Distribution Characteristics and Environmental Control Factors of Lipophilic Marine Algal Toxins in Changjiang Estuary and the Adjacent East China Sea

 ,
,
Abstract
:1. Introduction
2. Results and Discussion
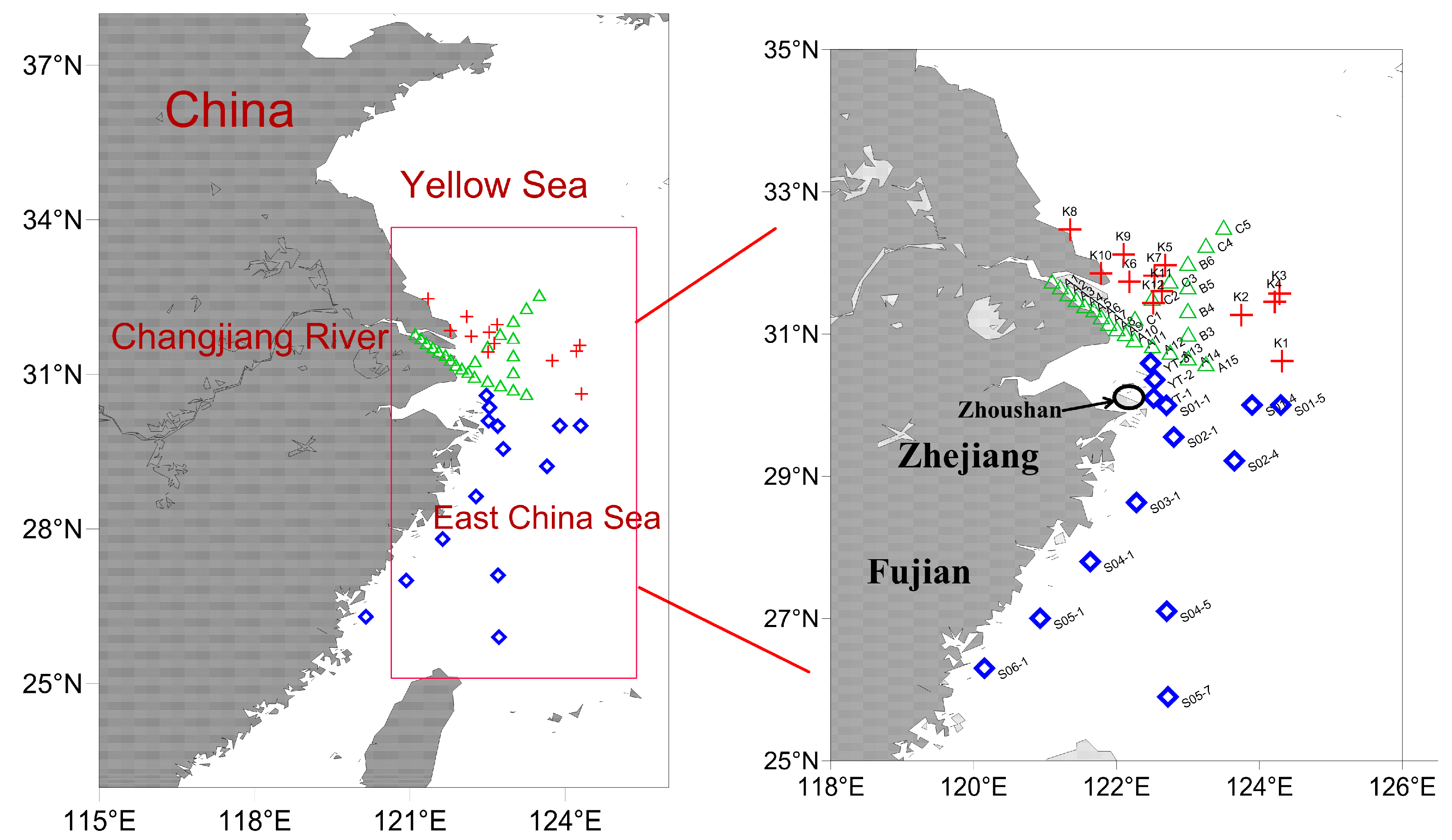
2.1. Study Area and Sample Collection
2.2. Identification of LMATs in Seawater and Phytoplankton
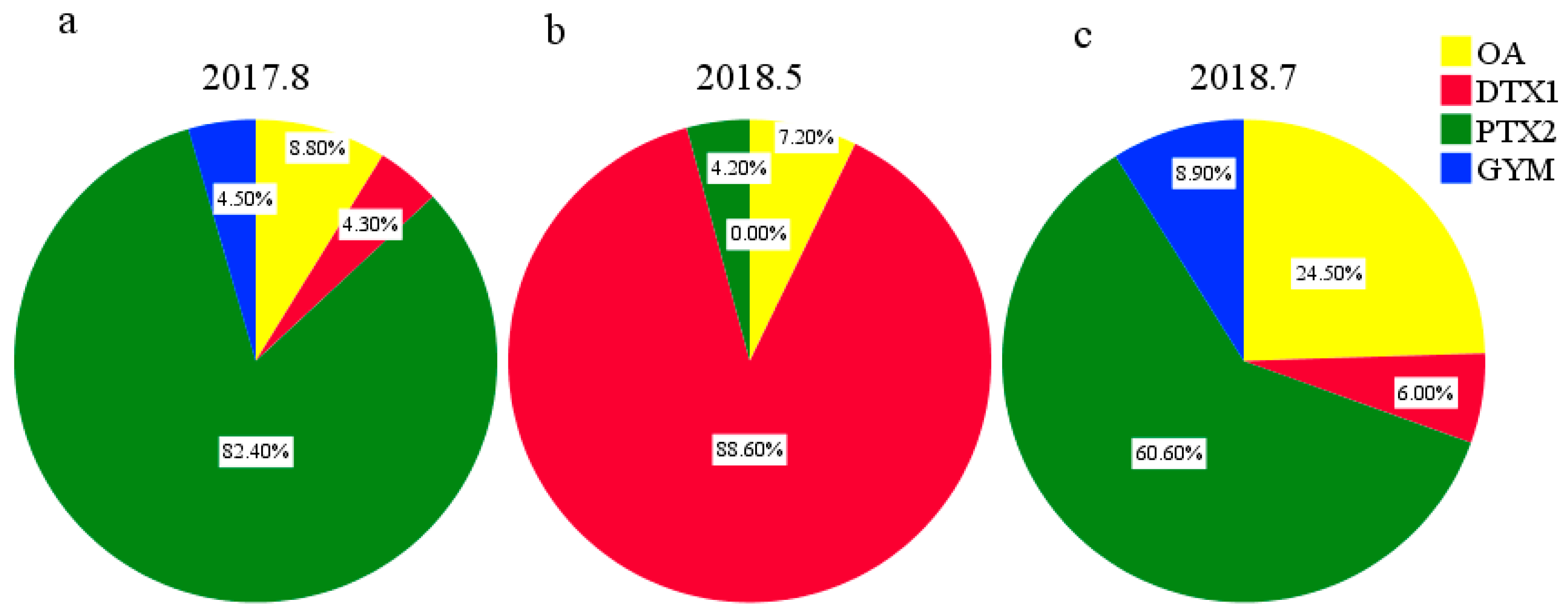
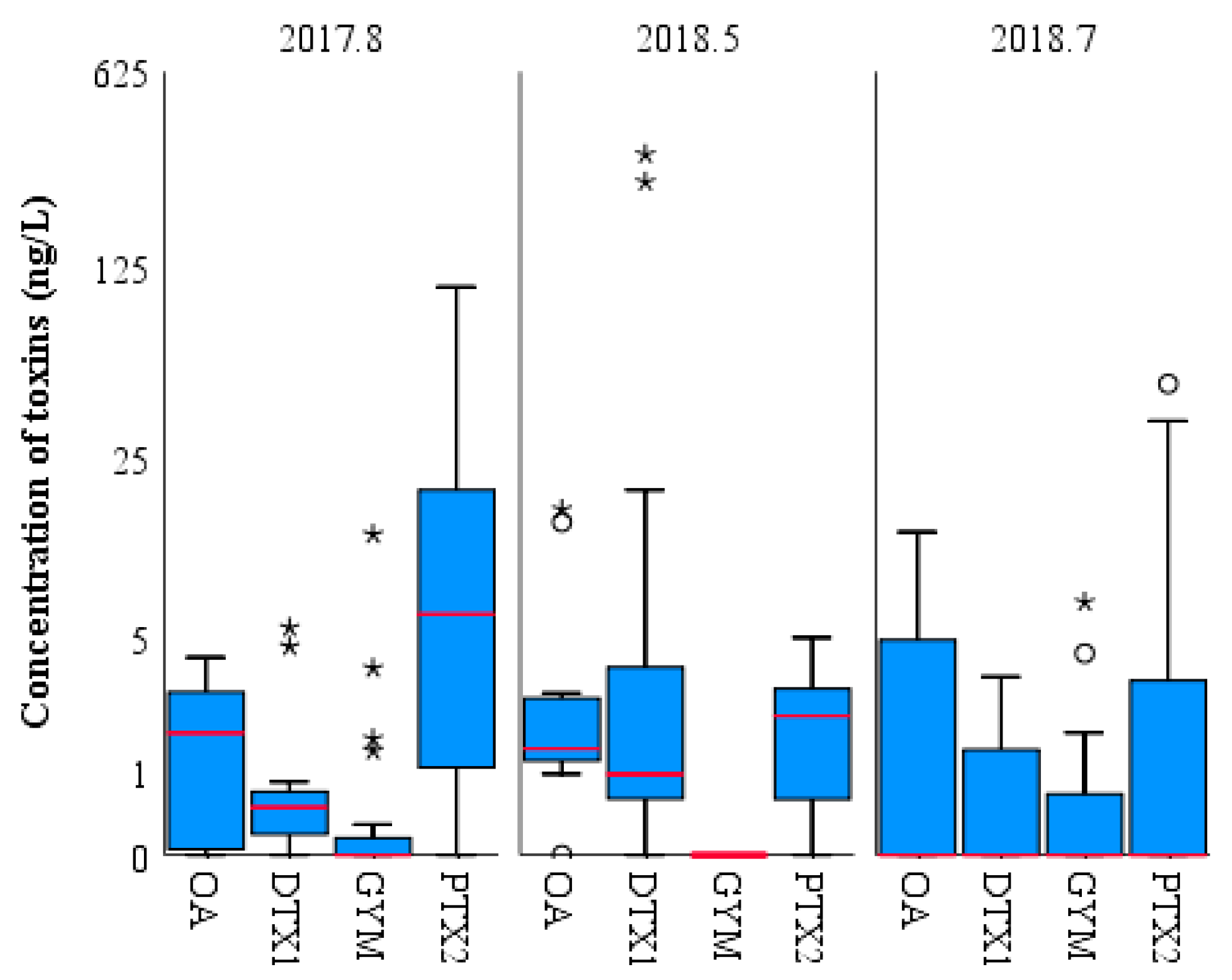
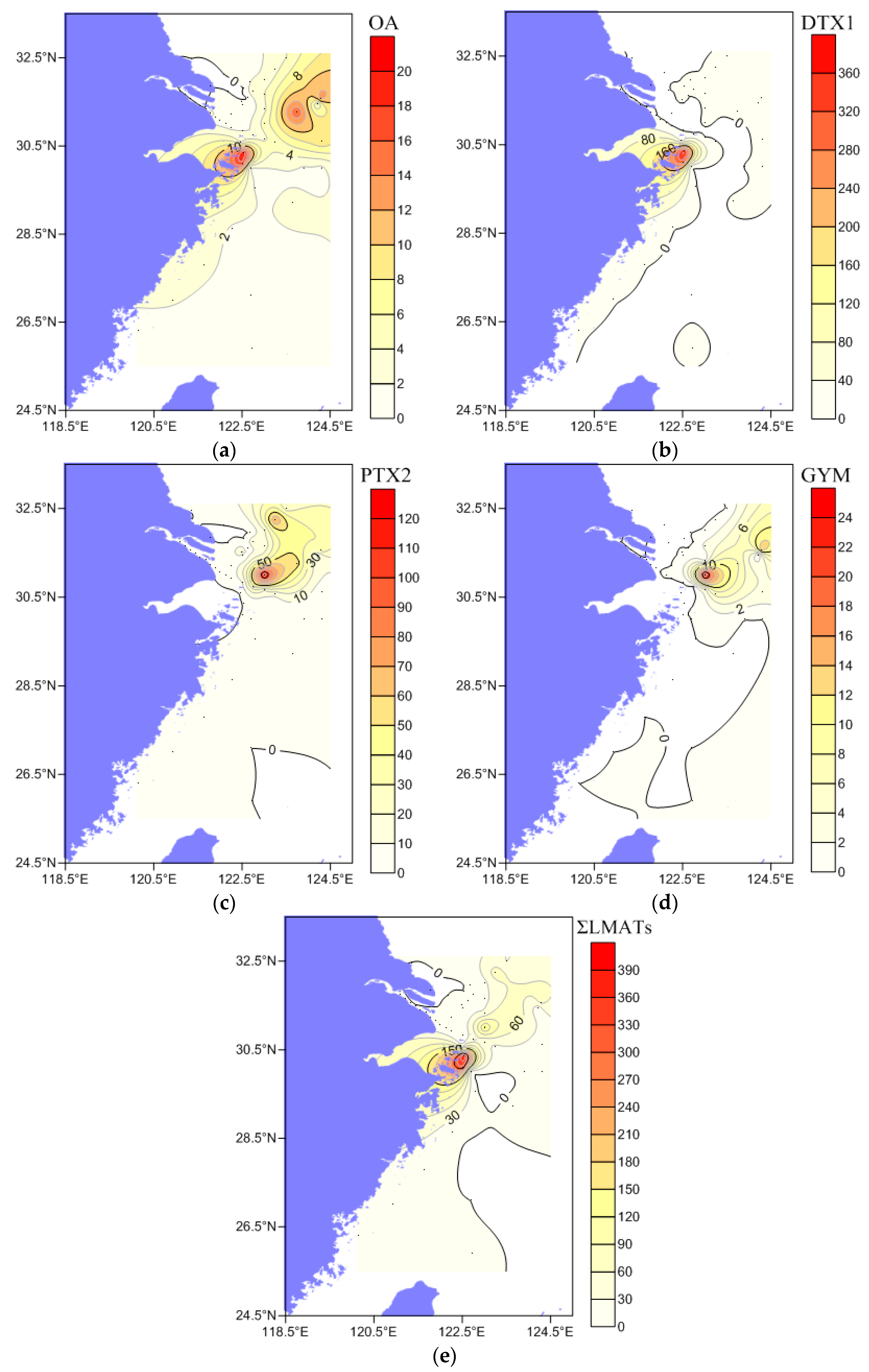
2.3. Composition, Concentration, and Distribution Characteristics of LMATs in Seawater
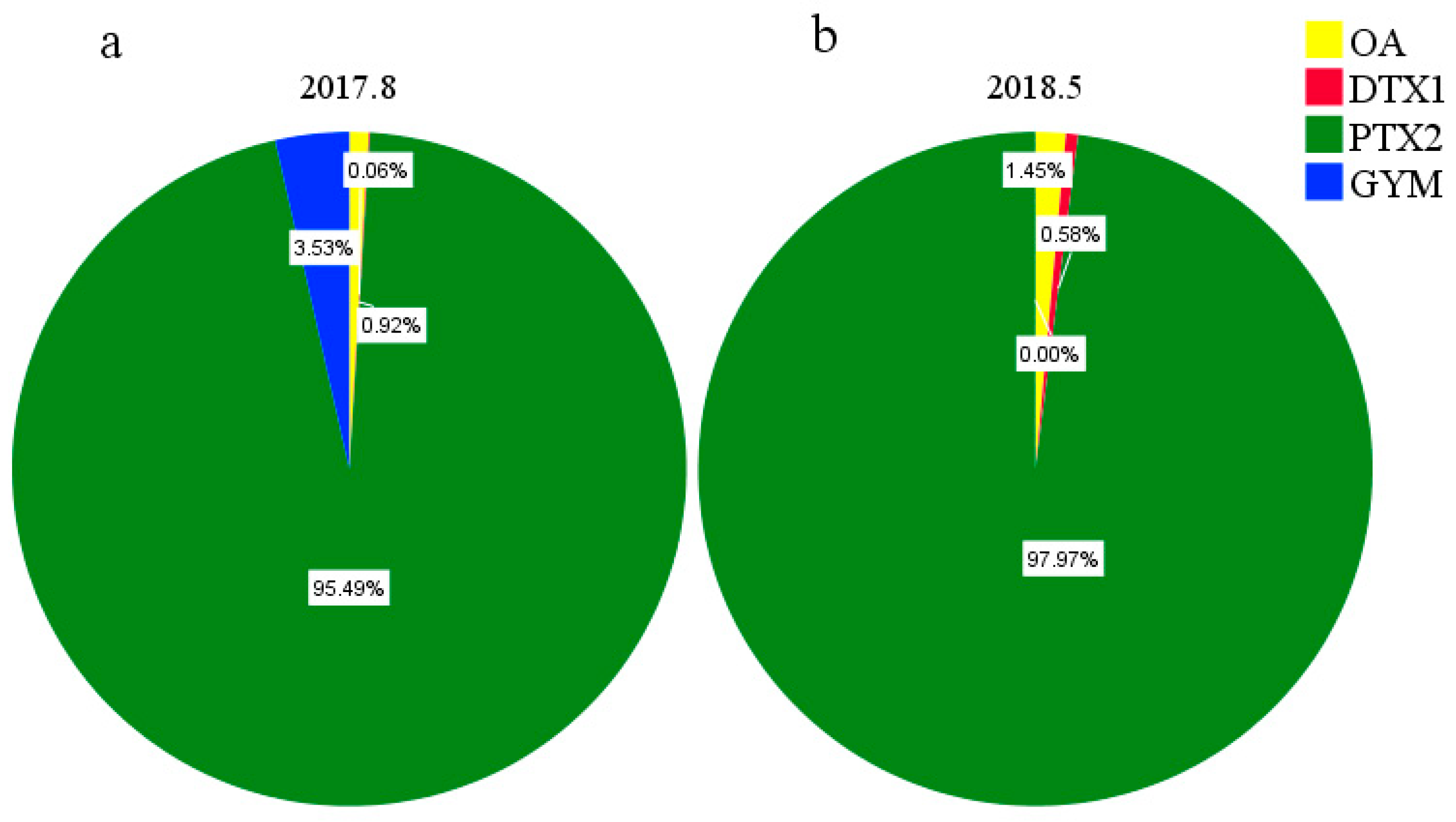
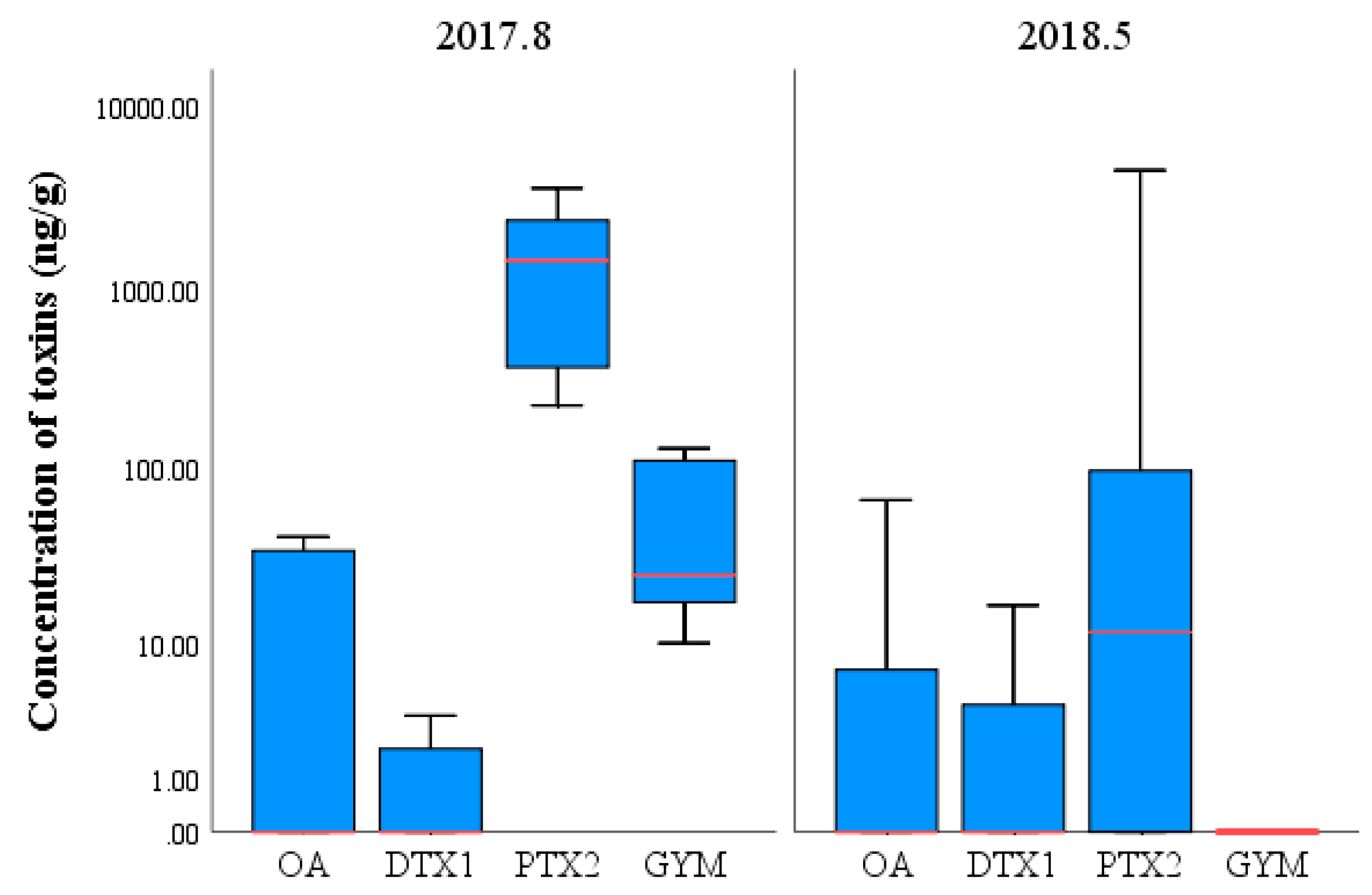
2.4. Composition and Concentration of LMATs in Phytoplankton
2.5. Phytoplankton Community Characteristics and LMAT Origins
2.6. Factors Affecting the Concentration and Distribution of LMATs in the CJE
3. Conclusions
4. Materials and Methods
4.1. Chemical Reagents
4.2. Preparation of LMAT Standard Solutions
4.3. Solid-Phase Extraction of Seawater Samples
4.4. Extraction of LMATs in Phytoplankton
4.5. HPLC-MS/MS Analysis Methods
4.6. Analytical Methods for Environmental Factors
4.7. Statistical Analysis
4.8. Quality Assurance and Quality Control (QA/QC)
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Behrenfeld, M.J.; O’Malley, R.T.; Siegel, D.A.; McClain, C.R.; Sarmiento, J.L.; Feldman, G.C.; Milligan, A.J.; Falkowski, P.G.; Letelier, R.M.; Boss, E.S. Climate-driven trends in contemporary ocean productivity. Nature 2006, 444, 752–755. [Google Scholar] [CrossRef]
- Hennon, G.M.; Dyhrman, S.T. Progress and promise of omics for predicting the impacts of climate change on harmful algal blooms. Harmful Algae 2019. [Google Scholar] [CrossRef]
- Du, W.; Lu, D.D. Harmful effects and detection of toxic algae and their algal toxins. J. Marin. Sci. 2008, 26, 89–97. [Google Scholar]
- Zhou, M.J.; Yu, R.C. Mechanisms and Impacts of Harmful Algal Blooms and the Count measures. Chin. J. Nat. 2007, 29, 72–77. [Google Scholar]
- Zingone, A.; Enevoldsen, H.O. The diversity of harmful algal blooms: A challenge for science and management. Ocean. Coast. Manag. 2000, 43, 725–748. [Google Scholar] [CrossRef]
- Li, Z.Y.; Chen, J.H.; Li, X.; Zhang, R.T.; Chen, C.; Shi, Q.; Wang, S.; Zheng, L.; Wang, X.R. Rapid screening and identification of paralytic shellfish poisoning toxins in red tide algae using hydrophilic interaction liquid chromatography-high resolution mass spectrometry with an accurate-mass database. Chin. J. Anal. Chem. 2013, 41, 979–985. [Google Scholar] [CrossRef]
- Toyofuku, H. Joint FAO/WHO/IOC activities to provide scientific advice on marine biotoxins. Mar. Pollut. Bull. 2006, 52, 1735–1745. [Google Scholar] [CrossRef]
- Chen, J.H.; Han, T.Z.; Li, X.T.; He, X.P.; Wang, Y.L.; Chen, F.R.; Song, X.C.; Zhou, D.S.; Wang, X.R. Occurrence and distribution of marine natural organic pollutants: Lipophilic marine algal toxins in the Yellow Sea and the Bohai Sea, China. Sci. Total. Environ. 2018, 612, 931–939. [Google Scholar] [CrossRef]
- Gerssen, A.; Mulder, P.P.J.; De Boer, J. Screening of lipophilic marine toxins in shellfish and algae: Development of a library using liquid chromatography coupled to orbitrap mass spectrometry. Anal. Chim. Acta 2011, 685, 176–185. [Google Scholar] [CrossRef]
- Chen, J.H.; Li, X.; Wang, S.; Chen, F.R.; Cao, W.; Sun, C.J.; Zheng, L.; Wang, X.R. Screening of lipophilic marine toxins in marine aquaculture environment using liquid chromatography-mass spectrometry. Chemosphere 2017, 168, 32–40. [Google Scholar] [CrossRef]
- Jauffrais, T.; Kilcoyne, J.; Herrenknecht, C.; Truquet, P.; Sechet, V.; Miles, C.O.; Hess, P. Dissolved azaspiracids are absorbed and metabolized by blue mussels (Mytilus edulis). Toxicon 2013, 65, 81–89. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, A.F.; Li, M.H.; Qiu, J.B.; Song, J.L.; Ji, Y.; Hu, Y.; Wang, S.Q.; Che, Y.J. Effect of Suspended Particulate Matter on the Accumulation of Dissolved Diarrhetic Shellfish Toxins by Mussels (Mytilus galloprovincialis) under Laboratory Conditions. Toxins 2018, 10, 273. [Google Scholar] [CrossRef]
- Mackenzie, L.A.; Selwood, A.I.; Mcnabb, P.; Rhodes, L. Benthic dinoflagellate toxins in two warm-temperate estuaries: Rangaunu and Parengarenga Harbours, Northland, New Zealand. Harmful Algae 2011, 10, 559–566. [Google Scholar] [CrossRef]
- Takahashi, E.; Yu, Q.M.; Eaglesham, G.; Connell, D.W.; McBroom, J.; Costanzo, S.; Shaw, G.R. Occurrence and seasonal variations of algal toxins in water, phytoplankton and shellfish from North Stradbroke Island, Queensland, Australia. Mar. Environ. Res. 2007, 64, 429–442. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roue, M.; Darius, H.T.; Chinain, M. Solid Phase Adsorption Toxin Tracking (SPATT) Technology for the Monitoring of Aquatic Toxins: A Review. Toxins 2018, 10, 167. [Google Scholar] [CrossRef] [PubMed]
- Rundberget, T.; Gustad, E.; Samdal, I.A.; Sandvik, M.; Miles, C.O. A convenient and cost-effective method for monitoring marine algal toxins with passive samplers. Toxicon 2009, 53, 542–550. [Google Scholar] [CrossRef]
- Fux, E.; Bire, R.; Hess, P. Comparative accumulation and composition of lipophilic marine biotoxins in passive samplers and in mussels (M. edulis) on the West Coast of Ireland. Harmful Algae 2009, 8, 523–537. [Google Scholar] [CrossRef]
- McCarthy, M.; van Pelt, F.N.A.M.; Bane, V.; O’Halloran, J.; Furey, A. Application of passive (SPATT) and active sampling methods in the profiling and monitoring of marine biotoxins. Toxicon 2014, 89, 77–86. [Google Scholar] [CrossRef]
- Boschorea, C.; Sanchis, J.; Farre, M.; Barcelo, D. Analysis of lipophilic marine biotoxins by liquid chromatography coupled with high-resolution mass spectrometry in seawater from the Catalan Coast. Anal. Bioanal. Chem. 2017, 409, 5451–5462. [Google Scholar] [CrossRef]
- Zendong, Z.; Bertrand, S.; Herrenknecht, C.; Abadie, E.; Jauzein, C.; Lemee, R.; Gouriou, J.; Amzil, Z.; Hess, P. Passive Sampling and High Resolution Mass Spectrometry for Chemical Profiling of French Coastal Areas with a Focus on Marine Biotoxins. Environ. Sci. Technol. 2016, 50, 8522–8529. [Google Scholar] [CrossRef]
- Li, X.; Li, Z.Y.; Chen, J.H.; Shi, Q.; Zhang, R.T.; Wang, S.; Wang, X.R. Detection, occurrence and monthly variations of typical lipophilic marine toxins associated with diarrhetic shellfish poisoning in the coastal seawater of Qingdao City, China. Chemosphere 2014, 111, 560–567. [Google Scholar] [CrossRef]
- Li, F.L.; Li, Z.X.; Guo, M.M.; Wu, H.Y.; Zhang, T.T.; Song, C.H. Investigation of diarrhetic shellfish toxins in Lingshan Bay, Yellow Sea, China, using solid-phase adsorption toxin tracking (SPATT). Food Addit. Contam Part A 2016, 33, 1367–1373. [Google Scholar] [CrossRef]
- Li, M.H.; Sun, G.; Qiu, J.B.; Li, A.F. Occurrence and variation of lipophilic shellfish toxins in phytoplankton, shellfish and seawater samples from the aquaculture zone in the Yellow Sea, China. Toxicon 2017, 127, 1–10. [Google Scholar] [CrossRef]
- Lane, J.Q.; Roddam, C.M.; Langlois, G.W.; Kudela, R.M. Application of solid phase adsorption toxin tracking (SPATT) for field detection of the hydrophilic phycotoxins domoic acid and saxitoxin in coastal California. Limnol. Oceanogr-Meth. 2010, 8, 645–660. [Google Scholar] [CrossRef]
- Fels-Klerx, H.J.V.D.; Olesen, J.E.; Naustvoll, L.J.; Friocourt, Y.; Mengelers, M.J.B.; Christensen, J.H. Climate change impacts on natural toxins in food production systems, exemplified by deoxynivalenol in wheat and diarrhetic shellfish toxins. Food Addit. Contam. A 2012, 29, 1647–1659. [Google Scholar] [CrossRef]
- Van Dolah, F.M. Marine algal toxins: Origins, health effects, and their increased occurrence. Environ. Health Perspect. 2000, 108 (Suppl. 1), 133–141. [Google Scholar] [CrossRef]
- Díaz, P.A.; Ruiz-Villarreal, M.; Pazos, Y.; Moita, T.; Reguera, B. Climate variability and Dinophysis acuta blooms in an upwelling system. Harmful Algae 2016, 53, 145–159. [Google Scholar] [CrossRef]
- Federica, F.L.; Blanco, L.; Rodríguez, L.P.; Vieites, J.M.; Cabado, A.G. Phycotoxins in Marine Shellfish: Origin, Occurrence and Effects on Humans. Mar. Drugs 2018, 16, 1–26. [Google Scholar]
- Gobler, C.J.; Doherty, O.M.; Hattenrath-Lehmann, T.K.; Griffith, A.W.; Kang, Y.; Litaker, R.W. Ocean warming since 1982 has expanded the niche of toxic algal blooms in the North Atlantic and North Pacific oceans. Proc. Natl. Acad. Sci. 2017, 114, 4975–4980. [Google Scholar] [CrossRef] [Green Version]
- Hattenrath-Lehmann, T.K.; Smith, J.L.; Wallace, R.B.; Merlo, L.R.; Koch, F.; Mittelsdorf, H.; Goleski, J.A.; Anderson, D.M.; Gobler, C.J. The effects of elevated CO2 on the growth and toxicity of field populations and cultures of the saxitoxin-producing dinoflagellate, Alexandrium fundyense. Limnol. Oceanogr. 2015, 60, 198–214. [Google Scholar] [CrossRef]
- Yu, R.C.; Liu, D.Y. Harmful Algal Blooms in the Coastal Waters of China: Current Situation, Long-term Changes and Prevention Strategies. Bull. Chin. Acad. Sci. 2016, 31, 1167–1174. [Google Scholar]
- Guo, H.; Ding, D.W.; Lin, F.G.; Guan, C.J. Characteristics and patterns of red tide in China coastal waters during the last 20a. Adv. Mar. Sci. 2015, 33, 547–558. [Google Scholar]
- Liu, R.Y.; Liu, L.; Liang, Y.B.; Yu, J.; Xu, D.Y.; Wei, N.; Yang, L.; Guo, H. The distribution, impacts and risks of toxic microalgae and phycotoxins in China. Mar. Environ. Sci. 2016, 35, 787–800. [Google Scholar]
- Yu, R.C.; Zhang, Q.C.; Kong, F.Z.; Zhou, Z.X.; Chen, Z.F.; Zhao, Y.; Geng, H.X.; Dai, L.; Yan, T.; Zhou, M.J. Status, impacts and long-term changes of harmful algae blooms in the sea area adjacent to the Changjiang river estuary. Oceanol. Limnol. Sin. 2017, 48, 1178–1186. [Google Scholar]
- Liu, M.Z.; Ma, J.R.; Kang, L.; Wei, Y.Y.; He, Q.; Hu, X.B.; Li, H. Strong turbulence benefits toxic and colonial cyanobacteria in water: A potential way of climate change impact on the expansion of Harmful Algal Blooms. Sci. Total. Environ. 2019, 670, 613–622. [Google Scholar] [CrossRef]
- Su, J.L.; Pan, Y.Q. On the shelf circulation north of Taiwan. Acta Oceanol. Sin. 1987, 6 (Suppl. I), 1–20. [Google Scholar]
- Tseng, Y.F.; Lin, J.; Dai, M.; Kao, S.J. Joint effect of freshwater plume and coastal upwelling on phytoplankton growth off the Changjiang River. Biogeosciences 2014, 11, 409–423. [Google Scholar] [CrossRef] [Green Version]
- Yang, D.Z.; Yin, B.S.; Su, J.C.; Zhang, Y. Numerical study on the origins and the forcing mechanism of the phosphate in upwelling areas off the coast of Zhejiang province, China in summer. J. Mar. Syst. 2013, 123, 1–18. [Google Scholar] [CrossRef]
- Zhu, R.; Yang, G.P.; Zhang, H.H. Temporal and spatial distributions of carbonyl sulfide, dimethyl sulfide, and carbon disulfide in seawater and marine atmosphere of the Changjiang Estuary and its adjacent East China Sea. Limnol. Oceanogr. 2018, 64, 1–18. [Google Scholar] [CrossRef]
- Zhou, M.J.; Shen, Z.L.; Yu, R.C. Responses of a coastal phytoplankton community to increased nutrient input from the Changjiang (Yangtze) River. Cont. Shelf Res. 2008, 28, 1483–1489. [Google Scholar] [CrossRef]
- Wang, Y.L.; Chen, J.H.; Li, Z.Y.; Wang, S.; Zheng, X.L.; Wang, X.R. Analysis of typical cyclic imine toxins by atmospheric pressure chemical ionization-mass spectrometry. J. Chin. Mass Spectr. Soc. 2015, 36, 551–558. [Google Scholar]
- European Commission. Commission Decision 2002/657/EC implementing Council Directive 96/23/EC Concerning the Performance of Analytical Methods and the Interpretation of Results. Off. J. Eur. Comm. 2002, 221, 8–36. [Google Scholar]
- Li, A.F.; Ma, J.G.; Cao, J.J.; McCarron, P. Toxins in mussels (Mytilus galloprovincialis) associated with diarrhetic shellfish poisoning episodes in China. Toxicon 2012, 60, 420–425. [Google Scholar] [CrossRef] [PubMed]
- Blanco, J.; Martín-Morales, E.; Álvarez, G. Stability of okadaic acid and 13-desmethyl spirolide C in seawater and sediment. Mar. Chem. 2008, 207, 21–25. [Google Scholar] [CrossRef]
- Mackenzie, L.A.; Holland, P.; Mcnabb, P.; Beuzenberg, V.; Selwood, A.I.; Suzuki, T. Complex toxin profiles in phytoplankton and Greenshell mussels (Perna canaliculus), revealed by LC-MS/MS analysis. Toxicon 2002, 40, 1321–1330. [Google Scholar] [CrossRef]
- Li, A.F.; Sun, G.; Qiu, J.B.; Li, F. Lipophilic shellfish toxins in Dinophysis caudata, picked cells and in shellfish from the East China Sea. Environ. Sci. Pollut. Res. 2015, 22, 3116–3126. [Google Scholar] [CrossRef]
- Yu, R.C.; Luo, X. Status and research perspectives on toxic algae and phycotoxins in the coastal waters of China. Stud. Mar. Sin. 2016, 51, 155–166. [Google Scholar]
- Stirling, D.J. Survey of historical New Zealand shellfish samples for accumulation of gymnodimine. New Zeal. J. Mar. Fresh. 2001, 35, 851–857. [Google Scholar] [CrossRef]
- Munday, R.; Towers, N.R.; MacKenzie, L.; Beuzenberg, V.; Holland, P.T.; Miles, C.O. Acute toxicity of gymnodimine in mice. Toxicon 2004, 44, 173–178. [Google Scholar] [CrossRef]
- Molgó, J.; Marchot, P.; Aráoz, R.; Benoit, E.; Iorga, B.I.; Zakarian, A.; Taylor, P.; Bourne, Y.; Servent, D. Cyclic imine toxins from dinoflagellates: A growing family of potent antagonists of the nicotinic acetylcholine receptors. J. Neurochem. 2017, 142, 41–51. [Google Scholar] [CrossRef]
- Zurhelle, C.; Nieva, J.; Tillmann, U.; Harder, T.; Krock, B.; Tebben, J. Identification of novel gymnodimines and spirolides from the marine dinoflagellate Alexandrium ostenfeldii. Mar. Drugs 2018, 16, 446. [Google Scholar] [CrossRef] [PubMed]
- Gu, H.F. Morphology, phylogenetic position, and ecophysiology of Alexandrium ostenfeldii (Dinophyceae) from the Bohai Sea, China. J. Syst. Evol. 2011, 49, 606–616. [Google Scholar] [CrossRef]
- Ajani, P.A.; Larsson, M.E.; Rubio-Zuazo, A.; Bush, S.; Brett, S.; Farrell, H. Modelling bloom formation of the toxic dinoflagellates Dinophysis acuminata and Dinophysis caudata in a highly modified estuary, south eastern Australia. Estuar. Coast. Shelf Sci. 2016, 183, 95–106. [Google Scholar] [CrossRef]
- Basti, L.; Uchida, H.; Matsushima, R.; Watanabe, R.; Suzuki, T.; Yamatogi, T.; Nagai, S. Influence of Temperature on Growth and Production of Pectenotoxin-2 by a Monoclonal Culture of Dinophysis caudata. Mar. Drugs 2015, 13, 7124–7137. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Basti, L.; Suzuki, T.; Uchida, H.; Kamiyama, T.; Nagai, S. Thermal acclimation affects growth and lipophilic toxin production in a strain of cosmopolitan harmful alga Dinophysis acuminata. Harmful Algae 2018, 73, 119–128. [Google Scholar] [CrossRef]
- Justic, D.; Rabalais, N.N.; Turner, R.E. Stioichiometry nutrient balance and origin of coastal eutrophication. Mar. Pollut. Bull. 1995, 30, 41–46. [Google Scholar] [CrossRef]
- Li, Y.; Li, D.J.; Tang, J.L.; Wang, Y.M.; Liu, Z.G.; He, S.Q. Long-term changes in the Changjiang estuary plankton community related to anthropogenic eutrophication. Aquat. Ecosyst. Health 2010, 13, 66–72. [Google Scholar] [CrossRef]
- Li, H.M.; Tang, H.J.; Shi, X.Y.; Zhang, C.S.; Wang, X.L. Increased nutrient loads from the Changjiang (Yangtze) River have led to increased Harmful Algal Blooms. Harmful Algae 2014, 39, 92–101. [Google Scholar] [CrossRef]
- Parsons, T.R.; Maita, Y.; Lalli, C.M. A Manual of Chemical and Biological Methods for Seawater Analysis; Pergamon Press: New York, NY, USA, 1984. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variables | OA | GYM | PTX2 | DTX1 |
|---|---|---|---|---|
| T | −0.480 * | −0.258 | −0.502 * | 0.244 |
| S | 0.770 ** | 0.308 | 0.555 ** | −0.186 |
| DO | 0.580 ** | 0.702 ** | 0.843 ** | −0.104 |
| pH | 0.236 | −0.176 | −0.239 | 0.195 |
| TSS | −0.265 | −0.171 | −0.278 | 0.105 |
| Chla | −0.008 | −0.021 | 0.107 | 0.037 |
| PO4 | −0.535 ** | −0.306 | −0.429 * | −0.118 |
| SiO3 | −0.805 ** | −0.335 | −0.566 ** | 0.253 |
| NH4 | −0.023 | 0.234 | 0.414 * | 0.080 |
| NO2 | −0.089 | −0.266 | −0.125 | 0.146 |
| NO3 | −0.821 ** | −0.348 | −0.584 ** | 0.208 |
| DIN | −0.823 ** | −0.348 | −0.583 ** | 0.210 |
| Compounds | Molecular Formula | Retention Time (min) | Segment Time (min) | Precursor Ion (m/z) | Qualitative/Quantitative Ion (m/z) | Collision Energy/V |
|---|---|---|---|---|---|---|
| OAa | C44H68O13 | 12.6 | 8.5–16.0 | 827.5 [M+Na]+ | 809.6/723.5 | 1.0 |
| YTXb | C55H82O21S2 | 20.2 | 16.0–24.5 | 1141.7 [M-H]− | 1123.7/1061.8 | 1.5 |
| DTX1c | C45H70O13 | 19.2 | 16.0–24.5 | 841.5 [M+H]+ | 823.5/737.5 | 1.5 |
| AZA1d | C47H71NO12 | 24.9 | 24.5–30.2 | 842.5 [M+H]+ | 824.4/806.4 | 0.9 |
| AZA2e | C48H73NO12 | 25.6 | 24.5–30.2 | 856.5 [M+H]+ | 838.5/820.7 | 1.1 |
| GYMf | C32H45NO4 | 32.0 | 30.2–34.8 | 508.4 [M+H]+ | 490.4/392.4 | 1.0 |
| SPX1g | C42H61NO7 | 35.3 | 34.8–48.0 | 692.4 [M+H]+ | 674.4/656.4 | 1.1 |
| PTX2h | C47H70O14 | 36.0 | 34.8–48.0 | 876.7 [M+NH4]+ | 805.7/823.6 | 1.0 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
He, X.; Chen, J.; Wu, D.; Sun, P.; Ma, X.; Wang, J.; Liu, L.; Chen, K.; Wang, B. Distribution Characteristics and Environmental Control Factors of Lipophilic Marine Algal Toxins in Changjiang Estuary and the Adjacent East China Sea. Toxins 2019, 11, 596. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins11100596
He X, Chen J, Wu D, Sun P, Ma X, Wang J, Liu L, Chen K, Wang B. Distribution Characteristics and Environmental Control Factors of Lipophilic Marine Algal Toxins in Changjiang Estuary and the Adjacent East China Sea. Toxins. 2019; 11(10):596. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins11100596
Chicago/Turabian StyleHe, Xiuping, Junhui Chen, Danni Wu, Ping Sun, Xin Ma, Jiuming Wang, Lijun Liu, Kan Chen, and Baodong Wang. 2019. "Distribution Characteristics and Environmental Control Factors of Lipophilic Marine Algal Toxins in Changjiang Estuary and the Adjacent East China Sea" Toxins 11, no. 10: 596. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins11100596