Degradation of Aflatoxin B1 and Zearalenone by Bacterial and Fungal Laccases in Presence of Structurally Defined Chemicals and Complex Natural Mediators

 ,
,
Abstract
:1. Introduction
2. Results
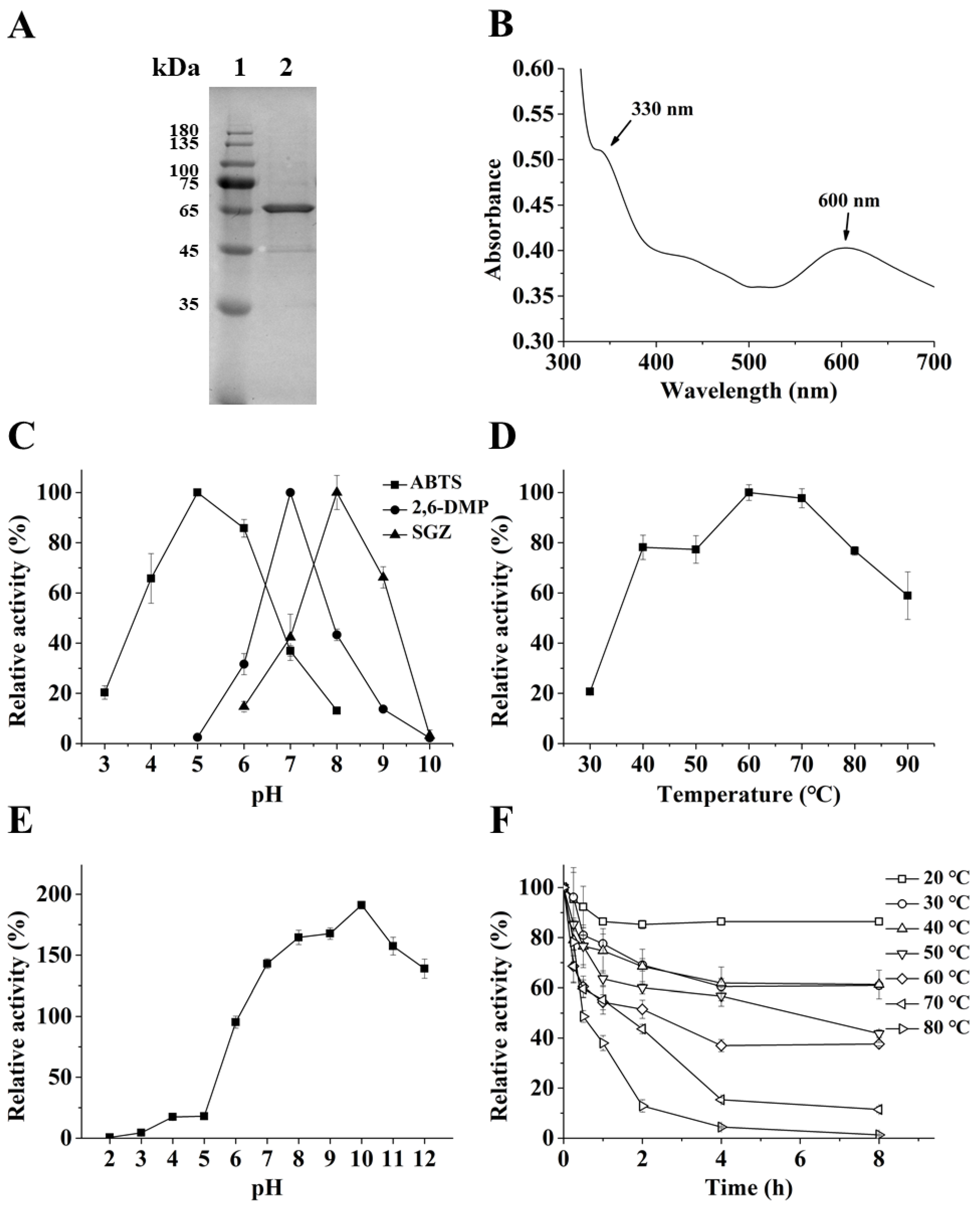
2.1. Biochemical Properties of the CotA laccase from Bacillus subtilis (BsCotA)
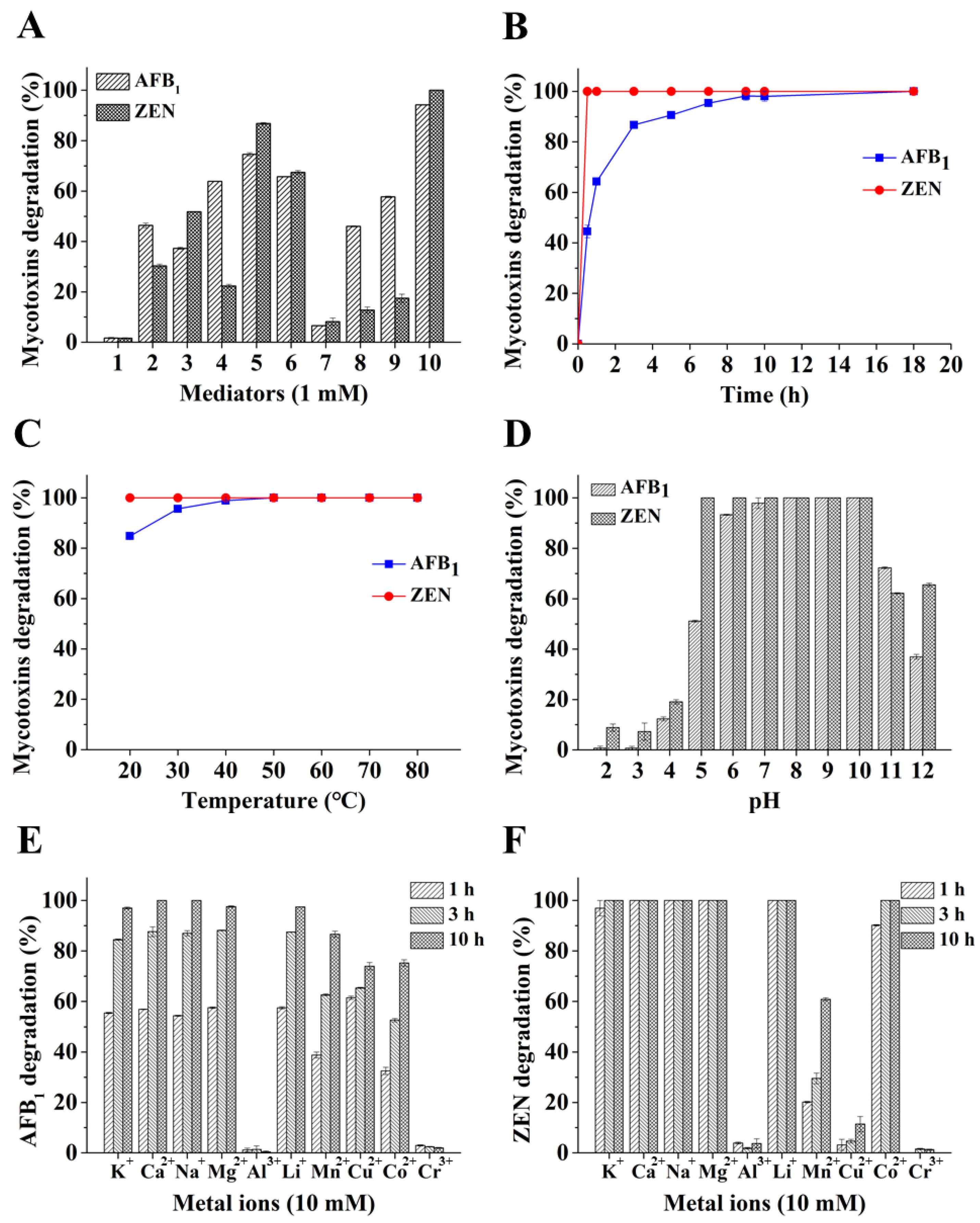
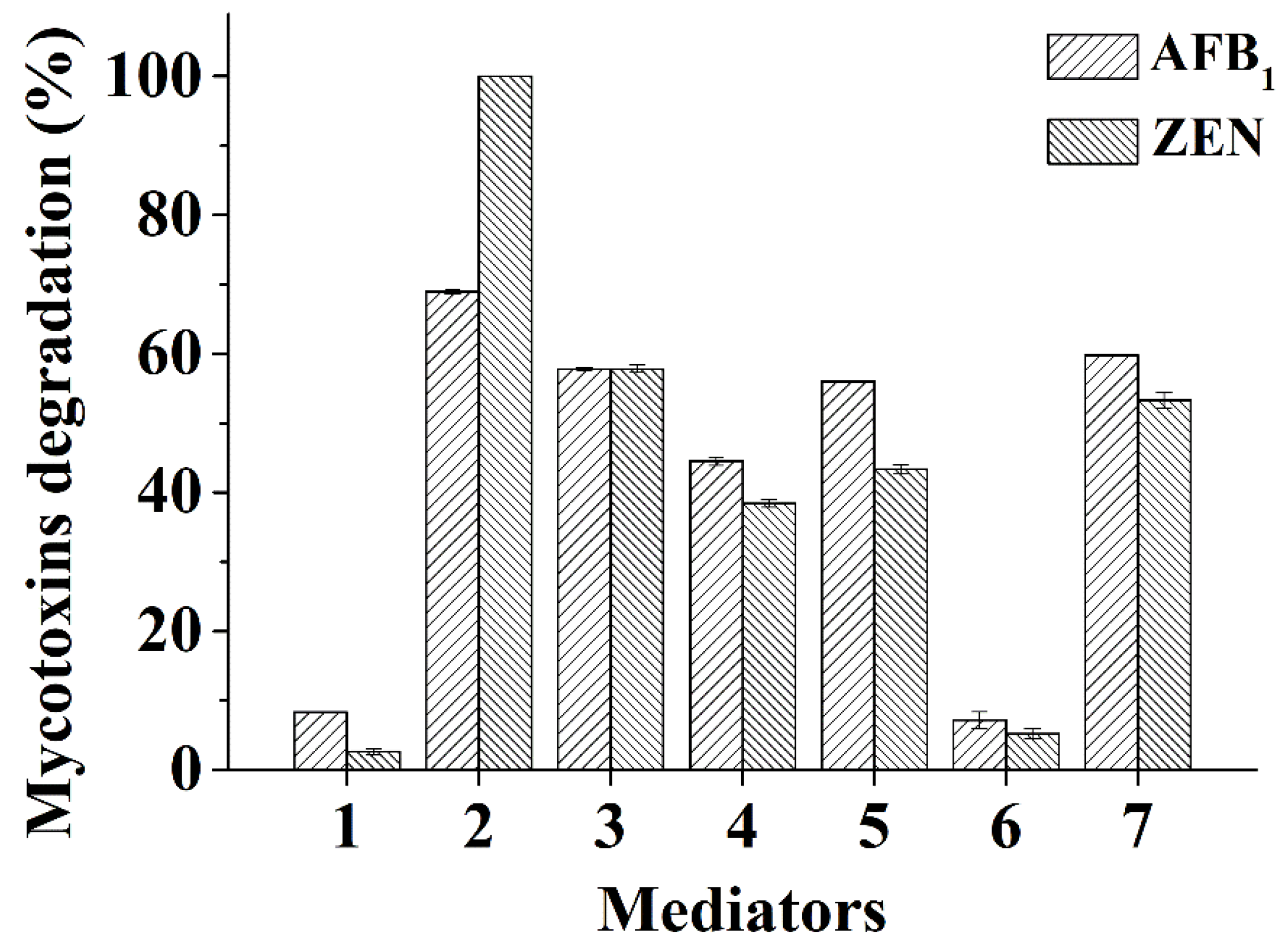
2.2. Degradation of Mycotoxins by BsCotA with Structurally Defined Chemical Compounds as Mediators
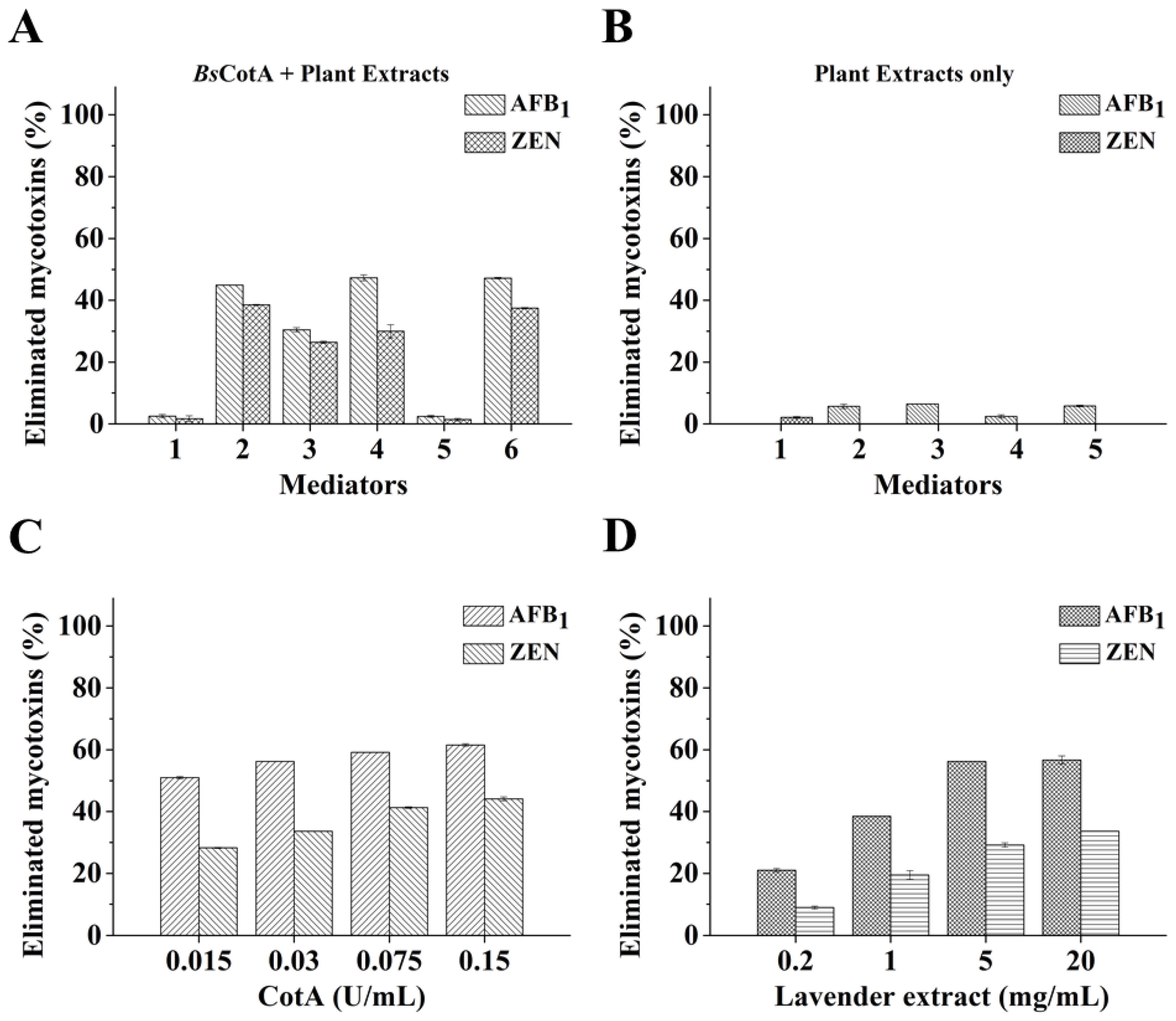
2.3. Degradation of Mycotoxins by BsCotA with Plant Extracts as a Natural Mediator
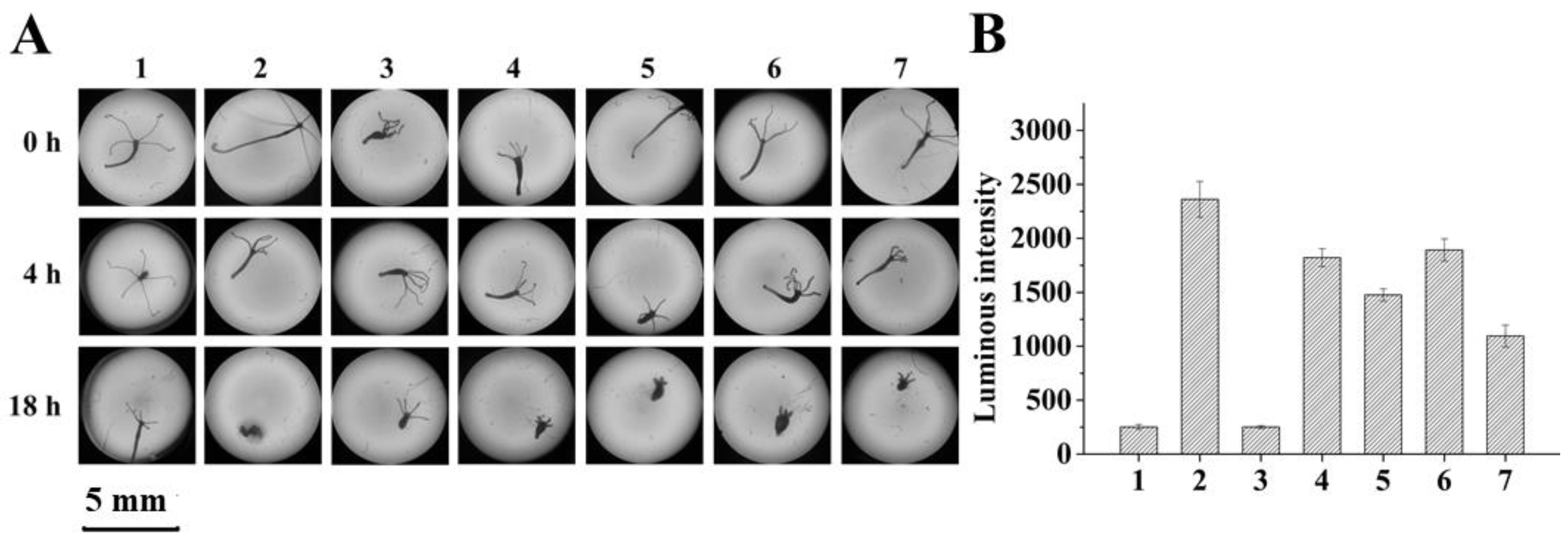
2.4. Evidence for AFB1 and ZEN Detoxification by BsCotA/Mediator Treatment
2.5. Degradation of AFB1 and ZEN by a Fungal Laccase
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Chemicals and Other Materials
5.2. Bacterial Strains and Culture Conditions
5.3. Cloning, Expression, and Purification of Recombinant BsCotA
5.4. Determining the Laccase Activity
5.5. Biochemical Characterization of Recombinant BsCotA
5.6. Mycotoxins Degradation by the Laccase/Mediator Systems
5.7. Hydra Assay
5.8. BLYES Assay
5.9. HPLC Analysis
5.10. Statistical Analyses
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| AFB1 | Aflatoxin B1 |
| ZEN | Zearalenone |
| DON | Deoxynivalenol |
| FB1 | Fumonisin B1 |
References
- Broom, L. Mycotoxins and the intestine. Anim. Nutr. 2015, 1, 262–265. [Google Scholar] [CrossRef] [PubMed]
- Ji, C.; Fan, Y.; Zhao, L. Review on biological degradation of mycotoxins. Anim. Nutr. 2016, 2, 127–133. [Google Scholar] [CrossRef] [PubMed]
- Guan, S.; Zhou, T.; Yin, Y.; Xie, M. Microbial strategies to control aflatoxins in food and feed. World Mycotoxin J. 2011, 4, 413–424. [Google Scholar] [CrossRef] [Green Version]
- Mitchell, N.J.; Bowers, E.; Hurburgh, C.; Wu, F. Potential economic losses to the US corn industry from aflatoxin contamination. Food Addit. Contam. Part A Chem. Anal. Control Expo. Risk Assess. 2016, 33, 540–550. [Google Scholar] [CrossRef]
- Zhou, G.; Chen, Y.; Kong, Q.; Ma, Y.; Liu, Y. Detoxification of aflatoxin B1 by Zygosaccharomyces rouxii with solid state fermentation in peanut meal. Toxins 2017, 9, 42. [Google Scholar] [CrossRef]
- Nielsen, L.K.; Cook, D.J.; Edwards, S.G.; Ray, R.V. The prevalence and impact of Fusarium head blight pathogens and mycotoxins on malting barley quality in UK. Int. J. Food Microbiol. 2014, 179, 38–49. [Google Scholar] [CrossRef] [Green Version]
- Stoev, S.D. Foodborne mycotoxicoses, risk assessment and underestimated hazard of masked mycotoxins and joint mycotoxin effects or interaction. Environ. Toxicol. Pharmacol. 2015, 39, 794–809. [Google Scholar] [CrossRef]
- Gromadzka, K.; Waskiewicz, A.; Chelkowski, J.; Golinski, P. Zearalenone and its metabolites: Occurrence, detection, toxicity and guidelines. World Mycotoxin J. 2008, 1, 209–220. [Google Scholar] [CrossRef]
- Schoevers, E.J.; Santos, R.R.; Colenbrander, B.; Fink-Gremmels, J.; Roelen, B.A.J. Transgenerational toxicity of zearalenone in pigs. Reprod. Toxicol. 2012, 34, 110–119. [Google Scholar] [CrossRef]
- Zinedine, A.; Soriano, J.M.; Molto, J.C.; Manes, J. Review on the toxicity, occurrence, metabolism, detoxification, regulations and intake of zearalenone: An oestrogenic mycotoxin. Food Chem. Toxicol. 2007, 45, 1–18. [Google Scholar] [CrossRef]
- Fernández Juri, M.G.; Dalcero, A.M.; Magnoli, C.E. In vitro aflatoxin B1 binding capacity by two Enterococcus faecium strains isolated from healthy dog faeces. J. Appl. Microbiol. 2015, 118, 574–582. [Google Scholar] [CrossRef] [PubMed]
- Yu, Y.; Wu, H.; Tang, Y.; Qiu, L. Cloning, expression of a peroxiredoxin gene from Acinetobacter sp. SM04 and characterization of its recombinant protein for zearalenone detoxification. Microbiol. Res. 2012, 167, 121–126. [Google Scholar] [CrossRef] [PubMed]
- Reiss, R.; Ihssen, J.; Richter, M.; Eichhorn, E.; Schilling, B.; Thöny-Meyer, L. Laccase versus laccase-like multi-copper oxidase: A comparative study of similar enzymes with diverse substrate spectra. PLoS ONE 2013, 8, e65633. [Google Scholar] [CrossRef] [PubMed]
- Karami, C.; Taher, M.A. A catechol biosensor based on immobilizing laccase to Fe3O4@ Au core-shell nanoparticles. Int. J. Biol. Macromol. 2019, 129, 84–90. [Google Scholar] [CrossRef]
- Alberts, J.F.; Gelderblom, W.C.A.; Botha, A.; Van Zyl, W.H. Degradation of aflatoxin B1 by fungal laccase enzymes. Int. J. Food Microbiol. 2009, 135, 47–52. [Google Scholar] [CrossRef]
- Banu, L.; Lupu, A.; Aprodu, L. Degradation of zearalenone by laccase enzyme. Sci. Study Res. Chem. Chem. Eng. Biotechnol. Food Ind. 2013, 14, 79–84. [Google Scholar]
- Jones, S.M.; Solomon, E.I. Electron transfer and reaction mechanism of laccases. Cell. Mol. Life Sci. 2015, 72, 869–883. [Google Scholar] [CrossRef] [Green Version]
- Arzola, K.G.; Arévalo, M.C.; Falcón, M.A. Catalytic efficiency of natural and synthetic compounds used as laccase-mediators in oxidising veratryl alcohol and a kraft lignin, estimated by electrochemical analysis. Electrochim. Acta 2009, 54, 2621–2629. [Google Scholar] [CrossRef]
- Couto, S.R.; Herrera, J.L.T. Industrial and biotechnological applications of laccases: A review. Biotechnol. Adv. 2006, 24, 500–513. [Google Scholar] [CrossRef]
- Liang, S.; Luo, Q.; Huang, Q. Degradation of sulfadimethoxine catalyzed by laccase with soybean meal extract as natural mediator: Mechanism and reaction pathway. Chemosphere 2017, 181, 320–327. [Google Scholar] [CrossRef] [Green Version]
- Camarero, S.; Garcıa, O.; Vidal, T.; Colom, J.; del Rıo, J.C.; Gutiérrez, A.; Gras, J.M.; Monje, R.; Martınez, M.J.; Martınez, Á.T. Efficient bleaching of non-wood high-quality paper pulp using laccase-mediator system. Enzym. Microb. Technol. 2004, 35, 113–120. [Google Scholar] [CrossRef] [Green Version]
- Riva, S. Laccases: Blue enzymes for green chemistry. Trends Biotechnol. 2006, 24, 219–226. [Google Scholar] [CrossRef] [PubMed]
- Loi, M.; Fanelli, F.; Cimmarusti, M.T.; Mirabelli, V.; Haidukowski, M.; Logrieco, A.F.; Caliandro, R.; Mule, G. In vitro single and combined mycotoxins degradation by Ery4 laccase from Pleurotus eryngii and redox mediators. Food Control 2018, 90, 401–406. [Google Scholar] [CrossRef]
- Martins, L.O.; Soares, C.M.; Pereira, M.M.; Teixeira, M.; Costa, T.; Jones, G.H.; Henriques, A.O. Molecular and biochemical characterization of a highly stable bacterial laccase that occurs as a structural component of the Bacillus subtilis endospore coat. J. Biol. Chem. 2002, 277, 18849–18859. [Google Scholar] [CrossRef]
- Grass, G.; Rensing, C. CueO is a multi-copper oxidase that confers copper tolerance in Escherichia coli. Biochem. Biophys. Res. Commun. 2001, 286, 902–908. [Google Scholar] [CrossRef]
- Hullo, M.F.; Moszer, I.; Danchin, A.; Isabelle, M.-V. CotA of Bacillus subtilis is a copper-dependent laccase. J. Bacteriol. 2001, 183, 5426–5430. [Google Scholar] [CrossRef]
- Johannes, C.; Majcherczyk, A. Laccase activity tests and laccase inhibitors. J. Biotechnol. 2000, 78, 193–199. [Google Scholar] [CrossRef]
- Wang, C.L.; Cui, D.Z.; Lu, L.; Zhang, N.; Yang, H.Y.; Zhao, M.; Dai, S.J. Cloning and characterization of CotA laccase from Bacillus subtilis WD23 decoloring dyes. Ann. Microbiol. 2016, 66, 461–467. [Google Scholar] [CrossRef]
- Lu, L.; Wang, T.N.; Xu, T.F.; Wang, J.Y.; Wang, C.L.; Zhao, M. Cloning and expression of thermo-alkali-stable laccase of Bacillus licheniformis in Pichia pastoris and its characterization. Bioresour. Technol. 2013, 134, 81–86. [Google Scholar] [CrossRef]
- Loi, M.; Fanelli, F.; Zucca, P.; Liuzzi, V.; Quintieri, L.; Cimmarusti, M.; Monaci, L.; Haidukowski, M.; Logrieco, A.; Sanjust, E.; et al. Aflatoxin B1 and M1 degradation by Lac2 from Pleurotus pulmonarius and redox mediators. Toxins 2016, 8, 245. [Google Scholar] [CrossRef]
- Kabak, B.; Var, I. Factors affecting the removal of aflatoxin M1 from food model by Lactobacillus and Bifidobacterium strains. J. Environ. Sci. Health B 2008, 43, 617–624. [Google Scholar] [CrossRef] [PubMed]
- Raters, M.; Matissek, R. Thermal stability of aflatoxin B1 and ochratoxin A. Mycotoxin Res. 2008, 24, 130–134. [Google Scholar] [CrossRef] [PubMed]
- Ryu, D.; Hanna, M.A.; Eskridge, K.M.; Bullerman, L.B. Heat stability of zearalenone in an aqueous buffered model system. J. Agric. Food Chem. 2003, 51, 1746–1748. [Google Scholar] [CrossRef] [PubMed]
- Krifaton, C.; Kriszt, B.; Risa, A.; Szoboszlay, S.; Cserháti, M.; Harkai, P.; Eldridge, M.; Wang, J.; Kukolya, J. Application of a yeast estrogen reporter system for screening zearalenone degrading microbes. J. Hazard. Mater. 2013, 244, 429–435. [Google Scholar] [CrossRef]
- Giardina, P.; Faraco, V.; Pezzella, C.; Piscitelli, A.; Vanhulle, S.; Sannia, G. Laccases: A never-ending story. Cell. Mol. Life Sci. 2010, 67, 369–385. [Google Scholar] [CrossRef]
- Liu, Y.; Huang, L.; Guo, W.; Jia, L.; Fu, Y.; Gui, S.; Lu, F. Cloning, expression, and characterization of a thermostable and pH-stable laccase from Klebsiella pneumoniae and its application to dye decolorization. Process Biochem. 2017, 53, 125–134. [Google Scholar] [CrossRef]
- Kumar, S.; Jain, K.K.; Rani, S.; Bhardwaj, K.N.; Goel, M.; Kuhad, R.C. In-vitro refolding and characterization of recombinant laccase (CotA) from Bacillus pumilus MK001 and its potential for phenolics degradation. Mol. Biotechnol. 2016, 58, 789–800. [Google Scholar] [CrossRef]
- Reiss, R.; Ihssen, J.; Thöny-Meyer, L. Bacillus pumilus laccase: A heat stable enzyme with a wide substrate spectrum. BMC Biotechnol. 2011, 11, 9. [Google Scholar] [CrossRef]
- Basheer, S.; Rashid, N.; Akram, M.S.; Akhtar, M. A highly stable laccase from Bacillus subtilis strain R5: Gene cloning and characterization. Biosci. Biotechnol. Biochem. 2019, 83, 436–445. [Google Scholar] [CrossRef]
- Meng, Y.T.; Zheng, Y.M.; Zhang, L.M.; He, J.Z. Biogenic Mn oxides for effective adsorption of Cd from aquatic environment. Environ. Pollut. 2009, 157, 2577–2583. [Google Scholar] [CrossRef]
- Navas, L.E.; Martinez, F.D.; Taverna, M.E.; Fetherolf, M.M.; Eltis, L.D.; Nicolau, V.; Estenoz, D.; Campos, E.; Benintende, G.B.; Berretta, M.F. A thermostable laccase from Thermus sp. 2.9 and its potential for delignification of Eucalyptus biomass. AMB Express 2019, 9, 24. [Google Scholar] [CrossRef] [PubMed]
- Guan, Z.; Song, C.; Zhang, N.; Zhou, W.; Xu, C.; Zhou, L.; Zhao, H.; Cai, Y.; Liao, X. Overexpression, characterization, and dye-decolorizing ability of a thermostable, pH-stable, and organic solvent-tolerant laccase from Bacillus pumilus W3. J. Mol. Catal. B Enzym. 2014, 101, 1–6. [Google Scholar] [CrossRef]
- Wang, T.N.; Lu, L.; Wang, J.Y.; Xu, T.F.; Li, J.; Zhao, M. Enhanced expression of an industry applicable CotA laccase from Bacillus subtilis in Pichia pastoris by non-repressing carbon sources together with pH adjustment: Recombinant enzyme characterization and dye decolorization. Process Biochem. 2015, 50, 97–103. [Google Scholar] [CrossRef]
- Fang, Z.M.; Zhou, P.; Chang, F.; Yin, Q.; Fang, W.; Yuan, J.; Zhang, X.C.; Xiao, Y.Z. Structure-based rational design to enhance the solubility and thermostability of a bacterial laccase Lac15. PLoS ONE 2014, 9, e102423. [Google Scholar] [CrossRef]
- Chauhan, P.S.; Goradia, B.; Saxena, A. Bacterial laccase: Recent update on production, properties and industrial applications. 3 Biotech 2017, 7, 323. [Google Scholar] [CrossRef]
- Singh, G.; Capalash, N.; Goel, R.; Sharma, P. A pH-stable laccase from alkali-tolerant γ-proteobacterium JB: Purification, characterization and indigo carmine degradation. Enzym. Microb. Technol. 2007, 41, 794–799. [Google Scholar] [CrossRef]
- Ashe, B.; Nguyen, L.N.; Hai, F.I.; Lee, D.J.; van de Merwe, J.P.; Leusch, F.D.L.; Price, W.E.; Nghiem, L.D. Impacts of redox-mediator type on trace organic contaminants degradation by laccase: Degradation efficiency, laccase stability and effluent toxicity. Int. Biodeterior. Biodegrad. 2016, 113, 169–176. [Google Scholar] [CrossRef] [Green Version]
- Astolfi, P.; Brandi, P.; Galli, C.; Gentili, P.; Gerini, M.F.; Greci, L.; Lanzalunga, O. New mediators for the enzyme laccase: Mechanistic features and selectivity in the oxidation of non-phenolic substrates. New J. Chem. 2005, 29, 1308–1317. [Google Scholar] [CrossRef]
- Guo, M.Y.; Xu, Y.Q.; Ren, L.; He, S.Z.; Pang, X.H. A systematic study on DNA barcoding of medicinally important genus Epimedium L. (Berberidaceae). Genes 2018, 9, 637. [Google Scholar] [CrossRef]
- Xiong, M.; Li, Y.; Tang, P.; Zhang, Y.; Cao, M.; Ni, J.; Xing, M. Effectiveness of aromatherapy massage and inhalation on symptoms of depression in Chinese community-dwelling older adults. J. Altern. Complement. Med. 2018, 24, 717–724. [Google Scholar] [CrossRef]
- Zhou, Y.; Tang, N.Y.; Huang, L.J.; Zhao, Y.J.; Tang, X.Q.; Wang, K.C. Effects of salt stress on plant growth, antioxidant capacity, glandular trichome density, and volatile exudates of Schizonepeta tenuifolia Briq. Int. J. Mol. Sci. 2018, 19, 252. [Google Scholar] [CrossRef] [PubMed]
- Zapata-Castillo, P.; Villalonga-Santana, L.; Islas-Flores, I.; Rivera-Muñoz, G.; Ancona-Escalante, W.; Solís-Pereira, S. Synergistic action of laccases from Trametes hirsuta Bm2 improves decolourization of indigo carmine. Lett. Appl. Microbiol. 2015, 61, 252–258. [Google Scholar] [CrossRef] [PubMed]
- Jeon, J.R.; Murugesan, K.; Kim, Y.M.; Kim, E.J.; Chang, Y.S. Synergistic effect of laccase mediators on pentachlorophenol removal by Ganoderma lucidum laccase. Appl. Microbiol. Biotechnol. 2008, 81, 783–790. [Google Scholar] [CrossRef] [PubMed]
- McKenzie, K.S.; Sarr, A.B.; Mayura, K.; Bailey, R.H.; Miller, D.R.; Rogers, T.D.; Norred, W.P.; Voss, K.A.; Plattner, R.D.; Kubena, L.F.; et al. Oxidative degradation and detoxification of mycotoxins using a novel source of ozone. Food Chem. Toxicol. 1997, 35, 807–820. [Google Scholar] [CrossRef]
- Briones-Reyes, D.; Gomez-Martinez, L.; Cueva-Rolon, R. Zearalenone contamination in corn for human consumption in the state of Tlaxcala, Mexico. Food Chem. 2007, 100, 693–698. [Google Scholar] [CrossRef]
- Seitz, L.M. Comparison of methods for aflatoxin analysis by high-pressure liquid chromatography. J. Chromatogr. 1975, 104, 81–89. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Substrate | kcat (s−1) | Km (μM) | kcat/Km (M−1·s−1) |
|---|---|---|---|
| ABTS | 7.72 ± 0.67 | 178.73 ± 49.53 | (4.56 ± 0.90) × 104 |
| DMP | 2.73 ± 0.18 | (1.35 ± 0.27) × 103 | (2.09 ± 0.31) × 103 |
| SGZ | 2.39 ± 0.08 | 118.80 ± 12.36 | (2.04 ± 0.25) × 104 |
| Class | Compound | Structure |
|---|---|---|
| Laccase Model Substrate | ||
| ABTS |  | |
| DMP |  | |
| SGZ |  | |
| Structurally Defined Chemicals as Potential Mediators | ||
| p-Coumaric acid |  | |
| Syringic acid |  | |
| Vanillin |  | |
| Syringaldehyde |  | |
| Caffeic acid |  | |
| 1-Hydroxybenzotriazole |  | |
| Gallic acid |  | |
| Vanillic acid |  | |
| Methyl syringate |  | |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, X.; Bai, Y.; Huang, H.; Tu, T.; Wang, Y.; Wang, Y.; Luo, H.; Yao, B.; Su, X. Degradation of Aflatoxin B1 and Zearalenone by Bacterial and Fungal Laccases in Presence of Structurally Defined Chemicals and Complex Natural Mediators. Toxins 2019, 11, 609. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins11100609
Wang X, Bai Y, Huang H, Tu T, Wang Y, Wang Y, Luo H, Yao B, Su X. Degradation of Aflatoxin B1 and Zearalenone by Bacterial and Fungal Laccases in Presence of Structurally Defined Chemicals and Complex Natural Mediators. Toxins. 2019; 11(10):609. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins11100609
Chicago/Turabian StyleWang, Xiaolu, Yingguo Bai, Huoqing Huang, Tao Tu, Yuan Wang, Yaru Wang, Huiying Luo, Bin Yao, and Xiaoyun Su. 2019. "Degradation of Aflatoxin B1 and Zearalenone by Bacterial and Fungal Laccases in Presence of Structurally Defined Chemicals and Complex Natural Mediators" Toxins 11, no. 10: 609. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins11100609