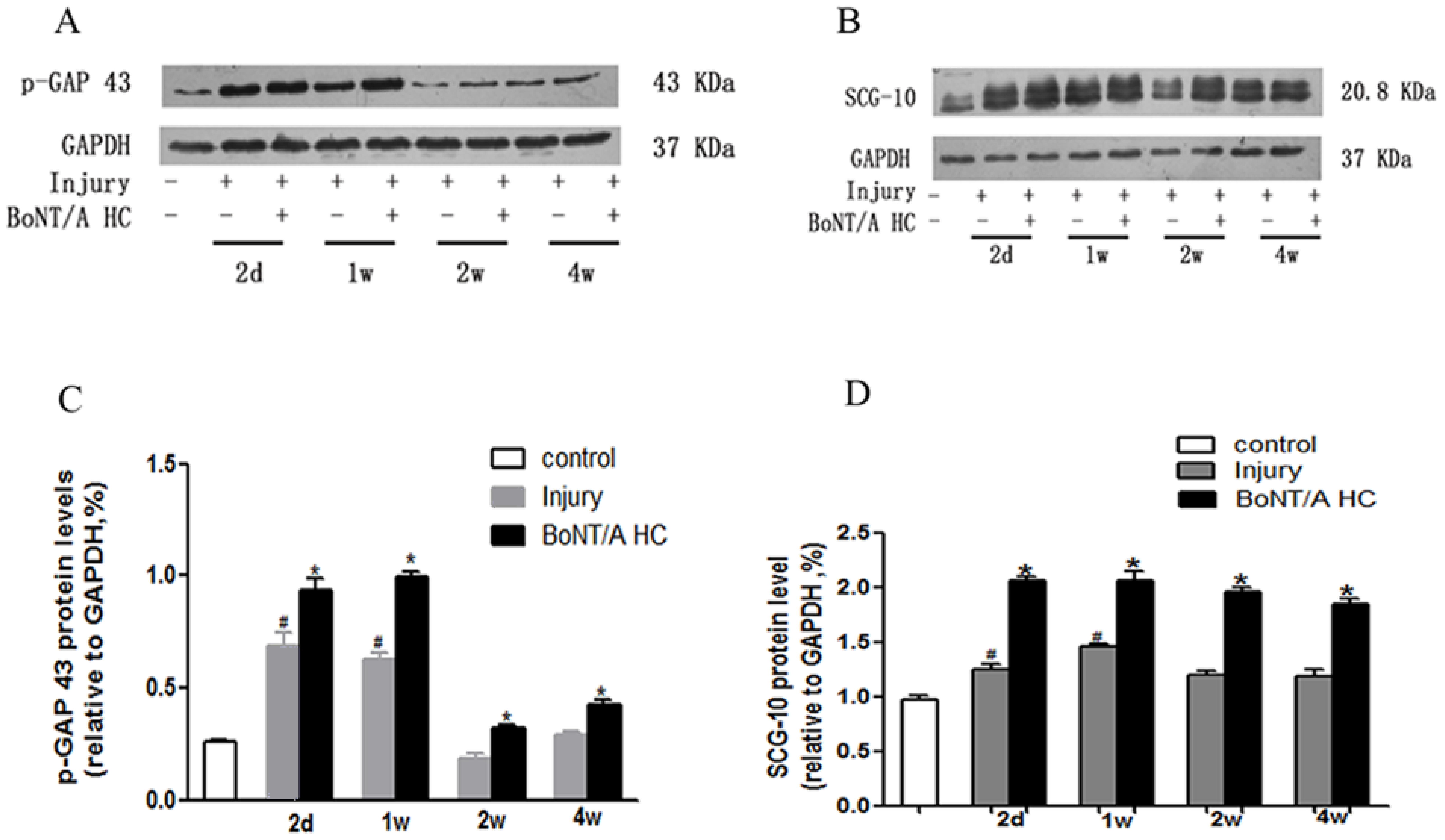
2.3. Effects of BoNT/A Heavy Chain on the Expression of Selective Growth-Associated Proteins
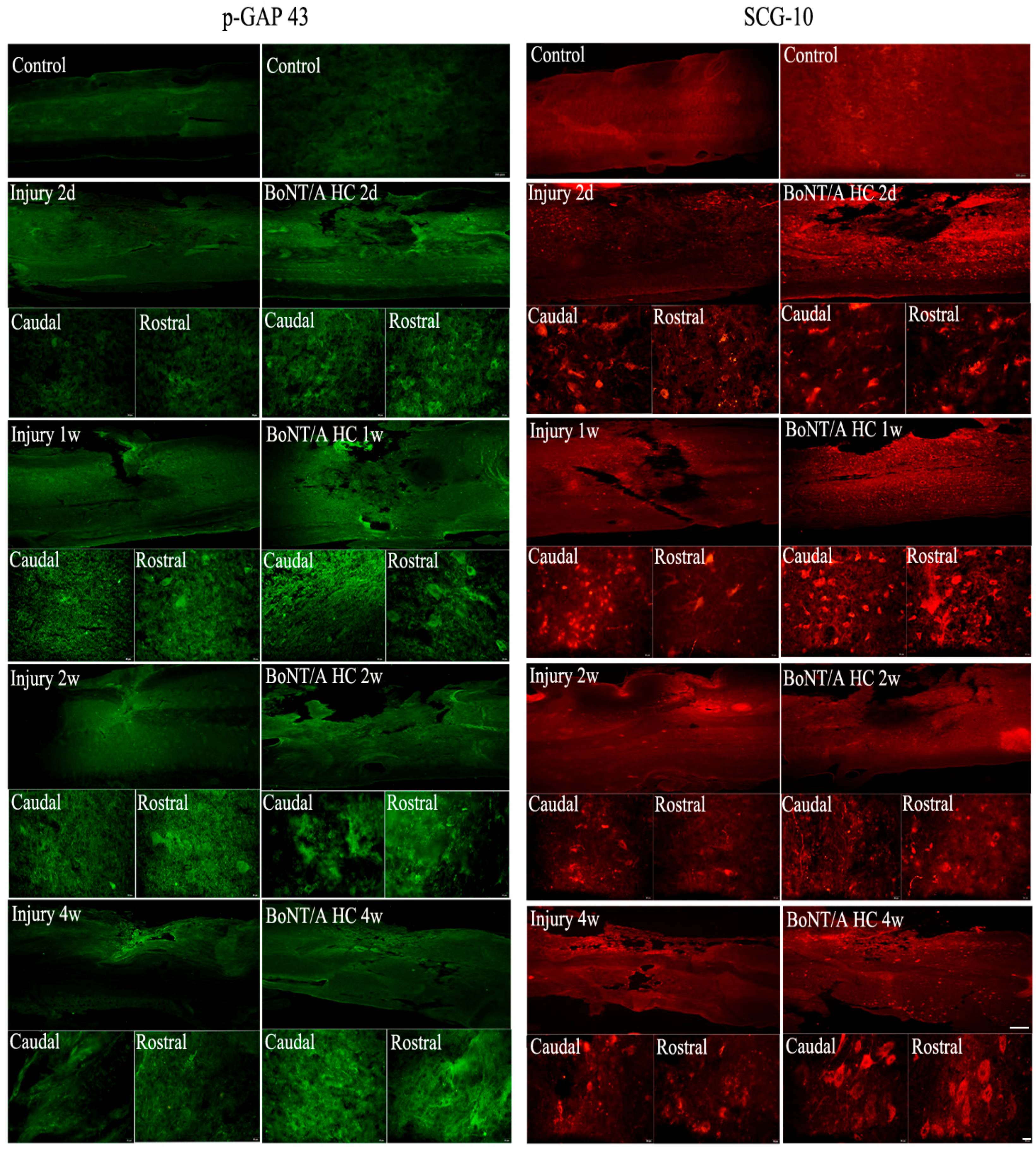
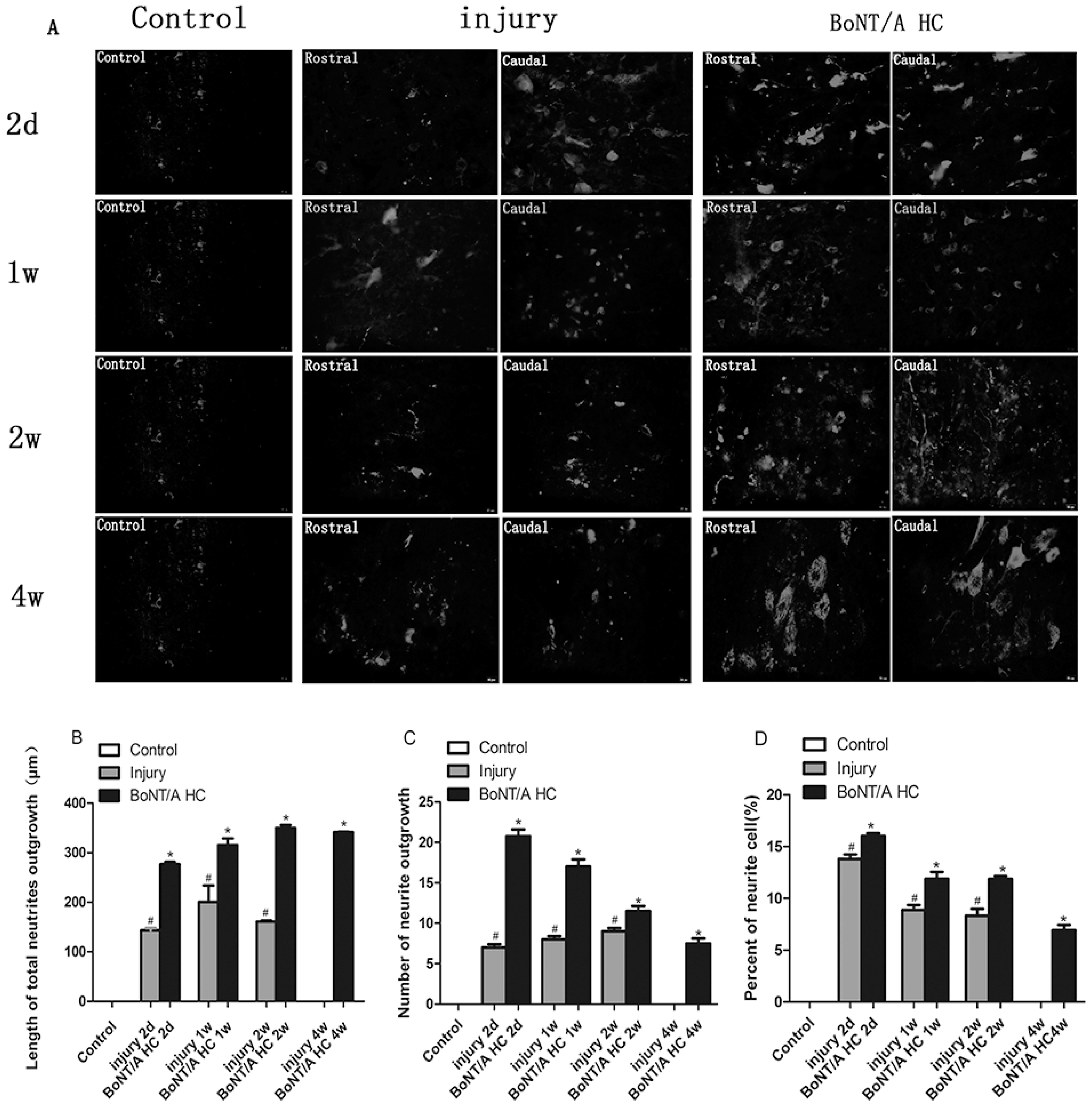
2.4. The Distribution of Positive GAP43 and SCG10 Immunofluorescence at the Injured Area after Administration of the BoNT/A HC
2.5. The Measurement of SCG 10 Positive Neuronal Processes
2.6. The Amelioration of Motor and Sensory Function by BoNT/A HC Treatment Based on SCI
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Establishing the Rat Spinal Cord Hemi-Section Injury Model
5.2. BoNT/A Heavy Chain Administration and Groups
5.3. Two-Dimensional Gel Electrophoresis
5.4. SDS-PAGE and Western Blot
5.5. Immunofluorescence Staining
5.6. Measurement of Neuronal Processes
5.7. Behavioral Evaluation
5.8. Statistical Analysis
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Silver, J.; Miller, J.H. Regeneration beyond the glial scar. Nat. Rev. Neurosci. 2004, 5, 146–156. [Google Scholar] [CrossRef] [PubMed]
- Giger, R.J.; Hollis, E.R.; Tuszynski, M.H. Guidance molecules in axon regeneration. Cold Spring Harb. Perspect. Biol. 2010, 2, 1–21. [Google Scholar] [CrossRef] [PubMed]
- Simpson, L.A.; Eng, J.J.; Hsieh, J.T.C.; Wolfe, D.L.; Team, T.S. The health and life priorities of individuals with spinal cord injury: A systematic review. J. Neurotrauma 2012, 29, 1548–1555. [Google Scholar] [CrossRef] [PubMed]
- Ferguson, T.A.; Son, Y.J. Extrinsic and intrinsic determinants of nerve regeneration. J. Tissue Eng. 2011, 2, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Baldwin, K.T.; Giger, R.J. Insights into the physiological role of CNS regeneration inhibitors. Front. Mol. Neurosci. 2015, 8, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Geoffroy, C.G.; Zheng, B. Myelin-associated inhibitors in axonal growth after CNS injury. Curr. Opin. Neurobiol. 2014, 27, 31–38. [Google Scholar] [CrossRef] [PubMed]
- He, Z. Intrinsic control of axon regeneration. J. Biomed. Res. 2010, 24, 2–5. [Google Scholar] [CrossRef] [Green Version]
- Mckerracher, L.; Rosen, K.M. MAG, myelin and overcoming growth inhibition in the CNS. Front. Mol. Neurosci. 2015, 8, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Amemori, T.; Ruzicka, J.; Romanyuk, N.; Jhanwaruniyal, M.; Sykova, E.; Jendelova, P. Comparison of intraspinal and intrathecal implantation of induced pluripotent stem cell-derived neural precursors for the treatment of spinal cord injury in rats. Stem Cell Res. Ther. 2015, 6, 257–268. [Google Scholar] [CrossRef] [PubMed]
- Yang, P.; Qin, Y.; Bian, C.; Zhao, Y.; Zhang, W. Intrathecal delivery of il-6 reactivates the intrinsic growth capacity of pyramidal cells in the sensorimotor cortex after spinal cord injury. PLoS ONE 2015, 10, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Zhang, P.; Ray, R.; Singh, B.R.; Ray, P. Mastoparan-7 rescues botulinum toxin-A poisoned neurons in a mouse spinal cord cell culture model. Toxicon 2013, 76, 37–43. [Google Scholar] [CrossRef] [PubMed]
- Zichel, R.; Mimran, A.; Keren, A.; Barnea, A.; Steinberger-Levy, I.; Marcus, D.; Turgeman, A.; Reuvenyet, S. Efficacy of a potential trivalent vaccine based on Hc fragments of botulinum toxins A, B, and E produced in a cell-free expression system. Clin. Vaccine Immunol. 2010, 17, 784–792. [Google Scholar] [CrossRef] [PubMed]
- Rossetto, O.; Pirazzini, M.; Montecucco, C. Botulinum neurotoxins: Genetic, structural and mechanistic insights. Nat. Rev. Microbiol. 2014, 12, 535–549. [Google Scholar] [CrossRef] [PubMed]
- Meunier, F.A.; Lisk, G.; Sesardic, D.; Dolly, J.O. Dynamics of motor nerve terminal remodeling unveiled using SNARE-cleaving botulinum toxins: The extent and duration are dictated by the sites of SNAP-25 truncation. Mol. Cell. Neurosci. 2003, 22, 454–466. [Google Scholar] [CrossRef]
- Dolly, O. Synaptic transmission: Inhibition of neurotransmitter release by Botulinum toxins. Headache 2003, 43, 16–24. [Google Scholar] [CrossRef]
- Jankovic, J. Botulinum toxin in clinical practice. J. Neurol. Neurosurg. Psychiatry 2004, 75, 951–957. [Google Scholar] [CrossRef] [PubMed]
- Beske, P.H.; Hoffman, K.M.; Machamer, J.B.; Eisen, M.R.; McNutt, P.M. Use-dependent potentiation of voltage-gated calcium channels rescues neurotransmission in nerve terminals intoxicated by botulinum neurotoxin serotype A. Sci. Rep. 2017, 7, 15862. [Google Scholar] [CrossRef] [PubMed]
- Durham, P.L.; Cady, R.; Cady, R. Regulation of Calcitonin Gene-Related Peptide Secretion From Trigeminal Nerve Cells by Botulinum Toxin Type A: Implications for Migraine Therapy. Headache 2004, 44, 35–43. [Google Scholar] [CrossRef] [PubMed]
- Guo, B.L.; Zheng, C.X.; Sui, B.D.; Li, Y.Q.; Wang, Y.Y.; Yang, Y.L. A closer look to botulinum neurotoxin type A-induced analgesia. Toxicon 2013, 71, 134–139. [Google Scholar] [CrossRef] [PubMed]
- Aoki, K.R. Review of a Proposed Mechanism for the Antinociceptive Action of Botulinum Toxin Type A. Neurotoxicology 2005, 26, 785–793. [Google Scholar] [CrossRef] [PubMed]
- Jeynes, L.C.; Gauci, C.A. Evidence for the use of botulinum toxin in the chronic pain setting: A review of the literature. Pain Pract. 2008, 8, 269–276. [Google Scholar] [CrossRef] [PubMed]
- Aoki, K.R. Future aspects of botulinum neurotoxins. J. Neural Trans. 2008, 115, 567–573. [Google Scholar] [CrossRef] [PubMed]
- Thenganatt, M.A.; Fahn, S. Botulinum toxin for the treatment of movement disorders. Curr. Neurol. Neurosci. Rep. 2012, 12, 399–409. [Google Scholar] [CrossRef] [PubMed]
- Foster, L.; Clapp, L.; Erickson, M.; Jabbari, B. Botulinum toxin A and chronic low back pain: A randomized, double-blind study. Neurology 2001, 56, 1290–1293. [Google Scholar] [CrossRef] [PubMed]
- Ayyar, B.V.; Tajhya, R.B.; Beeton, C.; Atassi, M.Z. Antigenic sites on the H N domain of botulinum neurotoxin A stimulate protective antibody responses against active toxin. Sci. Rep. 2015, 5, 15776. [Google Scholar] [CrossRef] [PubMed]
- Humeau, Y.; Doussau, F.; Grant, N.J.; Poulain, B. How botulinum and tetanus neurotoxins block neurotransmitter release. Biochimie 2002, 82, 427–446. [Google Scholar] [CrossRef]
- Juzans, P.; Comella, J.X.; Molgo, J.; Faille, L.; Angaut-Petit, D. Nerve terminal sprouting in botulinum type-A treated mouse levator auris longus muscle. Neuromuscul. Disord. 1996, 6, 177–185. [Google Scholar] [CrossRef]
- Foran, P.G.; Davletov, B.; Meunier, F.A. Getting musclesmoving again after botulinum toxin: Novel therapeuti challenges. Trends Mol. Med. 2003, 9, 291–299. [Google Scholar] [CrossRef]
- Ko, C.P. Do nerve terminal sprouts contribute to functional recovery from botulinum neurotoxin A? J. Physiol. 2008, 586, 3021. [Google Scholar] [CrossRef] [PubMed]
- Coffield, J.A.; Yan, X. Neuritogenic actions of botulinum neurotoxin A on cultured motor neurons. J. Pharmacol. Exp. Ther. 2009, 330, 352–358. [Google Scholar] [CrossRef] [PubMed]
- Mori, N.; Morii, H. SCG10-related neuronal growth-associated proteins in neural development, plasticity, degeneration, and aging. J. Neurosci. Res. 2002, 70, 264–273. [Google Scholar] [CrossRef] [PubMed]
- Pellier-Monnin, V.; Astic, L.; Bichet, S.; Riederer, B.M.; Grenningloh, G. Expression of SCG10 and stathmin proteins in the rat olfactory system during development and axonal regeneration. J. Comp. Neurol. 2001, 433, 239–254. [Google Scholar] [CrossRef] [PubMed]
- Holahan, M.R. A Shift from a Pivotal to Supporting Role for the Growth-Associated Protein (GAP-43) in the Coordination of Axonal Structural and Functional Plasticity. Front. Cell. Neurosci. 2017, 11, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Goldberg, J.L.; Klassen, M.P.; Hua, Y.; Barres, B.A. Amacrine-signaled loss of intrinsic axon growth ability by retinal ganglion cells. Science 2002, 296, 1860–1864. [Google Scholar] [CrossRef] [PubMed]
- Fernandes, K.J.; Fan, D.P.; Tsui, B.J.; Cassar, S.L.; Tetzlaff, W. Influence of the axotomy to cell body distance in rat rubrospinal and spinal motoneurons: Differential regulation of gap-43, tubulins, and neurofilamentm. J. Comp. Neurol. 1999, 414, 495–510. [Google Scholar] [CrossRef]
- Fagoe, N.D.; Heest, J.V.; Verhaagen, J. Spinal cord injury and the neuron-intrinsic regeneration-associated gene program. Neuromol. Med. 2014, 16, 799–813. [Google Scholar] [CrossRef] [PubMed]
- Lieberman, A.R. The axon reaction: A review of the principal features of perikaryal responses to axon injury. Int. Rev. Neurobiol. 1971, 14, 49–124. [Google Scholar] [PubMed]
- Grafstein, B. The nerve cell body response to axotomy. Exp. Neurol. 1975, 48, 32–51. [Google Scholar] [CrossRef]
- Skene, J.H.; Willard, M. Axonally Transported Proteins Associated with Axon Growth in Rabbit Central and Peripheral Nervous Systems. J. Cell Biol. 1981, 89, 96–103. [Google Scholar] [CrossRef] [PubMed]
- Skene, J.H. Axonal growth-associated proteins. Annu. Rev. Neurosci. 1989, 12, 127–156. [Google Scholar] [CrossRef] [PubMed]
- Tetzlaff, W.; Alexander, S.W.; Miller, F.D.; Bisby, M.A. Response of facial and rubrospinal neurons to axotomy: Changes in mrna expression for cytoskeletal proteins and gap-43. J. Neurosci. 1991, 11, 2528–2544. [Google Scholar] [PubMed]
- Shin, J.E.; Geisler, S.; Diantonio, A. Dynamic regulation of SCG10 in regenerating axons after injury. Exp. Neurol. 2014, 252, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Ozon, S.; Maucuer, A.; Sobel, A. The stathmin family—Molecular and biological characterization of novel mammalian proteins expressed in the nervous system. Eur. J. Biochem. 2010, 248, 794–806. [Google Scholar] [CrossRef]
- Riederer, B.M.; Pellier, V.; Antonsson, B.; Paolo, G.D.; Stimpson, S.A.; Lütjens, R.; Catsicas, S.; Grenningloh, G. Regulation of microtubule dynamics by the neuronal growth-associated protein SCG 10. Proc. Natl. Acad. Sci. USA 1997, 94, 741–745. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Shan, C.; Cao, W.; Zhang, C.; Teng, J.; Chen, J. SCG10 promotes non-amyloidogenic processing of amyloid precursor protein by facilitating its trafficking to the cell surface. Hum. Mol. Genet. 2013, 22, 4888–4900. [Google Scholar] [CrossRef] [PubMed]
- Prado, W.A. Antinociceptive potency of intrathecal morphine in the rat tail flick test: A comparative study using acute lumbar catheter in rats with or without a chronic atlanto-occipital catheter. J. Neurosci. Methods 2003, 129, 33–39. [Google Scholar] [CrossRef]
- Ulmann, L.; Rodeau, J.L.; Danoux, L.; Contet-Audonneau, J.L.; Pauly, G.; Schlichter, R. Dehydroepiandrosterone and neurotrophins favor axonal growth in a sensory neuron-keratinocyte coculture model. Neuroscience 2009, 159, 514–525. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Groups | 0 d | 2 d | 1 w | 2 w | 4 w |
|---|---|---|---|---|---|
| Control | 8.5 ± 0.2 | 8.5 ± 0.2 | 8.3 ± 0.1 | 7.8 ± 0.2 | 8.2 ± 0.2 |
| Injury | 8.5 ± 0.2 | 22.8 ± 1.0 # | 22.1 ± 0.9 # | 22.4 ± 1.0 # | 21.5 ± 0.9 # |
| BoNT/A HC | 8.5 ± 0.2 | 24.4 ± 0.3 | 24.5 ± 0.3 | 24.1 ± 0.4 | 22.5 ± 0.9 |
| Name of Group | Pseudo Surgery | SCI without BoNT/A HC | SCI with BoNT/A HC | ||||||
|---|---|---|---|---|---|---|---|---|---|
| Time | 0 | 2 d | 1 wk | 2 wks | 4 wks | 2 d | 1 wk | 2 wks | 4 wks |
| No. of animals | 6 | 6 | 6 | 6 | 6 | 6 | 6 | 6 | 6 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, Y.-F.; Liu, F.; Lan, J.; Bai, J.; Li, X.-Q. The Effect of Botulinum Neurotoxin Serotype a Heavy Chain on the Growth Related Proteins and Neurite Outgrowth after Spinal Cord Injury in Rats. Toxins 2018, 10, 66. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins10020066
Wang Y-F, Liu F, Lan J, Bai J, Li X-Q. The Effect of Botulinum Neurotoxin Serotype a Heavy Chain on the Growth Related Proteins and Neurite Outgrowth after Spinal Cord Injury in Rats. Toxins. 2018; 10(2):66. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins10020066
Chicago/Turabian StyleWang, Ya-Fang, Fu Liu, Jing Lan, Juan Bai, and Xia-Qing Li. 2018. "The Effect of Botulinum Neurotoxin Serotype a Heavy Chain on the Growth Related Proteins and Neurite Outgrowth after Spinal Cord Injury in Rats" Toxins 10, no. 2: 66. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins10020066