Development of Real-Time Immuno-PCR Based on Phage Displayed an Anti-Idiotypic Nanobody for Quantitative Determination of Citrinin in Monascus

Abstract
:1. Introduction
2. Results
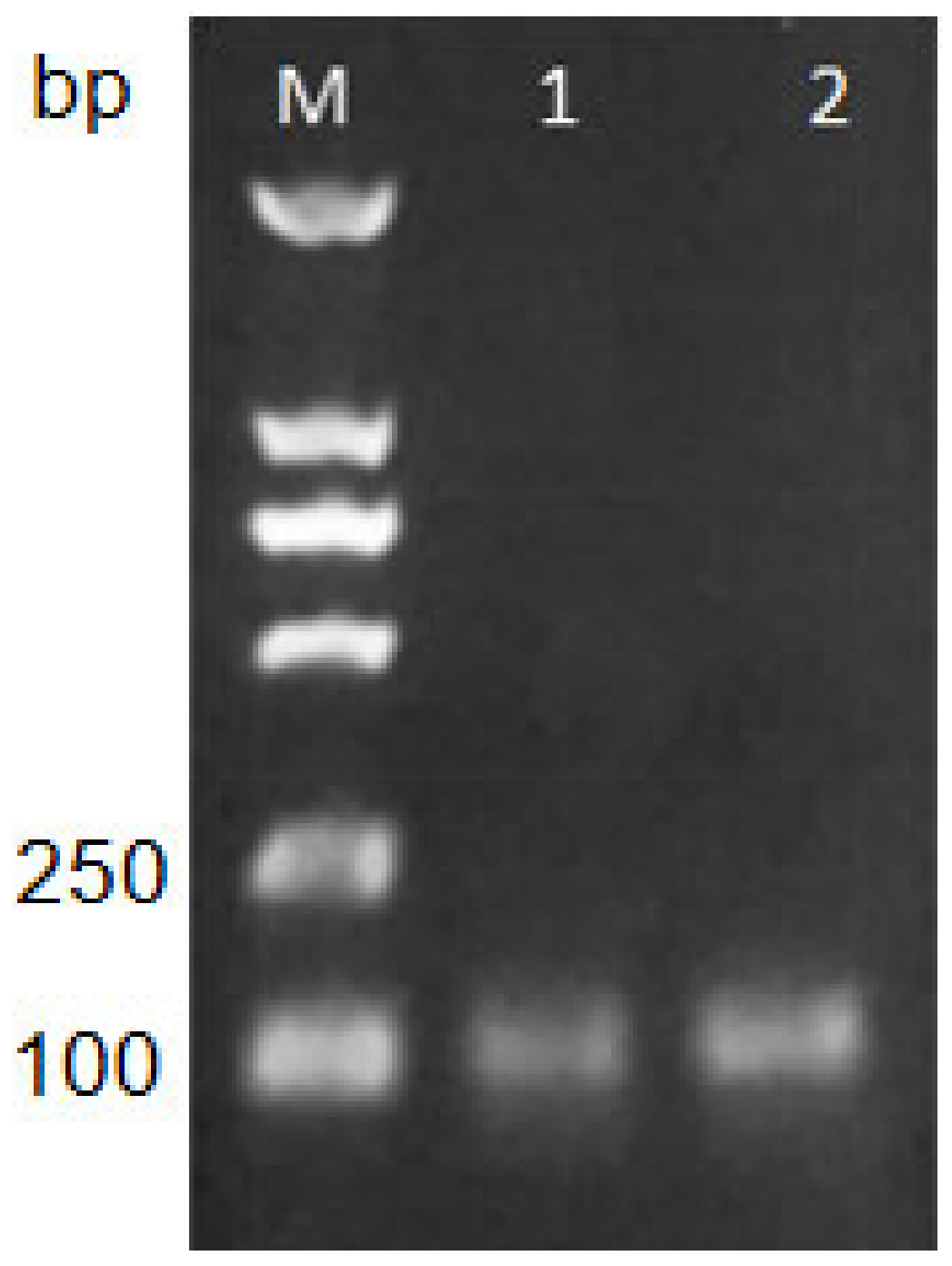
2.1. Verification of Correctness of Primers
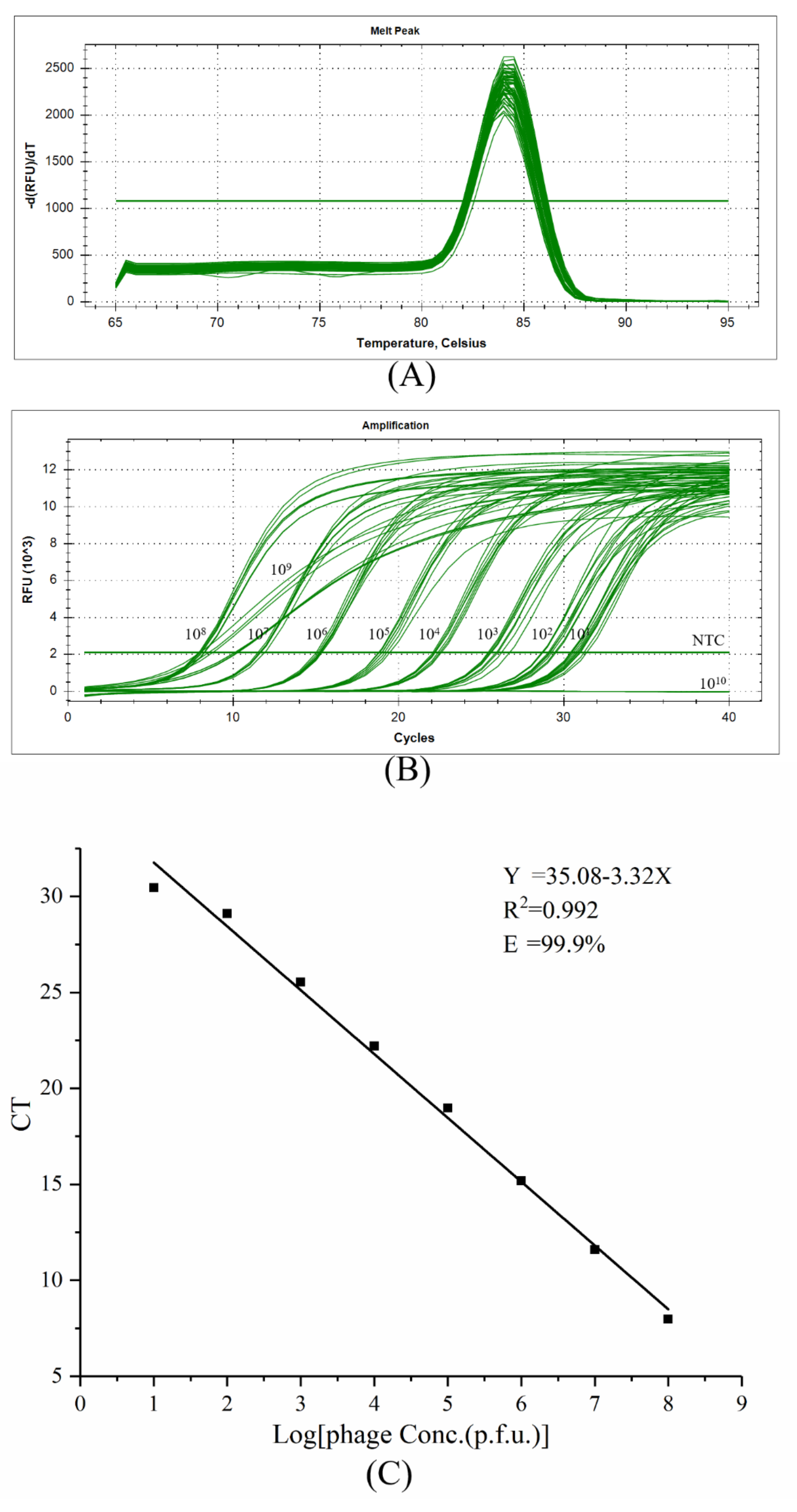
2.2. Optimization of Real-Time PCR Reaction Conditions
2.3. Establishment of rtIPCR for CIT
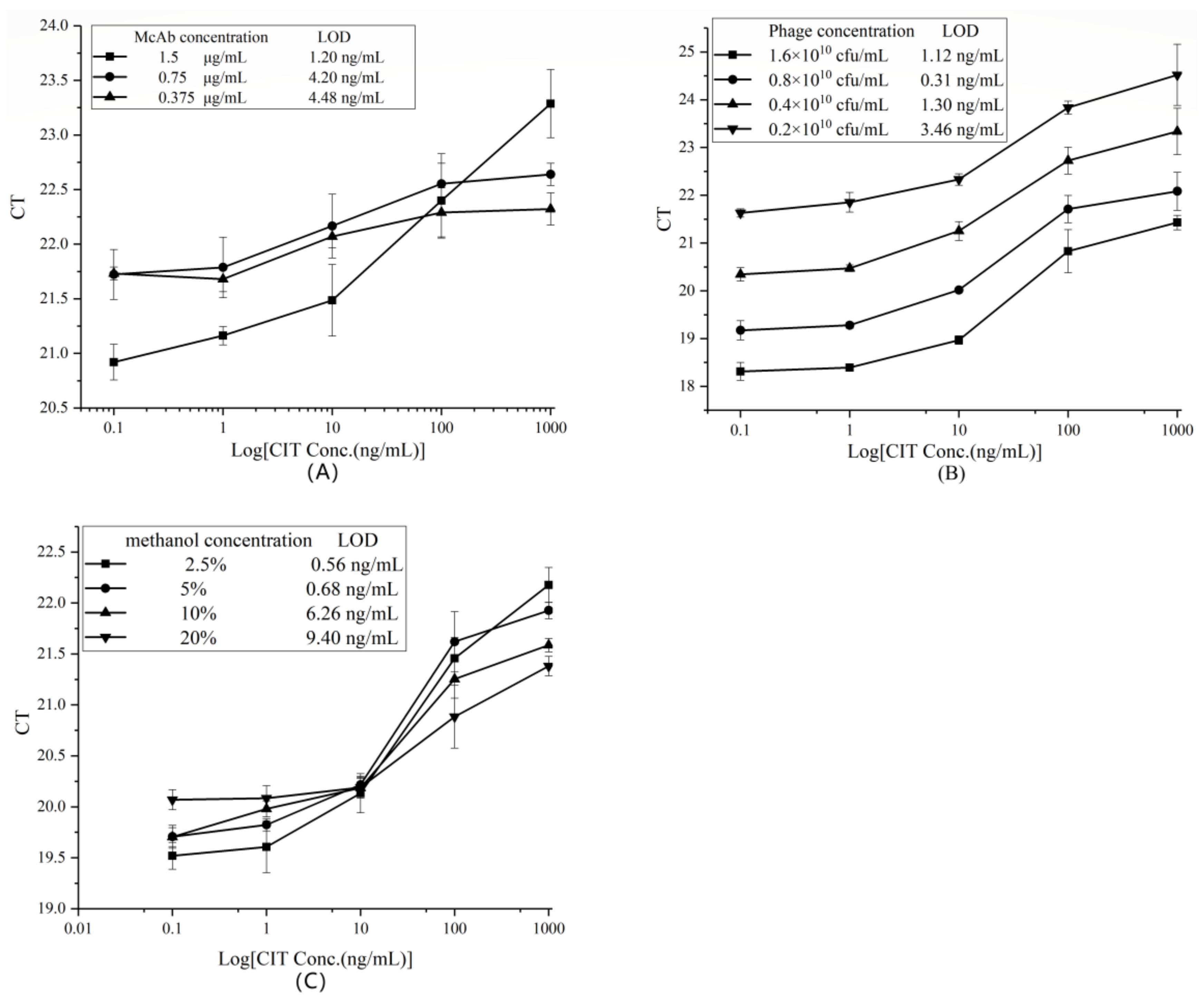
2.3.1. Optimization of the Concentration of Monoclonal Antibodies (McAbs)
2.3.2. Optimization of Phage Input
2.3.3. The Effect of Methanol Concentration
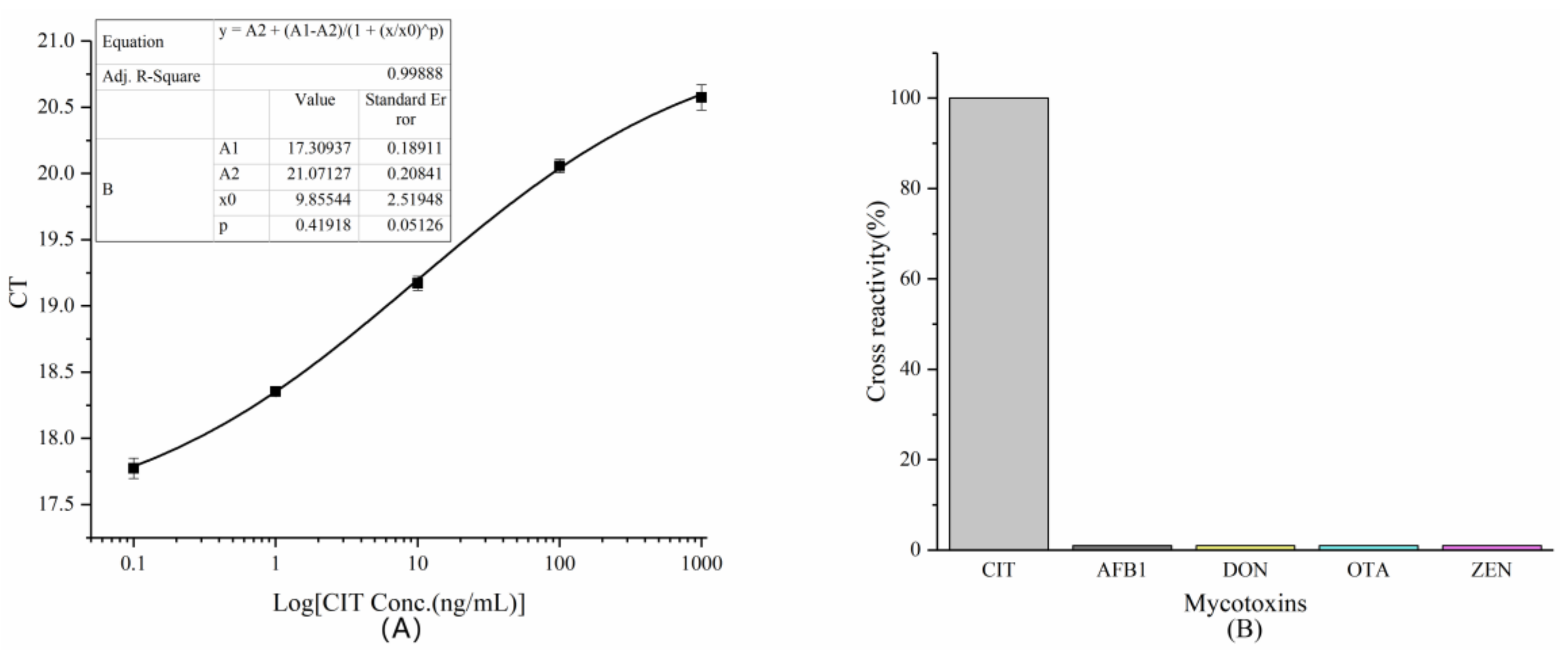
2.3.4. Establishment of A Standard Curve
2.3.5. Cross-Reactivity
2.4. Assay Validation
2.5. Sample Analysis
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Chemicals and Reagents
5.2. Amplification and Purification of Phage Displaying Anti-Idiotypic Nanobodies
5.3. Phage Titer Determination
5.4. PCR Validation of Primer
5.5. Efficiency Assessment of Real-Time PCR
5.6. Phage-Mediated rtIPCR
5.7. Spike and Recovery Analysis
5.8. Sample Analysis
Author Contributions
Funding
Conflicts of Interest
References
- Blanc, P.J.; Laussac, J.; Le Bars, J.; Le Bars, P.; Loret, M.; Pareilleux, A.; Prome, D.; Prome, J.; Santerre, A.; Goma, G. Characterization of monascidin A from Monascus as citrinin. Int. J. Food Microbiol. 1995, 27, 201–213. [Google Scholar] [CrossRef]
- Flajs, D.; Peraica, M. Toxicological properties of citrinin. Arch. Ind. Hyg. Toxicol. 2009, 60, 457–464. [Google Scholar] [CrossRef] [PubMed]
- Liu, B.-H.; Wu, T.-S.; Su, M.-C.; Chung, C.P.; Yu, F.-Y. Evaluation of citrinin occurrence and cytotoxicity in Monascus fermentation products. J. Agric. Food Chem. 2005, 53, 170–175. [Google Scholar] [CrossRef] [PubMed]
- Wong, H.C.; Koehler, P.E. Production and isolation of an antibiotic from Monascus purpureus and its relationship to pigment production. J. Food Sci. 1981, 46, 589–592. [Google Scholar] [CrossRef]
- Rašić, D.; Mladinić, M.; Želježić, D.; Pizent, A.; Stefanović, S.; Milićević, D.; Konjevoda, P.; Peraica, M. Effects of combined treatment with ochratoxin A and citrinin on oxidative damage in kidneys and liver of rats. Toxicon 2018, 146, 99–105. [Google Scholar] [CrossRef] [PubMed]
- Pleadin, J.; Zadravec, M.; Lešić, T.; Vahčić, N.; Frece, J.; Mitak, M.; Markov, K. Co-occurrence of ochratoxin A and citrinin in unprocessed cereals established during a three-year investigation period. Food Addit. Contam. Part B 2018, 11, 20–25. [Google Scholar] [CrossRef] [PubMed]
- Limay-Rios, V.; Miller, J.D.; Schaafsma, A.W. Occurrence of Penicillium verrucosum, ochratoxin A, ochratoxin B and citrinin in on-farm stored winter wheat from the Canadian Great Lakes Region. PLoS ONE 2017, 12, e0181239. [Google Scholar] [CrossRef]
- Wawrzyniak, J.; Waśkiewicz, A.; Ryniecki, A. Evaluation of critical points of mould growth and mycotoxin production in the stored barley ecosystem with a hazardous initial microbiological state of grain. J. Stored Prod. Res. 2018, 77, 166–176. [Google Scholar] [CrossRef]
- Čulig, B.; Bevardi, M.; Bošnir, J.; Serdar, S.; Lasić, D.; Racz, A.; Galić, A.; Kuharić, Ž. Presence of citrinin in grains and its possible health effects. Afr. J. Tradit. Complement. Altern. Med. 2017, 14, 22–30. [Google Scholar] [CrossRef]
- Ali, N. Co-occurrence of citrinin and ochratoxin A in rice in Asia and its implications for human health. J. Sci. Food Agric. 2018, 98, 2055–2059. [Google Scholar] [CrossRef] [PubMed]
- Arévalo, F.J.; Granero, A.M.; Fernández, H.; Raba, J.; Zón, M.A. Citrinin (CIT) determination in rice samples using a micro fluidic electrochemical immunosensor. Talanta 2011, 83, 966–973. [Google Scholar] [CrossRef] [PubMed]
- Pleadin, J.; Frece, J.; Kudumija, N.; Petrović, D.; Vasilj, V.; Zadravec, M.; Škrivanko, M.; Perković, I.; Markov, K. Citrinin in cereals and feedstuffs coming from Croatia and Bosnia & Herzegovina. Food Addit. Contam. Part B 2016, 9, 268–274. [Google Scholar]
- Ning, Z.-Q.; Cui, H.; Xu, Y.; Huang, Z.-B.; Tu, Z.; Li, Y.-P. Deleting the citrinin biosynthesis-related gene, ctnE, to greatly reduce citrinin production in Monascus aurantiacus Li AS3. 4384. Int. J. Food Microbiol. 2017, 241, 325–330. [Google Scholar] [CrossRef] [PubMed]
- Huang, Z.; Zhang, L.; Wang, Y.; Gao, H.; Li, X.; Huang, X.; Huang, T. Effects of rutin and its derivatives on citrinin production by Monascus aurantiacus Li AS3. 4384 in liquid fermentation using different types of media. Food Chem. 2019, 284, 205–212. [Google Scholar] [CrossRef] [PubMed]
- Tao, J.; Jing, Z.; Honglei, L.; Rong, J. Determination of citrinin in rice by thin layer chromatography. J. Hyg. Res. 2002, 6, 25. [Google Scholar]
- Gimeno, A. Determination of citrinin in corn and barley on thin layer chromatographic plates impregnated with glycolic acid. J. Assoc. Off. Anal. Chem. 1984, 67, 194–196. [Google Scholar]
- Chalam, R.; Stahr, H. Thin layer chromatographic determination of citrinin. J. Assoc. Off. Anal. Chem. 1979, 62, 570–572. [Google Scholar]
- Gimeno, A.; Martins, M.L. Rapid thin layer chromatographic determination of patulin, citrinin, and aflatoxin in apples and pears, and their juices and jams. J. Assoc. Off. Anal. Chem. 1983, 66, 85–91. [Google Scholar]
- Geng, X.; Wang, N.; Gao, Y.; Ning, H.; Guan, Y. A novel HPLC flow cell integrated UV light emitting diode induced fluorescence detector as alternative for sensitive determination of aflatoxins. Anal. Chim. Acta 2018, 1033, 81–86. [Google Scholar] [CrossRef]
- Tan, G.H.; Wong, R.C. Method validation in the determination of aflatoxins in noodle samples using the QuEChERS method (Quick, Easy, Cheap, Effective, Rugged and Safe) and high performance liquid chromatography coupled to a fluorescence detector (HPLC–FLD). Food Control 2011, 22, 1807–1813. [Google Scholar]
- Frisvad, J.C.; Thrane, U. Standardized high-performance liquid chromatography of 182 mycotoxins and other fungal metabolites based on alkylphenone retention indices and UV—VIS spectra (diodearray detection). J. Chromatogr. A 1987, 404, 195–214. [Google Scholar] [CrossRef]
- Ji, X.; Xu, J.; Wang, X.; Qi, P.; Wei, W.; Chen, X.; Li, R.; Zhou, Y. Citrinin determination in red fermented rice products by optimized extraction method coupled to liquid chromatography tandem mass spectrometry (LC-MS/MS). J. Food Sci. 2015, 80, T1438–T1444. [Google Scholar] [CrossRef] [PubMed]
- Sadi, B.S.; Bayat, M.; Tajik, P.; Hashemi, S.J. Citrinin detection by intensified fluorescence signal of a FRET-based immunosensor using magnetic/silica core–shell. Saudi J. Biol. Sci. 2018, 25, 171–177. [Google Scholar] [CrossRef] [PubMed]
- Atar, N.; Eren, T.; Yola, M.L. A molecular imprinted SPR biosensor for sensitive determination of citrinin in red yeast rice. Food Chem. 2015, 184, 7–11. [Google Scholar] [CrossRef] [PubMed]
- Zachetti, V.G.L.; Granero, A.M.; Robledo, S.N.; Zon, M.A.; Fernández, H. Development of an amperometric biosensor based on peroxidases to quantify citrinin in rice samples. Bioelectrochemistry 2013, 91, 37–43. [Google Scholar] [CrossRef]
- De Baere, S.; Osselaere, A.; Devreese, M.; Vanhaecke, L.; De Backer, P.; Croubels, S. Development of a liquid–chromatography tandem mass spectrometry and ultra-high-performance liquid chromatography high-resolution mass spectrometry method for the quantitative determination of zearalenone and its major metabolites in chicken and pig plasma. Anal. Chim. Acta 2012, 756, 37–48. [Google Scholar] [CrossRef] [PubMed]
- Sun, W.; Han, Z.; Aerts, J.; Nie, D.; Jin, M.; Shi, W.; Zhao, Z.; De Saeger, S.; Zhao, Y.; Wu, A. A reliable liquid chromatography–tandem mass spectrometry method for simultaneous determination of multiple mycotoxins in fresh fish and dried seafoods. J. Chromatogr. A 2015, 1387, 42–48. [Google Scholar] [CrossRef]
- Li, Y.; Wu, H.; Guo, L.; Zheng, Y.; Guo, Y. Microsphere-based flow cytometric immunoassay for the determination of citrinin in red yeast rice. Food Chem. 2012, 134, 2540–2545. [Google Scholar] [CrossRef]
- Cheng, H.; Chen, Y.; Yang, Y.; Chen, X.; Guo, X.; Du, A. Characterization of anti-citrinin specific scFvs selected from non-immunized mouse splenocytes by eukaryotic ribosome display. PLoS ONE 2015, 10, e0131482. [Google Scholar] [CrossRef]
- Xu, Y.; Xiong, L.; Li, Y.; Xiong, Y.; Tu, Z.; Fu, J.; Tang, X. Citrinin detection using phage-displayed anti-idiotypic single-domain antibody for antigen mimicry. Food Chem. 2015, 177, 97–101. [Google Scholar] [CrossRef]
- Xu, Y.; Xiong, L.; Li, Y.; Xiong, Y.; Tu, Z.; Fu, J.; Chen, B. Anti-idiotypic nanobody as citrinin mimotope from a naive alpaca heavy chain single domain antibody library. Anal. Bioanal. Chem. 2015, 407, 5333–5341. [Google Scholar] [CrossRef] [PubMed]
- Kalayu Yirga, S.; Ling, S.; Yang, Y.; Yuan, J.; Wang, S. The preparation and identification of a monoclonal antibody against citrinin and the development of detection via indirect competitive ELISA. Toxins 2017, 9, 110. [Google Scholar] [CrossRef] [PubMed]
- Wu, S.-W.; Yu, Y.-A.; Liu, B.-H.; Yu, F.-Y. Development of a Sensitive Enzyme-Linked Immunosorbent Assay and Rapid Gold Nanoparticle Immunochromatographic Strip for Detecting Citrinin in Monascus Fermented Food. Toxins 2018, 10, 354. [Google Scholar] [CrossRef] [PubMed]
- Shu, M.; Xu, Y.; Liu, X.; Li, Y.; He, Q.; Tu, Z.; Fu, J.; Gee, S.J.; Hammock, B.D. Anti-idiotypic nanobody-alkaline phosphatase fusion proteins: Development of a one-step competitive enzyme immunoassay for fumonisin B1 detection in cereal. Anal. Chim. Acta 2016, 924, 53–59. [Google Scholar] [CrossRef] [PubMed]
- Sun, Z.; Wang, X.; Chen, Q.; Yun, Y.; Tang, Z.; Liu, X. Nanobody-Alkaline Phosphatase Fusion Protein-Based Enzyme-Linked Immunosorbent Assay for One-Step Detection of Ochratoxin A in Rice. Sensors 2018, 18, 4044. [Google Scholar] [CrossRef]
- Guo, Y.-C.; Zhou, Y.-F.; Zhang, X.-E.; Zhang, Z.-P.; Qiao, Y.-M.; Bi, L.-J.; Wen, J.-K.; Liang, M.-F.; Zhang, J.-B. Phage display mediated immuno-PCR. Nucleic Acids Res. 2006, 34, e62. [Google Scholar] [CrossRef] [PubMed]
- Lei, J.; Li, P.; Zhang, Q.; Wang, Y.; Zhang, Z.; Ding, X.; Zang, W. Anti-idiotypic nanobody-phage based real-time immuno-PCR for detection of hepatocarcinogen aflatoxin in grains and feedstuffs. Anal. Chem. 2014, 86, 10841–10846. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | CIT Add Amount (μg/kg) | rtIPCR (n = 3) | ||
|---|---|---|---|---|
| Average Content ± Deviation (μg/kg) | Recovery Rate (%) | Coefficient of Variation (%) | ||
| Rice flour | 0.1 | 0.09 ± 0.01 | 90.0 | 11.2 |
| 1 | 1.05 ± 0.16 | 104.6 | 14.8 | |
| 10 | 10.4 ± 1.0 | 103.9 | 9.8 | |
| 100 | 97.3 ± 6.5 | 97.3 | 6.7 | |
| 1000 | 916.4 ± 116.6 | 91.6 | 12.7 | |
| Flour | 0.1 | 0.09 ± 0.01 | 85.6 | 14.3 |
| 1 | 1.10 ± 0.14 | 110.0 | 13.2 | |
| 10 | 9.5 ± 1.2 | 94.7 | 12.3 | |
| 100 | 102.9 ± 12.9 | 103.0 | 12.5 | |
| 1000 | 757.9 ± 93.1 | 75.8 | 12.3 | |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Huang, W.; Tu, Z.; Ning, Z.; He, Q.; Li, Y. Development of Real-Time Immuno-PCR Based on Phage Displayed an Anti-Idiotypic Nanobody for Quantitative Determination of Citrinin in Monascus. Toxins 2019, 11, 572. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins11100572
Huang W, Tu Z, Ning Z, He Q, Li Y. Development of Real-Time Immuno-PCR Based on Phage Displayed an Anti-Idiotypic Nanobody for Quantitative Determination of Citrinin in Monascus. Toxins. 2019; 11(10):572. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins11100572
Chicago/Turabian StyleHuang, Wenping, Zhui Tu, Zhenqiang Ning, Qinghua He, and Yanping Li. 2019. "Development of Real-Time Immuno-PCR Based on Phage Displayed an Anti-Idiotypic Nanobody for Quantitative Determination of Citrinin in Monascus" Toxins 11, no. 10: 572. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins11100572