Fusarium graminearum FgCWM1 Encodes a Cell Wall Mannoprotein Conferring Sensitivity to Salicylic Acid and Virulence to Wheat

 , , add
Show full author list
, , add
Show full author list
Abstract
:1. Introduction
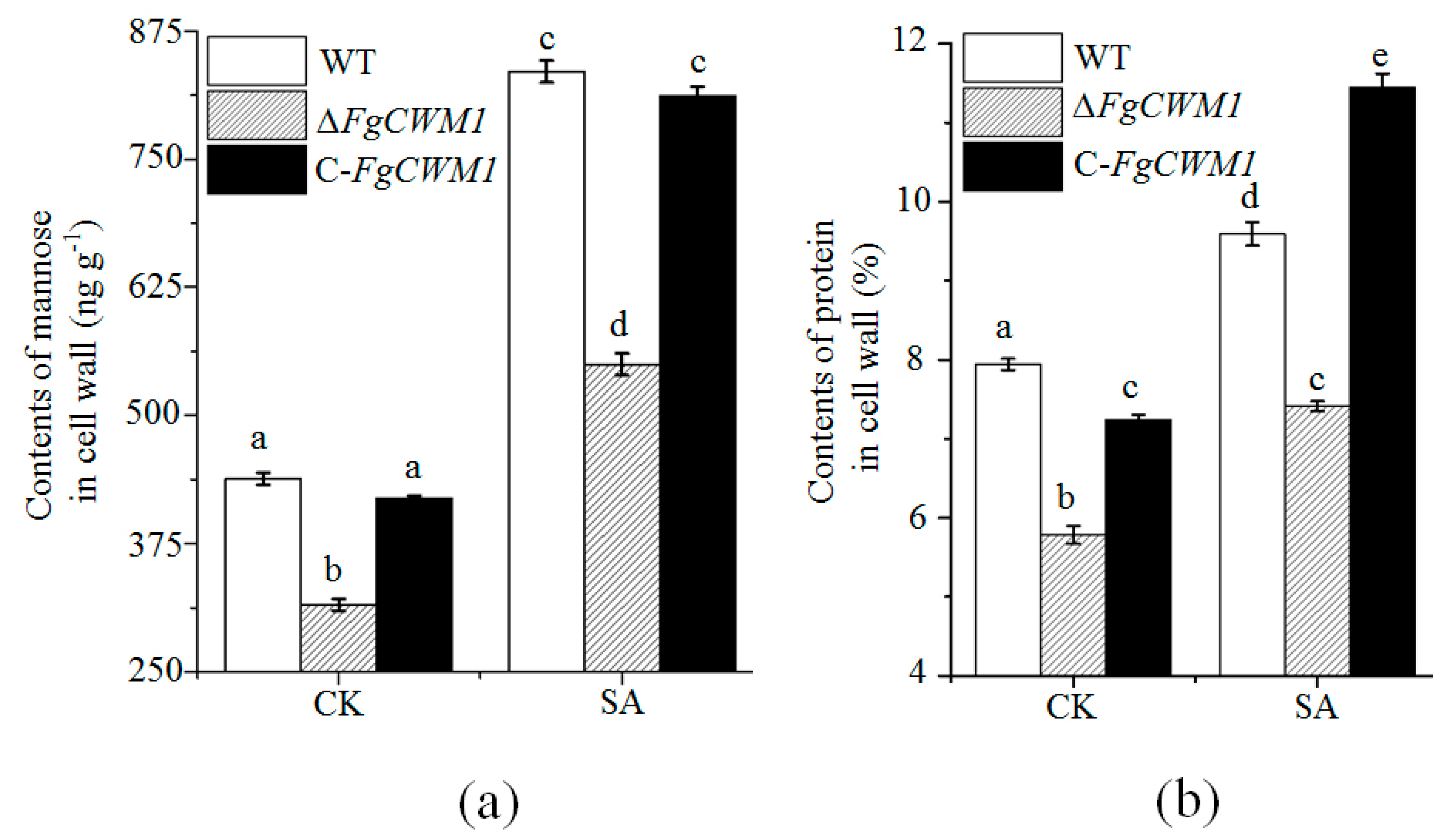
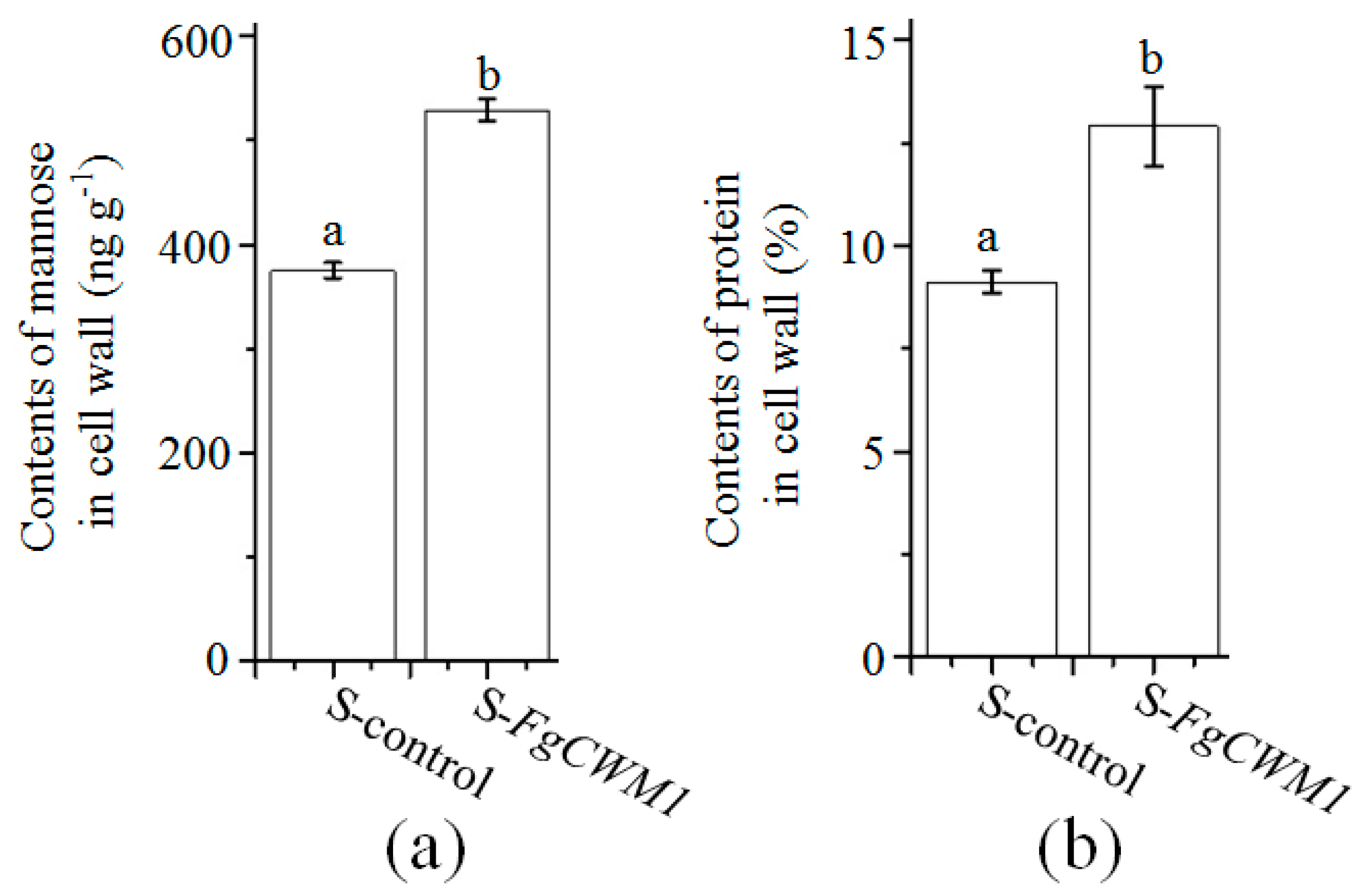
2. Results
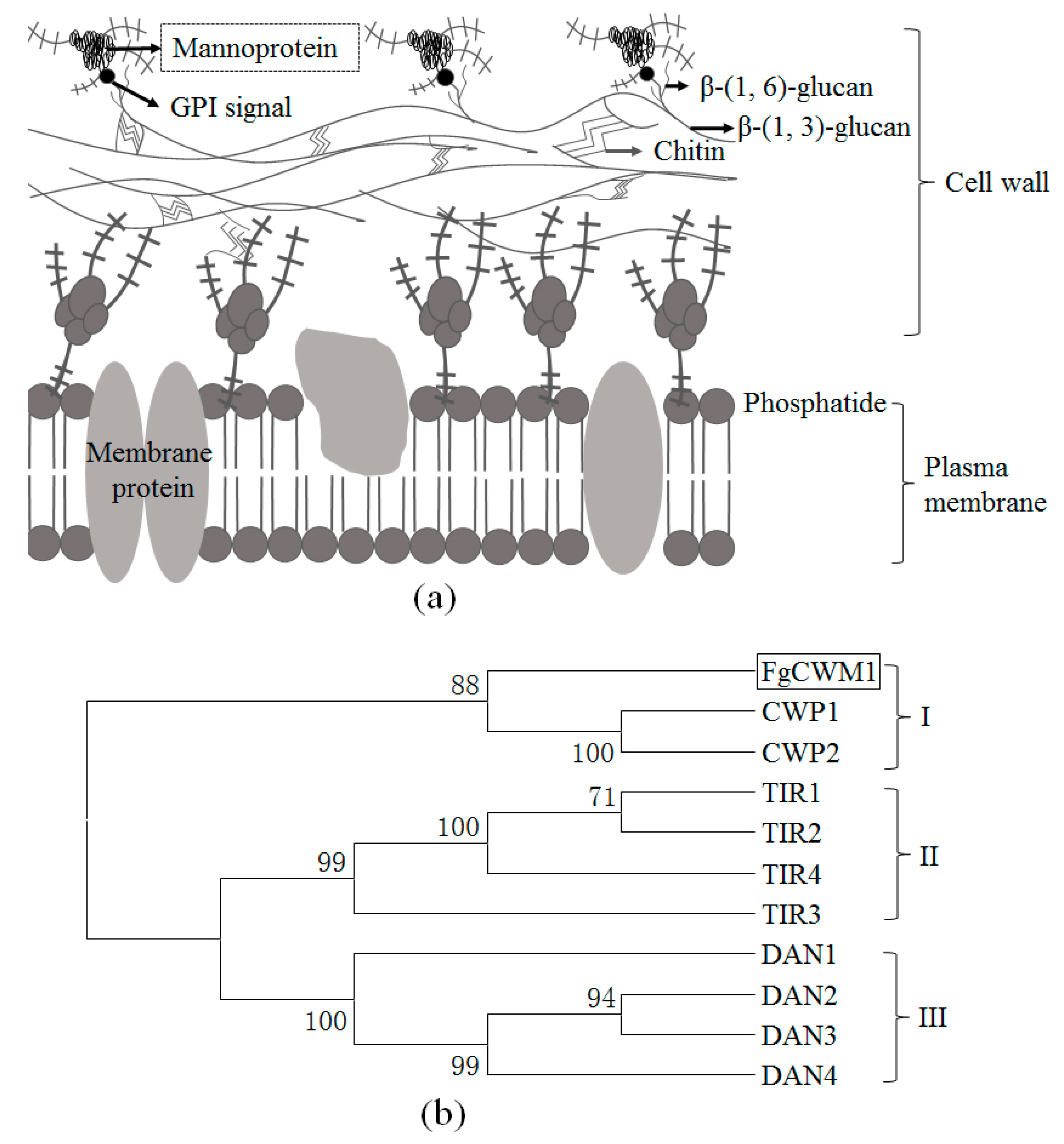
2.1. Sequence Analysis
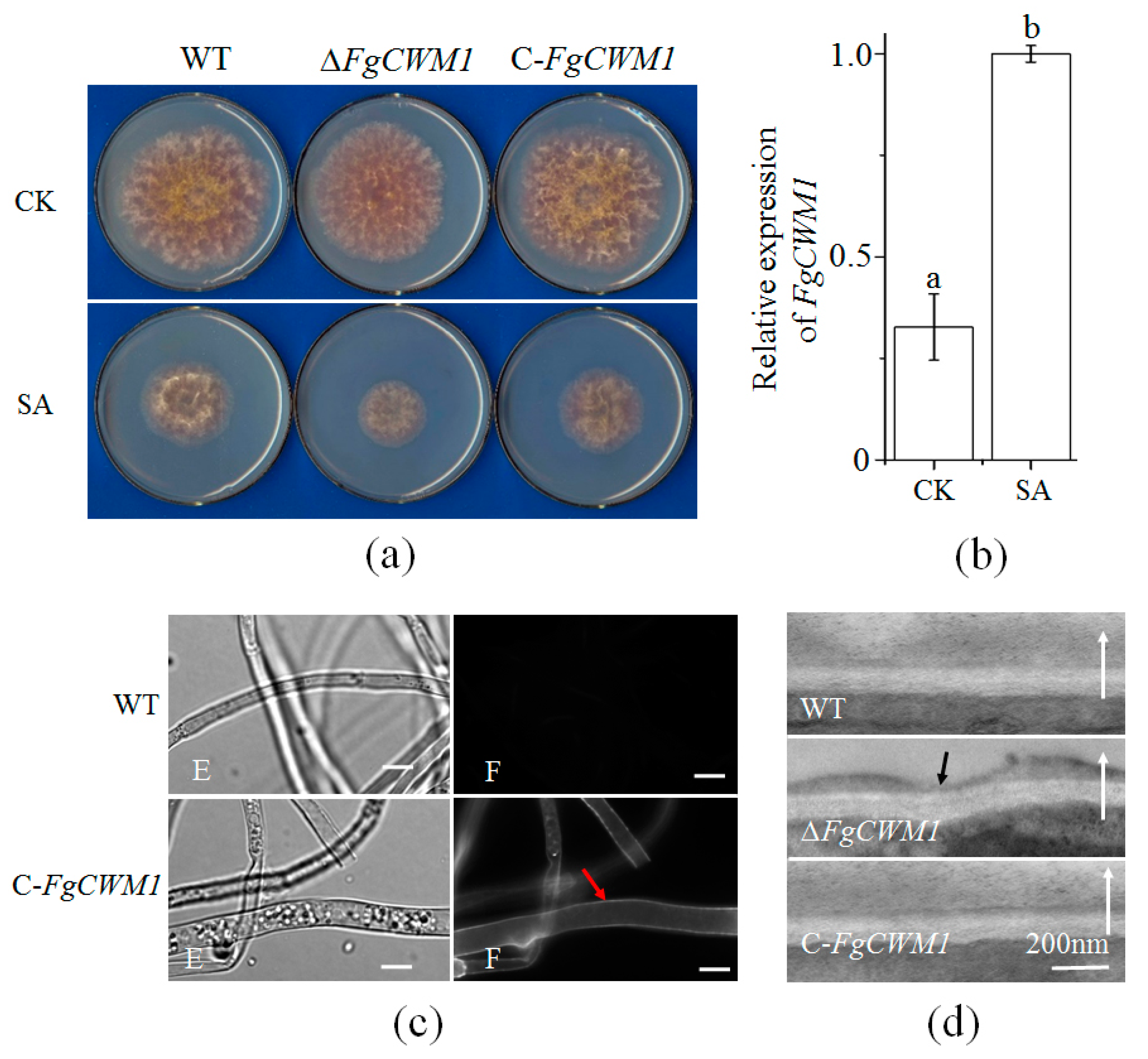
2.2. Creation of ΔFgCWM1 and C-FgCWM1
2.3. Effect of FgCWM1 on Mycelial Growth
2.4. Effect of FgCWM1 on Fungal Cell Wall Development
2.5. FgCWM1 Encodes a Cell Wall Mannoprotein
2.6. Effect of FgCWM1 on Pathogenicity
3. Discussion
4. Materials and Methods
4.1. Materials and Growth Conditions
4.2. Sequence Analysis
4.3. Construction of Deletion and Complementation Mutants
4.4. FgCWM1 Expression in S. cerevisiae
4.5. Determination of Mannose and Protein Contents in Fungal Cell Wall
4.6. Microscopic Assay
4.7. Virulence Assay and DON Measurement
4.8. Gene Expression Analysis
4.9. Quantification of SA in Wheat Spikes
4.10. Statistical Analysis
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Goswami, R.S.; Kistler, H.C. Heading for disaster: Fusarium graminearum on cereal crops. Mol. Plant Pathol. 2004, 5, 515–525. [Google Scholar] [CrossRef] [PubMed]
- Desjardins, A.E.; Hohn, T.M.; McCormick, S.P. Trichothecene biosynthesis in Fusarium: Chemistry, genetics, and significance. Microbiol. Rev. 1993, 57, 595–604. [Google Scholar] [PubMed]
- Fernando, W.G.; Paulitz, T.C.; Seaman, W.L.; Dutilleul, P.; Miller, J.D. Head blight gradients caused by Gibberella zeae from area sources of inoculum in wheat field plots. Phytopathology 1997, 87, 414–421. [Google Scholar] [CrossRef] [PubMed]
- McMullen, M.; Bergstrom, G.; De Wolf, E.; Dill-Macky, R.; Hershman, D.; Shaner, G.; Van Sanford, D. A unified effort to fight an enemy of wheat and barley: Fusarium Head Blight. Plant Dis. 2012, 96, 1712–1728. [Google Scholar] [CrossRef]
- Legrand, F.; Picot, A.; Cobo-Díaz, J.F.; Chen, W.; Le Floch, G. Challenges facing the biological control strategies for the management of Fusarium Head Blight of cereals caused by F. graminearum. Biol. Control 2017, 113, 26–38. [Google Scholar] [CrossRef]
- De Groot, P.W.; Ram, A.F.; Klis, F.M. Features and functions of covalently linked proteins in fungal cell walls. Fungal Genet. Biol. 2005, 42, 657–675. [Google Scholar] [CrossRef]
- Lionetti, V.; Giancaspro, A.; Fabri, E.; Giove, S.L.; Reem, N.; Zabotina, O.A.; Blanco, A.; Gadaleta, A.; Bellincampi, D. Cell wall traits as potential resources to improve resistance of durum wheat against Fusarium graminearum. BMC Plant Biol. 2015, 15, 6. [Google Scholar] [CrossRef]
- Chow, J.; Notaro, M.; Prabhakar, A.; Free, S.J.; Cullen, P.J. Impact of fungal MAPK pathway targets on the Cell Wall. J. Fungi 2018, 4, 93. [Google Scholar] [CrossRef]
- Schoffelmeer, E.A.; Klis, F.M.; Sietsma, J.H.; Cornelissen, B.J. The cell wall of Fusarium oxysporum. Fungal Genet. Biol. 1999, 27, 275–282. [Google Scholar] [CrossRef]
- Karkowska-Kuleta, J.; Kozik, A. Cell wall proteome of pathogenic fungi. Acta. Biochim. Pol. 2015, 62, 339–351. [Google Scholar] [CrossRef] [Green Version]
- Abramova, N.; Sertil, O.; Mehta, S.; Lowry, C.V. Reciprocal regulation of anaerobic and aerobic cell wall mannoprotein gene expression in Saccharomyces cerevisiae. J. Bacteriol. 2001, 183, 2881–2887. [Google Scholar] [CrossRef] [PubMed]
- Klis, F.M.; de Groot, P.; Hellingwerf, K. Molecular organization of the cell wall of Candida albicans. Med. Mycol. 2001, 39, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Klis, F.M.; Mol, P.; Hellingwerf, K.; Brul, S. Dynamics of cell wall structure in Saccharomyces cerevisiae. FEMS Microbiol. Rev. 2002, 26, 239–256. [Google Scholar] [CrossRef] [PubMed]
- Francois, J.M. Cell surface interference with plasma membrane and transport processes in yeasts. Adv. Exp. Med. Biol. 2016, 892, 11–31. [Google Scholar] [CrossRef]
- Van der Vaart, J.M.; Caro, L.H.; Chapman, J.W.; Klis, F.M.; Verrips, C.T. Identification of three mannoproteins in the cell wall of Saccharomyces cerevisiae. J. Bacteriol. 1995, 177, 3104–3110. [Google Scholar] [CrossRef]
- Dielbandhoesing, S.K.; Zhang, H.; Caro, L.H.; van der Vaart, J.M.; Klis, F.M.; Verrips, C.T.; Brul, S. Specific cell wall proteins confer resistance to nisin upon yeast cells. Appl. Environ. Microbiol. 1998, 64, 4047–4052. [Google Scholar]
- Zhang, M.; Liang, Y.P.; Zhang, X.H.; Xu, Y.; Dai, H.P.; Xiao, W. Deletion of yeast CWP genes enhances cell permeability to genotoxic agents. Toxicol. Sci. 2008, 103, 68–76. [Google Scholar] [CrossRef]
- Henry, C.; Fontaine, T.; Heddergott, C.; Robinet, P.; Aimanianda, V.; Beau, R.; Beauvais, A.; Mouyna, I.; Prevost, M.C.; Fekkar, A.; et al. Biosynthesis of cell wall mannan in the conidium and the mycelium of Aspergillus fumigatus. Cell. Microbiol. 2016, 18, 1881–1891. [Google Scholar] [CrossRef]
- Du, T.; Ouyang, H.; Voglmeir, J.; Wilson, I.B.H.; Jin, C. Aspergillus fumigatus Mnn9 is responsible for mannan synthesis and required for covalent linkage of mannoprotein to the cell wall. Fungal Genet. Biol. 2019, 128, 20–28. [Google Scholar] [CrossRef]
- Sandini, S.; La Valle, R.; De Bernardis, F.; Macri, C.; Cassone, A. The 65 kDa mannoprotein gene of Candida albicans encodes a putative beta-glucanase adhesin required for hyphal morphogenesis and experimental pathogenicity. Cell. Microbiol. 2007, 9, 1223–1238. [Google Scholar] [CrossRef]
- Makandar, R.; Essig, J.S.; Schapaugh, M.A.; Trick, H.N.; Shah, J. Genetically engineered resistance to Fusarium head blight in wheat by expression of Arabidopsis NPR1. Mol. Plant Microbe Interact. 2006, 19, 123–129. [Google Scholar] [CrossRef] [PubMed]
- Qi, P.F.; Johnston, A.; Balcerzak, M.; Rocheleau, H.; Harris, L.J.; Long, X.Y.; Wei, Y.M.; Zheng, Y.L.; Ouellet, T. Effect of salicylic acid on Fusarium graminearum, the major causal agent of fusarium head blight in wheat. Fungal Biol. 2012, 116, 413–426. [Google Scholar] [CrossRef] [PubMed]
- Qi, P.F.; Zhang, Y.Z.; Liu, C.H.; Zhu, J.; Chen, Q.; Guo, Z.R.; Wang, Y.; Xu, B.J.; Zheng, T.; Jiang, Y.F.; et al. Fusarium graminearum ATP-Binding cassette transporter gene FgABCC9 is required for its transportation of salicylic acid, fungicide resistance, mycelial growth and pathogenicity towards wheat. Int. J. Mol. Sci. 2018, 19, 2351. [Google Scholar] [CrossRef] [PubMed]
- Qi, P.F.; Zhang, Y.Z.; Liu, C.H.; Chen, Q.; Guo, Z.R.; Wang, Y.; Xu, B.J.; Jiang, Y.F.; Zheng, T.; Gong, X.; et al. Functional analysis of FgNahG clarifies the contribution of salicylic acid to wheat (Triticum aestivum) resistance against Fusarium Head Blight. Toxins 2019, 11, 59. [Google Scholar] [CrossRef] [PubMed]
- Qi, P.F.; Balcerzak, M.; Rocheleau, H.; Leung, W.; Wei, Y.M.; Zheng, Y.L.; Ouellet, T. Jasmonic acid and abscisic acid play important roles in host-pathogen interaction between Fusarium graminearum and wheat during the early stages of fusarium head blight. Physiol. Mol. Plant. Pathol. 2016, 93, 39–48. [Google Scholar] [CrossRef]
- Uknes, S.; Mauch-Mani, B.; Moyer, M.; Potter, S.; Williams, S.; Dincher, S.; Chandler, D.; Slusarenko, A.; Ward, E.; Ryals, J. Acquired resistance in Arabidopsis. Plant Cell 1992, 4, 645–656. [Google Scholar] [CrossRef]
- Sorahinobar, M.; Niknamet, V.; Ebrahimzadeh, H.; Soltanloo, H.; Behmanesh, M.; Enferadi, S.T. Central role of salicylic acid in resistance of wheat against Fusarium graminearum. J. Plant Growth Regul. 2016, 35, 477–491. [Google Scholar] [CrossRef]
- Zhang, Y.Z.; Chen, Q.; Liu, C.H.; Liu, Y.B.; Yi, P.; Niu, K.X.; Wang, Y.Q.; Wang, A.Q.; Yu, H.Y.; Pu, Z.E.; et al. Chitin synthase gene FgCHS8 affects virulence and fungal cell wall sensitivity to environmental stress in Fusarium graminearum. Fungal Biol. 2016, 120, 764–774. [Google Scholar] [CrossRef]
- Zhang, Y.Z.; Wei, Z.Z.; Liu, C.H.; Chen, Q.; Xu, B.J.; Guo, Z.R.; Cao, Y.L.; Wang, Y.; Han, Y.N.; Chen, C.; et al. Linoleic acid isomerase gene FgLAI12 affects sensitivity to salicylic acid, mycelial growth and virulence of Fusarium graminearum. Sci. Rep. 2017, 7, 46129. [Google Scholar] [CrossRef]
- Rocheleau, H.; Al-harthi, R.; Ouellet, T. Degradation of salicylic acid by Fusarium graminearum. Fungal Biol. 2019, 123, 77–86. [Google Scholar] [CrossRef]
- Harris, L.J.; Balcerzak, M.; Johnston, A.; Schneiderman, D.; Ouellet, T. Host-preferential Fusarium graminearum gene expression during infection of wheat, barley, and maize. Fungal Biol. 2016, 120, 111–123. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.B.; Li, H.P.; Zhang, J.B.; Song, B.; Chen, F.F.; Duan, X.J.; Xu, H.Q.; Liao, Y.C. Disruption of the chitin synthase gene CHS1 from Fusarium asiaticum results in an altered structure of cell walls and reduced virulence. Fungal Genet. Biol. 2010, 47, 205–215. [Google Scholar] [CrossRef] [PubMed]
- Li, G.Q.; Zhou, J.Y.; Jia, H.Y.; Gao, Z.X.; Fan, M.; Luo, Y.J.; Zhao, P.T.; Xue, S.; Li, N.; Yuan, Y.; et al. Mutation of a histidine-rich calcium-binding-protein gene in wheat confers resistance to Fusarium head blight. Nat. Genet. 2019, 51, 1106–1112. [Google Scholar] [CrossRef] [PubMed]
- Su, Z.Q.; Bernardo, A.; Tian, B.; Chen, H.; Wang, S.; Ma, H.X.; Cai, S.B.; Liu, D.T.; Zhang, D.D.; Li, T.; et al. A deletion mutation in TaHRC confers Fhb1 resistance to Fusarium head blight in wheat. Nat. Genet. 2019, 51, 1099–1105. [Google Scholar] [CrossRef] [PubMed]
- Koch, A.; Kumar, N.; Weber, L.; Keller, H.; Imani, J.; Kogel, K.H. Host-induced gene silencing of cytochrome P450 lanosterol C14 α-demethylase-encoding genes confers strong resistance to Fusarium species. Proc. Natl. Acad. Sci. USA 2013, 110, 19324–19329. [Google Scholar] [CrossRef] [PubMed]
- Cheng, W.; Song, X.S.; Li, H.P.; Cao, L.H.; Sun, K.; Qiu, X.L.; Xu, Y.B.; Yang, P.; Huang, T.; Zhang, J.B.; et al. Host-induced gene silencing of an essential chitin synthase gene confers durable resistance to Fusarium head blight and seedling blight in wheat. Plant Biotechnol. J. 2015, 13, 1335–1345. [Google Scholar] [CrossRef] [PubMed]
- Ding, L.; Xu, H.; Yi, H.; Yang, L.; Kong, Z.; Zhang, L.; Xue, S.; Jia, H.; Ma, Z. Resistance to hemi-biotrophic F. graminearum infection is associated with coordinated and ordered expression of diverse defense signaling pathways. PLoS ONE 2011, 6, e19008. [Google Scholar] [CrossRef]
- Ameye, M.; Audenaert, K.; De Zutter, N.; Steppe, K.; Van Meulebroek, L.; Vanhaecke, L.; De Vleesschauwer, D.; Haesaert, G.; Smagghe, G. Priming of wheat with the green leaf volatile Z-3-hexenyl acetate enhances defense against Fusarium graminearum but boosts deoxynivalenol production. Plant Physiol. 2015, 167, 1671–1684. [Google Scholar] [CrossRef]
- Qi, P.F.; Jiang, Y.F.; Guo, Z.R.; Chen, Q.; Ouellet, T.; Zong, L.J.; Wei, Z.Z.; Wang, Y.; Zhang, Y.Z.; Xu, B.J.; et al. Transcriptional reference map of hormone responses in wheat spikes. BMC Genom. 2019, 20, 390. [Google Scholar] [CrossRef]
- Mesterházy, A. Types and components of resistance to Fusarium head blight of wheat. Plant Breed. 1995, 114, 377–386. [Google Scholar] [CrossRef]
- Cappellini, R.A.; Peterson, J.L. Macroconidium formation in submerged cultures by a nonsporulating strain of Gibberella zeae. Mycologia 1965, 57, 962–966. [Google Scholar] [CrossRef]
- Tamura, K.; Peterson, D.; Peterson, N.; Stecher, G.; Nei, M.; Kumar, S. MEGA5: Molecular evolutionary genetics analysis using maximum likelihood, evolutionary distance, and maximum parsimony methods. Mol. Biol. Evol. 2011, 28, 2731–2739. [Google Scholar] [CrossRef] [PubMed]
- Lodhi, M.A.; Ye, G.N.; Weeden, N.F.; Reisch, B.I. A simple and efficient method for DNA extraction from grapevine cultivars and Vitis species. Plant Mol. Biol. Rep. 1994, 12, 6–13. [Google Scholar] [CrossRef]
- Frandsen, R.J.; Andersson, J.A.; Kristensen, M.B.; Giese, H. Efficient four fragment cloning for the construction of vectors for targeted gene replacement in filamentous fungi. BMC Mol. Biol. 2008, 9, 70. [Google Scholar] [CrossRef]
- Frandsen, R.J.; Frandsen, M.; Giese, H. Targeted gene replacement in fungal pathogens via Agrobacterium tumefaciens-mediated transformation. Methods Mol. Biol. 2012, 835, 17–45. [Google Scholar] [CrossRef]
- Maier, F.J.; Malz, S.; Losch, A.P.; Lacour, T.; Schafer, W. Development of a highly efficient gene targeting system for Fusarium graminearum using the disruption of a polyketide synthase gene as a visible marker. FEMS Yeast Res. 2005, 5, 653–662. [Google Scholar] [CrossRef]
- Kollár, R.; Petrakova, E.; Ashwell, G.; Robbins, P.W.; Cabib, E. Architecture of the yeast cell wall, the linkage between chitin and β(1→3)-glucan. J. Biol. Chem. 1995, 270, 1170–1178. [Google Scholar] [CrossRef]
- Cameron, D.R.; Cooper, D.G.; Neufeld, R.J. The mannoprotein of Saccharomyces cerevisiae is an effective bioemulsifier. Appl. Environ. Microbiol. 1988, 54, 1420–1425. [Google Scholar]
- Jung, S.; Rickert, D.A.; Deak, N.A.; Aldin, E.D.; Recknor, J.; Johnson, L.A.; Murphy, P.A. Comparison of kjeldahl and dumas methods for determining protein contents of soybean products. J. Am. Oil Chem. Soc. 2003, 80, 1169. [Google Scholar] [CrossRef]
- Martin-Urdiroz, M.; Roncero, M.I.; Gonzalez-Reyes, J.A.; Ruiz-Roldan, C. ChsVb, a class VII chitin synthase involved in septation, is critical for pathogenicity in Fusarium oxysporum. Eukaryot. Cell 2008, 7, 112–121. [Google Scholar] [CrossRef]
- Miller, D.; Blackwell, B.A. Biosynthesis of 3-acetyldeoxynivalenol and other metabolites by Fusarium culmorum HLK 1503 in a stirred jar fermentor. Can. J. Bot. 1986, 64, 1–5. [Google Scholar] [CrossRef]
- Siciliano, I.; Amaral Carneiro, G.; Spadaro, D.; Garibaldi, A.; Gullino, M.L. Jasmonic acid, abscisic acid, and salicylic acid are involved in the phytoalexin responses of rice to Fusarium fujikuroi, a high gibberellin producer pathogen. J. Agric. Food Chem. 2015, 63, 8134–8142. [Google Scholar] [CrossRef] [PubMed]
- Tang, Q.Y.; Zhang, C.X. Data Processing System (DPS) software with experimental design, statistical analysis and data mining developed for use in entomological research. Insect Sci. 2013, 20, 254–260. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer | Sequence (5′– 3′) | Reference |
|---|---|---|
| FgCWM1-Up-F | GCGGGCCCTACTCAGGGTAACGGAAAGG | This study |
| FgCWM1-Up-R | GCGAGCTCACAATGCAGCTCAATGTCG | |
| FgCWM1-Down-F | GGAAGCTTCAACCCAGACCTACCCA | This study |
| FgCWM1-Down-R | GGACTAGTGAAGAGCAGCGAACCAG | |
| P5 | TGATAATAATGTCCTCGTTCC | This study |
| P6 | TGACGAACTGTAAGTCGGATA | |
| P7 | ACCGAACTTCAAGACACCA | This study |
| P8 | CAACGGCCTCAACCTACT | |
| R-FgCWM1-F | AACCATGGATGAAGTTCTCCGCTGC | This study |
| R-FgCWM1-R | GGCCATGGAAGCATCCTTCAGAAGAGGT | |
| SS-FgCWM1-F | GCGAGCTCGCATGAAGTTCTCCGCTGC | This study |
| SS-FgCWM1-R | GGGGATCCAAGCATCCTTCAGAAGAGGT | |
| Fg-GAPDH-F | TGACTTGACTGTTCGCCTCGAGAA | [22] |
| Fg-GAPDH-R | ATGGAGGAGTTGGTGTTGCCGTTA | |
| Fg-β-tubulin-F | GTTGATCTCCAAGATCCGTG | [22] |
| Fg-β-tubulin-R | CATGCAAATGTCGTAGAGGG | |
| Fg-Factor1-F | CCTCCAGGATGTCTACAAGA | [22] |
| Fg-Factor1-R | CTCAACGGACTTGACTTCAG | |
| RJ-FgCWM1-F | GCTGGTGCCGAGGCTGTT | This study |
| RJ-FgCWM1-R | GGTCATCGCAGGTGTTTCCA | |
| Aox-F | GACTTGTCATGGTAGATGCCTG | [22] |
| Aox-R | CAGGACGAGCATAACCATTCTC | |
| w-GAPDH-F | AACTGTTCATGCCATCACTGCCAC | [22] |
| w-GAPDH-R | AGGACATACCAGTGAGCTTGCCAT | |
| hn-RNP-Q-F | TCACCTTCGCCAAGCTCAGAACTA | [22] |
| hn-RNP-Q-R | AGTTGAACTTGCCCGAAACATGCC |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, Y.-Z.; Chen, Q.; Liu, C.-H.; Lei, L.; Li, Y.; Zhao, K.; Wei, M.-Q.; Guo, Z.-R.; Wang, Y.; Xu, B.-J.; et al. Fusarium graminearum FgCWM1 Encodes a Cell Wall Mannoprotein Conferring Sensitivity to Salicylic Acid and Virulence to Wheat. Toxins 2019, 11, 628. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins11110628
Zhang Y-Z, Chen Q, Liu C-H, Lei L, Li Y, Zhao K, Wei M-Q, Guo Z-R, Wang Y, Xu B-J, et al. Fusarium graminearum FgCWM1 Encodes a Cell Wall Mannoprotein Conferring Sensitivity to Salicylic Acid and Virulence to Wheat. Toxins. 2019; 11(11):628. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins11110628
Chicago/Turabian StyleZhang, Ya-Zhou, Qing Chen, Cai-Hong Liu, Lu Lei, Yang Li, Kan Zhao, Mei-Qiao Wei, Zhen-Ru Guo, Yan Wang, Bin-Jie Xu, and et al. 2019. "Fusarium graminearum FgCWM1 Encodes a Cell Wall Mannoprotein Conferring Sensitivity to Salicylic Acid and Virulence to Wheat" Toxins 11, no. 11: 628. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins11110628