A Simple LC–MS Method for the Quantitation of Alkaloids in Endophyte-Infected Perennial Ryegrass

 , ,
, ,
Abstract
:
1. Introduction
2. Results
2.1. Method Performance
2.2. LOD, LOQ and Linearity
2.3. Accuracy (Bias), Precision and Selectivity
2.4. Extraction of Alkaloids
2.5. Matrix Effect and Recovery
2.6. Method Robustness and Application
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Chemicals
5.2. Plant Material
5.3. Sample Preparation
5.4. Sample Extraction
5.5. Method Validation
5.6. Analytical Instruments and Conditions
5.7. Method Application
5.8. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Jones, M.; Lazenby, A. The Grass Crop: The Physiological Basis of Production; Chapman and Hall: London, UK, 1988. [Google Scholar]
- Moate, P.; Williams, S.; Grainger, C.; Hannah, M.; Mapleson, D.; Auldist, M.; Greenwood, J.; Popay, A.; Hume, D.; Mace, W. Effects of wild-type, AR1 and AR37 endophyte-infected perennial ryegrass on dairy production in Victoria, Australia. Anim. Prod. Sci. 2012, 52, 1117–1130. [Google Scholar] [CrossRef]
- Liu, J.; Nagabhyru, P.; Schardl, C.L. Epichloë festucae endophytic growth in florets, seeds, and seedlings of perennial ryegrass (Lolium perenne). Mycologia 2017, 109, 691–700. [Google Scholar] [CrossRef] [PubMed]
- Gagic, M.; Faville, M.J.; Zhang, W.; Forester, N.T.; Rolston, M.P.; Johnson, R.D.; Ganesh, S.; Koolaard, J.P.; Easton, H.; Hudson, D. Seed transmission of Epichloë endophytes in Lolium perenne is heavily influenced by host genetics. Front. Plant Sci. 2018, 9, 1580. [Google Scholar] [CrossRef] [PubMed]
- Schardl, C.L.; Leuchtmann, A.; Spiering, M.J. Symbioses of grasses with seedborne fungal endophytes. Annu. Rev. Plant Biol. 2004, 55, 315–340. [Google Scholar] [CrossRef]
- Scott, B.; Schardl, C. Fungal symbionts of grasses: Evolutionary insights and agricultural potential. Trends Microbiol. 1993, 1, 196–200. [Google Scholar] [CrossRef]
- Hume, D.; Sewell, J. Agronomic advantages conferred by endophyte infection of perennial ryegrass (Lolium perenne L.) and tall fescue (Festuca arundinacea Schreb.) in Australia. Crop Pasture Sci. 2014, 65, 747–757. [Google Scholar] [CrossRef]
- Malinowski, D.P.; Belesky, D.P. Adaptations of endophyte-infected cool-season grasses to environmental stresses: Mechanisms of drought and mineral stress tolerance. Crop Sci. 2000, 40, 923–940. [Google Scholar] [CrossRef]
- Clay, K.; Schardl, C. Evolutionary origins and ecological consequences of endophyte symbiosis with grasses. Am. Nat. 2002, 160, S99–S127. [Google Scholar] [CrossRef]
- Zain, M.E. Impact of mycotoxins on humans and animals. J. Saudi Chem. Soc. 2011, 15, 129–144. [Google Scholar] [CrossRef] [Green Version]
- Wilson, D. Fungal endophytes: Out of sight but should not be out of mind. Oikos 1993, 68, 379–384. [Google Scholar] [CrossRef]
- Smith, B.; McLeay, L.; Embling, P. Effect of the mycotoxins penitrem, paxilline and lolitrem B on the electromyographic activity of skeletal and gastrointestinal smooth muscle of sheep. Res. Vet. Sci. 1997, 62, 111–116. [Google Scholar] [CrossRef]
- Paterson, J.; Forcherio, C.; Larson, B.; Samford, M.; Kerley, M. The effects of fescue toxicosis on beef cattle productivity. J. Anim. Sci. 1995, 73, 889–898. [Google Scholar] [CrossRef] [PubMed]
- Porter, J. Analysis of endophyte toxins: Fescue and other grasses toxic to livestock. J. Anim. Sci. 1995, 73, 871–880. [Google Scholar] [CrossRef] [PubMed]
- Gallagher, R.; White, E.; Mortimer, P. Ryegrass staggers: Isolation of potent neurotoxins lolitrem A and lolitrem B from staggers-producing pastures. N. Z. Vet. J. 1981, 29, 189–190. [Google Scholar] [CrossRef] [PubMed]
- Craig, A.M.; Blythe, L.L.; Duringer, J.M. The role of the Oregon State University endophyte service laboratory in diagnosing clinical cases of endophyte toxicoses. J. Agric. Food Chem. 2014, 62, 7376–7381. [Google Scholar] [CrossRef] [PubMed]
- Blythe, L.; Estill, C.; Males, J.; Craig, A. Determination of the toxic threshold of lolitrem B in cattle eating endophyte-infected perennial ryegrass. In Proceedings of the 6th International Symposium on Fungal Endophytes of Grasses; New Zealand Grassland Association: Christchurch, New Zealand, 2007; pp. 399–402. [Google Scholar]
- Fisher, M.J.; Bohnert, D.; Ackerman, C.; Schauer, C.; DelCurto, T.; Craig, A.; Vanzant, E.; Harmon, D.; Schrick, F. Evaluation of perennial ryegrass straw as a forage source for ruminants. J. Anim. Sci. 2004, 82, 2175–2184. [Google Scholar] [CrossRef] [PubMed]
- Bush, L.P.; Wilkinson, H.H.; Schardl, C.L. Bioprotective alkaloids of grass-fungal endophyte symbioses. Plant Physiol. 1997, 114, 1. [Google Scholar] [CrossRef]
- Rowan, D.D.; Hunt, M.B.; Gaynor, D.L. Peramine, a novel insect feeding deterrent from ryegrass infected with the endophyte Acremonium loliae. J. Chem. Soc. Chem. Commun. 1986, 12, 935–936. [Google Scholar] [CrossRef]
- Siegel, M.; Latch, G.; Bush, L.; Fannin, F.; Rowan, D.; Tapper, B.; Bacon, C.; Johnson, M. Fungal endophyte-infected grasses: Alkaloid accumulation and aphid response. J. Chem. Ecol. 1990, 16, 3301–3315. [Google Scholar] [CrossRef]
- Rowan, D.D. Lolitrems, peramine and paxilline: Mycotoxins of the ryegrass/endophyte interaction. Agric. Ecosyst. Environ. 1993, 44, 103–122. [Google Scholar] [CrossRef]
- Gallagher, R.T.; Hawkes, A.D. High-performance liquid chromatography with stop-flow ultraviolet spectral chracterization of lolitrem neurotoxins from perennial ryegrass. J. Chromatogr. A 1985, 322, 159–167. [Google Scholar] [CrossRef]
- Gallagher, R.T.; Hawkes, A.D.; Stewart, J.M. Rapid determination of the neurotoxin lolitrem B in perennial ryegrass by high-performance liquid chromatography with fluorescence detection. J. Chromatogr. A 1985, 321, 217–226. [Google Scholar] [CrossRef]
- Hovermale, J.T.; Craig, A.M. Correlation of ergovaline and lolitrem B levels in endophyte-infected perennial ryegrass (Lolium perenne). J. Vet. Diagn. Investig. 2001, 13, 323–327. [Google Scholar] [CrossRef] [PubMed]
- Rottinghaus, G.E.; Garner, G.B.; Cornell, C.N.; Ellis, J.L. HPLC method for quantitating ergovaline in endophyte-infested tall fescue: Seasonal variation of ergovaline levels in stems with leaf sheaths, leaf blades, and seed heads. J. Agric. Food Chem. 1991, 39, 112–115. [Google Scholar] [CrossRef]
- Craig, A.M.; Bilich, D.; Hovermale, J.T.; Welty, R.E. Improved extraction and HPLC methods for ergovaline from plant material and rumen fluid. J. Vet. Diagn. Investig. 1994, 6, 348–352. [Google Scholar] [CrossRef] [PubMed]
- Hill, N.S.; Rottinghaus, G.E.; Agee, C.S.; Schultz, L.M. Simplified sample preparation for HPLC analysis of ergovaline in tall fescue. Crop Sci. 1993, 33, 331–333. [Google Scholar] [CrossRef]
- Garner, G.B.; Rottinghaus, G.E.; Cornell, C.N.; Testereci, H. Chemistry of compounds associated with endophyte/grass interaction: Ergovaline-and ergopeptine-related alkaloids. Agric. Ecosyst. Environ. 1993, 44, 65–80. [Google Scholar] [CrossRef]
- Spiering, M.J.; Davies, E.; Tapper, B.A.; Schmid, J.; Lane, G.A. Simplified extraction of ergovaline and peramine for analysis of tissue distribution in endophyte-infected grass tillers. J. Agric. Food Chem. 2002, 50, 5856–5862. [Google Scholar] [CrossRef]
- Walker, K.; Duringer, J.; Craig, A.M. Determination of the ergot alkaloid ergovaline in tall fescue seed and straw using a QuEChERS extraction method with High-Performance Liquid Chromatography–Fluorescence Detection. J. Agric. Food Chem. 2015, 63, 4236–4242. [Google Scholar] [CrossRef]
- Baldauf, M.W.; Mace, W.J.; Richmond, D.S. Endophyte-mediated resistance to black cutworm as a function of plant cultivar and endophyte strain in tall fescue. Environ. Entomol. 2011, 40, 639–647. [Google Scholar] [CrossRef]
- Blankenship, J.D.; Spiering, M.J.; Wilkinson, H.H.; Fannin, F.F.; Bush, L.P.; Schardl, C.L. Production of loline alkaloids by the grass endophyte, Neotyphodium uncinatum, in defined media. Phytochemistry 2001, 58, 395–401. [Google Scholar] [CrossRef]
- Schardl, C.L.; Grossman, R.B.; Nagabhyru, P.; Faulkner, J.R.; Mallik, U.P. Loline alkaloids: Currencies of mutualism. Phytochemistry 2007, 68, 980–996. [Google Scholar] [CrossRef] [PubMed]
- Moore, J.R.; Pratley, J.E.; Mace, W.J.; Weston, L.A. Variation in alkaloid production from genetically diverse Lolium accessions infected with Epichloë species. J. Agric. Food Chem. 2015, 63, 10355–10365. [Google Scholar] [CrossRef] [PubMed]
- Rasmussen, S.; Lane, G.A.; Mace, W.; Parsons, A.J.; Fraser, K.; Xue, H. The use of genomics and metabolomics methods to quantify fungal endosymbionts and alkaloids in grasses. Methods Mol. Biol. 2011, 860, 213–226. [Google Scholar]
- Soto-Barajas, M.C.; Zabalgogeazcoa, I.; González-Martin, I.; Vázquez-de-Aldana, B.R. Qualitative and quantitative analysis of endophyte alkaloids in perennial ryegrass using near-infrared spectroscopy. J. Sci. Food Agric. 2017, 97, 5028–5036. [Google Scholar] [CrossRef]
- Laganà, A. Introduction to the toxins special issue on LC-MS/MS methods for mycotoxin analysis. Toxins 2017, 9, 325. [Google Scholar] [CrossRef]
- Reddy, P.; Deseo, M.; Ezernieks, V.; Guthridge, K.; Spangenberg, G.; Rochfort, S. Toxic indole diterpenes from endophyte-infected perennial ryegrass Lolium perenne L.: Isolation and stability. Toxins 2019, 11, 16. [Google Scholar] [CrossRef]
- Young, C.A.; Tapper, B.A.; May, K.; Moon, C.D.; Schardl, C.L.; Scott, B. Indole-diterpene biosynthetic capability of Epichloë endophytes as predicted by ltm gene analysis. Appl. Environ. Microbiol. 2009, 75, 2200–2211. [Google Scholar] [CrossRef]
- Salminen, S.O.; Grewal, P.S.; Quigley, M.F. Does mowing height influence alkaloid production in endophytic tall fescue and perennial ryegrass? J. Chem. Ecol. 2003, 29, 1319–1328. [Google Scholar] [CrossRef]
- Salminen, S.O.; Richmond, D.S.; Grewal, S.K.; Grewal, P.S. Influence of temperature on alkaloid levels and fall armyworm performance in endophytic tall fescue and perennial ryegrass. Entomol. Exp. Appl. 2005, 115, 417–426. [Google Scholar] [CrossRef]
- Hettiarachchige, I.K.; Elkins, A.C.; Reddy, P.; Mann, R.C.; Guthridge, K.M.; Sawbridge, T.I.; Forster, J.W.; Spangenberg, G.C. Genetic modification of asexual Epichloë endophytes with the perA gene for peramine biosynthesis. Mol. Genet. Genom. 2018, 294, 315–328. [Google Scholar] [CrossRef] [PubMed]
- Fuchs, B.; Krischke, M.; Mueller, M.J.; Krauss, J. Peramine and lolitrem B from endophyte-grass associations cascade up the food chain. J. Chem. Ecol. 2013, 39, 1385–1389. [Google Scholar] [CrossRef] [PubMed]
- Fuchs, B.; Krischke, M.; Mueller, M.J.; Krauss, J. Herbivore-specific induction of defence metabolites in a grass–endophyte association. Funct. Ecol. 2017, 31, 318–324. [Google Scholar] [CrossRef]
- Peters, F.T.; Drummer, O.H.; Musshoff, F. Validation of new methods. Forensic Sci. Int. 2007, 165, 216–224. [Google Scholar] [CrossRef] [PubMed]
- Lea, K.; Smith, L.; Gaskill, C.; Coleman, R.; Smith, S. Ergovaline stability in tall fescue based on sample handling and storage methods. Front. Chem. 2014, 2, 76. [Google Scholar] [CrossRef]
- Hafner, M.; Sulyok, M.; Schuhmacher, R.; Crews, C.; Krska, R. Stability and epimerisation behaviour of ergot alkaloids in various solvents. World Mycotoxin J. 2008, 1, 67–78. [Google Scholar] [CrossRef]
- Smith, D.; Shappell, N. Epimerization of ergopeptine alkaloids in organic and aqueous solvents. J. Anim. Sci. 2002, 80, 1616–1622. [Google Scholar] [CrossRef]
- Fuchs, B.; Krischke, M.; Mueller, M.J.; Krauss, J. Plant age and seasonal timing determine endophyte growth and alkaloid biosynthesis. Fungal Ecol. 2017, 29, 52–58. [Google Scholar] [CrossRef]
- Shah, V.P.; Midha, K.K.; Dighe, S.; McGilveray, I.J.; Skelly, J.P.; Yacobi, A.; Layloff, T.; Viswanathan, C.; Cook, C.E.; McDowall, R. Analytical methods validation: Bioavailability, bioequivalence, and pharmacokinetic studies. Conference report. Pharm. Sci. 1992, 9, 588–592. [Google Scholar]
- Adhikari, K.B.; Boelt, B.; Fomsgaard, I.S. Identification and quantification of loline-type alkaloids in endophyte-infected grasses by LC-MS/MS. J. Agric. Food Chem. 2016, 64, 6212–6218. [Google Scholar] [CrossRef]
- Kaur, J.; Ekanayake, P.N.; Tian, P.; De Jong, E.V.Z.; Dobrowolski, M.P.; Rochfort, S.J.; Mann, R.C.; Smith, K.F.; Forster, J.W.; Guthridge, K.M. Discovery and characterisation of novel asexual Epichloë endophytes from perennial ryegrass (Lolium perenne L.). Crop Pasture Sci. 2015, 66, 1058–1070. [Google Scholar] [CrossRef]
- Ekanayake, P.N.; Hand, M.L.; Spangenberg, G.C.; Forster, J.W.; Guthridge, K.M. Genetic diversity and host specificity of fungal endophyte taxa in fescue pasture grasses. Crop Sci. 2012, 52, 2243–2252. [Google Scholar] [CrossRef]
- Eady, C.; Corkran, J.; Bailey, K.; Kerr, G.; Nicol, A. Estimation of ergovaline intake of cows from grazed perennial ryegrass containing NEA2 or standard endophyte. J. N. Z. Grassl. 2017, 79, 189–196. [Google Scholar]



{kind=link}
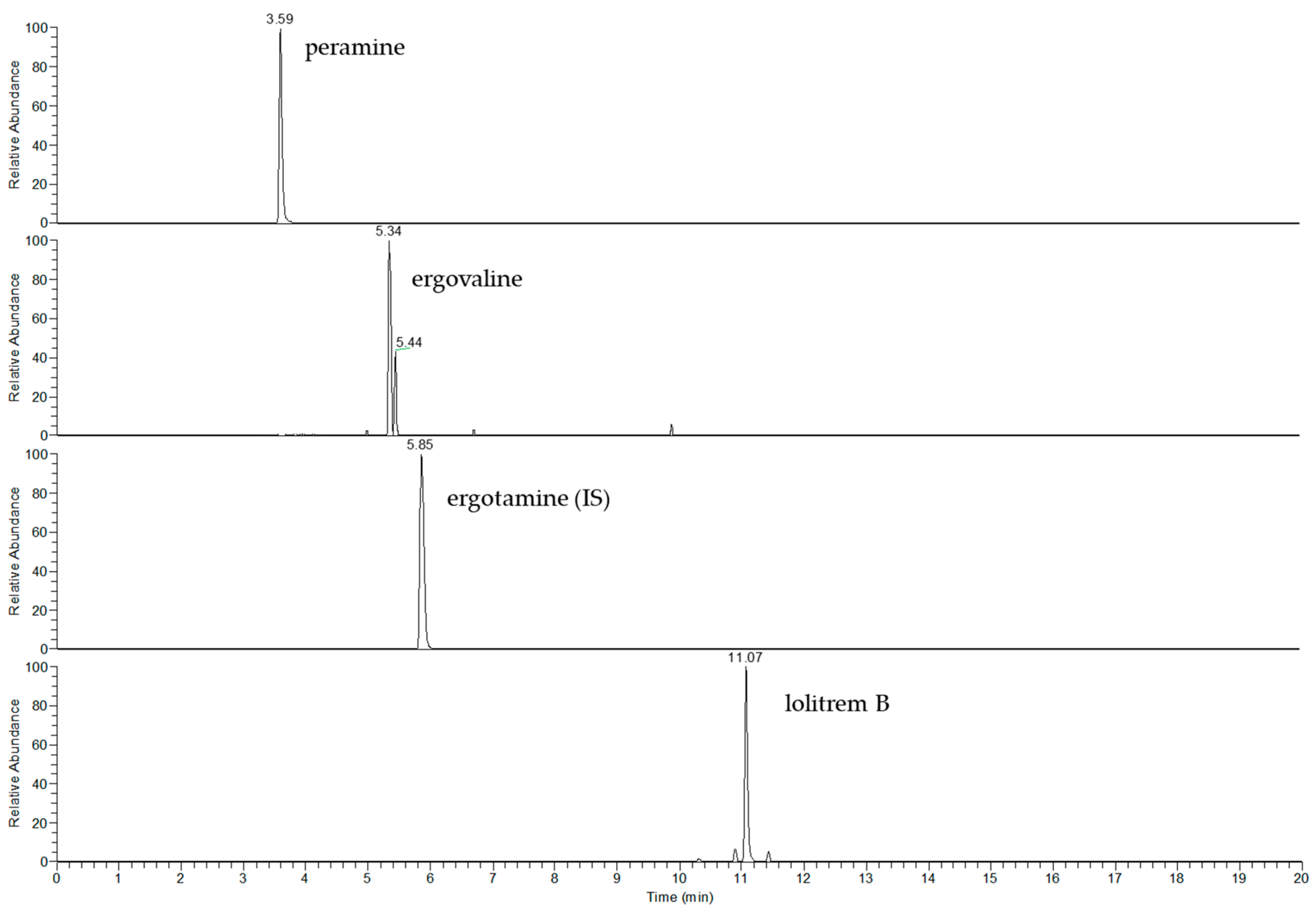{kind=link}
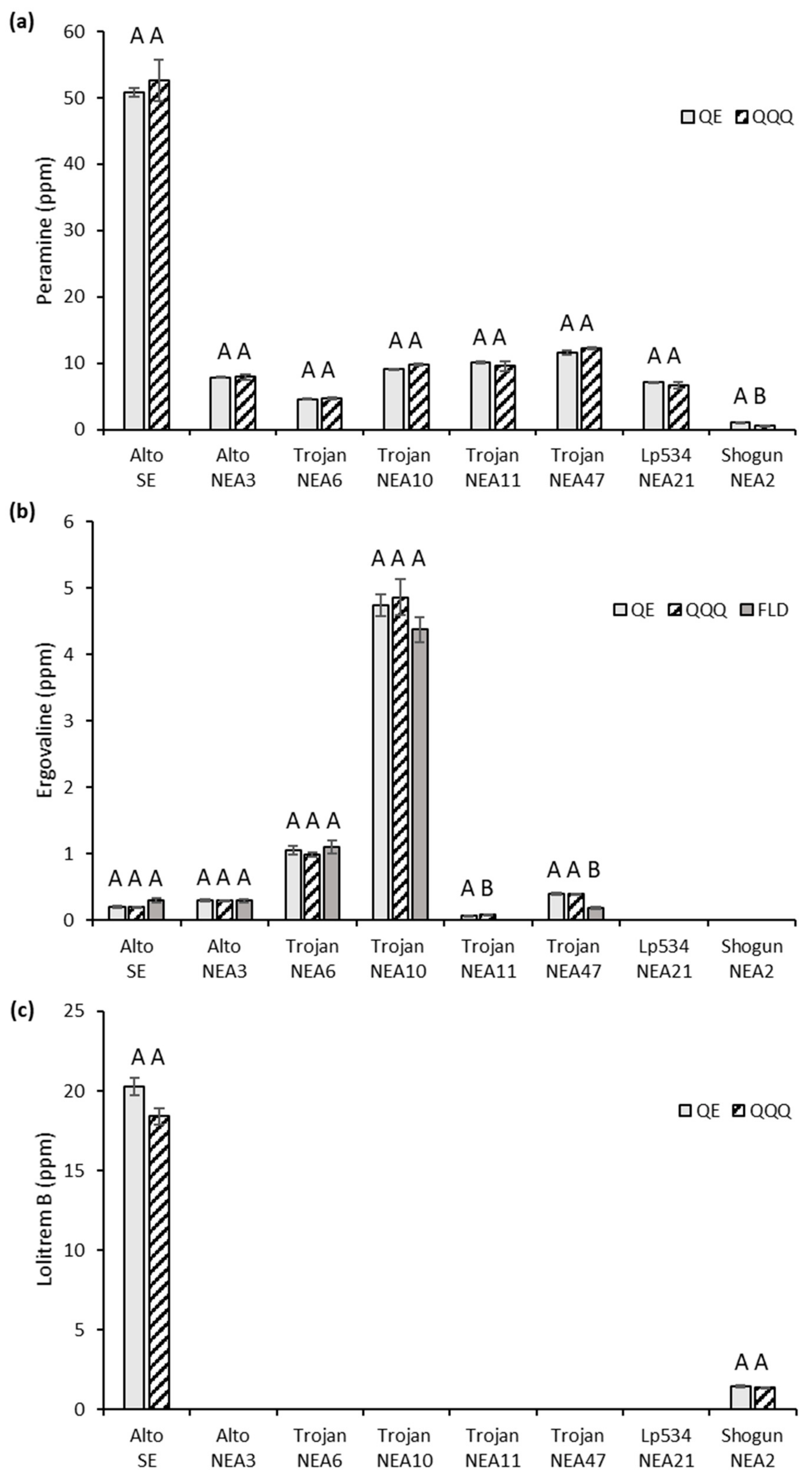{kind=link}
| Compound | RT | Ion Mass | LOD | LOQ | Standard Concentrations (ng/mL) | Equation | R2 | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| (min) | (m/z) | (ng/mL) | (ng/mL) | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | |||
| peramine | 3.59 | 248.1501 | 0.2 | 0.8 | 0.8 | 8.0 | 19.9 | 39.8 | 79.7 | 199.2 | 398.5 | 796.9 | 1593.9 | y = 217004x | 0.9912 |
| ergovaline/ergotamine 1 | 5.34/5.85 | 534.2709/582.2701 | 0.2 | 0.8 | 0.8 | 8.4 | 21.0 | 42.1 | 84.2 | 210.5 | 421.0 | 842.0 | 1684.0 | y = 54031x | 0.9998 |
| lolitrem B | 11.07 | 686.4037 | 1.8 | 6.0 | 6.0 | 12.0 | 30.0 | 60.0 | 120.0 | 300.0 | 600.0 | 1200.0 | 2400.0 | y = 12113x | 0.9970 |
| Standard | Actual Concentration (ng/mL) | Measured Concentration (ng/mL) | Accuracy (bias) (%) | Precision (% RSD) | |
|---|---|---|---|---|---|
| peramine | Low | 8.0 | 7.9 | 100.2 | 3.5 |
| Med | 79.7 | 79.2 | 99.7 | 3.4 | |
| High | 796.9 | 792.3 | 99.6 | 1.0 | |
| ergotamine | Low | 8.4 | 7.9 | 99.1 | 1.4 |
| Med | 84.2 | 83.2 | 99.4 | 2.4 | |
| High | 842.0 | 843.9 | 99.9 | 2.6 | |
| lolitrem B | Low | 12.0 | 11.9 | 99.3 | 2.3 |
| Med | 120.0 | 119.3 | 99.5 | 3.1 | |
| High | 1200.0 | 1194.8 | 100.0 | 2.8 |
| Peramine | Ergovaline | Lolitrem B | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Extract | Mean ± SD | RSD (%) | Σ | Mean Conc. (ng/mL) | RSD (%) | Σ | Mean Conc. (ng/mL) | RSD (%) | Σ | |
| Method 1 | 1 | 32.2 ± 0.1 | 0.4 | 39.5 | 0.1 ± 0.01 | 2.2 | 0.1 | 13.6 ± 0.2 | 1.2 | 15.3 |
| 2 | 7.3 ± 0.2 | 3.3 | - | - | - | - | 1.7 ± 0.1 * | 4.2 | - | |
| 3 | 2.8 ± 0.1 * | 5.0 | - | - | - | - | 0.2 ± 0.02 * | 7.5 | - | |
| 4 | 1.2 ± 0.1 * | 5.6 | - | - | - | - | - | - | - | |
| 5 | 0.6 ± 0.1 * | 8.3 | - | - | - | - | - | - | - | |
| Method 2 | 1 & 2 | 33.2 ± 0.1 | 0.4 | - | 0.1 ± 0.003 | 2.0 | - | 14.3 ± 0.8 | 5.4 | - |
| QC Sample | Spike 1 | Peramine | Ergotamine | Lolitrem B | |||
|---|---|---|---|---|---|---|---|
| RE | ME | RE | ME | RE | ME | ||
| E− | low | 77 | 66 | 85 | 76 | 87 | 84 |
| med | 79 | 70 | 87 | 77 | 86 | 80 | |
| high | 81 | 81 | 85 | 76 | 80 | 71 | |
| E+ | low | 88 | 61 | 85 | 74 | 84 | 88 |
| med | 85 | 67 | 87 | 74 | 90 | 78 | |
| high | 83 | 67 | 86 | 73 | 86 | 67 | |
| Cultivar/Variety | Endophyte Strain | Qualitative Alkaloid Profile | Supplier |
|---|---|---|---|
| Alto | SE | P, E, L | Barenbrug Agriseeds |
| Alto | NEA3 | P, E | Barenbrug Agriseeds |
| Trojan | WE 1 | nil | Barenbrug Agriseeds |
| Trojan | NEA6 | P, E | Barenbrug Agriseeds |
| Trojan | NEA10 | P, E | Barenbrug Agriseeds |
| Trojan | NEA11 | P, E | Barenbrug Agriseeds |
| Trojan | NEA47 | P, E | Barenbrug Agriseeds |
| LP534 | NEA21 | P, L | Barenbrug Agriseeds |
| Shogun 2 | NEA2 | P, E, L (low) | Barenbrug Agriseeds |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vassiliadis, S.; Elkins, A.C.; Reddy, P.; Guthridge, K.M.; Spangenberg, G.C.; Rochfort, S.J. A Simple LC–MS Method for the Quantitation of Alkaloids in Endophyte-Infected Perennial Ryegrass. Toxins 2019, 11, 649. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins11110649
Vassiliadis S, Elkins AC, Reddy P, Guthridge KM, Spangenberg GC, Rochfort SJ. A Simple LC–MS Method for the Quantitation of Alkaloids in Endophyte-Infected Perennial Ryegrass. Toxins. 2019; 11(11):649. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins11110649
Chicago/Turabian StyleVassiliadis, Simone, Aaron C. Elkins, Priyanka Reddy, Kathryn M. Guthridge, German C. Spangenberg, and Simone J. Rochfort. 2019. "A Simple LC–MS Method for the Quantitation of Alkaloids in Endophyte-Infected Perennial Ryegrass" Toxins 11, no. 11: 649. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins11110649