Proteopeptidomic, Functional and Immunoreactivity Characterization of Bothrops moojeni Snake Venom: Influence of Snake Gender on Venom Composition

 , , and
, , and
Abstract
:
1. Introduction
2. Results and Discussion
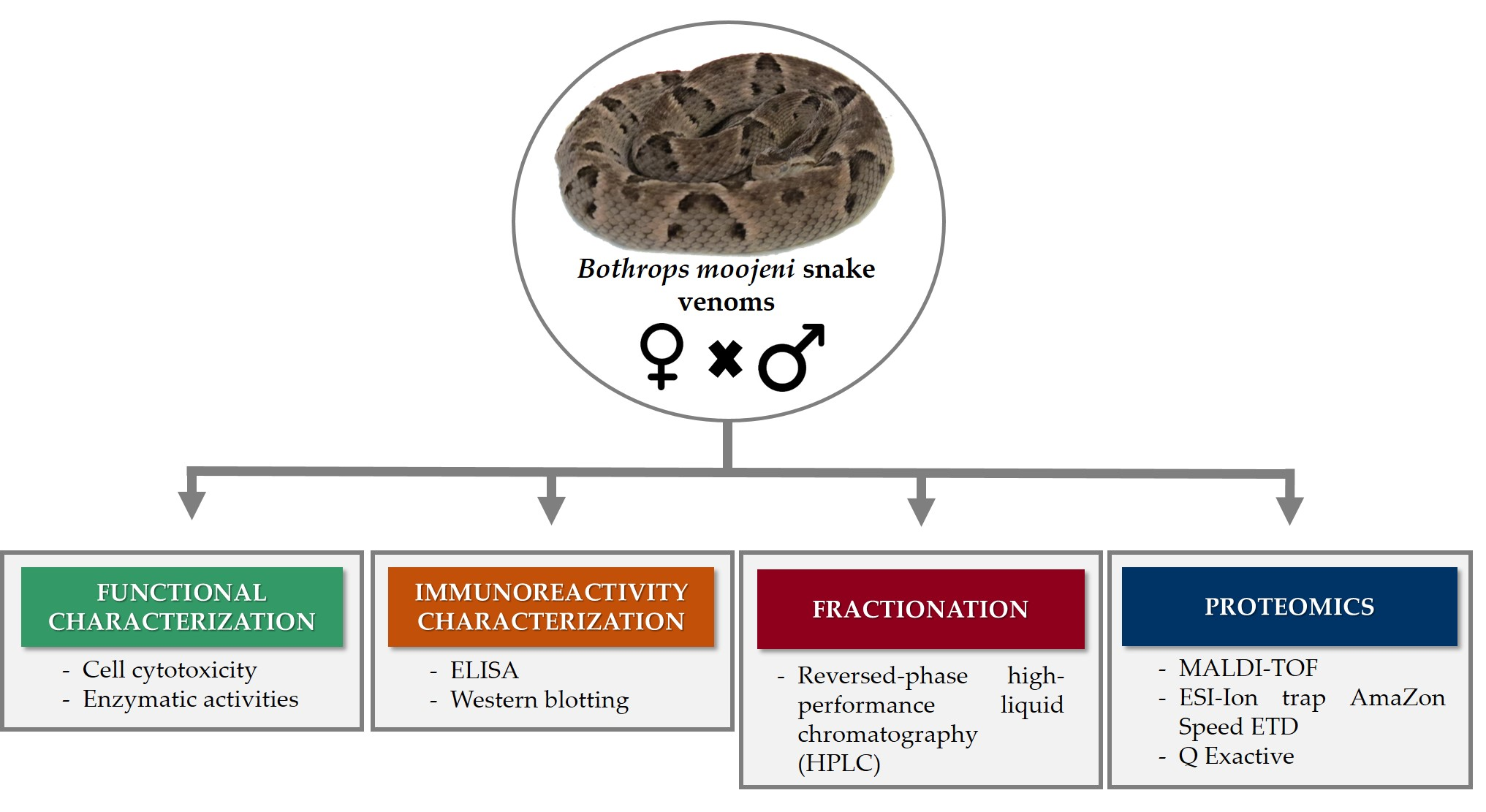
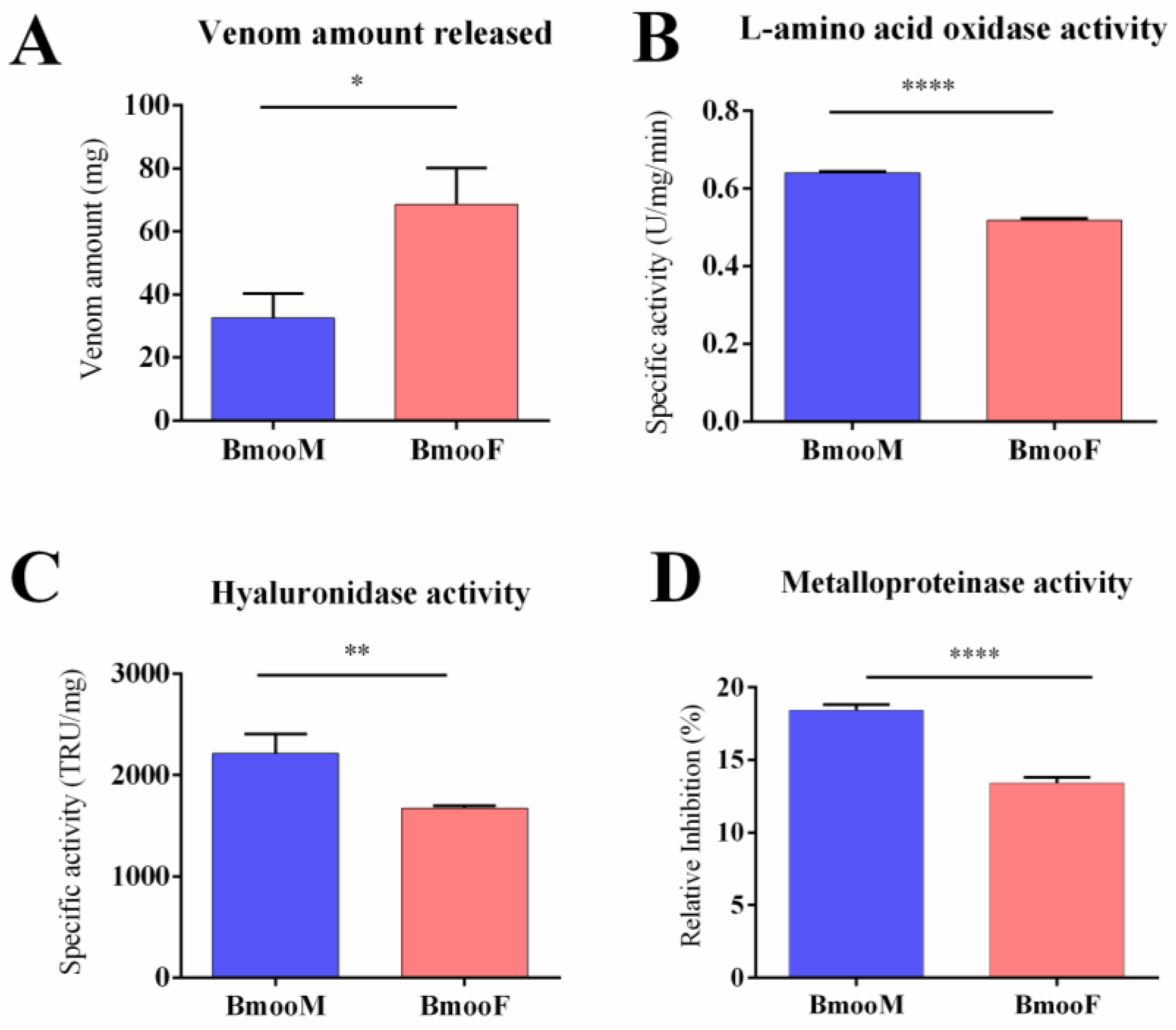
2.1. Functional and Cytotoxic Characterization
2.2. Immunoreactivity Characterization
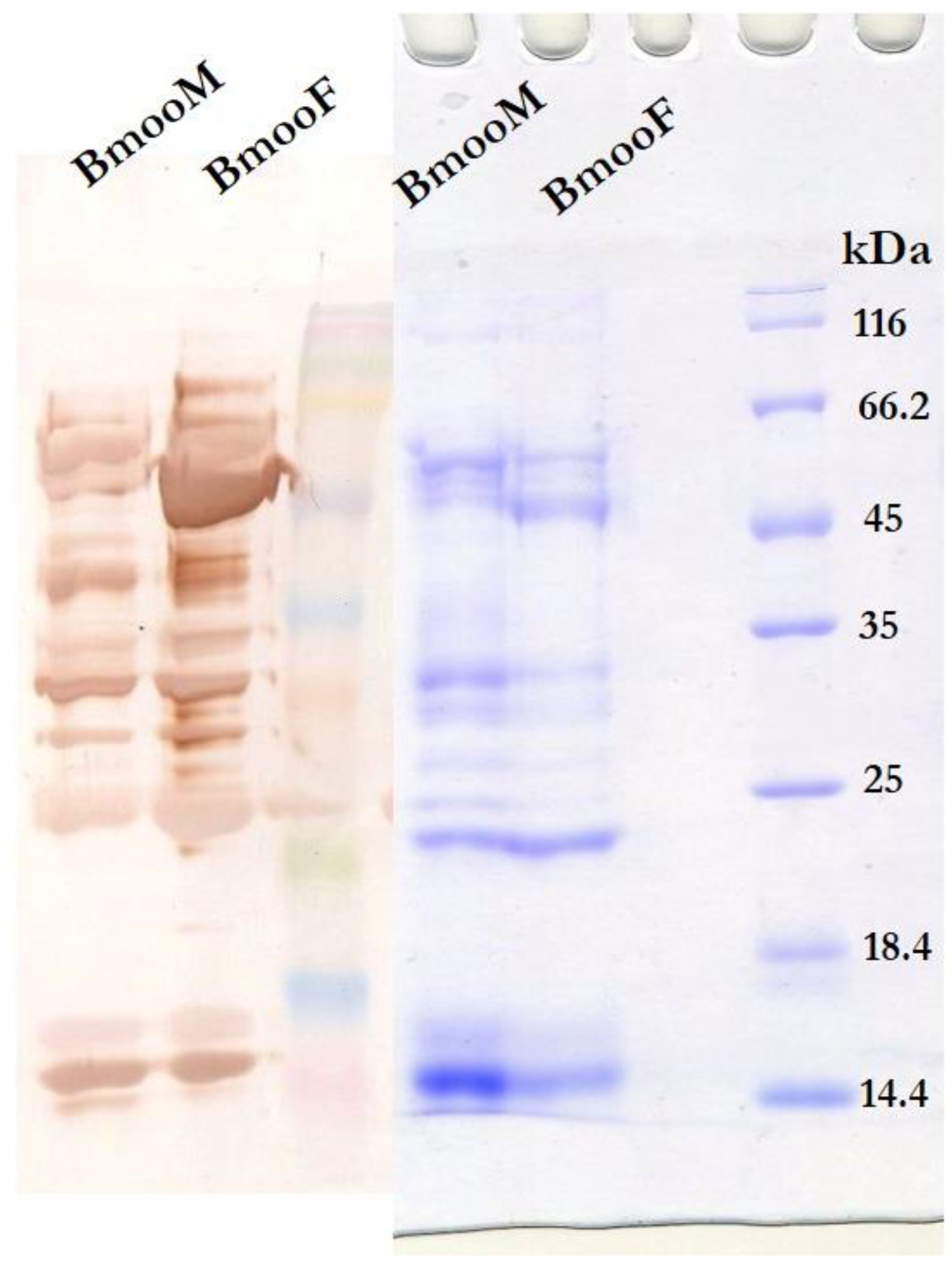
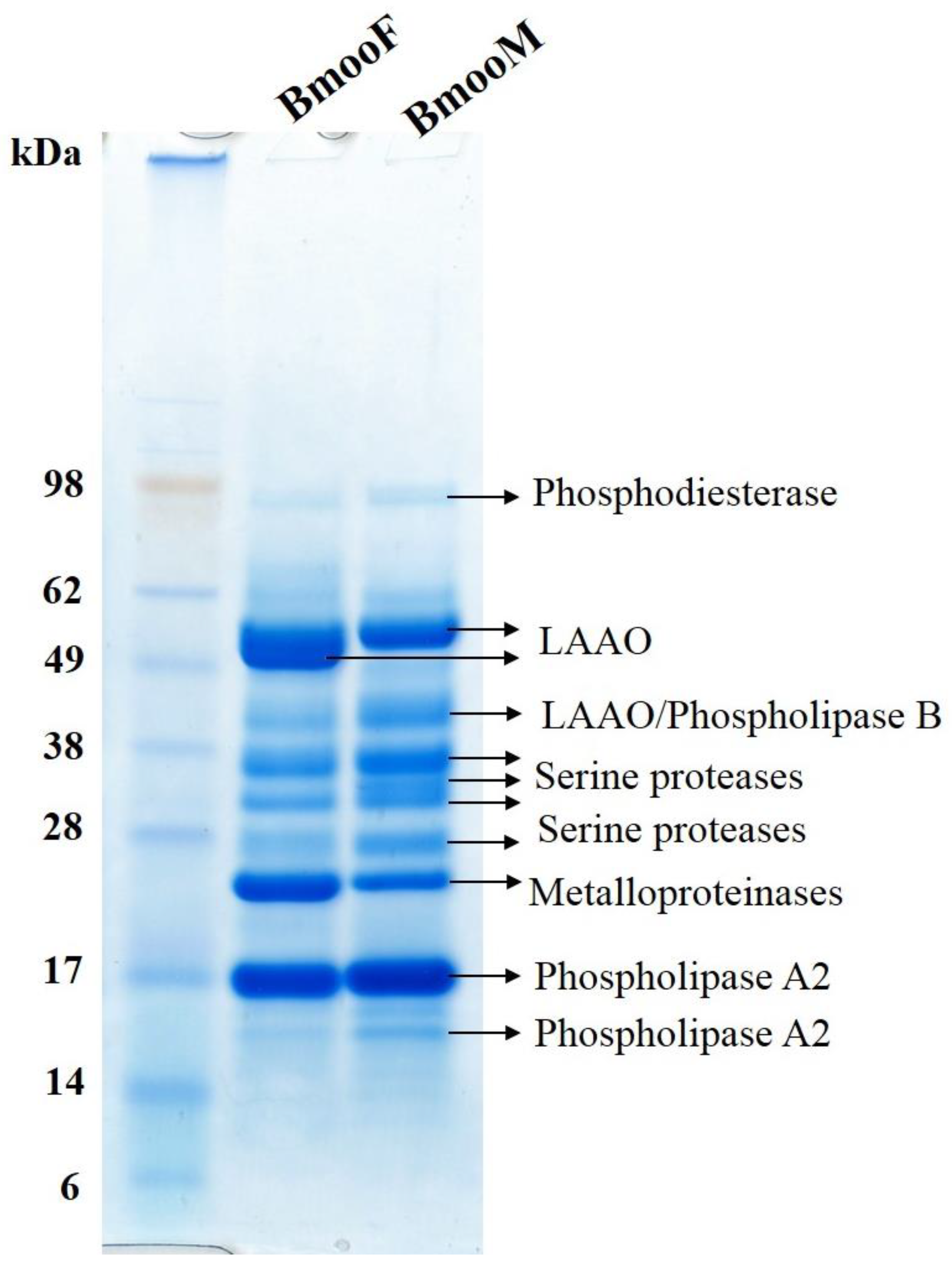
2.3. Proteomic Analyses
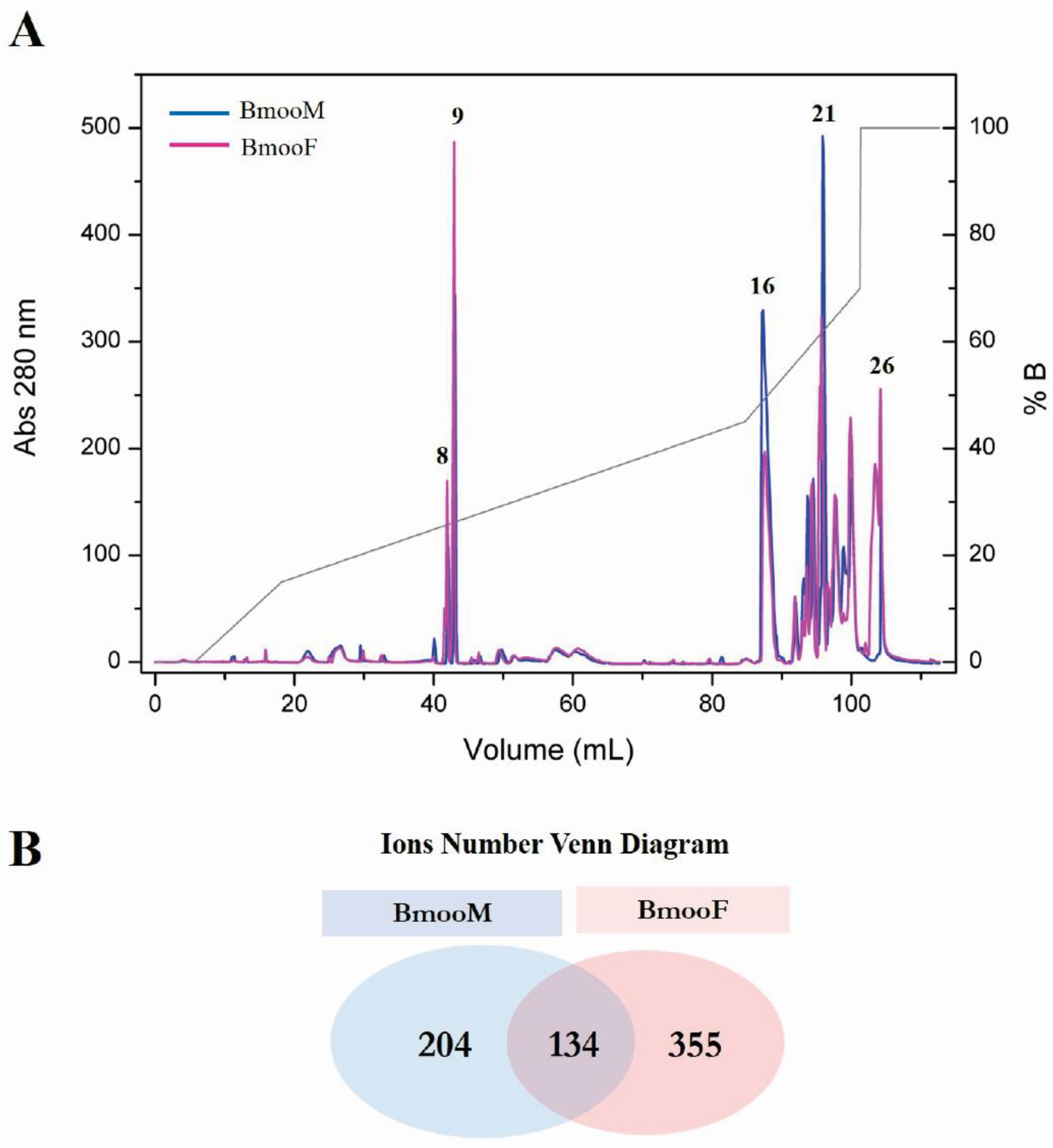
2.3.1. Fractionation by Reversed-Phase High-Performance Liquid Chromatography (HPLC) and Matrix Assisted Laser Desorption Ionization-Time of Flight (MALDI-TOF) Mass Spectrometry
2.3.2. Analysis in the AmaZon Speed electron transfer dissociation (ETD) Ion Trap Mass Spectrometer
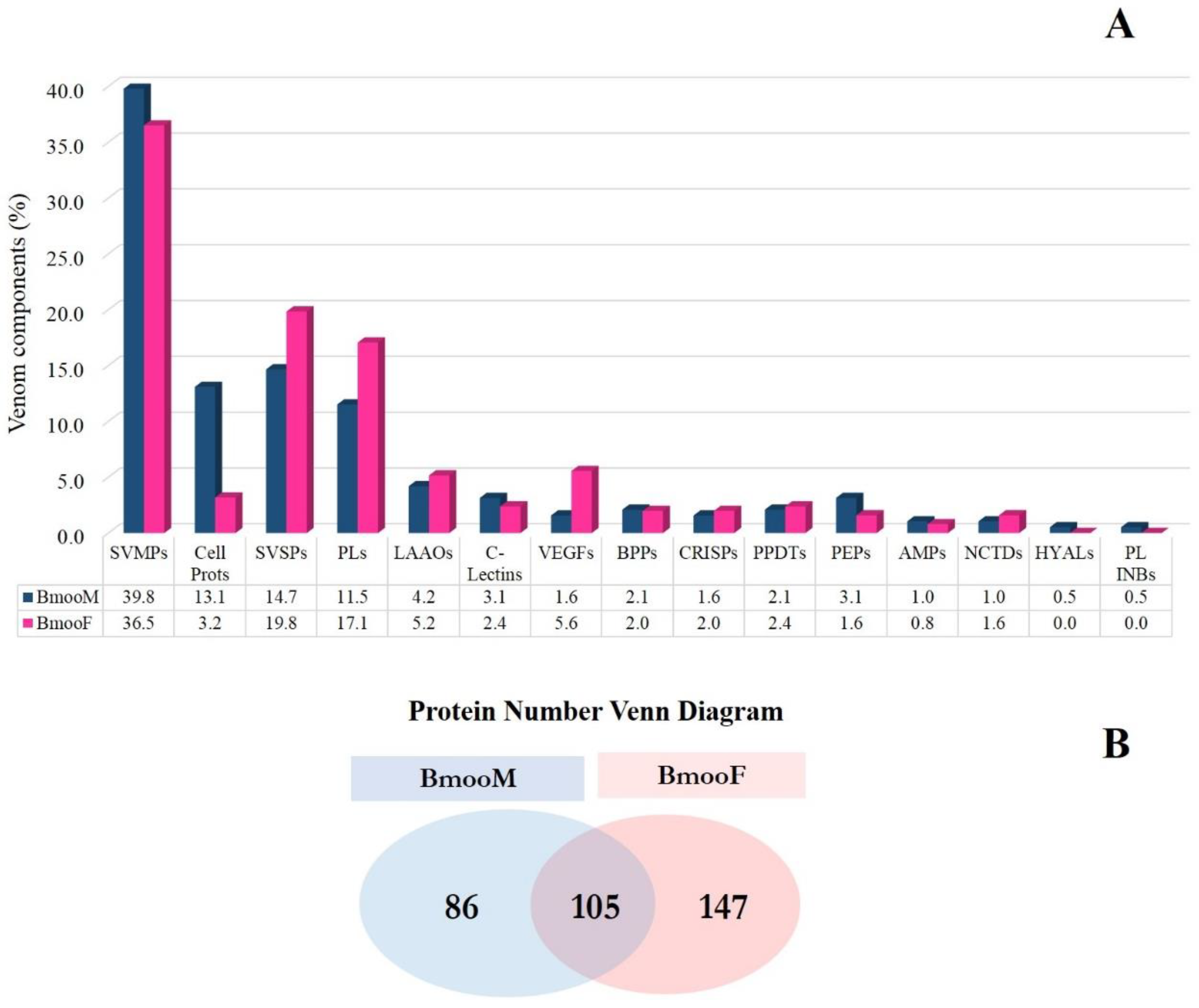
2.3.3. Shotgun Proteomics on Q-Exactive Orbitrap Mass Spectrometer
Analysis Against the Transcriptome Database
Analysis Against Uniprot Database
3. Discussion
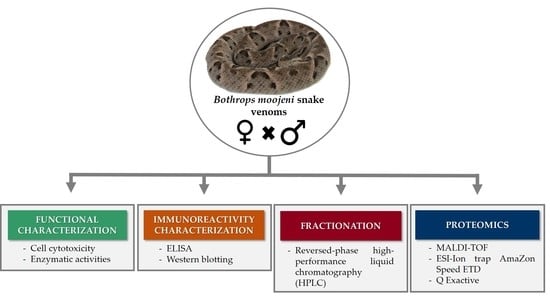
4. Methods
4.1. Chemicals and Materials
4.2. Venoms
4.3. Functional Characterization
4.3.1. L-Amino Acid Oxidase (LAAO) Activity
4.3.2. Hyaluronidase Activity
4.3.3. Metalloproteinase Activity
4.3.4. Phospholipase Activity
4.3.5. Serine Protease Activity
4.4. Cytotoxicity Assay
4.5. Immunoreactivity Characterization
4.5.1. Western Blotting
4.5.2. Enzyme-Linked Immunosorbent Assay (ELISA)
4.6. Statistical Analyses
4.7. Fractionation of Crude Venom by Reversed-Phase High-Performance Liquid Chromatography (HPLC)
4.8. Proteomics Analyses
4.8.1. Matrix Assisted Laser Desorption Ionization-Time of Flight (MALDI-TOF) Mass Spectrometry
4.8.2. Analysis in the AmaZon Speed ETD Ion Trap Mass Spectrometer
4.8.3. Shotgun Proteomics Using the Q-Exactive Orbitrap Mass Spectrometer
Supplementary Materials
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Calvete, J.J.; Sanz, L.; Angulo, Y.; Lomonte, B.; Gutiérrez, J.M. Venoms, venomics, antivenomics. FEBS Lett. 2009, 583, 1736–1743. [Google Scholar] [CrossRef] [PubMed]
- Von Reumont, B.M.; Campbell, L.I.; Jenner, R.A. Quo vadis venomics? A roadmap to neglected venomous invertebrates. Toxins (Basel) 2014, 6, 3488–3551. [Google Scholar] [CrossRef] [PubMed]
- Chippaux, J.-P. Snakebite envenomation turns again into a neglected tropical disease! J. Venom. Anim. Toxins Incl. Trop. Dis. 2017, 23, 38. [Google Scholar] [CrossRef] [PubMed]
- Sartim, M.A.; Costa, T.R.; Laure, H.J.; Espíndola, M.S.; Frantz, F.G.; Sorgi, C.A.; Cintra, A.C.; Arantes, E.C.; Faccioli, L.H.; Rosa, J.C.; et al. Moojenactivase, a novel pro-coagulant piiid metalloprotease isolated from bothrops moojeni snake venom, activates coagulation factors ii and x and induces tissue factor up-regulation in leukocytes. Arch. Toxicol. 2016, 90, 1261–1278. [Google Scholar] [CrossRef] [PubMed]
- Chippaux, J.P. Incidence and mortality due to snakebite in the americas. PLoS Negl. Trop. Dis. 2017, 11, e0005662. [Google Scholar] [CrossRef] [PubMed]
- Mamede, C.C.; de Sousa, B.B.; Pereira, D.F.; Matias, M.S.; de Queiroz, M.R.; de Morais, N.C.; Vieira, S.A.; Stanziola, L.; de Oliveira, F. Comparative analysis of local effects caused by bothrops alternatus and bothrops moojeni snake venoms: Enzymatic contributions and inflammatory modulations. Toxicon 2016, 117, 37–45. [Google Scholar] [CrossRef] [PubMed]
- Nogueira, C.; Sawaya, R.J.; Martins, M. Ecology of the pitviper, bothrops moojeni, in the brazilian cerrado. J. Herpetol. 2003, 37, 653–659. [Google Scholar] [CrossRef]
- Prashanth, J.R.; Brust, A.; Jin, A.H.; Alewood, P.F.; Dutertre, S.; Lewis, R.J. Cone snail venomics: From novel biology to novel therapeutics. Future Med. Chem. 2014, 6, 1659–1675. [Google Scholar] [CrossRef] [PubMed]
- Calvete, J.J. Venomics: Integrative venom proteomics and beyond. Biochem. J. 2017, 474, 611–634. [Google Scholar] [CrossRef] [PubMed]
- Lomonte, B.; Calvete, J.J. Strategies in ‘snake venomics’ aiming at an integrative view of compositional, functional, and immunological characteristics of venoms. J. Venom. Anim. Toxins Incl. Trop. Dis. 2017, 23, 26. [Google Scholar] [CrossRef] [PubMed]
- Zancolli, G.; Baker, T.G.; Barlow, A.; Bradley, R.K.; Calvete, J.J.; Carter, K.C.; de Jager, K.; Owens, J.B.; Price, J.F.; Sanz, L.; et al. Is hybridization a source of adaptive venom variation in rattlesnakes? A test, using a crotalus scutulatus × viridis hybrid zone in southwestern new mexico. Toxins (Basel) 2016, 8, 188. [Google Scholar] [CrossRef] [PubMed]
- Dias, G.S.; Kitano, E.S.; Pagotto, A.H.; Sant’anna, S.S.; Rocha, M.M.; Zelanis, A.; Serrano, S.M. Individual variability in the venom proteome of juvenile bothrops jararaca specimens. J. Proteome Res. 2013, 12, 4585–4598. [Google Scholar] [CrossRef] [PubMed]
- Gonçalves-Machado, L.; Pla, D.; Sanz, L.; Jorge, R.J.B.; Leitão-De-Araújo, M.; Alves, M.L.M.; Alvares, D.J.; De Miranda, J.; Nowatzki, J.; de Morais-Zani, K.; et al. Combined venomics, venom gland transcriptomics, bioactivities, and antivenomics of two bothrops jararaca populations from geographic isolated regions within the brazilian atlantic rainforest. J. Proteom. 2016, 135, 73–89. [Google Scholar] [CrossRef] [PubMed]
- Sousa, L.F.; Portes-Junior, J.A.; Nicolau, C.A.; Bernardoni, J.L.; Nishiyama, M.Y., Jr.; Amazonas, D.R.; Freitas-de-Sousa, L.A.; Mourão, R.H.; Chalkidis, H.M.; Valente, R.H.; et al. Functional proteomic analyses of bothrops atrox venom reveals phenotypes associated with habitat variation in the amazon. J. Proteom. 2017, 159, 32–46. [Google Scholar] [CrossRef] [PubMed]
- Furtado, M.F.; Maruyama, M.; Kamiguti, A.S.; Antonio, L.C. Comparative study of nine bothrops snake venoms from adult female snakes and their offspring. Toxicon 1991, 29, 219–226. [Google Scholar] [CrossRef]
- Zelanis, A.; Tashima, A.K.; Rocha, M.M.; Furtado, M.F.; Camargo, A.C.; Ho, P.L.; Serrano, S.M. Analysis of the ontogenetic variation in the venom proteome/peptidome of bothrops jararaca reveals different strategies to deal with prey. J. Proteom. Res. 2010, 9, 2278–2291. [Google Scholar] [CrossRef] [PubMed]
- Zelanis, A.; Andrade-Silva, D.; Rocha, M.M.; Furtado, M.F.; Serrano, S.M.; Junqueira-de-Azevedo, I.L.; Ho, P.L. A transcriptomic view of the proteome variability of newborn and adult bothrops jararaca snake venoms. PLoS Negl. Trop. Dis. 2012, 6, e1554. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Antunes, T.C.; Yamashita, K.M.; Barbaro, K.C.; Saiki, M.; Santoro, M.L. Comparative analysis of newborn and adult bothrops jararaca snake venoms. Toxicon 2010, 56, 1443–1458. [Google Scholar] [CrossRef] [PubMed]
- Zelanis, A.; Tashima, A.K.; Pinto, A.F.; Paes Leme, A.F.; Stuginski, D.R.; Furtado, M.F.; Sherman, N.E.; Ho, P.L.; Fox, J.W.; Serrano, S.M. Bothrops jararaca venom proteome rearrangement upon neonate to adult transition. Proteomics 2011, 11, 4218–4228. [Google Scholar] [CrossRef] [PubMed]
- Pimenta, D.C.; Prezoto, B.C.; Konno, K.; Melo, R.L.; Furtado, M.F.; Camargo, A.C.; Serrano, S.M. Mass spectrometric analysis of the individual variability of bothrops jararaca venom peptide fraction. Evidence for sex-based variation among the bradykinin-potentiating peptides. Rapid Commun. Mass Spectrom. 2007, 21, 1034–1042. [Google Scholar] [CrossRef] [PubMed]
- Menezes, M.C.; Furtado, M.F.; Travaglia-Cardoso, S.R.; Camargo, A.C.; Serrano, S.M. Sex-based individual variation of snake venom proteome among eighteen bothrops jararaca siblings. Toxicon 2006, 47, 304–312. [Google Scholar] [CrossRef] [PubMed]
- Zelanis, A.; Menezes, M.C.; Kitano, E.S.; Liberato, T.; Tashima, A.K.; Pinto, A.F.; Sherman, N.E.; Ho, P.L.; Fox, J.W.; Serrano, S.M. Proteomic identification of gender molecular markers in bothrops jararaca venom. J. Proteom. 2016, 139, 26–37. [Google Scholar] [CrossRef] [PubMed]
- Furtado, M.F.; Travaglia-Cardoso, S.R.; Rocha, M.M. Sexual dimorphism in venom of bothrops jararaca(serpentes: Viperidae). Toxicon 2006, 48, 401–410. [Google Scholar] [CrossRef] [PubMed]
- Tan, C.H.; Tan, K.Y.; Yap, M.K.; Tan, N.H. Venomics of tropidolaemus wagleri, the sexually dimorphic temple pit viper: Unveiling a deeply conserved atypical toxin arsenal. Sci. Rep. 2017, 7, 43237. [Google Scholar] [CrossRef] [PubMed]
- Nicoleti, A.F.; Medeiros, C.R.; Duarte, M.R.; França, F.O. Comparison of bothropoides jararaca bites with and without envenoming treated at the vital brazil hospital of the butantan institute, state of são paulo, brazil. Rev. Soc. Bras. Med. Trop. 2010, 43, 657–661. [Google Scholar] [CrossRef] [PubMed]
- Kouyoumdjian, J.A.; Polizelli, C. Acidentes ofídicos causados por bothrops moojeni: Correlaçäo do quadro clínico com o tamanho da serpente. Rev. Inst. Med. Trop. São Paulo 1989, 31, 84–90. [Google Scholar] [CrossRef] [PubMed]
- Kouyoumdjian, J.A.; Polizelli, C. Acidentes ofídicos causados por bothrops moojeni: Relato de 37 casos. Rev. Inst. Med. Trop. São Paulo 1988, 30, 424–432. [Google Scholar] [CrossRef]
- Melani, R.D.; Goto-Silva, L.; Nogueira, F.C.S.; Junqueira, M.; Domont, G.B. Shotgun approaches for venom analysis. In Venom Genomics and Proteomics; Springer: Heidelberg/Berlin, Germany, 2016; p. 13. [Google Scholar]
- Amorim, F.G.; Morandi-Filho, R.; Fujimura, P.T.; Ueira-Vieira, C.; Sampaio, S.V. New findings from the first transcriptome of the bothrops moojeni snake venom gland. Toxicon 2017, 140, 105–117. [Google Scholar] [CrossRef] [PubMed]
- Shine, R.; Charles, N. Ecology of the australian elapid snake tropidechis carinatus. J. Herpetol. 1982, 16, 383–387. [Google Scholar] [CrossRef]
- Lourenço, A.; Zorzella Creste, C.F.; de Barros, L.C.; Delazari dos Santos, L.; Pimenta, D.C.; Barraviera, B.; Ferreira, R.S. Individual venom profiling of crotalus durissus terrificus specimens from a geographically limited region: Crotamine assessment and captivity evaluation on the biological activities. Toxicon 2013, 69, 75–81. [Google Scholar] [CrossRef] [PubMed]
- Augusto-de-Oliveira, C.; Stuginski, D.R.; Kitano, E.S.; Andrade-Silva, D.; Liberato, T.; Fukushima, I.; Serrano, S.M.; Zelanis, A. Dynamic rearrangement in snake venom gland proteome: Insights into bothrops jararaca intraspecific venom variation. J. Proteom. Res. 2016, 15, 3752–3762. [Google Scholar] [CrossRef] [PubMed]
- Young, B.A.; Lee, C.E.; Daley, K.M. Do snakes meter venom? BioScience 2002, 52, 6. [Google Scholar] [CrossRef]
- Warrell, D.A. Snake bite. Lancet 2010, 375, 77–88. [Google Scholar] [CrossRef]
- Naik, B.S. “Dry bite” In venomous snakes: A review. Toxicon 2017, 133, 63–67. [Google Scholar] [CrossRef] [PubMed]
- Bordon, K.C.; Wiezel, G.A.; Cabral, H.; Arantes, E.C. Bordonein-L, a new L-amino acid oxidase from crotalus durissus terrificus snake venom: Isolation, preliminary characterization and enzyme stability. J. Venom. Anim. Toxins Incl. Trop. Dis. 2015, 21, 26. [Google Scholar] [CrossRef] [PubMed]
- Du, X.Y.; Clemetson, K.J. Snake venom L-amino acid oxidases. Toxicon 2002, 40, 659–665. [Google Scholar] [CrossRef]
- Aloulou, A.; Ali, Y.B.; Bezzine, S.; Gargouri, Y.; Gelb, M.H. Phospholipases: An overview. Methods Mol. Biol. 2012, 861, 63–85. [Google Scholar] [PubMed]
- Boldrini-França, J.; Cologna, C.T.; Pucca, M.B.; Bordon, K.C.; Amorim, F.G.; Anjolette, F.A.; Cordeiro, F.A.; Wiezel, G.A.; Cerni, F.A.; Pinheiro-Junior, E.L.; et al. Minor snake venom proteins: Structure, function and potential applications. Biochim. Biophys. Acta 2017, 1861, 824–838. [Google Scholar] [CrossRef] [PubMed]
- Soares, A.M.; Marcussi, S.; Stábeli, R.G.; França, S.C.; Giglio, J.R.; Ward, R.J.; Arantes, E.C. Structural and functional analysis of bmjmip, a phospholipase a2 myotoxin inhibitor protein from bothrops moojeni snake plasma. Biochem. Biophys. Res. Commun. 2003, 302, 193–200. [Google Scholar] [CrossRef]
- Zainal Abidin, S.A.; Rajadurai, P.; Chowdhury, M.E.; Ahmad Rusmili, M.R.; Othman, I.; Naidu, R. Proteomic characterization and comparison of malaysian tropidolaemus wagleri and cryptelytrops purpureomaculatus venom using shotgun-proteomics. Toxins (Basel) 2016, 8, 299. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Fonslow, B.R.; Shan, B.; Baek, M.C.; Yates, J.R. Protein analysis by shotgun/bottom-up proteomics. Chem. Rev. 2013, 113, 2343–2394. [Google Scholar] [CrossRef] [PubMed]
- Link, A.J.; Eng, J.; Schieltz, D.M.; Carmack, E.; Mize, G.J.; Morris, D.R.; Garvik, B.M.; Yates, J.R. Direct analysis of protein complexes using mass spectrometry. Nat. Biotechnol. 1999, 17, 676–682. [Google Scholar] [CrossRef] [PubMed]
- Hebert, A.S.; Richards, A.L.; Bailey, D.J.; Ulbrich, A.; Coughlin, E.E.; Westphall, M.S.; Coon, J.J. The one hour yeast proteome. Mol. Cell. Proteom. 2014, 13, 339–347. [Google Scholar] [CrossRef] [PubMed]
- Thakur, S.S.; Geiger, T.; Chatterjee, B.; Bandilla, P.; Fröhlich, F.; Cox, J.; Mann, M. Deep and highly sensitive proteome coverage by lc-ms/ms without prefractionation. Mol. Cell. Proteom. 2011, 10. [Google Scholar] [CrossRef] [PubMed]
- Margres, M.J.; McGivern, J.J.; Wray, K.P.; Seavy, M.; Calvin, K.; Rokyta, D.R. Linking the transcriptome and proteome to characterize the venom of the eastern diamondback rattlesnake (crotalus adamanteus). J. Proteom. 2014, 96, 145–158. [Google Scholar] [CrossRef] [PubMed]
- Pukrittayakamee, S.; Warrell, D.A.; Desakorn, V.; McMichael, A.J.; White, N.J.; Bunnag, D. The hyaluronidase activities of some southeast asian snake venoms. Toxicon 1988, 26, 629–637. [Google Scholar] [CrossRef]
- Bordon, K.C.; Perino, M.G.; Giglio, J.R.; Arantes, E.C. Isolation, enzymatic characterization and antiedematogenic activity of the first reported rattlesnake hyaluronidase from crotalus durissus terrificus venom. Biochimie 2012, 94, 2740–2748. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.J.; Shih, C.H.; Huang, T.F. A novel p-i class metalloproteinase with broad substrate-cleaving activity, agkislysin, from agkistrodon acutus venom. Biochem. Biophys. Res. Commun. 2004, 324, 224–230. [Google Scholar] [CrossRef] [PubMed]
- Gutiérrez, J.M.; Avila, C.; Rojas, E.; Cerdas, L. An alternative in vitro method for testing the potency of the polyvalent antivenom produced in costa rica. Toxicon 1988, 26, 411–413. [Google Scholar] [CrossRef]
- Hummel, B.C. A modified spectrophotometric determination of chymotrypsin, trypsin, and thrombin. Can. J. Biochem. Physiol. 1959, 37, 1393–1399. [Google Scholar] [CrossRef] [PubMed]
- Mosmann, T. Rapid colorimetric assay for cellular growth and survival: Application to proliferation and cytotoxicity assays. J. Immunol. Methods 1983, 65, 55–63. [Google Scholar] [CrossRef]
- Costa, T.R.; Menaldo, D.L.; Prinholato da Silva, C.; Sorrechia, R.; de Albuquerque, S.; Pietro, R.C.; Ghisla, S.; Antunes, L.M.; Sampaio, S.V. Evaluating the microbicidal, antiparasitic and antitumor effects of cr-laao from calloselasma rhodostoma venom. Int. J. Biol. Macromol. 2015, 80, 489–497. [Google Scholar] [CrossRef] [PubMed]
- Laemmli, U.K.; Beguin, F.; Gujer-Kellenberger, G. A factor preventing the major head protein of bacteriophage t4 from random aggregation. J. Mol. Biol. 1970, 47, 69–85. [Google Scholar] [CrossRef]
- Bernardes, C.P.; Menaldo, D.L.; Camacho, E.; Rosa, J.C.; Escalante, T.; Rucavado, A.; Lomonte, B.; Gutiérrez, J.M.; Sampaio, S.V. Proteomic analysis of bothrops pirajai snake venom and characterization of bpirmp, a new P-I metalloproteinase. J. Proteom. 2013, 80, 250–267. [Google Scholar] [CrossRef] [PubMed]
- Ma, B.; Zhang, K.; Hendrie, C.; Liang, C.; Li, M.; Doherty-Kirby, A.; Lajoie, G. Peaks: Powerful software for peptide de novo sequencing by tandem mass spectrometry. Rapid Commun. Mass Spectrom. 2003, 17, 2337–2342. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Accession | −10lgP | Coverage (%) | Number of Peptides | Number of Unique Peptides | Average Mass | Description |
|---|---|---|---|---|---|---|
| ATU85535.1 | 489.98 | 93 | 316 | 316 | 56,840 | L-amino acid oxidase |
| ATU85523.1 | 426.4 | 94 | 164 | 164 | 24,669 | Snake venom metalloproteinase BmooMPalpha-I-like isoform |
| ATU85528.1 | 376.13 | 88 | 71 | 71 | 15,699 | Basic phospholipase A2 myotoxin |
| ATU85541.1 | 334.57 | 85 | 35 | 35 | 26,837 | Cysteine-rich secretory protein Moojin |
| ATU85526.1 | 322.31 | 54 | 43 | 43 | 63,875 | Phospholipase B |
| ATU85531.1 | 272.92 | 61 | 23 | 23 | 16,334 | Snake venom vascular endothelial growth factor toxin |
| ATU85542.1 | 172.96 | 16 | 7 | 7 | 52,550 | Hyaluronidase BmooHyal-1 |
| ATU85551.1 | 172.41 | 38 | 4 | 4 | 9767 | Waprin 1 |
| ATU85533.1 | 109.82 | 30 | 5 | 4 | 16,657 | C-type lectin isoform 1 |
| Statistics Data | ||
| BmooM | BmooF | |
| Number of MS scans | 6676 | 6586 |
| Number of MS/MS scans | 42,843 | 39,953 |
| Statistics of filtered result | ||
| Peptide-Spectrum Matches | 12,453 | 9421 |
| Peptide Sequences | 3148 | 3349 |
| Protein Groups | 142 | 206 |
| Proteins | 191 | 252 |
| Proteins (number of Unique Peptides) | 103 (>2); 88 (=2); 0 (=1) | 149 (>2); 103 (=2); 0 (=1) |
| De novo Only Spectra | 1445 | 1040 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Amorim, F.G.; Costa, T.R.; Baiwir, D.; De Pauw, E.; Quinton, L.; Sampaio, S.V. Proteopeptidomic, Functional and Immunoreactivity Characterization of Bothrops moojeni Snake Venom: Influence of Snake Gender on Venom Composition. Toxins 2018, 10, 177. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins10050177
Amorim FG, Costa TR, Baiwir D, De Pauw E, Quinton L, Sampaio SV. Proteopeptidomic, Functional and Immunoreactivity Characterization of Bothrops moojeni Snake Venom: Influence of Snake Gender on Venom Composition. Toxins. 2018; 10(5):177. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins10050177
Chicago/Turabian StyleAmorim, Fernanda Gobbi, Tassia Rafaela Costa, Dominique Baiwir, Edwin De Pauw, Loic Quinton, and Suely Vilela Sampaio. 2018. "Proteopeptidomic, Functional and Immunoreactivity Characterization of Bothrops moojeni Snake Venom: Influence of Snake Gender on Venom Composition" Toxins 10, no. 5: 177. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins10050177