Accumulation and Biotransformation of Dinophysis Toxins by the Surf Clam Mesodesma donacium

 , ,
, ,
Abstract
:1. Introduction
2. Results
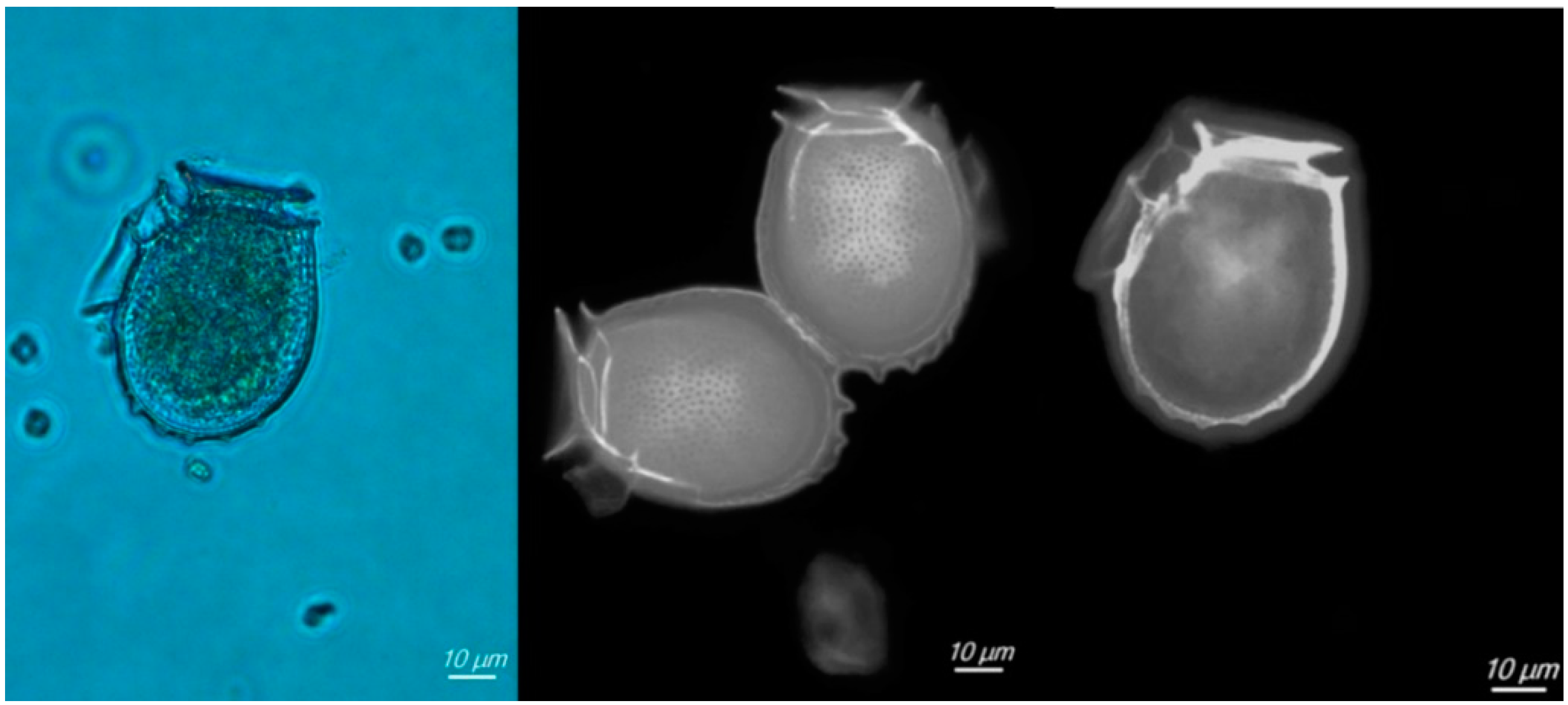
2.1. Abundance and Composition of Dinophysis Populations
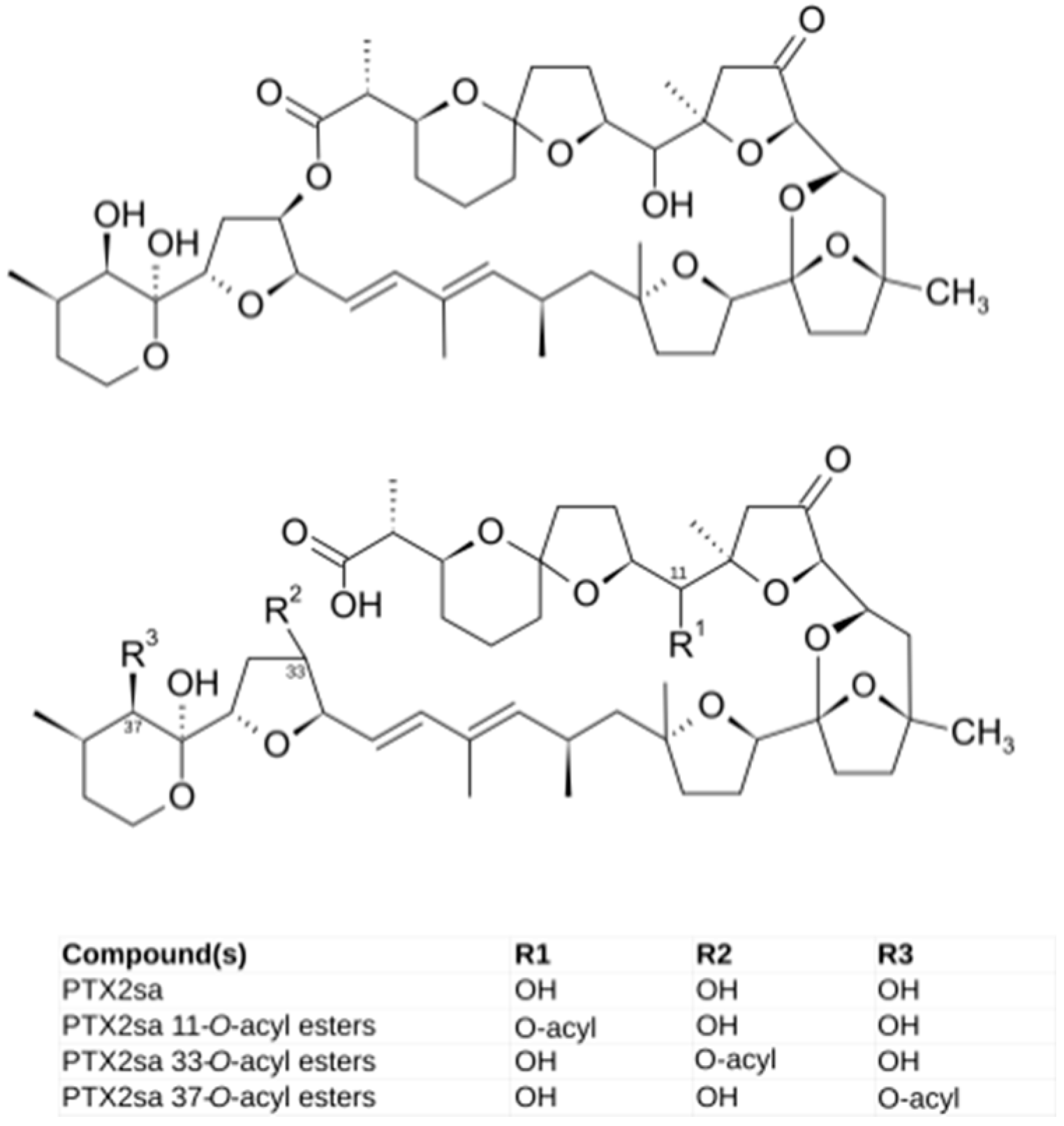
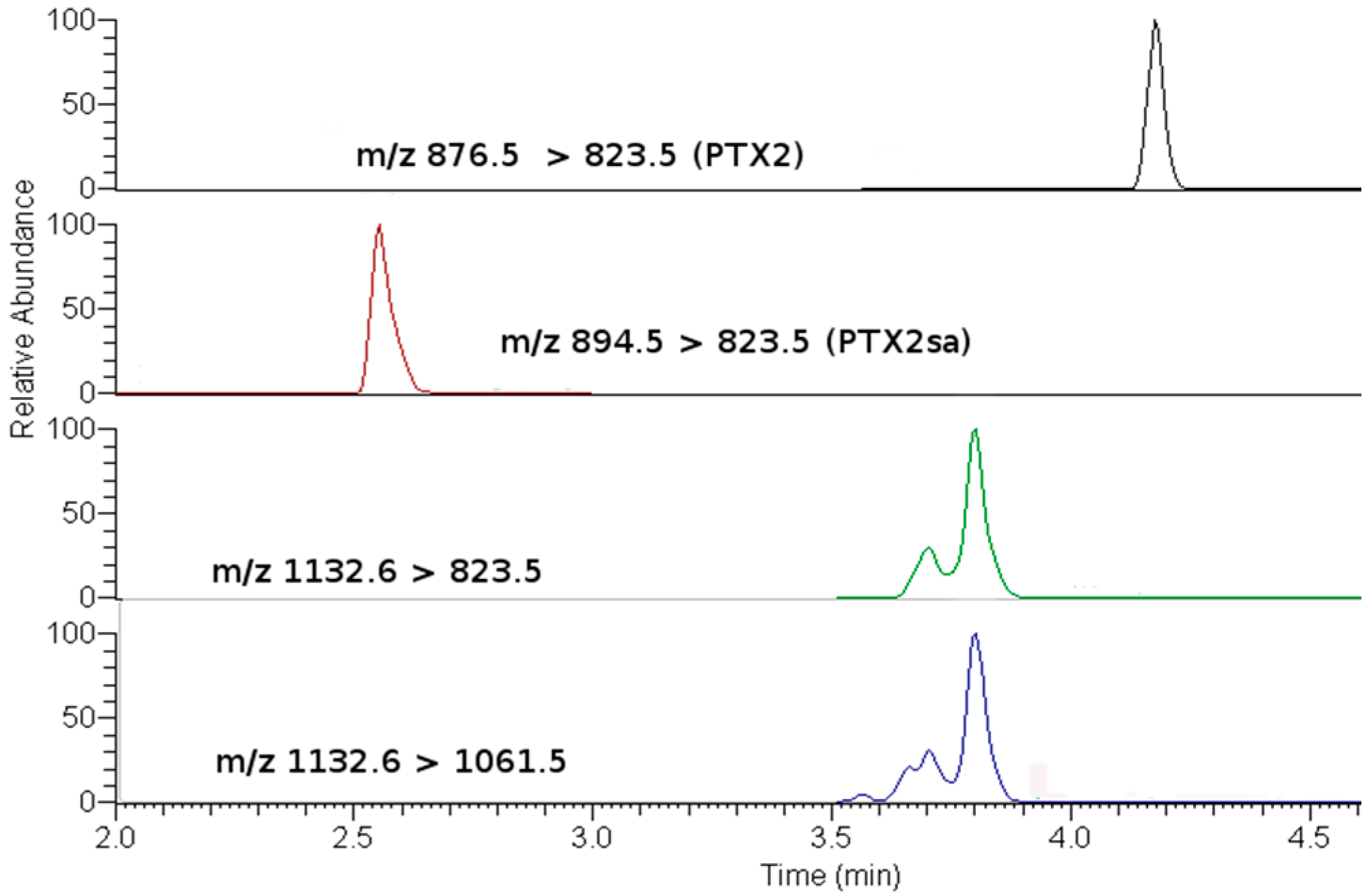
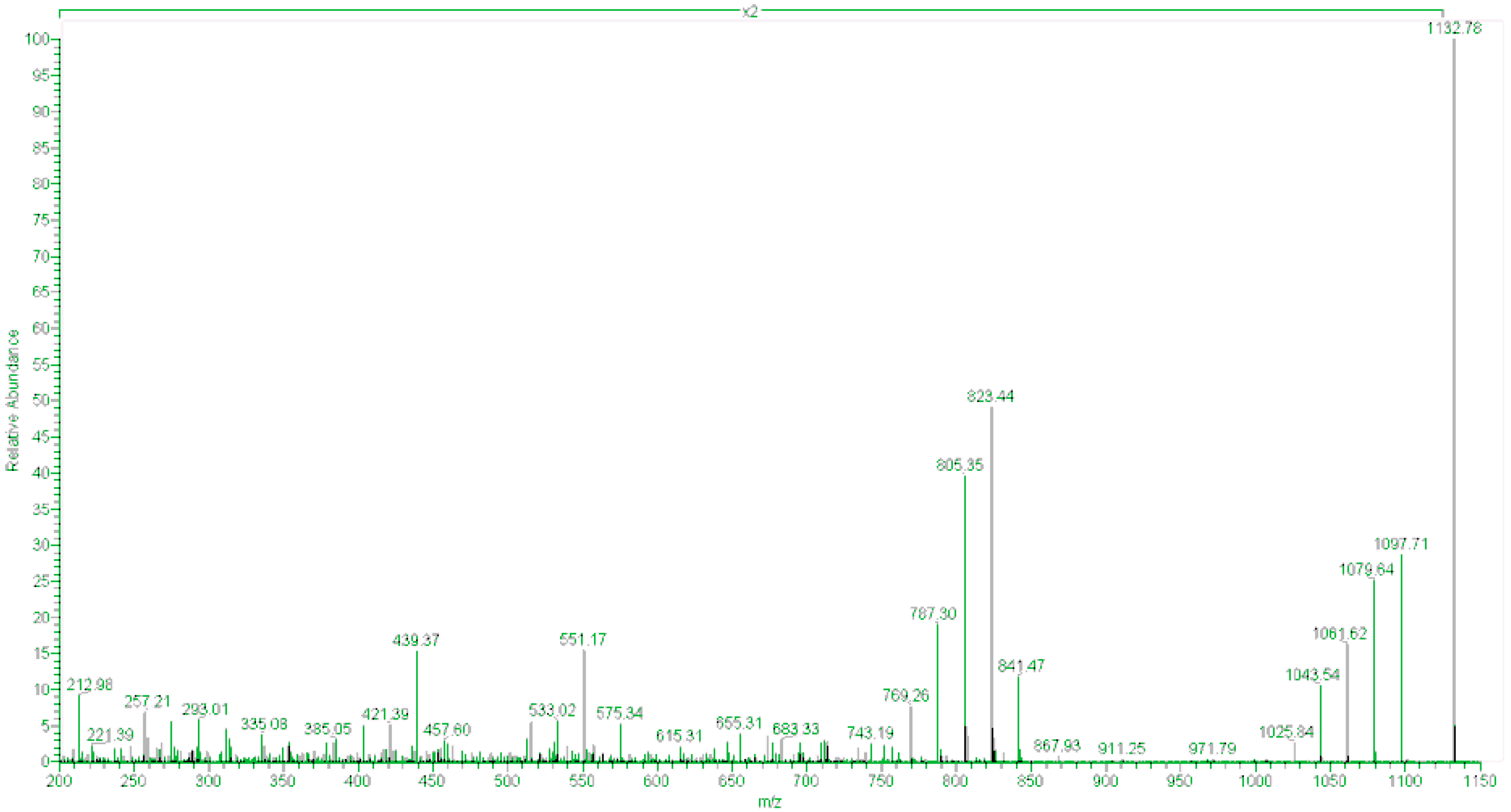
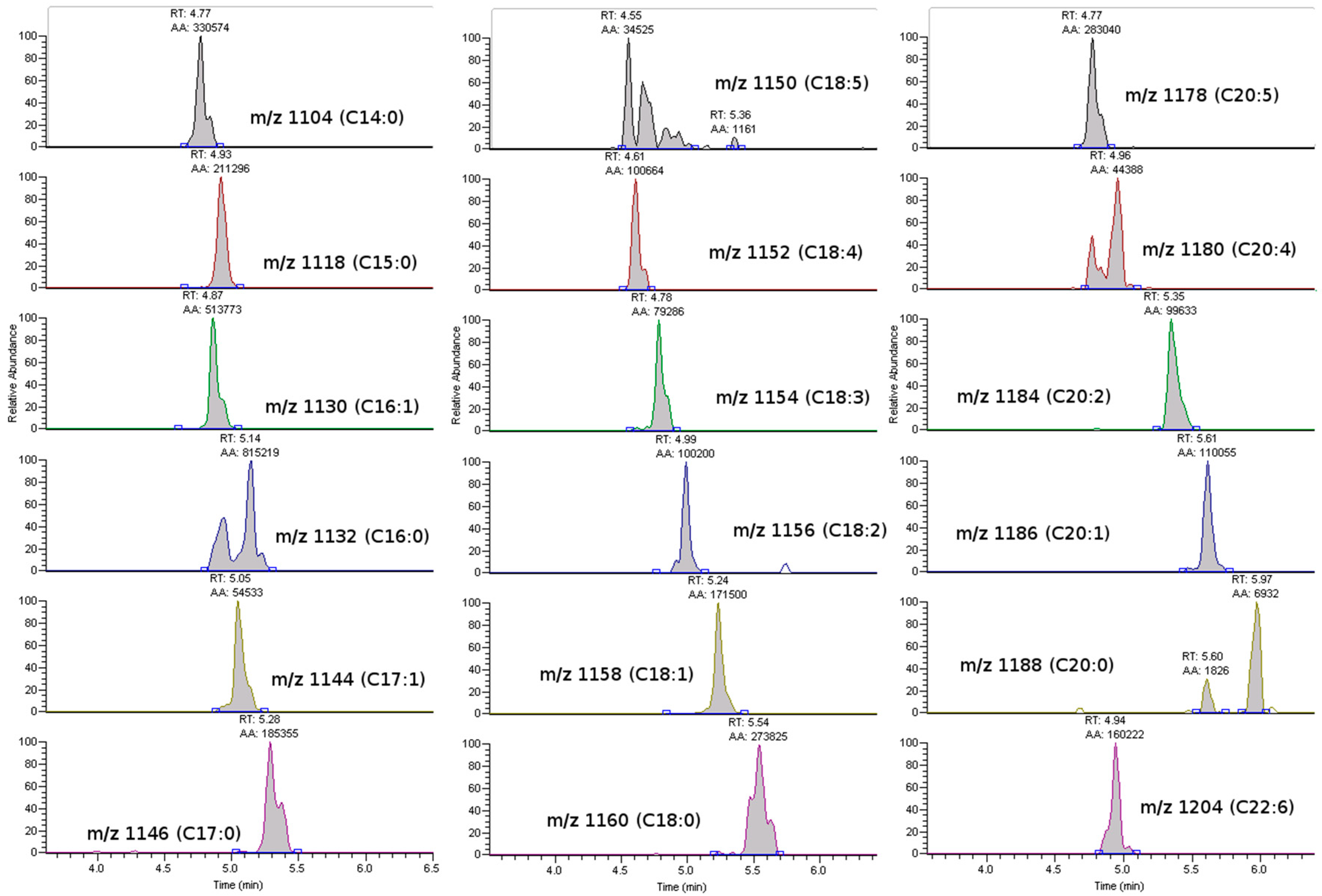
2.2. Toxin Profiles
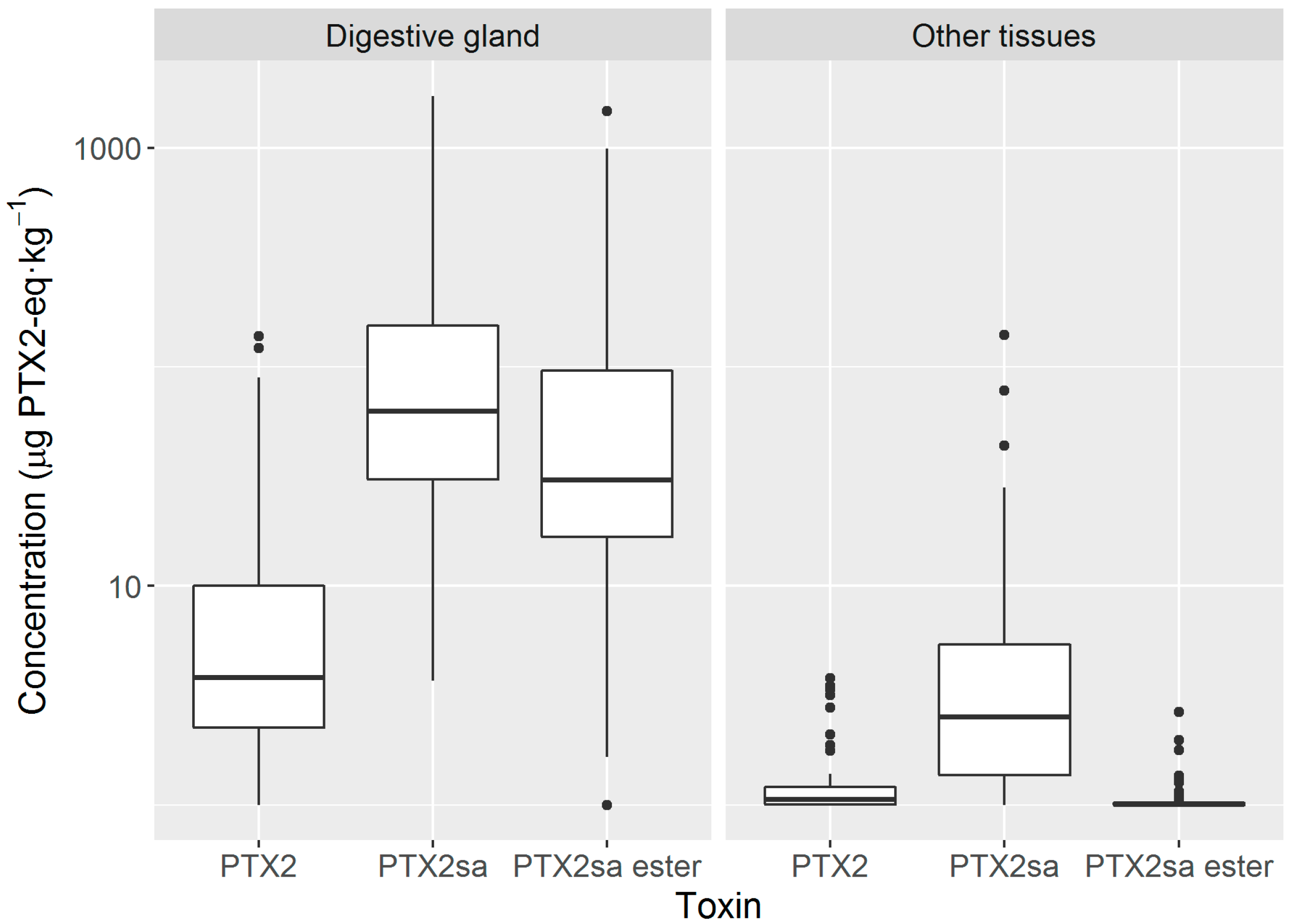
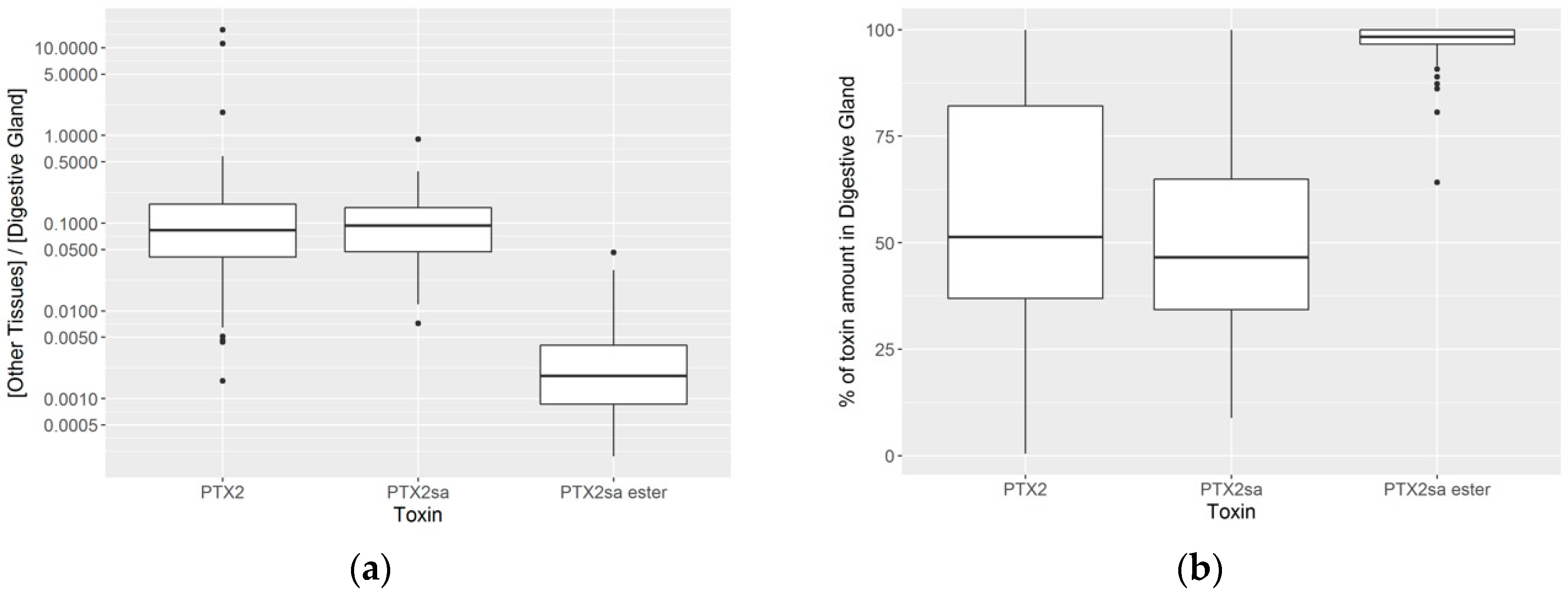
2.3. Anatomical Distribution of Toxins
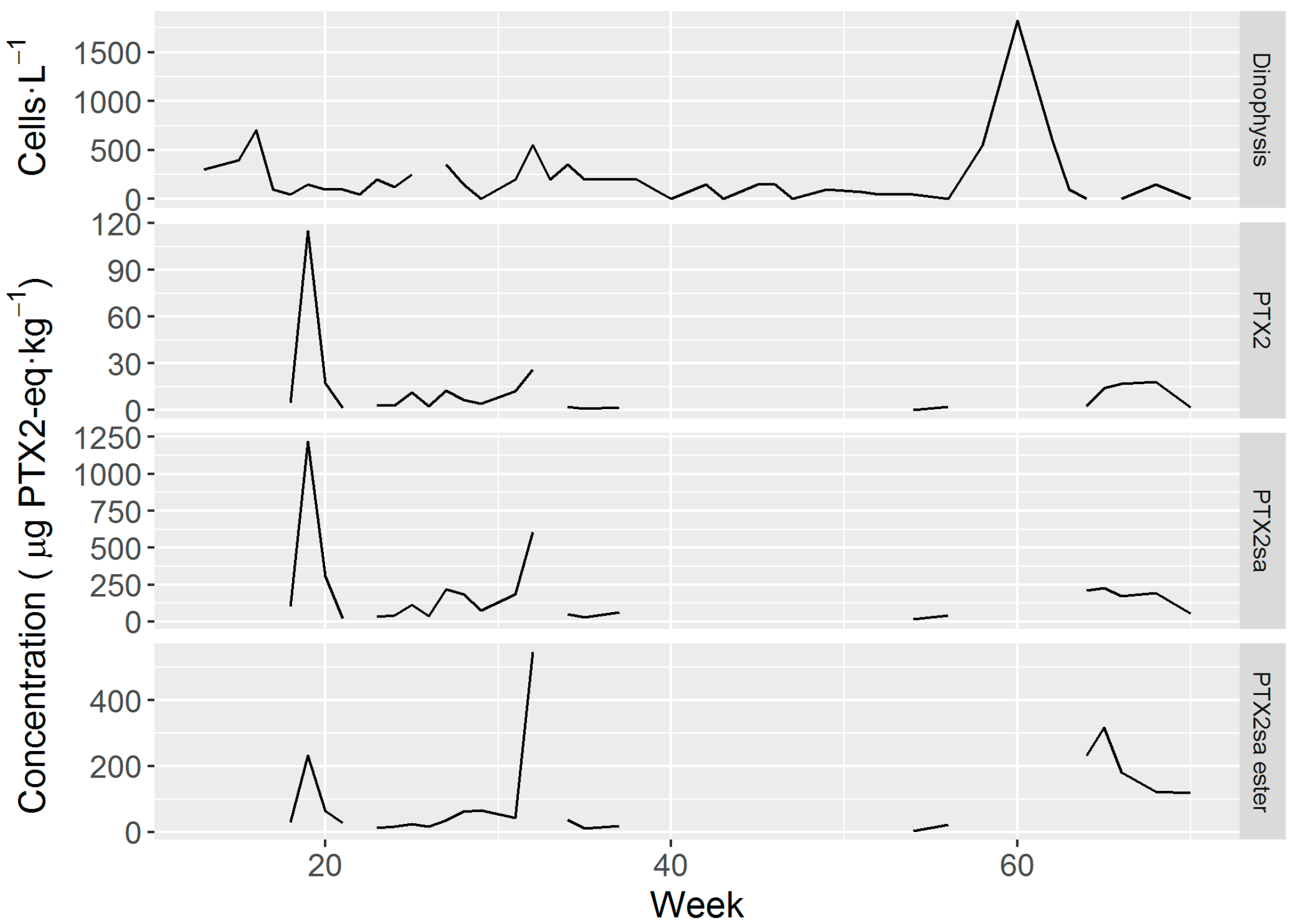
2.4. Dinophysis Abundance and Toxin Concentration
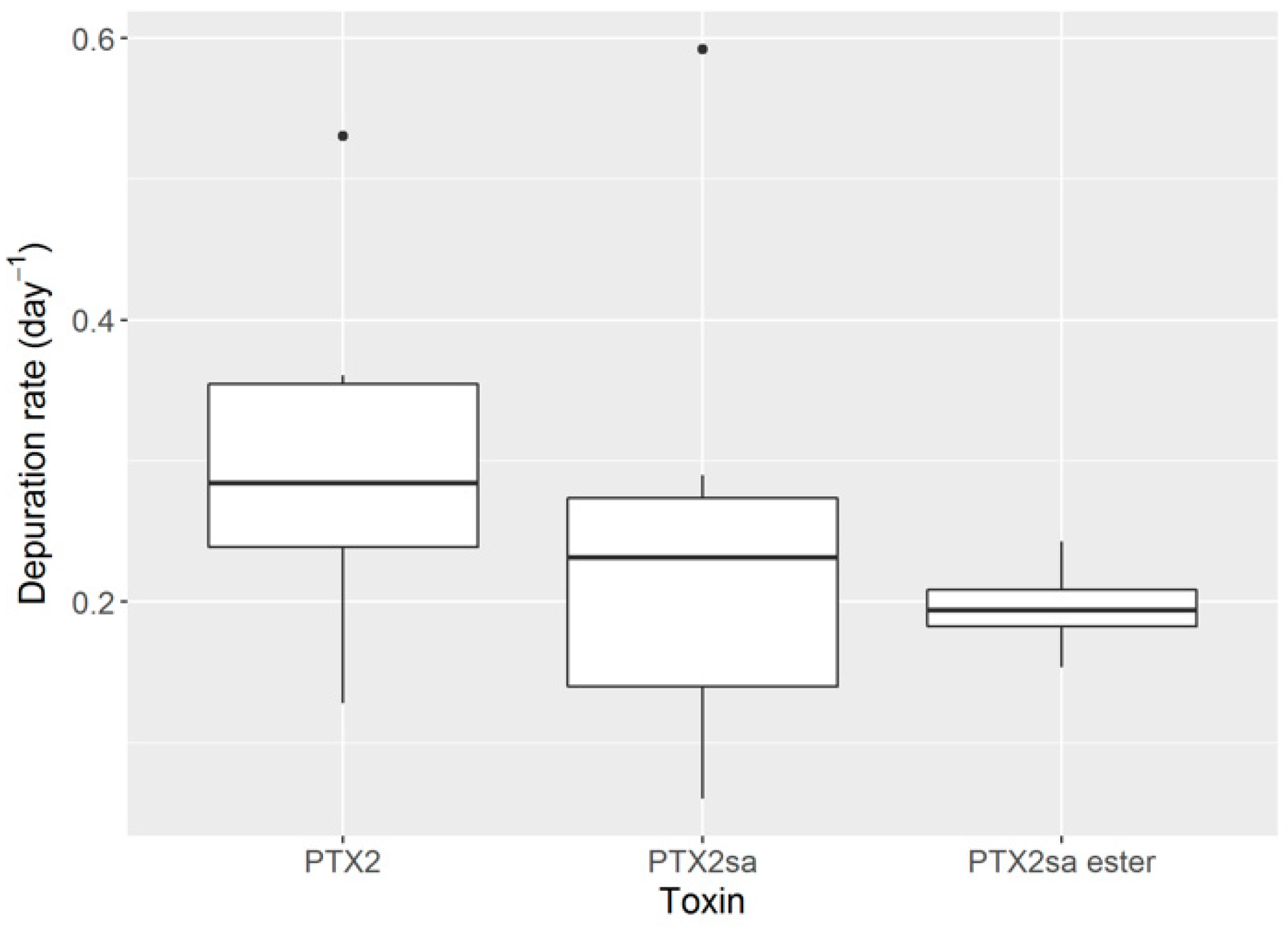
2.5. Depuration Rates
3. Discussion
4. Materials and Methods
4.1. Area of Study, Phytoplankton Sampling, Quantification and Taxonomic Analyses
4.2. Shellfish Sampling, Toxin Extraction and Hydrolysis
4.3. Toxin Detection and Quantification
4.4. Estimation of Depuration Rates
4.5. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Guzmán, L.; Campodonico, I. Marea roja en la región de Magallanes. Pub. Inst. Patagon. Ser. Monogr. 1975, 9, 44. [Google Scholar]
- Lembeye, G.; Yasumoto, Y.; Zhao, J.; Fernández, R. DSP outbreak in Chilean fjords. In Toxic Phytoplankton Blooms in the Sea; Smayda, T.J., Shimizu, Y., Eds.; Elservier: Amsterdam, The Netherlands, 1993; pp. 525–529. [Google Scholar]
- Zhao, J.; Lembeye, G.; Cenci, G.; Wall, B.; Yasumoto, T. Determination of okadaic acid and dinophysistoxin-1 in mussels from Chile, Italy and Ireland. In Toxic Phytoplankton Blooms in the Sea; Smayda, T.J., Shimizu, Y., Eds.; Elsevier: Amsterdam, The Netherlands, 1993; pp. 587–592. [Google Scholar]
- IOC. Second IOC Regional Science Planning Workshop on Harmful Algal Blooms in South America; IOC: Mar del Plata, Argentina, 1995; p. 75. [Google Scholar]
- Uribe, J.C.; García, C.; Rivas, M.; Lagos, N. First report of diarrhetic shellfish toxins in Magellanic fjord, Southern Chile. J. Shellfish Res. 2001, 20, 69–74. [Google Scholar]
- Villarroel, O. Detección de toxina paralizante, diarreica y amnésica en mariscos de la XI región por Cromatografía de Alta Resolución (HPLC) y bioensayo de ratón. Ciencia y Tecnologia del Mar 2004, 27, 33–42. [Google Scholar]
- Reguera, B.; Riobó, P.; Rodríguez, F.; Díaz, P.; Pizarro, G.; Paz, B.; Franco, J.; Blanco, J. Dinophysis Toxins: Causative Organisms, Distribution and Fate in Shellfish. Mar. Drugs 2014, 12, 394–461. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blanco, J.; Correa, J.; Muñíz, S.; Mariño, C.; Martín, H.; Arévalo, F. Evaluación del impacto de los métodos y niveles utilizados para el control de toxinas en el mejillón. Revista Galega dos Recursos Mariños 2013, 3, 1–55. Available online: https://www.researchgate.net/publication/236842103_Evaluacion_del_impacto_de_los_metodos_y_niveles_utilizados_para_el_control_de_toxinas_en_el_mejillon (accessed on 24 June 2018).
- Gestal Otero, J.J. Epidemiology of marine toxins. In Seafood and Freshwater Toxins. Physiology, Pharmacology and Detection, 3rd ed.; Botana, L.M., Ed.; CRC Press, Taylor and Francis Group: Boca Ratón, FL, USA, 2014; pp. 123–195. [Google Scholar]
- Munday, R. Toxicology of seafood toxins: A critical review. In Seafood and Freshwater Toxins: Pharmacology, Physiology, and Detection; CRC Press, Taylor and Francis Group: Boca Ratón, FL, USA, 2014; pp. 197–290. [Google Scholar]
- EFSA Panel on Contaminants in the Food Chain. Marine biotoxins in shellfish—Pectenotoxin group: Marine biotoxins in shellfish—Pectenotoxin group. EFSA J. 2009, 7. [Google Scholar] [CrossRef]
- EFSA Panel on Contaminants in the Food Chain. Scientific Opinion of the Panel on Contaminants in the Food Chain on a request from the European Commission on Marine Biotoxins in Shellfish—Summary on regulated marine biotoxins. EFSA J. 2009, 1306, 1–23. [Google Scholar]
- Suzuki, T.; Ota, H.; Yamasaki, M. Direct evidence of transformation of dinophysistoxin-1 to 7-O-acyl-dinophysistoxin-1 (dinophysistoxin-3) in the scallop Patinopecten yessoensis. Toxicon 1999, 37, 187–198. [Google Scholar] [CrossRef]
- Doucet, E.; Ross, N.N.; Quilliam, M.A. Enzymatic hydrolysis of esterified diarrhetic shellfish poisoning toxins and pectenotoxins. Anal. Bioanal. Chem. 2007, 389, 335–342. [Google Scholar] [CrossRef] [PubMed]
- Torgersen, T.; Sandvik, M.; Lundve, B.; Lindegarth, S. Profiles and levels of fatty acid esters of okadaic acid group toxins and pectenotoxins during toxin depuration. Part II: Blue mussels (Mytilus edulis) and flat oyster (Ostrea edulis). Toxicon 2008, 52, 418–427. [Google Scholar] [CrossRef] [PubMed]
- MacKenzie, L.A.; Selwood, A.I.; Marshall, C. Isolation and characterization of an enzyme from the Greenshell (TM) mussel Perna canaliculus that hydrolyses pectenotoxins and esters of okadaic acid. Toxicon 2012, 60, 406–419. [Google Scholar] [CrossRef] [PubMed]
- Wilkins, A.L.; Rehmann, N.; Torgersen, T.; Rundberget, T.; Keogh, M.; Petersen, D.; Hess, P.; Rise, F.; Miles, C.O. Identification of fatty acid esters of pectenotoxin-2 seco acid in blue mussels (Mytilus edulis) from Ireland. J. Agric. Food Chem. 2006, 54, 5672–5678. [Google Scholar] [CrossRef] [PubMed]
- Blanco, J.; Alvarez, G.; Uribe, E. Identification of pectenotoxins in plankton, filter feeders, and isolated cells of a Dinophysis acuminata with an atypical toxin profile, from Chile. Toxicon 2007, 49, 710–716. [Google Scholar] [CrossRef] [PubMed]
- Avaria, S.; Muñoz, P. Composición y biomasa del fitoplancton marino del norte de Chile en mayo de 1981 (operación oceanográfica MarChile XII-ERFEN III). Ciencia y Tecnología del Mar CONA 1983, 7, 109–140. [Google Scholar]
- Avaria, S.; Muñoz, P. Efectos del fenómeno “El Niño” sobre el fitoplancton marino del norte de Chile en diciembre de 1982. Ciencia y Tecnologia del Mar 1985, 9, 3–30. [Google Scholar]
- Avaria, S.; Muñoz, P.; Uribe, E. Composición y biomasa del fitoplancton marino del norte de Chile en Diciembre de 1980 (Operación oceanográfica MARCHILE XI-ERFEN II). Ciencia y Tecología del Mar CONA 1982, 6, 5–36. [Google Scholar]
- Santander, E.; Herrera, L.; Merino, C. Fluctuación diaria del fitoplancton en la capa superficial del océano durante la primavera de 1997 en el norte de Chile (20 18 S): II. Composición específica y abundancia celular. Revista de Biología Marina y Oceanografía 2003, 38, 13–25. [Google Scholar] [CrossRef]
- Faust, M.A.; Gulledge, R.A. Identifying harmful marine dinoflagellates. Contrib. USA Natl. Herb. 2002, 42, 1–144. [Google Scholar]
- Lebour, M.V. The Dinoflagellates of Northern Seas; Marine Biological Association of the United Kingdom: Plymouth, UK, 1925; Volume 250. [Google Scholar]
- Dodge, J.D.; Hart-Jones, B. Marine Dinoflagellates of the British Isles; Her Majesty’s Stationery Office (HMSO): London, UK, 1982. [Google Scholar]
- Olenina, I.; Hajdu, S.; Edler, L.; Andersson, A.; Wasmund, N.; Busch, S.; Göbel, J.; Gromisz, S.; Huseby, S.; Huttunen, M.; et al. Biovolumes and Size-Classes of Phytoplankton in the Baltic Sea; Baltic Marine Environment Protection Commission: Helsinki, Filand, 2006. [Google Scholar]
- Reguera, B. Biología, Autoecología y Toxinología de las Principales Especies del Género” Dinophysis” Asociadas a Episodios de Intoxicación Diarreogénica por Bivalvos (DSP). Ph.D. Thesis, Universidad de Barcelona, Barcelona, Spain, 2003. [Google Scholar]
- Sar, E.A.; Sunesen, I.; Lavigne, A.; Goya, A. Dinophysis spp. asociadas a detección de toxinas diarreicas (DSTs) en moluscos y a intoxicación diarreica en humanos (Provincia de Buenos Aires, Argentina). Revista de Biología Marina y Oceanografía 2010, 45, 451–460. [Google Scholar] [CrossRef]
- Fabro, E.; Almandoz, G.O.; Ferrario, M.; Tillmann, U.; Cembella, A.; Krock, B. Distribution of Dinophysis species and their association with lipophilic phycotoxins in plankton from the Argentine Sea. Harmful Algae 2016, 59, 31–41. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suzuki, T.; Mackenzie, L.; Stirling, D.; Adamson, J. Pectenotoxin-2 seco acid: A toxin converted from pectenotoxin-2 by the New Zealand Greenshell mussel, Perna canaliculus. Toxicon 2001, 39, 507–514. [Google Scholar] [CrossRef]
- Suzuki, T.; Mackenzie, L.; Stirling, D.; Adamson, J. Conversion of pectenotoxin-2 to pectenotoxin-2 seco acid in the New Zealand scallop, Pecten novaezelandiae. Fish. Sci. 2001, 67, 506–510. [Google Scholar] [CrossRef]
- Miles, C.O.; Wilkins, A.L.; Munday, R.; Dines, M.H.; Hawkes, A.D.; Briggs, L.R.; Sandvik, M.; Jensen, D.J.; Cooney, J.M.; Holland, P.T.; et al. Isolation of Pectenotoxin-2 From Dinophysis acuta and Its Conversion to Pectenotoxin-2 Seco Acid, and Preliminary Assessment of Their Acute Toxicities. Toxicon 2004, 43, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Burgess, V.; Shaw, G. Investigations into the Toxicology of Pectenotoxin-2-Seco Acid and 7-Epi Pectenotoxin 2-Seco Acid to Aid in a Health Risk Assessment for the Consumption of Shellfish Contaminated with These Shellfish Toxins in Australia; Report on Project No. 2001/258; Fisheries Research and Development Corporation: Canberra, Australia, 2003; ISBN 0975025910.
- Suzuki, T.; Mitsuya, T.; Matsubara, H.; Yamasaki, M. Determination of pectenotoxin-2 after solid-phase extraction from seawater and from the dinoflagellate Dinophysis fortii by liquid chromatography with electrospray mass spectrometry and ultraviolet detection. Evidence of oxidation of pectenotoxin-2 to pectenotoxin-6 in scallops. J. Chromatogr. A 1998, 815, 155–160. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, T. Chemistry and Detection of Okadaic Acid/Dinophysistoxins, Pectenotoxins and Yessotoxins. In Toxins and Biologically Active Compound from Microalgae. Vol 1 Origin, Chemistry and Detection; Rossini, G.P., Ed.; CRC Press: Boca Raton, FL, USA, 2014; pp. 99–152. [Google Scholar]
- Rossignoli, A.E.; Fernandez, D.; Regueiro, J.; Marino, C.; Blanco, J. Esterification of okadaic acid in the mussel Mytilus galloprovincialis. Toxicon 2011, 57, 712–720. [Google Scholar] [CrossRef] [PubMed]
- Marr, J.C.; Hu, T.; Pleasance, S.; Quilliam, M.A.; Wright, J.L.C. Detection of new 7-O-acyl derivatives of diarrhetic shellfish poisoning toxins by liquid chromatography-mass spectrometry. Toxicon 1992, 30, 1621–1630. [Google Scholar] [CrossRef]
- Morohashi, A.; Satake, M.; Murata, K.; Naoki, H.; Kaspar, H.F.; Yasumoto, T. Brevetoxin B3, a new brevetoxin analog isolated from the greenshell mussel Perna canaliculus involved in neurotoxic shellfish poisoning in new zealand. Tetrahedron Lett. 1995, 36, 8995–8998. [Google Scholar] [CrossRef]
- Aasen, J.A.; Hardstaff, W.; Aune, T.; Quilliam, M.A. Discovery of fatty acid ester metabolites of spirolide toxins in mussels from Norway using liquid chromatography/tandem mass spectrometry. Rapid Commun. Mass Spectrom 2006, 20, 1531–1537. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De la Iglesia, P.; McCarron, P.; Diogene, J.; Quilliam, M.A. Discovery of gymnodimine fatty acid ester metabolites in shellfish using liquid chromatography/mass spectrometry. Rapid Commun. Mass Spectrom. 2013, 27, 643–653. [Google Scholar] [CrossRef] [PubMed]
- Janer, G.; Lavado, R.; Thibaut, R.; Porte, C. Effects of 17β-estradiol exposure in the mussel Mytilus galloprovincialis: A possible regulating role for steroid acyltransferases. Aquat. Toxicol. 2005, 75, 32–42. [Google Scholar] [CrossRef] [PubMed]
- Janer, G.; Mesia-Vela, S.; Porte, C.; Kauffman, F.C. Esterification of vertebrate-type steroids in the Eastern oyster (Crassostrea virginica). Steroids 2004, 69, 129–136. [Google Scholar] [CrossRef] [PubMed]
- Furumochi, S.; Onoda, T.; Cho, Y.; Fuwa, H.; Sasaki, M.; Yotsu-Yamashita, M.; Konoki, K. Effect of carbon chain length in acyl coenzyme A on the efficiency of enzymatic transformation of okadaic acid to 7-O-acyl okadaic acid. Bioorg. Med. Chem. Lett. 2016, 26, 2992–2996. [Google Scholar] [CrossRef] [PubMed]
- Blanco, J.; Fernández, M.L.; Míguez, A.; Moroño, A. Okadaic acid depuration in the mussel Mytilus galloprovincialis: One- and two-compartment models and the effect of environmental conditions. Mar. Ecol. Prog. Ser. 1999, 176, 153–163. [Google Scholar] [CrossRef]
- Moroño, A.; Arévalo, F.; Fernández, M.L.; Maneiro, J.; Pazos, Y.; Salgado, C.; Blanco, J. Accumulation and transformation of DSP toxins in mussels Mytilus galloprovincialis LMK during a toxic episode caused by Dinophysis acuminata. Aquat. Toxicol. 2003, 62, 269–280. [Google Scholar] [CrossRef]
- Lindegarth, S.; Torgersen, T.; Lundve, B.; Sandvik, M. Differential Retention of okadaic acid (OA) group toxins and pectenotoxins (PTX) in the blue mussel, Mytilus edulis (L.), and european glat oyster, Ostrea edulis (L.). J. Shellfish Res. 2009, 28, 313–323. [Google Scholar] [CrossRef]
- Vale, P. Differential Dynamics of Dinophysistoxins and Pectenotoxins Between Blue Mussel and Common Cockle: A Phenomenon Originating From the Complex Toxin Profile of Dinophysis acuta. Toxicon 2004, 44, 123–134. [Google Scholar] [CrossRef] [PubMed]
- Valle-Levinson, A.; Moraga, J.; Olivares, J.; Blanco, J.L. Tidal and residual circulation in a semi-arid bay: Coquimbo Bay, Chile. Cont. Shelf Res. 2000, 20, 2009–2028. [Google Scholar] [CrossRef]
- Berríos, M.; Olivares, J. Caracterización granulométrica y contenido de carbono orgánico de los sedimentos marinos superficiales, en el sistema de bahías de la IV Región. Coquimbo. Cienc. Tecnol. Mar. 1996, 19, 37–45. [Google Scholar]
- Fritz, L.; Triemer, R.E. A rapid simple technique utilizing calcofluor white M2R for the visualization of dinoflagellate thecal plates. J. Phycol. 1985, 21, 662–664. [Google Scholar] [CrossRef]
- Hasle, R.G. The inverted microscope method. In Phytoplankton Manual; UNESCO: Paris, France, 1978; pp. 88–96. [Google Scholar]
- EURLMB. EU-Harmonised Standard Operating Procedure for Determination of Lipophilic Marine Biotoxins in Molluscs by LC-MS/MS. Version 5. Available online: http://aesan.msssi.gob.es/CRLMB/docs/docs/metodos_analiticos_de_desarrollo/EU-Harmonised-SOP-LIPO-LCMSMS_Version5.pdf (accessed on 30 June 2015).
- Regueiro, J.; Rossignoli, A.E.; Alvarez, G.; Blanco, J. Automated on-line solid-phase extraction coupled to liquid chromatography tandem mass spectrometry for determination of lipophilic marine toxins in shellfish. Food Chem. 2011, 129, 533–540. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2014. [Google Scholar]
- Wickham, H. Ggplot2: Elegant Graphics for Data Analysis; Springer: New York, NY, USA, 2016. [Google Scholar]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Reference | Parent | Product | CE |
|---|---|---|---|
| PTXs method | |||
| OA_DTX-2 | 803.5 | 255.2 | 48 |
| OA_DTX-2 | 803.5 | 563.4 | 43 |
| DTX-1 | 817.5 | 255.2 | 48 |
| DTX-1 | 817.5 | 563.5 | 43 |
| PTX-2 | 876.5 | 805.5 | 23 |
| PTX-2 | 876.5 | 823.5 | 21 |
| PTX1 | 892.5 | 839.5 | 23 |
| PTX6 | 906.5 | 853.5 | 23 |
| PTX12 | 874.5 | 821.5 | 23 |
| PTX2sa | 894.5 | 823.5 | 21 |
| PTX2sa | 894.5 | 805.5 | 21 |
| PTX11sa | 910.5 | 179.2 | 50 |
| PTX11sa | 910.5 | 137.2 | 50 |
| C16-PTX2sa C33,37 | 1132.6 | 823.5 | 23 |
| C16-PTX2sa C11 | 1132.6 | 1061.5 | 23 |
| Acyl derivatives method | |||
| C14:0-PTX2sa | 1104.6 | 823.5 | 23 |
| C15:0-PTX2sa | 1118.6 | 823.5 | 23 |
| C16:1-PTX2sa | 1130.6 | 823.5 | 23 |
| C16:0-PTX2sa | 1132.6 | 823.5 | 23 |
| C17:1-PTX2sa | 1144.6 | 823.5 | 23 |
| C17:0-PTX2sa | 1146.6 | 823.5 | 23 |
| C18:5-PTX2sa | 1150.6 | 823.5 | 23 |
| C18:4-PTX2sa | 1152.6 | 823.5 | 23 |
| C18:3-PTX2sa | 1154.6 | 823.5 | 23 |
| C18:2-PTX2sa | 1156.6 | 823.5 | 23 |
| C18:1-PTX2sa | 1158.6 | 823.5 | 23 |
| C18:0-PTX2sa | 1160.6 | 823.5 | 23 |
| C20:5-PTX2sa | 1178.6 | 823.5 | 23 |
| C20:4-PTX2sa | 1180.6 | 823.5 | 23 |
| C20:2-PTX2sa | 1184.6 | 823.5 | 23 |
| C20:1-PTX2sa | 1186.6 | 823.5 | 23 |
| C20:0-PTX2sa | 1188.6 | 823.5 | 23 |
| C22:6-PTX2sa | 1204.6 | 823.5 | 23 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Blanco, J.; Álvarez, G.; Rengel, J.; Díaz, R.; Mariño, C.; Martín, H.; Uribe, E. Accumulation and Biotransformation of Dinophysis Toxins by the Surf Clam Mesodesma donacium. Toxins 2018, 10, 314. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins10080314
Blanco J, Álvarez G, Rengel J, Díaz R, Mariño C, Martín H, Uribe E. Accumulation and Biotransformation of Dinophysis Toxins by the Surf Clam Mesodesma donacium. Toxins. 2018; 10(8):314. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins10080314
Chicago/Turabian StyleBlanco, Juan, Gonzalo Álvarez, José Rengel, Rosario Díaz, Carmen Mariño, Helena Martín, and Eduardo Uribe. 2018. "Accumulation and Biotransformation of Dinophysis Toxins by the Surf Clam Mesodesma donacium" Toxins 10, no. 8: 314. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins10080314