Genotoxic and Cytotoxic Effects on the Immune Cells of the Freshwater Bivalve Dreissena polymorpha Exposed to the Environmental Neurotoxin BMAA

 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
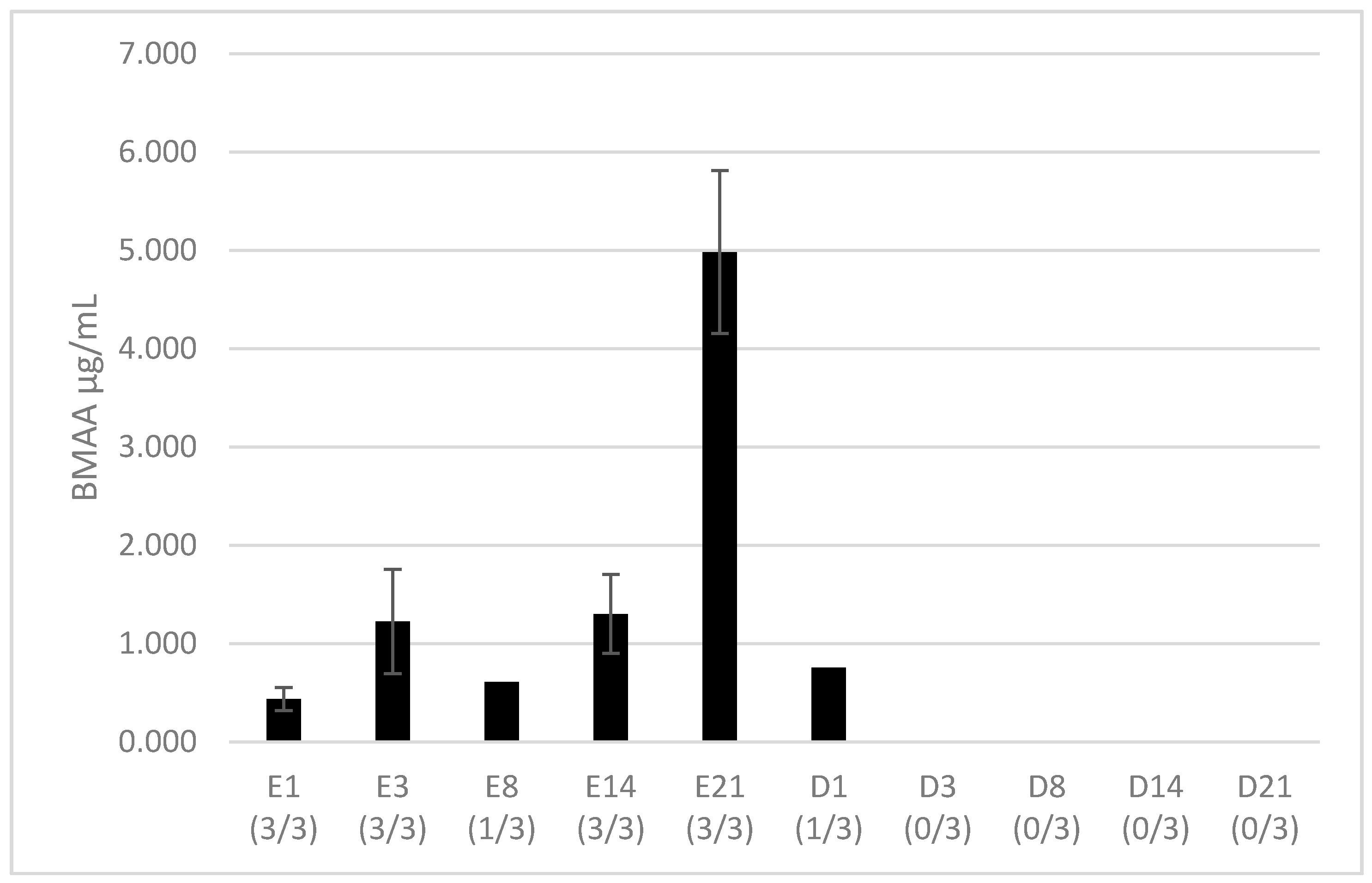
2.1. BMAA Accumulation in Hemolymph
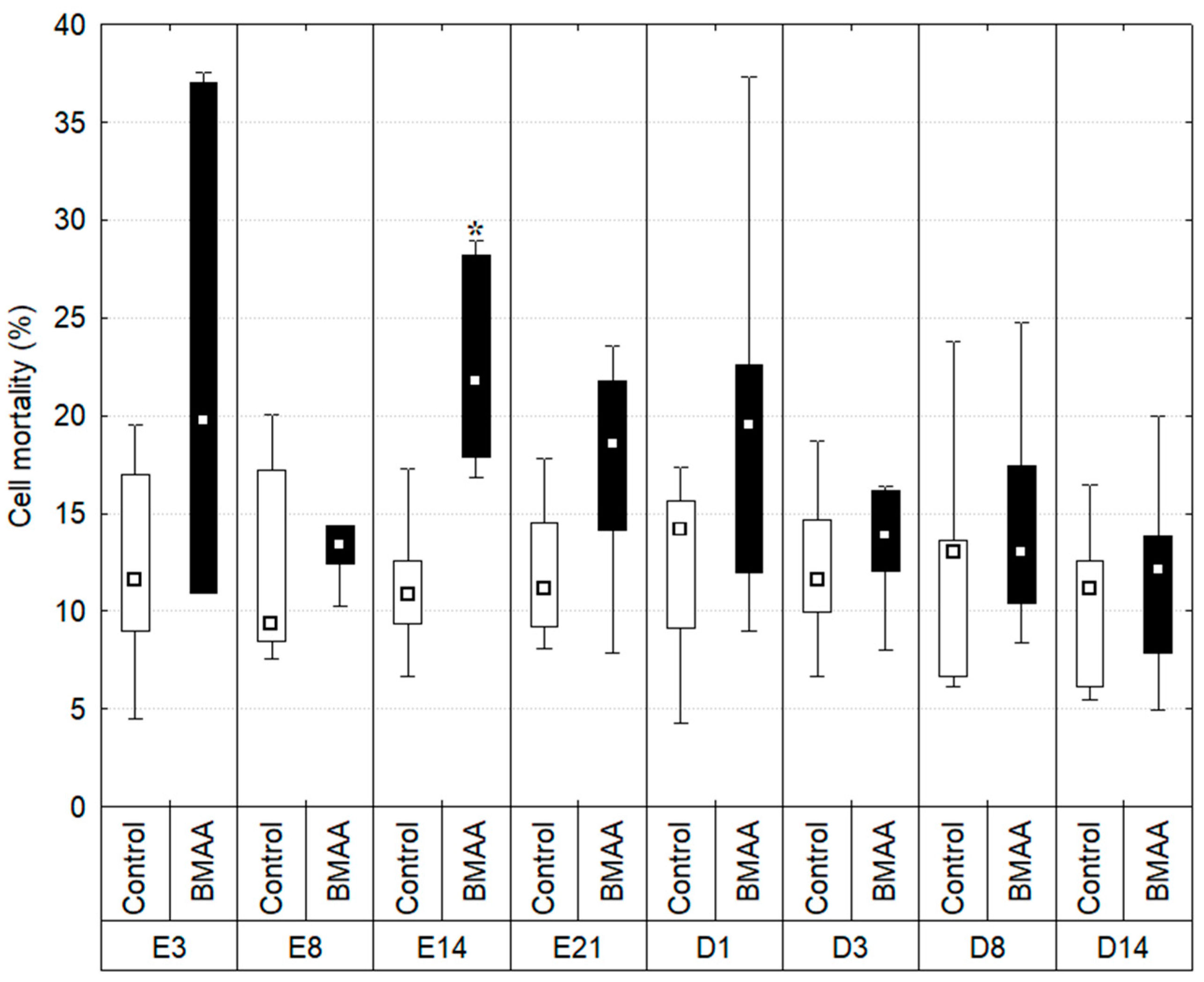
2.2. Hemocytes Mortality
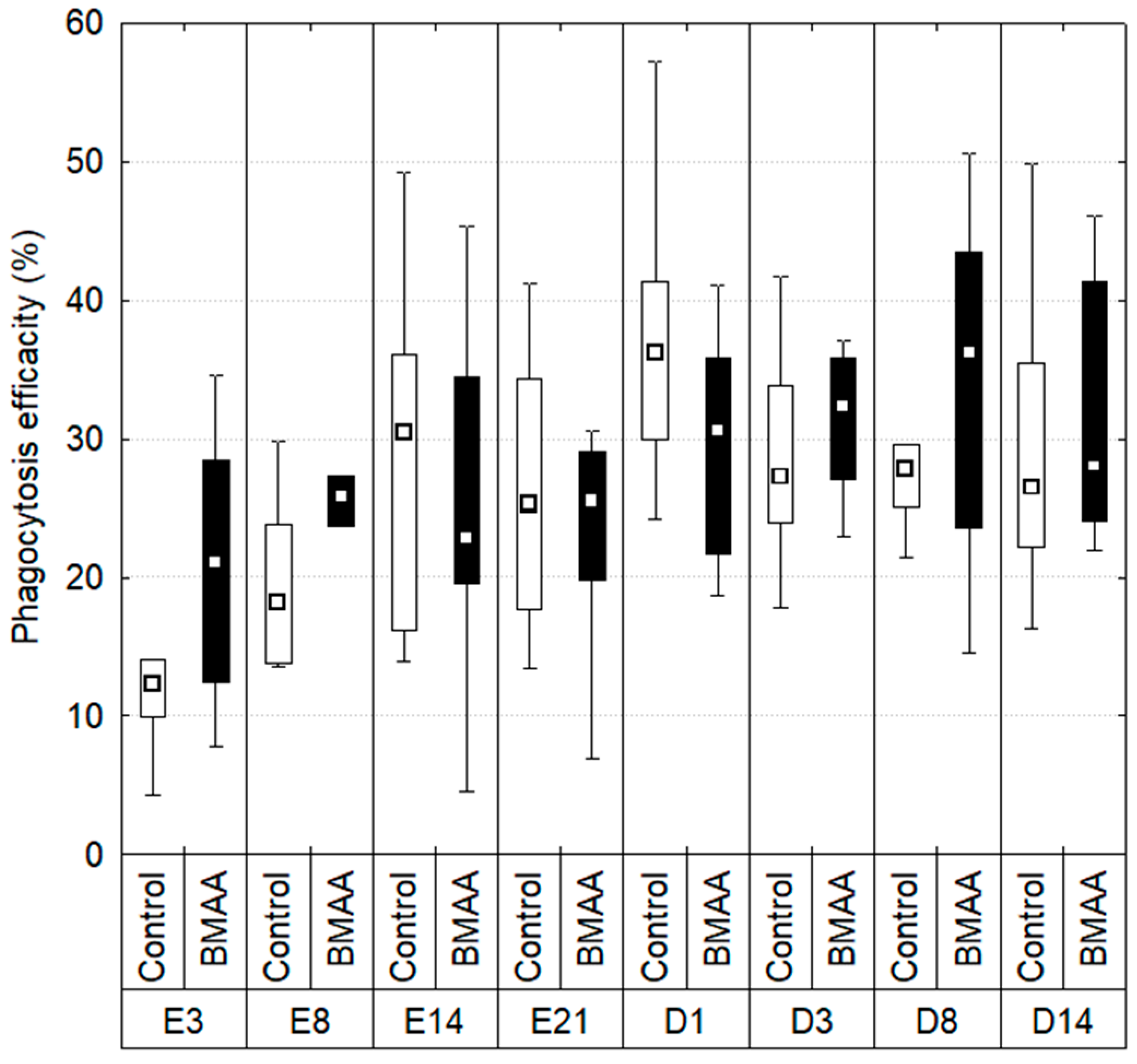
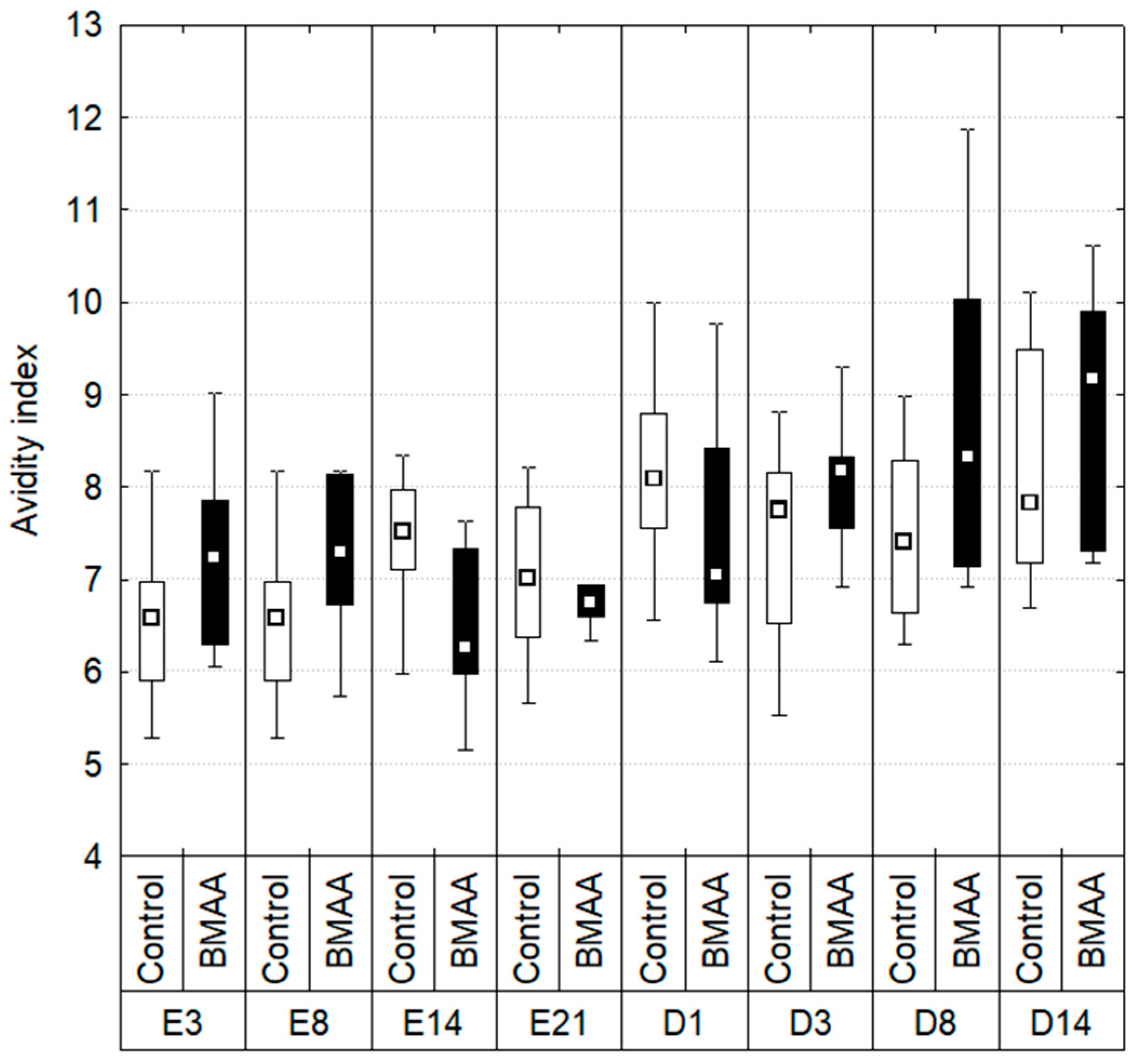
2.3. Hemocytes Phagocytosis
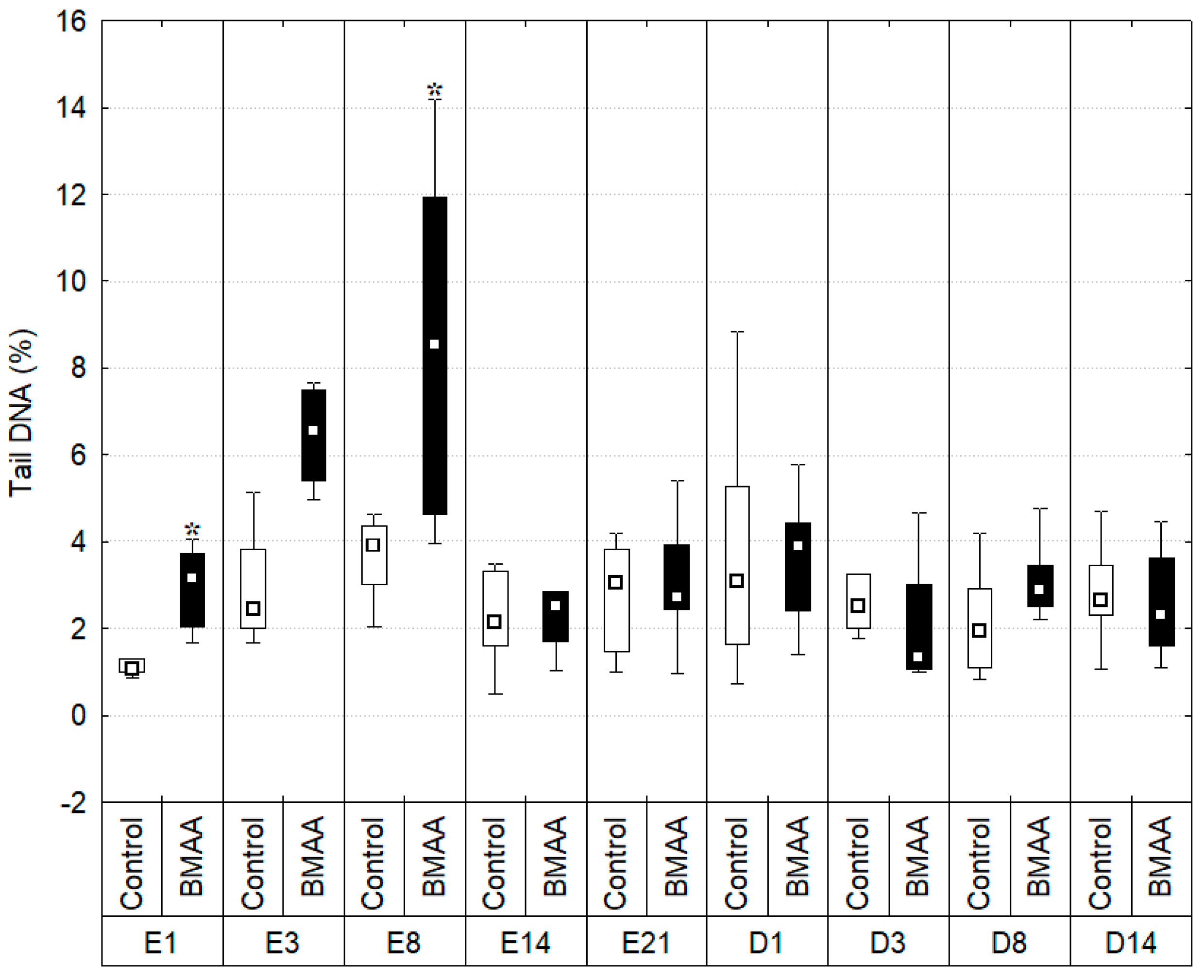
2.4. DNA Integrity
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Mussels Sampling and Acclimation
5.2. Exposure
5.3. BMAA in Hemolymph
5.4. Preparation of Samples for Flow Cytometry and COMET Assay Analysis
5.4.1. Flow Cytometry
- Mortality assay
- Phagocytosis efficacity and avidity index
5.4.2. DNA Integrity Measurements
5.5. Statistics
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Heisler, J.; Glibert, P.M.; Burkholder, J.M.; Anderson, D.M.; Cochlan, W.; Dennison, W.C.; Dortch, Q.; Gobler, C.J.; Heil, C.A.; Humphries, E.; et al. Eutrophication and Harmful Algal Blooms: A scientific consensus. Harmful Algae 2008, 8, 3–13. [Google Scholar] [CrossRef] [PubMed]
- Paerl, H.W.; Otten, T.G. Harmful Cyanobacterial Blooms: Causes, Consequences, and Controls. Microb. Ecol. 2013, 65, 995–1010. [Google Scholar] [CrossRef] [PubMed]
- Cox, P.A.; Banack, S.A.; Murch, S.J. Biomagnification of Cyanobacterial Neurotoxins and Neurodegenerative Disease among the Chamorro People of Guam. Proc. Natl. Acad. Sci. USA 2003, 100, 13380–13383. [Google Scholar] [CrossRef] [PubMed]
- Murch, S.J.; Cox, P.A.; Banack, S.A.; Steele, J.C.; Sacks, O.W. Occurrence of β-N-methylamino-l-alanine (BMAA) in ALS/PDC Patients from Guam. Acta Neurol. Scand. 2004, 110, 267–269. [Google Scholar] [CrossRef] [PubMed]
- Murch, S.J.; Cox, P.A.; Banack, S.A. A Mechanism for Slow Release of Biomagnified Cyanobacterial Neurotoxins and Neurodegenerative Disease in Guam. Proc. Natl. Acad. Sci. USA 2004, 101, 12228–12231. [Google Scholar] [CrossRef] [PubMed]
- Chernoff, N.; Hill, D.J.; Diggs, D.L.; Faison, B.D.; Francis, B.M.; Lang, J.R.; Larue, M.M.; Le, T.-T.; Loftin, K.A.; Lugo, J.N.; et al. A Critical Review of the Postulated Role of the Non-essential Amino Acid, β-N-methylamino-l-alanine, in Neurodegenerative Disease in Humans. J. Toxicol. Environ. Heal. B 2017, 20, 183–229. [Google Scholar] [CrossRef] [PubMed]
- Weiss, J.H.; Choi, D.W. Beta-N-methylamino-l-alanine Neurotoxicity: Requirement for Bicarbonate as a Cofactor. Science 1988, 241, 973–975. [Google Scholar] [CrossRef] [PubMed]
- Lobner, D. Mechanisms of beta-N-methylamino-l-alanine Induced Neurotoxicity. Amyotroph. Lateral Scler. 2009, 10 (Suppl. 2), 56–60. [Google Scholar] [CrossRef] [PubMed]
- Popova, A.A.; Koksharova, O.A. Neurotoxic Non-proteinogenic Amino Acid β-N-Methylamino-l-alanine and Its Role in Biological Systems. Biochemistry 2016, 81, 794–805. [Google Scholar] [CrossRef] [PubMed]
- Nunn, P.B.; Ponnusamy, M. β-N-Methylaminoalanine (BMAA): Metabolism and Metabolic Effects in Model Systems and in Neural and Other Tissues of the Rat in vitro. Toxicon 2009, 54, 85–94. [Google Scholar] [CrossRef] [PubMed]
- Dunlop, R.A.; Cox, P.A.; Banack, S.A.; Rodgers, K.J. The Non-protein Amino Acid BMAA is Misincorporated into Human Proteins in Place of l-Serine Causing Protein Misfolding and Aggregation. PLoS ONE 2013, 8, e75376. [Google Scholar] [CrossRef] [PubMed]
- Onselen, R.; Downing, S.; Kemp, G.; Downing, T. Investigating β-N-Methylamino-l-alanine Misincorporation in Human Cell Cultures: A Comparative Study with Known Amino Acid Analogues. Toxins 2017, 9, 400. [Google Scholar] [CrossRef] [PubMed]
- Faassen, E. Presence of the Neurotoxin BMAA in Aquatic Ecosystems: What Do We Really Know? Toxins 2014, 6, 1109–1138. [Google Scholar] [CrossRef] [PubMed]
- Monteiro, M.; Costa, M.; Moreira, C.; Vasconcelos, V.M.; Baptista, M.S. Screening of BMAA-producing cyanobacteria in cultured isolates and in in situ blooms. J. Appl. Phycol. 2016. [Google Scholar] [CrossRef]
- Réveillon, D.; Abadie, E.; Séchet, V.; Brient, L.; Savar, V.; Bardouil, M.; Hess, P.; Amzil, Z. Beta-N-methylamino-l-alanine: LC-MS/MS Optimization, Screening of Cyanobacterial Strains and Occurrence in Shellfish from Thau, a French Mediterranean Lagoon. Mar. Drugs 2014, 12, 5441–5467. [Google Scholar] [CrossRef] [PubMed]
- Réveillon, D.; Abadie, E.; Séchet, V.; Masseret, E.; Hess, P.; Amzil, Z. β-N-methylamino-l-alanine (BMAA) and Isomers: Distribution in Different Food Web Compartments of Thau Lagoon, French Mediterranean Sea. Mar. Environ. Res. 2015, 110, 8–18. [Google Scholar] [CrossRef] [PubMed]
- Réveillon, D.; Séchet, V.; Hess, P.; Amzil, Z. Production of BMAA and DAB by Diatoms (Phaeodactylum tricornutum, Chaetoceros sp., Chaetoceros calcitrans and Thalassiosira pseudonana) and Bacteria Isolated from a Diatom Culture. Harmful Algae 2016, 58, 45–50. [Google Scholar] [CrossRef] [PubMed]
- Lage, S.; Costa, P.R.; Moita, T.; Eriksson, J.; Rasmussen, U.; Rydberg, S.J. BMAA in Shellfish from Two Portuguese Transitional Water Bodies Suggests the Marine Dinoflagellate Gymnodinium Catenatum as a Potential BMAA Source. Aquat. Toxicol. 2014, 152, 131–138. [Google Scholar] [CrossRef] [PubMed]
- Mondo, K.; Broc Glover, W.; Murch, S.J.; Liu, G.; Cai, Y.; Davis, D.A.; Mash, D.C. Environmental Neurotoxins β-N-Methylamino-l-Alanine (BMAA) and Mercury in Shark Cartilage Dietary Supplements. Food Chem. Toxicol. 2014, 70, 26–32. [Google Scholar] [CrossRef] [PubMed]
- Salomonsson, M.L.; Hansson, A.; Bondesson, U. Development and in-house Validation of a Method for Quantification of BMAA in Mussels Using Dansyl Chloride Derivatization and Ultra Performance Liquid Chromatography Tandem Mass Spectrometry. Anal. Methods 2013, 5, 4865. [Google Scholar] [CrossRef]
- Salomonsson, M.L.; Fredriksson, E.; Alfjorden, A.; Hedeland, M.; Bondesson, U. Seafood sold in Sweden contains BMAA: A Study of Free and Total Concentrations with UHPLC-MS/MS and Dansyl Chloride Derivatization. Toxicol. Rep. 2015, 2, 1473–1481. [Google Scholar] [CrossRef] [PubMed]
- Jiang, L.; Kiselova, N.; Rosén, J.; Ilag, L.L. Quantification of Neurotoxin BMAA (β-N-methylamino-l-alanine) in Seafood from Swedish Markets. Sci. Rep. 2014, 4, 6931. [Google Scholar] [CrossRef] [PubMed]
- Christensen, S.J.; Hemscheidt, T.K.; Trapido-Rosenthal, H.; Laws, E.A.; Bidigare, R.R. Detection and Quantification of β-methylamino-l-alanine in Aquatic Invertebrates. Limnol. Oceanogr. Methods 2012, 10, 891–898. [Google Scholar] [CrossRef]
- Réveillon, D.; Séchet, V.; Hess, P.; Amzil, Z. Systematic Detection of BMAA (β-N-methylamino-l-alanine) and DAB (2,4-diaminobutyric acid) in Mollusks Collected in Shellfish Production Areas along the French Coasts. Toxicon 2016, 110, 35–46. [Google Scholar] [CrossRef] [PubMed]
- Masseret, E.; Banack, S.; Boumédiène, F.; Abadie, E.; Brient, L.; Pernet, F.; Juntas-Morales, R.; Pageot, N.; Metcalf, J.; Cox, P.; et al. Dietary BMAA exposure in an amyotrophic lateral sclerosis cluster from southern France. PLoS ONE 2013, 8, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Esterhuizen-Londt, M.; Wiegand, C.; Downing, T.G. β-N-methylamino-l-alanine (BMAA) Uptake by the Animal Model, Daphnia Magna and Subsequent Oxidative Stress. Toxicon 2015, 100, 20–26. [Google Scholar] [CrossRef] [PubMed]
- Lürling, M.; Faassen, E.J.; Van Eenennaam, J.S. Effects of the Cyanobacterial Neurotoxin β-N-methylamino-l-alanine (BMAA) on the Survival, Mobility and Reproduction of Daphnia magna. J. Plankton Res. 2011, 33, 333–342. [Google Scholar] [CrossRef]
- Faassen, E.J.; García-Altares, M.; Mendes e Mello, M.; Lürling, M. Trans Generational Effects of the Neurotoxin BMAA on the Aquatic Grazer Daphnia Magna. Aquat. Toxicol. 2015, 168, 98–107. [Google Scholar] [CrossRef] [PubMed]
- Downing, S.; Contardo-Jara, V.; Pflugmacher, S.; Downing, T.G. The Fate of the Cyanobacterial Toxin β-N-methylamino-l-alanine in Freshwater Mussels. Ecotoxicol. Environ. Saf. 2014, 101, 51–58. [Google Scholar] [CrossRef] [PubMed]
- Baptista, M.S.; Vasconcelos, R.G.W.; Ferreira, P.C.; Almeida, C.M.R.; Vasconcelos, V.M. Assessment of the Non-protein Amino Acid BMAA in Mediterranean mussel Mytilus Galloprovincialis after Feeding with Estuarine Cyanobacteria. Environ. Sci. Pollut. Res. 2015, 22, 12501–12510. [Google Scholar] [CrossRef] [PubMed]
- Son, M. Native Range of the Zebra Mussel and Quagga Mussel and New Data on Their Invasions within the Ponto-Caspian Region. Aquat. Invasions 2007, 2, 174–184. [Google Scholar] [CrossRef]
- Oehlmann, J.; Schulte-Oehlmann, U. Molluscs as bioindicators. Trace Met. other Contam. Environ. 2003, 6, 577–635. [Google Scholar] [CrossRef]
- White, J.D.; Sarnelle, O. Size-structured Vulnerability of the Colonial Cyanobacterium, Microcystis Aeruginosa, to Grazing by Zebra Mussels (Dreissena polymorpha). Freshw. Biol. 2014, 59, 514–525. [Google Scholar] [CrossRef]
- Aldridge, D.C.; Elliott, P.; Moggridge, G.D. Microencapsulated Biobulletse for the Control of Biofouling Zebra Mussels. Environ. Sci. Technol. 2006, 40, 975–979. [Google Scholar] [CrossRef] [PubMed]
- Winkel, E.H.T.; Davids, C. Food selection by Dreissena polymorpha Pallas (Mollusca: Bivalvia). Freshw. Biol. 1982, 12, 553–558. [Google Scholar] [CrossRef]
- Baines, S.B.; Fisher, N.S.; Cole, J.J. Uptake of Dissolved Organic Matter (DOM) and its Importance to Metabolic Requirements of the Zebra Mussel, Dreissena polymorpha. Limnol. Oceanogr. 1996, 50, 36–47. [Google Scholar] [CrossRef]
- Binelli, A.; Della Torre, C.; Magni, S.; Parolini, M. Does Zebra Mussel (Dreissena polymorpha) Represent the Freshwater Counterpart of Mytilus in Ecotoxicological Studies? A Critical Review. Environ. Pollut. 2015, 196, 386–403. [Google Scholar] [CrossRef] [PubMed]
- Cheng, T.C.; Bivalves, R. Invertebrate Blood Cells; Ratcliffe, N.A., Rowley, A.F., Eds.; Academic Press Inc.: New York, NY, USA, 1981; pp. 233–300. ISBN 0-12-582101-8. [Google Scholar]
- Mydlarz, L.D.; Jones, L.E.; Harvell, C.D. Innate Immunity, Environmental and Disease Ecology of Marine and Freshwater Invertebrates. Annu. Rev. Ecol. Evol. Syst. 2006, 37, 251–288. [Google Scholar] [CrossRef]
- Donaghy, L.; Lambert, C.; Choi, K.-S.; Soudant, P. Hemocytes of the Carpet Shell Clam (Ruditapes decussatus) and the Manila Clam (Ruditapes philippinarum): Current Knowledge and Future Prospects. Aquaculture 2009, 297, 10–24. [Google Scholar] [CrossRef]
- Lance, E.; Arnich, N.; Maignien, T.; Biré, R. Occurrence of β-N-methylamino-l-alanine (BMAA) and Isomers in Aquatic Environments and Aquatic Food Sources for Humans. Toxins 2018, 10, 83. [Google Scholar] [CrossRef] [PubMed]
- Roy-Lachapelle, A.; Solliec, M.; Sauvé, S. Determination of BMAA and Three Alkaloid Cyanotoxins in Lake Water Using Dansyl Chloride Derivatization and High-resolution Mass Spectrometry. Anal. Bioanal. Chem. 2015, 407, 5487–5501. [Google Scholar] [CrossRef] [PubMed]
- Baldwin, B.S.; Mayer, M.S.; Dayton, J.; Pau, N.; Mendilla, J.; Sullivan, M.; Moore, A.; Ma, A.; Mills, E.L. Comparative Growth and Feeding in Zebra and Quagga Mussels (Dreissena polymorpha and Dreissena bugensis): Implications for North American Lakes. Can. J. Fish. Aquat. Sci. 2002, 59, 680–694. [Google Scholar] [CrossRef]
- Faassen, E.J. Extraction and LC-MS/MS Analysis of Underivatised BMAA. In Handbook of Cyanobacterial Monitoring and Cyanotoxin Analysis; John Wiley & Sons, Ltd.: Chichester, UK, 2017; pp. 439–446. [Google Scholar]
- Souba, W.W.; Pacitti, A.J. Review: How Amino Acids Get Into Cells: Mechanisms, Models, Menus, and Mediators. J. Parenter. Enter. Nutr. 1992, 16, 569–578. [Google Scholar] [CrossRef] [PubMed]
- Smith, Q.R.; Nagura, H.; Takada, Y.; Duncan, M.W. Facilitated Transport of the Neurotoxin, Beta-N-methylamino-l-alanine, Across the Blood-brain Barrier. J. Neurochem. 1992, 58, 1330–1337. [Google Scholar] [CrossRef] [PubMed]
- Albano, R.; Lobner, D. Transport of BMAA into Neurons and Astrocytes by System x c. Neurotox. Res. 2017, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Novak, M.; Hercog, K.; Žegura, B. Assessment of the Mutagenic and Genotoxic Activity of Cyanobacterial Toxin Beta-N-methyl-amino-l-alanine in Salmonella typhimurium. Toxicon 2016, 118, 134–140. [Google Scholar] [CrossRef] [PubMed]
- Potjewyd, G.; Day, P.J.; Shangula, S.; Margison, G.P.; Povey, A.C. l-β-N-methylamino-l-alanine (BMAA) Nitrosation Generates a Cytotoxic DNA Damaging Alkylating Agent: An Unexplored Mechanism for Neurodegenerative Disease. Neurotoxicology 2017, 59, 105–109. [Google Scholar] [CrossRef] [PubMed]
- Chiu, A.S.; Gehringer, M.M.; Braidy, N.; Guillemin, G.J.; Welch, J.H.; Neilan, B.A. Gliotoxicity of the Cyanotoxin, β-methyl-amino-l-alanine (BMAA). Sci. Rep. 2013, 3, 1482. [Google Scholar] [CrossRef] [PubMed]
- Contardo-Jara, V.; Otterstein, S.K.B.; Downing, S.; Downing, T.G.; Pflugmacher, S. Response of Antioxidant and Biotransformation Systems of Selected Freshwater Mussels (Dreissena polymorpha, Anadonta cygnea, Unio tumidus, and Corbicula javanicus) to the Cyanobacterial Neurotoxin β-N-methylamino-l-alanine. Toxicol. Environ. Chem. 2014, 96, 451–465. [Google Scholar] [CrossRef]
- Barzilai, A.; Yamamoto, K.I. DNA Damage Responses to Oxidative Stress. DNA Repair 2004, 3, 1109–1115. [Google Scholar] [CrossRef] [PubMed]
- Smith, J.A.; Weidemann, M.J. Further Characterization of the Neutrophil Oxidative Burst by Flow Cytometry. J. Immunol. Methods 1993, 162, 261–268. [Google Scholar] [CrossRef]
- Shigenaga, M.K.; Ames, B.N. Assays for 8-hydroxy-2′-deoxyguanosine: A Biomarker of in vivo Oxidative DNA Damage. Free Radic. Biol. Med. 1991, 10, 211–216. [Google Scholar] [CrossRef]
- Karlsson, O.; Jiang, L.; Andersson, M.; Ilag, L.L.; Brittebo, E.B. Protein Association of the Neurotoxin and Non-protein Amino Acid BMAA (β-N-methylamino-l-alanine) in the Liver and Brain Following Neonatal Administration in Rats. Toxicol. Lett. 2014, 226, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Rao, R.V.; Bredesen, D.E. Misfolded Proteins, Endoplasmic Reticulum Stress and Neurodegeneration. Curr. Opin. Cell Biol. 2004, 16, 653–662. [Google Scholar] [CrossRef] [PubMed]
- Rodgers, K.J. Non-protein Amino Acids and Neurodegeneration: The Enemy within. Exp. Neurol. 2014, 253, 192–196. [Google Scholar] [CrossRef] [PubMed]
- Wootton, E.C.; Dyrynda, E.A.; Ratcliffe, N.A. Bivalve immunity: Comparisons between the Marine Mussel (Mytilus edulis), the Edible Cockle (Cerastoderma edule) and the Razor-shell (Ensis siliqua). Fish Shellfish Immunol. 2003, 15, 195–210. [Google Scholar] [CrossRef]
- Shapiro, H.M. Parameters and Probes. In Practical Flow Cytometry; John Wiley & Sons, Ltd.: Chichester, UK, 1988; pp. 273–410. ISBN 9780471722731. [Google Scholar]
- Johnson, I.; Spence, M.T. Assays for Cell Viability, Their Amine-Reactive Proliferation and Function; A Guide to Fluorescent Probes and Labeling Technologies; Thermo Fisher Scientific: Shanghai, China, 2011; pp. 650–738. [Google Scholar]
- Davey, H.M.; Hexley, P. Red but not dead? Membranes of Stressed Saccharomyces Cerevisiae are Permeable to Propidium Iodide. Environ. Microbiol. 2011, 13, 163–171. [Google Scholar] [CrossRef] [PubMed]
- David, S.S.; O’Shea, V.L.; Kundu, S. Base-excision Repair of Oxidative DNA Damage. Nature 2007, 447, 941–950. [Google Scholar] [CrossRef] [PubMed]
- Villela, I.V.; de Oliveira, I.M.; da Silva, J.; Henriques, J.A.P. DNA Damage and Repair in Haemolymph Cells of Golden Mussel (Limnoperna fortunei) Exposed to Environmental Contaminants. Mutat. Res. Toxicol. Environ. Mutagen. 2006, 605, 78–86. [Google Scholar] [CrossRef] [PubMed]
- Rosén, J.; Westerberg, E.; Hellenäs, K.E.; Salomonsson, M.L. A new method for analysis of underivatized free β-methylamino-alanine: Validation and method comparison. Toxicon 2016, 121, 105–108. [Google Scholar] [CrossRef] [PubMed]
- Evariste, L.; Auffret, M.; Audonnet, S.; Alain, G.; David, E.; Brousseau, P.; Fournier, M.; Betoulle, S. Functional features of hemocyte subpopulations of the invasive mollusk species Dreissena polymorpha. Fish Shellfish Immunol. 2016, 56, 144–154. [Google Scholar] [CrossRef] [PubMed]
- Singh, N.P.; McCoy, M.T.; Tice, R.R.; Schneider, E.L. A Simple Technique for Quantitation of Low Levels of DNA Damage in Individual Cells. Exp. Cell Res. 1988, 175, 184–191. [Google Scholar] [CrossRef]





© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lepoutre, A.; Milliote, N.; Bonnard, M.; Palos-Ladeiro, M.; Rioult, D.; Bonnard, I.; Bastien, F.; Faassen, E.; Geffard, A.; Lance, E. Genotoxic and Cytotoxic Effects on the Immune Cells of the Freshwater Bivalve Dreissena polymorpha Exposed to the Environmental Neurotoxin BMAA. Toxins 2018, 10, 106. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins10030106
Lepoutre A, Milliote N, Bonnard M, Palos-Ladeiro M, Rioult D, Bonnard I, Bastien F, Faassen E, Geffard A, Lance E. Genotoxic and Cytotoxic Effects on the Immune Cells of the Freshwater Bivalve Dreissena polymorpha Exposed to the Environmental Neurotoxin BMAA. Toxins. 2018; 10(3):106. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins10030106
Chicago/Turabian StyleLepoutre, Alexandra, Nadia Milliote, Marc Bonnard, Mélissa Palos-Ladeiro, Damien Rioult, Isabelle Bonnard, Fanny Bastien, Elisabeth Faassen, Alain Geffard, and Emilie Lance. 2018. "Genotoxic and Cytotoxic Effects on the Immune Cells of the Freshwater Bivalve Dreissena polymorpha Exposed to the Environmental Neurotoxin BMAA" Toxins 10, no. 3: 106. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins10030106