Fusarium Mycotoxins Disrupt the Barrier and Induce IL-6 Release in a Human Placental Epithelium Cell Line

 , , ,
, , ,
Abstract
:1. Introduction
2. Results
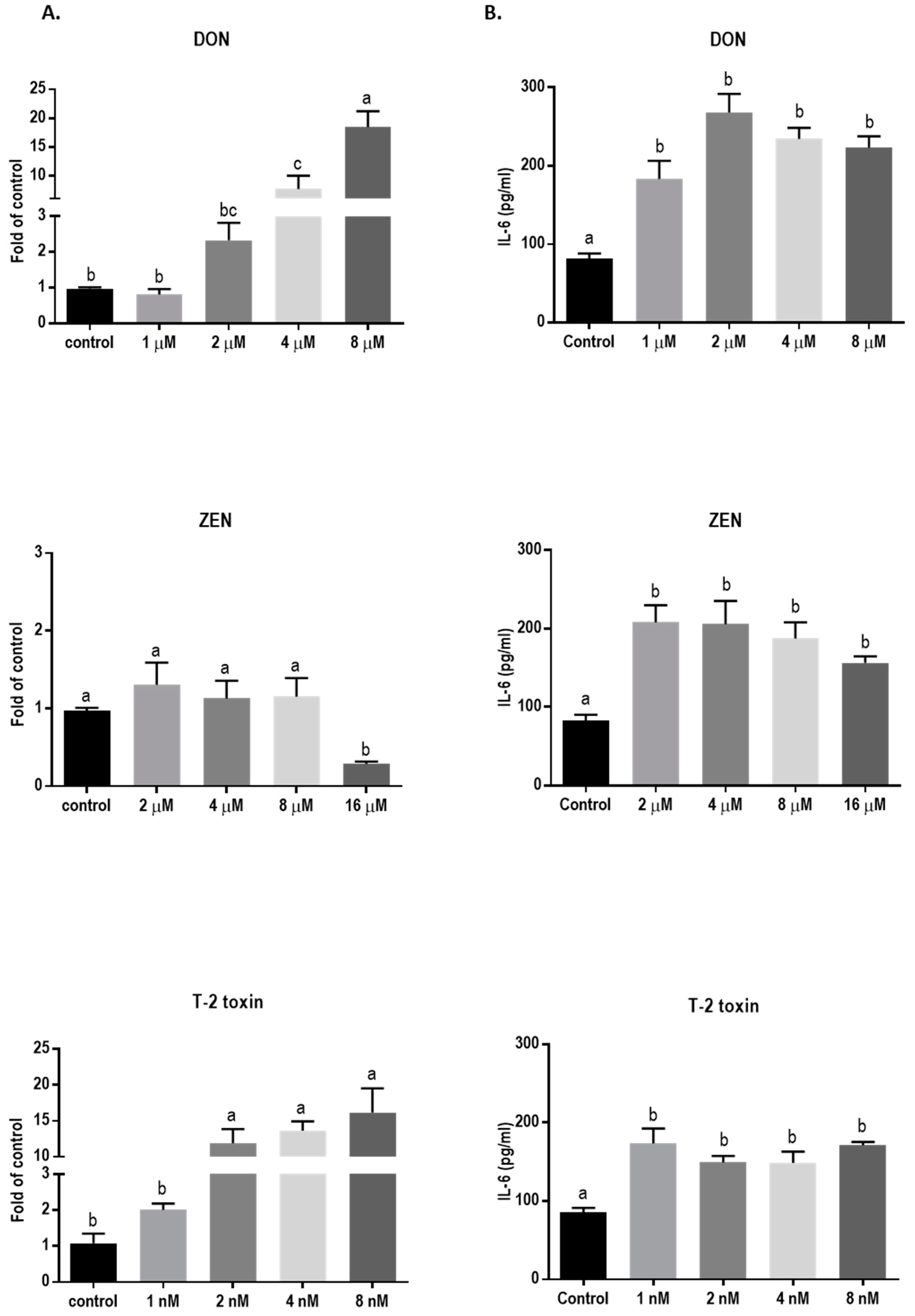
2.1. Concentrations of 8 µM DON, 16 µM ZEN, and 8 nM T-2 Toxin are Cytotoxic for BeWo Cells
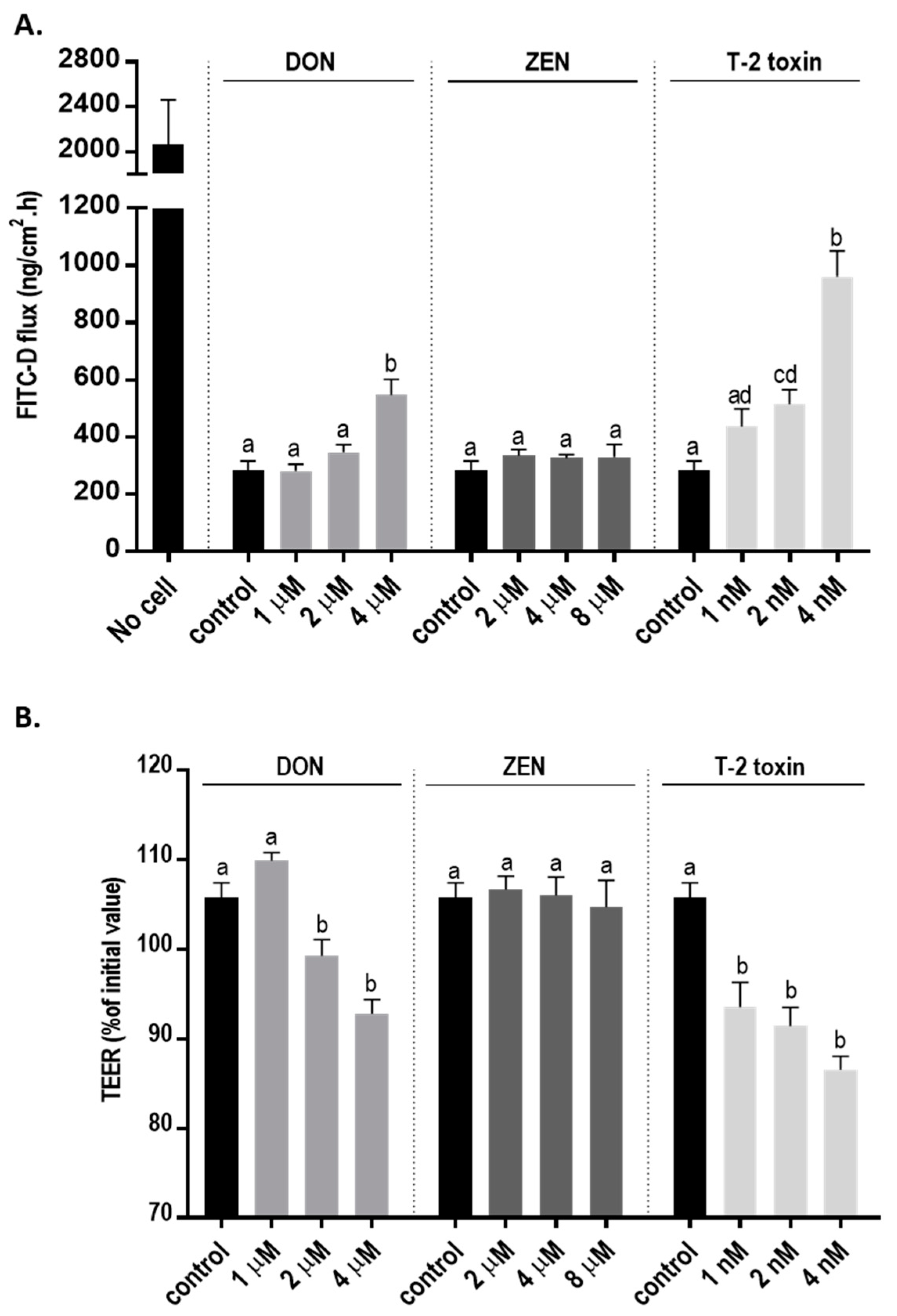
2.2. Mycotoxins Exposure Disrupts the Integrity and Increases Permeability of the BeWo Cell Layer
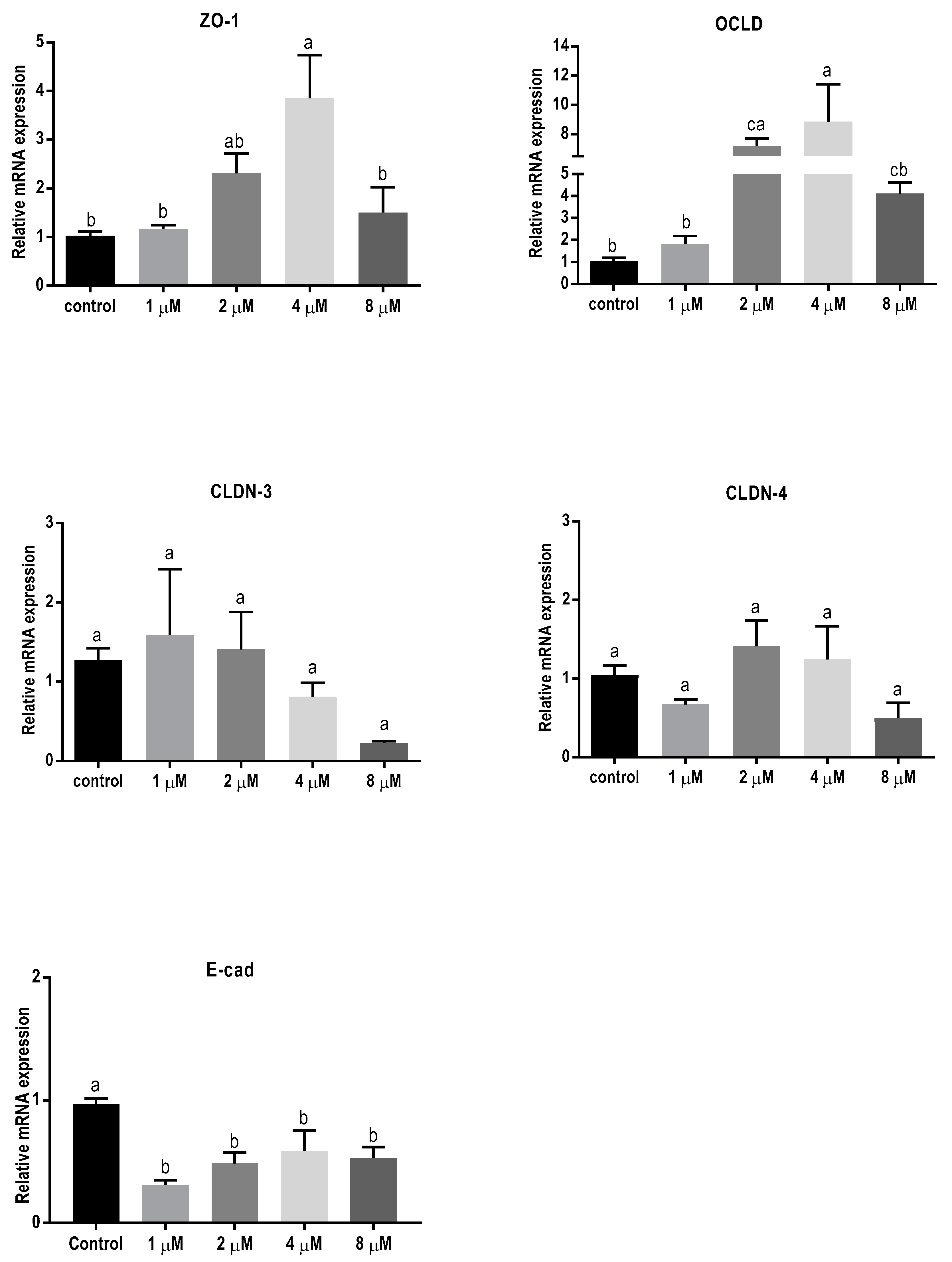
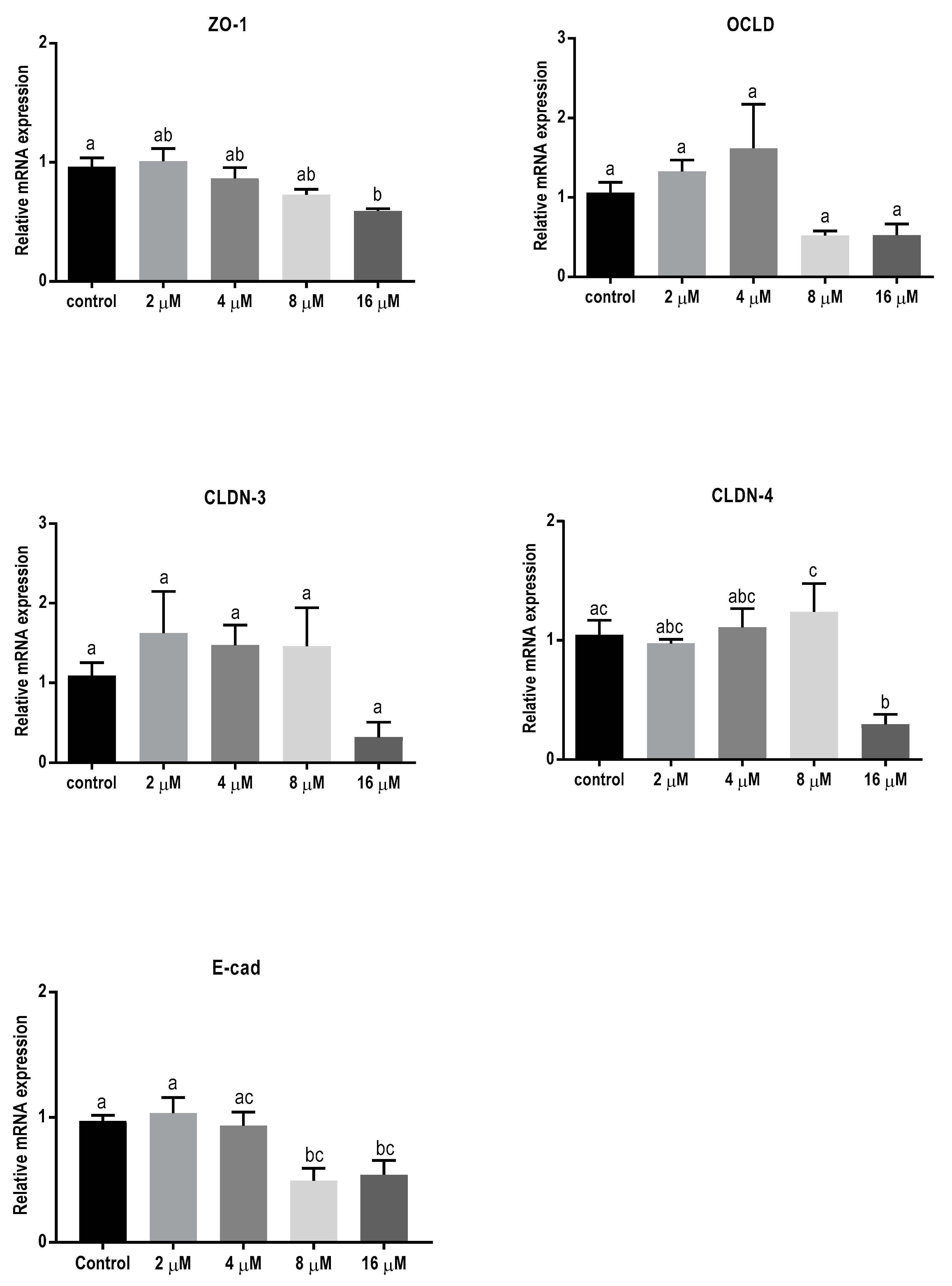
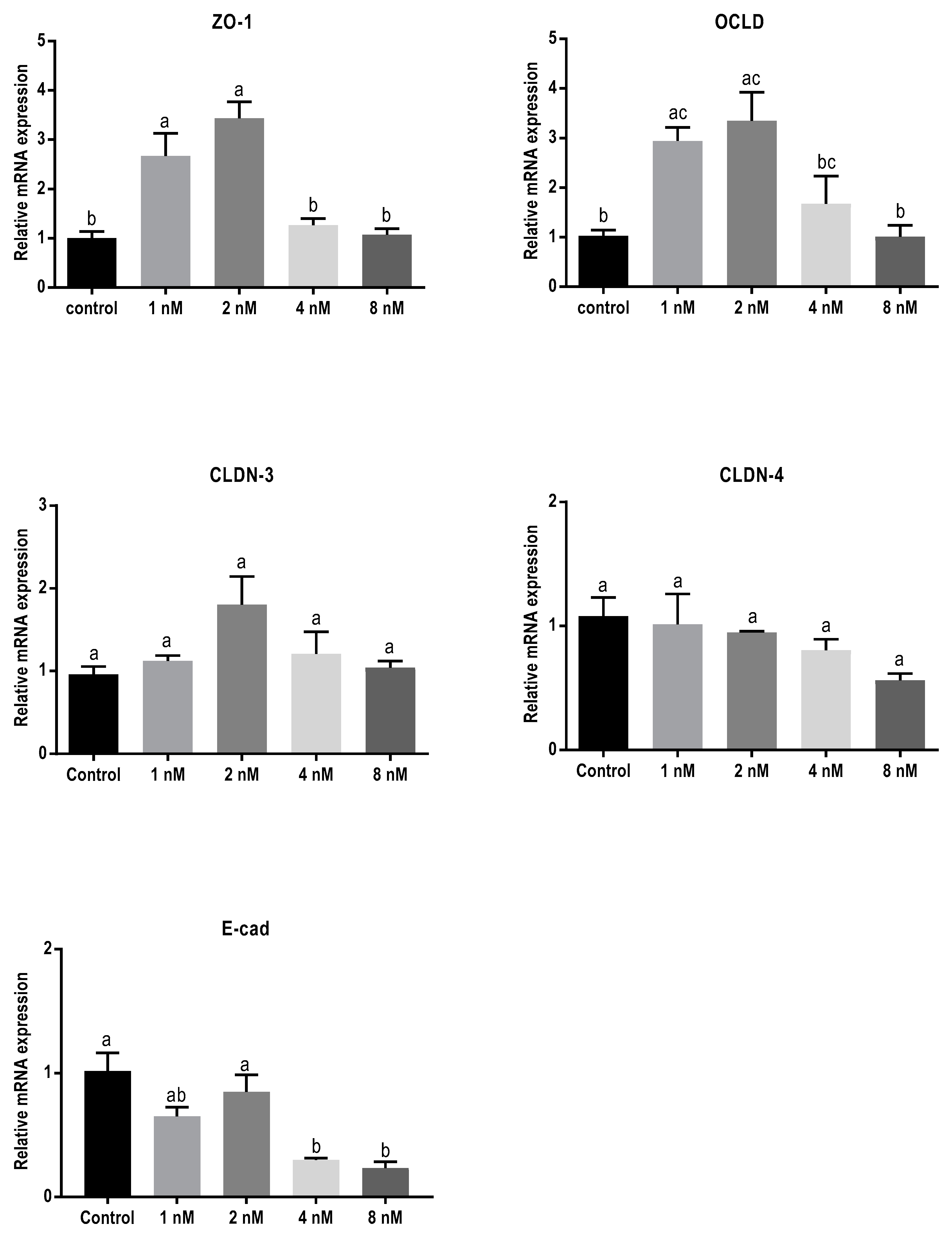
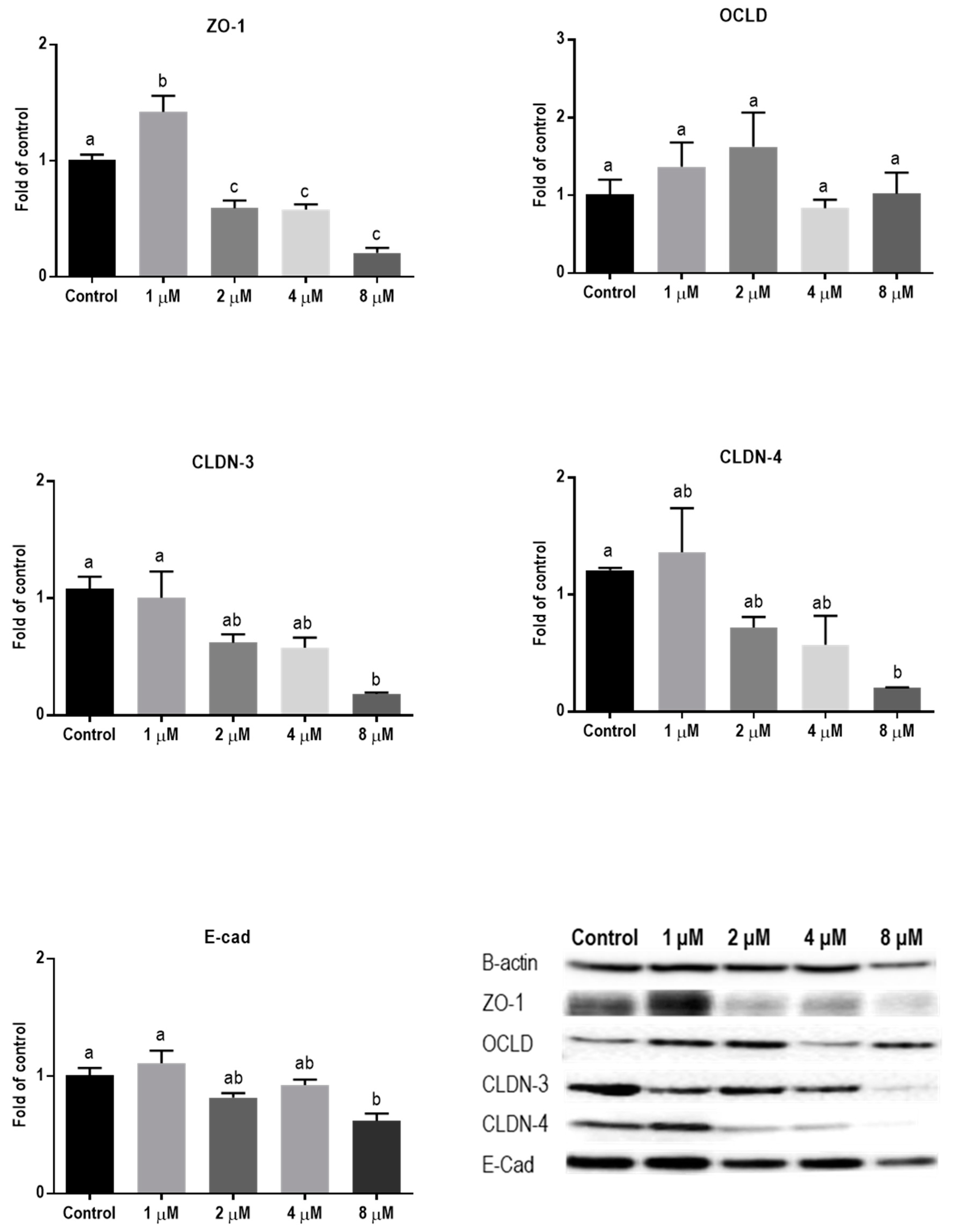
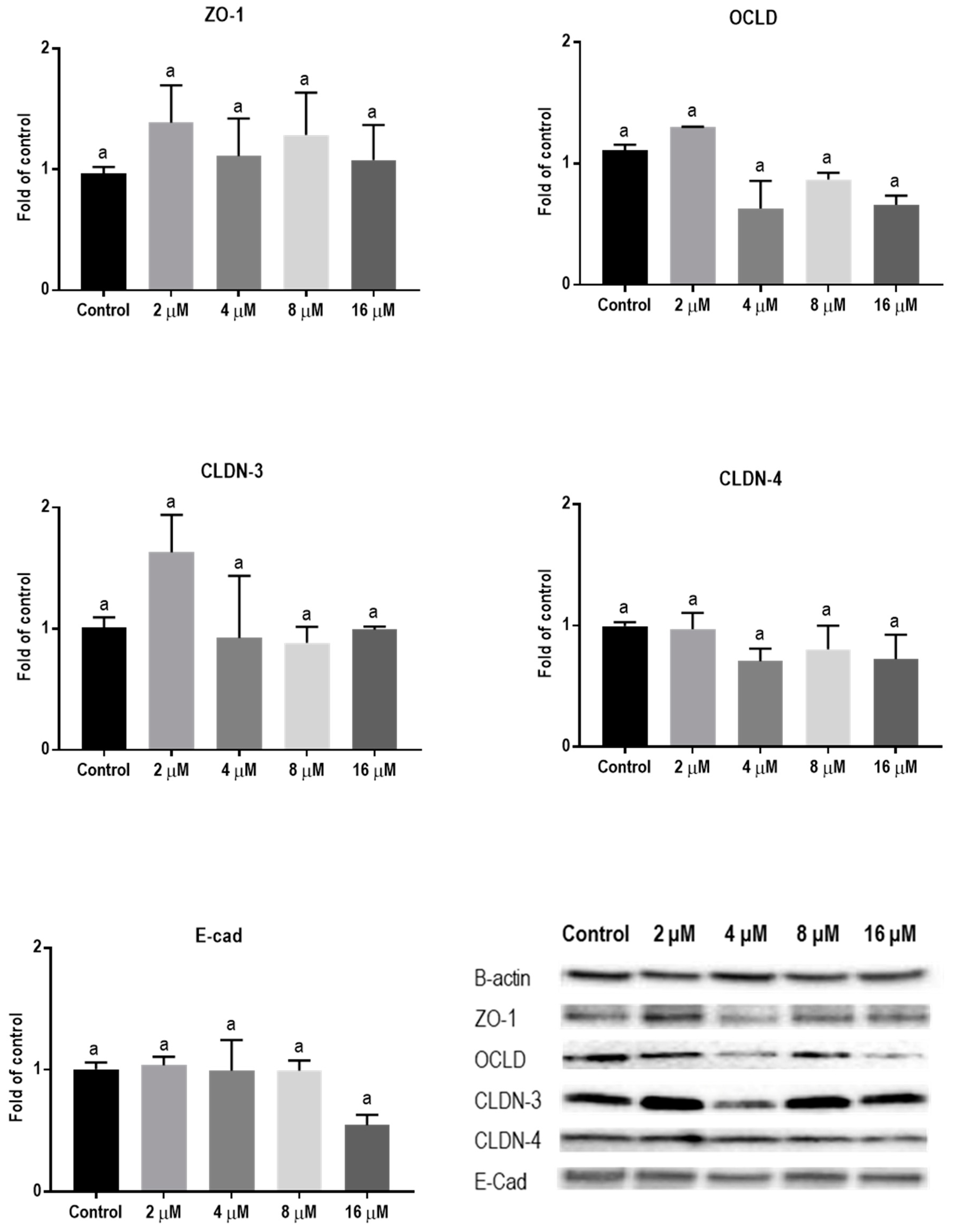
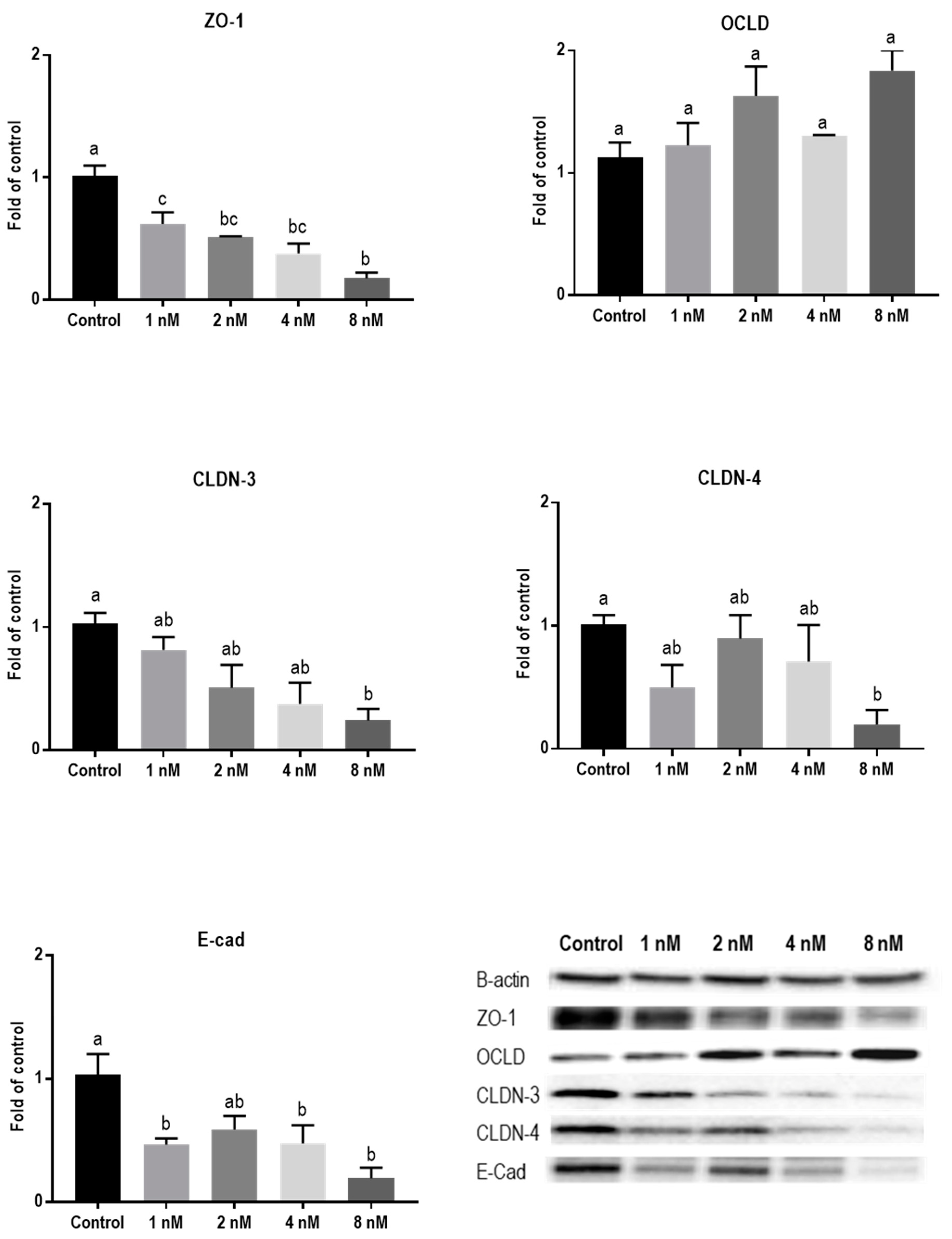
2.3. Mycotoxins Exposure Alters Gene Expression of Junctional Proteins
2.4. Mycotoxins Exposure Modulates the Protein Expression of Junctional Proteins
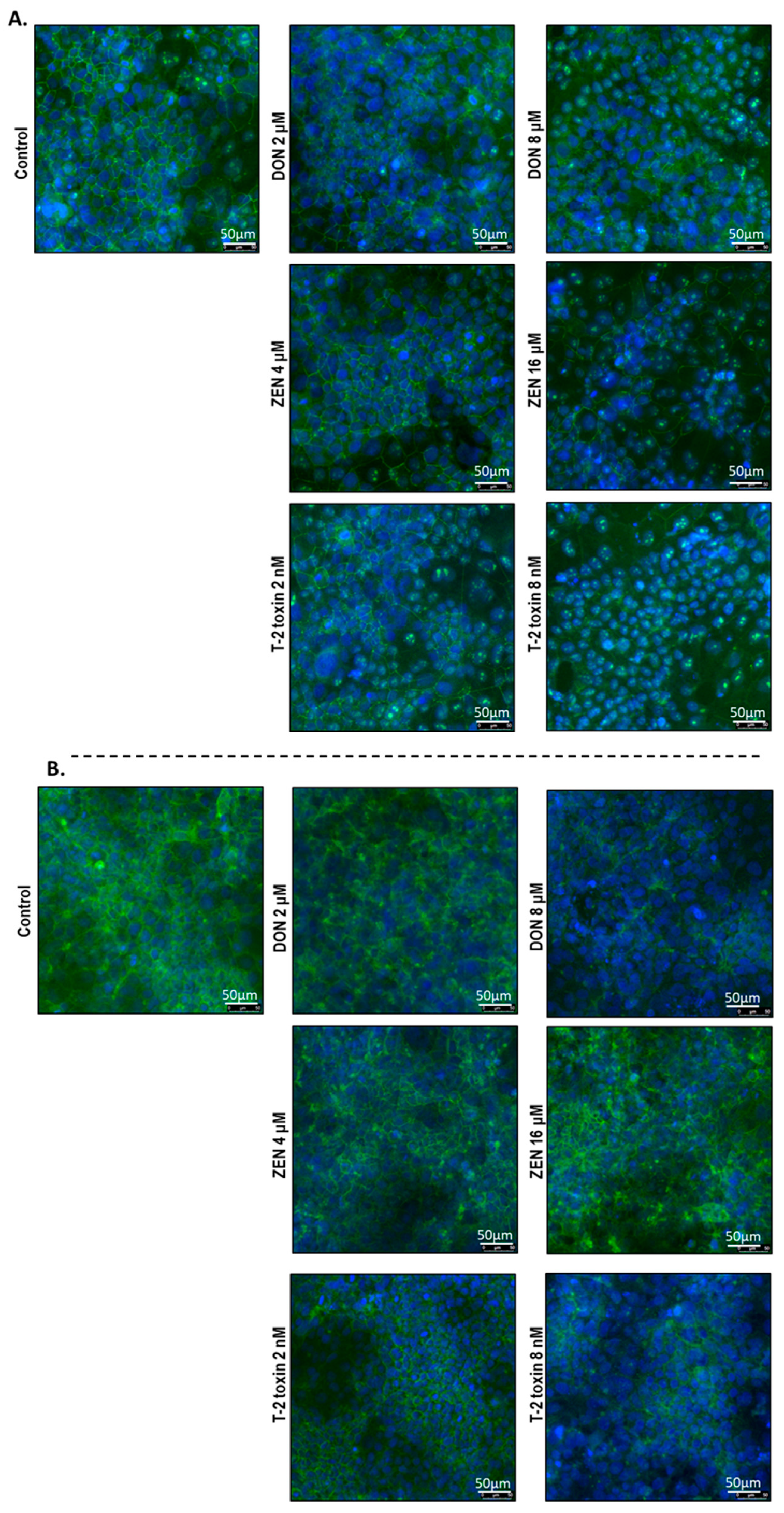
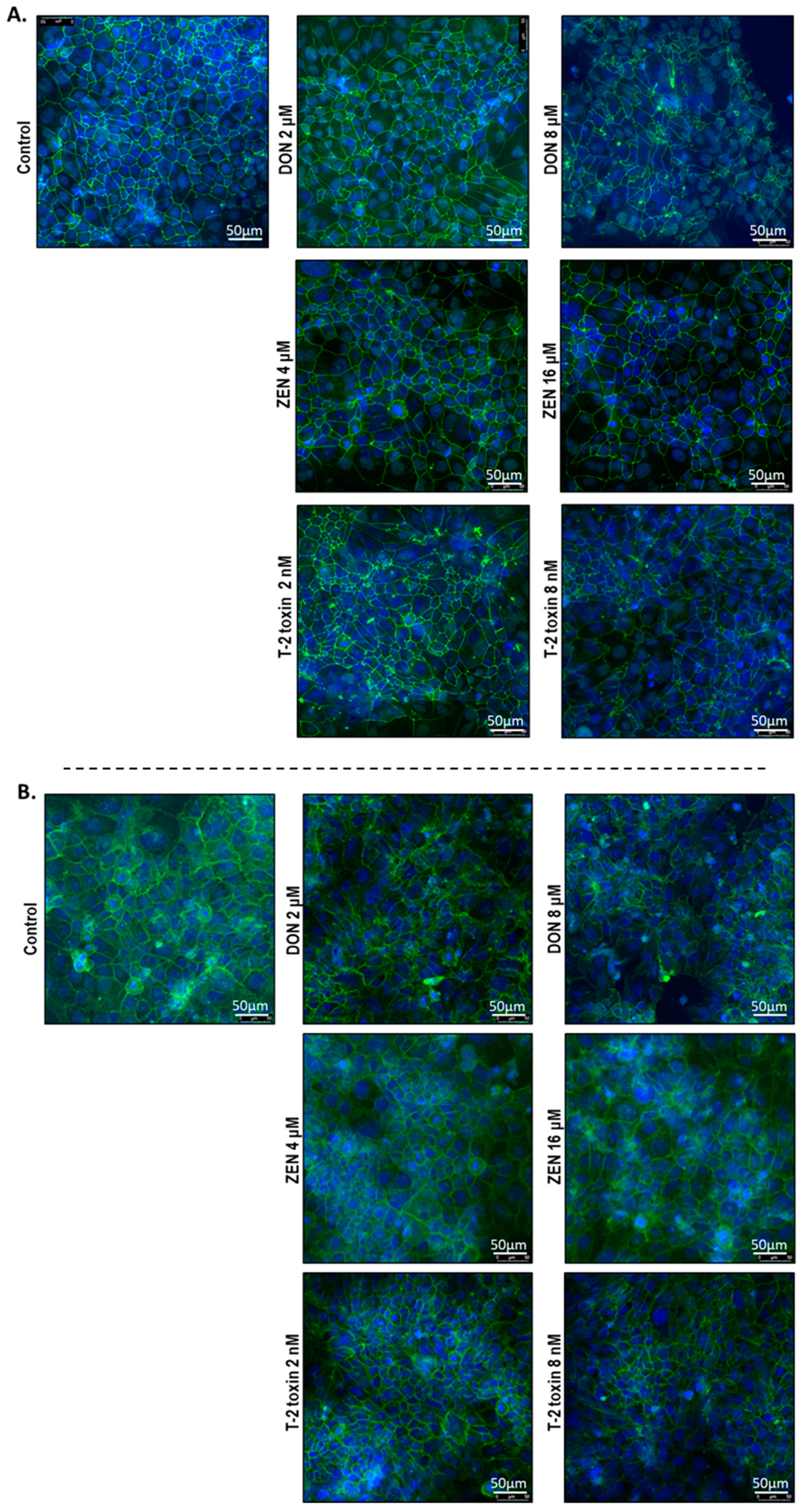
2.5. Mycotoxins Exposure Alters the Localization of Junctional Proteins
2.6. Mycotoxins Exposure Increases Interleukin-6 (IL-6) mRNA Expression and Protein Release
3. Discussion
4. Materials and Methods
4.1. Mycotoxins
4.2. Cell Culture
4.3. Cell Viability Assay
4.4. TEER Measurement and Paracellular Tracer Flux Assay
4.5. RNA Extraction and Quantitative qPCR
4.6. Western Blot Analysis
4.7. Immunofluorescence Staining
4.8. Quantitative Determination of Interleukin-6 (IL-6), IL-8, and IL-1β
4.9. Statistical Analysis
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Reddy, K.R.N.; Salleh, B.; Saad, B.; Abbas, H.K.; Abel, C.A.; Shier, W.T. An overview of mycotoxin contamination in foods and its implications for human health. Toxin Rev. 2010, 29, 3–26. [Google Scholar] [CrossRef]
- Logrieco, A.; Mule, G.; Moretti, A.; Bottalico, A. Toxigenic Fusarium Species and Mycotoxins Associated with Maize Ear Rot in Europe. In Mycotoxins in Plant Disease; Springer: Berlin/Heidelberg, Germany, 2002; pp. 597–609. [Google Scholar]
- Murphy, P.A.; Hendrich, S.; Landgren, C.; Bryant, C.M. Food Mycotoxins: An Update. J. Food Sci. 2006, 71, R51–R65. [Google Scholar] [CrossRef]
- Stoev, S.D. Foodborne mycotoxicoses, risk assessment and underestimated hazard of masked mycotoxins and joint mycotoxin effects or interaction. Environ. Toxicol. Pharmacol. 2015, 39, 794–809. [Google Scholar] [CrossRef] [PubMed]
- Broekaert, N.; Devreese, M.; De Baere, S.; De Backer, P.; Croubels, S. Modified Fusarium mycotoxins unmasked: From occurrence in cereals to animal and human excretion. Food Chem. Toxicol. 2015, 80, 17–31. [Google Scholar] [CrossRef] [PubMed]
- Raiola, A.; Tenore, G.C.; Manyes, L.; Meca, G.; Ritieni, A. Risk analysis of main mycotoxins occurring in food for children: An overview. Food Chem. Toxicol. 2015, 84, 169–180. [Google Scholar] [CrossRef] [PubMed]
- Nielsen, J.K.; Vikstrom, A.C.; Turner, P.; Knudsen, L.E. Deoxynivalenol transport across the human placental barrier. Food Chem. Toxicol 2011, 49, 2046–2052. [Google Scholar] [CrossRef] [PubMed]
- Bernhoft, A.; Behrens, G.H.G.; Ingebrigtsen, K.; Langseth, W.; Berndt, S.; Haugen, T.B.; Grotmol, T. Placental transfer of the estrogenic mycotoxin zearalenone in rats. Reprod. Toxicol. 2001, 15, 545–550. [Google Scholar] [CrossRef]
- Lafarge-Frayssinet, C.; Chakor, K.; Lafont, P.; Frayssinet, C. Transplacental transfer of T2-toxin: Pathological effect. J. Environ. Pathol. Toxicol. Oncol. 1990, 10, 64–68. [Google Scholar]
- Dänicke, S.; Brüssow, K.P.; Goyarts, T.; Valenta, H.; Ueberschär, K.H.; Tiemann, U. On the transfer of the Fusarium toxins deoxynivalenol (DON) and zearalenone (ZON) from the sow to the full-term piglet during the last third of gestation. Food Chem. Toxicol. 2007, 45, 1565–1574. [Google Scholar] [CrossRef]
- Goyarts, T.; Dänicke, S.; Brüssow, K.P.; Valenta, H.; Ueberschär, K.H.; Tiemann, U. On the transfer of the Fusarium toxins deoxynivalenol (DON) and zearalenone (ZON) from sows to their fetuses during days 35–70 of gestation. Toxicol. Lett. 2007, 171, 38–49. [Google Scholar] [CrossRef]
- Zhao, Y.; Zhu, X.; Wu, H.; Zhuang, D.; Yu, G.; Li, X.; Li, F.; Yu, A. Evaluation of Fetal Skeletal Malformations in Deoxynivalenol-Treated Mice Using Microarray Analysis. Arch. Environ. Contam. Toxicol. 2012, 63, 445–452. [Google Scholar] [CrossRef] [PubMed]
- Yu, M.; Chen, L.; Peng, Z.; Wang, D.; Song, Y.; Wang, H.; Yao, P.; Yan, H.; Nüssler, A.; Liu, L.; et al. Embryotoxicity Caused by DON-Induced Oxidative Stress Mediated by Nrf2/HO-1 Pathway. Toxins 2017, 9, 188. [Google Scholar] [CrossRef] [PubMed]
- Gao, X.; Sun, L.; Zhang, N.; Li, C.; Zhang, J.; Xiao, Z.; Qi, D. Gestational Zearalenone Exposure Causes Reproductive and Developmental Toxicity in Pregnant Rats and Female Offspring. Toxins 2017, 9, 21. [Google Scholar] [CrossRef] [PubMed]
- Prouillac, C.; Videmann, B.; Mazallon, M.; Lecoeur, S. Induction of cells differentiation and ABC transporters expression by a myco-estrogen, zearalenone, in human choriocarcinoma cell line (BeWo). Toxicology 2009, 263, 100–107. [Google Scholar] [CrossRef]
- Prouillac, C.; Koraichi, F.; Videmann, B.; Mazallon, M.; Rodriguez, F.; Baltas, M.; Lecoeur, S. In vitro toxicological effects of estrogenic mycotoxins on human placental cells: Structure activity relationships. Toxicol. Appl. Pharm. 2012, 259, 366–375. [Google Scholar] [CrossRef]
- Tanaka, T.; Abe, H.; Kimura, M.; Onda, N.; Mizukami, S.; Yoshida, T.; Shibutani, M. Developmental exposure to T-2 toxin reversibly affects postnatal hippocampal neurogenesis and reduces neural stem cells and progenitor cells in mice. Arch. Toxicol. 2016, 90, 2009–2024. [Google Scholar] [CrossRef]
- Sehata, S.; Kiyosawa, N.; Makino, T.; Atsumi, F.; Ito, K.; Yamoto, T.; Teranishi, M.; Baba, Y.; Uetsuka, K.; Nakayama, H.; et al. Morphological and microarray analysis of T-2 toxin-induced rat fetal brain lesion. Food Chem. Toxicol. 2004, 42, 1727–1736. [Google Scholar] [CrossRef]
- Rousseaux, C.G.; Schiefer, H.B. Maternal toxicity, embryolethality and abnormal fetal development in CD-1 mice following one oral dose of T-2 toxin. J. Appl. Toxicol. 1987, 7, 281–288. [Google Scholar] [CrossRef]
- Holladay, S.D.; Blaylock, B.L.; Comment, C.E.; Heindel, J.J.; Luster, M.I. Fetal thymic atrophy after exposure to T-2 toxin: Selectivity for lymphoid progenitor cells. Toxicol. Appl. Pharm. 1993, 121, 8–14. [Google Scholar] [CrossRef]
- Holladay, S.D.; Smith, B.J.; Luster, M.I. B lymphocyte precursor cells represent sensitive targets of T2 mycotoxin exposure. Toxicol. Appl. Pharm. 1995, 131, 309–315. [Google Scholar] [CrossRef]
- Bode, C.J.; Jin, H.; Rytting, E.; Silverstein, P.S.; Young, A.M.; Audus, K.L. In Vitro Models for Studying Trophoblast Transcellular Transport. Methods Mol. Med. 2006, 122, 225–239. [Google Scholar] [PubMed]
- Poulsen, M.S.; Rytting, E.; Mose, T.; Knudsen, L.E. Modeling placental transport: Correlation of in vitro BeWo cell permeability and ex vivo human placental perfusion. Toxicol. Vitr. 2009, 23, 1380–1386. [Google Scholar] [CrossRef]
- Smith, C.; Moe, A.; Ganapathy, V. Nutrient transport pathways across the epithelium of the placenta. Annu. Rev. Nutr. 1992, 12, 183–206. [Google Scholar] [CrossRef] [PubMed]
- Pidoux, G.; Gerbaud, P.; Gnidehou, S.; Grynberg, M.; Geneau, G.; Guibourdenche, J.; Carette, D.; Cronier, L.; Evain-Brion, D.; Malassine, A.; et al. ZO-1 is involved in trophoblastic cell differentiation in human placenta. Am. J. Physiol. Cell Physiol. 2010, 298, C1517–C1526. [Google Scholar] [CrossRef] [PubMed]
- Marzioni, D.; Banita, M.; Felici, A.; Paradinas, F.; Newlands, E.; De Nictolis, M.; Mühlhauser, J.; Castellucci, M. Expression of ZO-1 and occludin in normal human placenta and in hydatidiform moles. Mol. Hum. Reprod. 2001, 7, 279–285. [Google Scholar] [CrossRef] [PubMed]
- Ahn, C.; Yang, H.; Lee, D.; An, B.S.; Jeung, E.B. Placental claudin expression and its regulation by endogenous sex steroid hormones. Steroids 2015, 100, 44–51. [Google Scholar] [CrossRef]
- Tossetta, G.; Paolinelli, F.; Avellini, C.; Salvolini, E.; Ciarmela, P.; Lorenzi, T.; Emanuelli, M.; Toti, P.; Giuliante, R.; Gesuita, R.; et al. IL-1beta and TGF-beta weaken the placental barrier through destruction of tight junctions: An in vivo and in vitro study. Placenta 2014, 35, 509–516. [Google Scholar] [CrossRef]
- Hannan, N.J.; Paiva, P.; Dimitriadis, E.; Salamonsen, L.A. Models for Study of Human Embryo Implantation: Choice of Cell Lines? Biol. Reprod. 2010, 82, 235–245. [Google Scholar] [CrossRef]
- Li, H.; van Ravenzwaay, B.; Rietjens, I.M.; Louisse, J. Assessment of an in vitro transport model using BeWo b30 cells to predict placental transfer of compounds. Arch. Toxicol. 2013, 87, 1661–1669. [Google Scholar] [CrossRef]
- Griffiths, S.K.; Campbell, J.P. Placental structure, function and drug transfer. Contin. Educ. Anaesth. Crit. Care Pain 2015, 15, 84–89. [Google Scholar] [CrossRef]
- Tossetta, G.; Paolinelli, F.; Lorenzi, T.; Ciarmela, P.; Avellini, C.; Salvolini, E.; Toti, P.; Petraglia, F.; Castellucci, M.; Marzioni, D. Analysis of tight junctions in placentas affected by chorioamnionitis: In vivo and in vitro analysis. Ital. J. Anat. Embryol. 2012, 117, 193. [Google Scholar]
- Hartsock, A.; Nelson, W.J. Adherens and tight junctions: Structure, function and connections to the actin cytoskeleton. Biochim. Biophys. Acta BBA Biomembr. 2008, 1778, 660–669. [Google Scholar] [CrossRef] [PubMed]
- Massart, F.; Meucci, V.; Saggese, G.; Soldani, G. High Growth Rate of Girls with Precocious Puberty Exposed to Estrogenic Mycotoxins. J. Pediatrics 2008, 152, 690–695. [Google Scholar] [CrossRef] [PubMed]
- Deng, F.; Tao, F.B.; Liu, D.Y.; Xu, Y.Y.; Hao, J.H.; Sun, Y.; Su, P.Y. Effects of growth environments and two environmental endocrine disruptors on children with idiopathic precocious puberty. Eur. J. Endocrinol. 2012, 166, 803–809. [Google Scholar] [CrossRef] [Green Version]
- Chan-Hon-Tong, A.; Charles, M.-A.; Forhan, A.; Heude, B.; Sirot, V. Exposure to food contaminants during pregnancy. Sci. Total Environ. 2013, 458–460, 27–35. [Google Scholar] [CrossRef]
- González-Osnaya, L.; Cortés, C.; Soriano, J.M.; Moltó, J.C.; Mañes, J. Occurrence of deoxynivalenol and T-2 toxin in bread and pasta commercialised in Spain. Food Chem. 2011, 124, 156–161. [Google Scholar] [CrossRef]
- Schothorst, R.C.; van Egmond, H.P. Report from SCOOP task 3.2.10 “collection of occurrence data of Fusarium toxins in food and assessment of dietary intake by the population of EU member states”: Subtask: Trichothecenes. Toxicol. Lett. 2004, 153, 133–143. [Google Scholar] [CrossRef]
- Steinkellner, H.; Binaglia, M.; Dall’Asta, C.; Gutleb, A.C.; Metzler, M.; Oswald, I.P.; Parent-Massin, D.; Alexander, J. Combined hazard assessment of mycotoxins and their modified forms applying relative potency factors: Zearalenone and T2/HT2 toxin. Food Chem. Toxicol. 2019, 131, 110599. [Google Scholar] [CrossRef]
- Al-Jaal, B.A.; Jaganjac, M.; Barcaru, A.; Horvatovich, P.; Latiff, A. Aflatoxin, fumonisin, ochratoxin, zearalenone and deoxynivalenol biomarkers in human biological fluids: A systematic literature review, 2001–2018. Food Chem. Toxicol. 2019, 129, 211–228. [Google Scholar] [CrossRef] [Green Version]
- Alassane-Kpembi, I.; Schatzmayr, G.; Taranu, I.; Marin, D.; Puel, O.; Oswald, I.P. Mycotoxins co-contamination: Methodological aspects and biological relevance of combined toxicity studies. Crit. Rev. Food Sci. Nutr. 2017, 57, 3489–3507. [Google Scholar] [CrossRef]
- Alassane-Kpembi, I.; Kolf-Clauw, M.; Gauthier, T.; Abrami, R.; Abiola, F.A.; Oswald, I.P.; Puel, O. New insights into mycotoxin mixtures: The toxicity of low doses of Type B trichothecenes on intestinal epithelial cells is synergistic. Toxicol. Appl. Pharmacol. 2013, 272, 191–198. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Wang, W.; Cheng, G.; Huang, L.; Chen, D.; Tao, Y.; Pan, Y.; Hao, H.; Wu, Q.; Wan, D.; et al. High Risk of Embryo-Fetal Toxicity: Placental Transfer of T-2 Toxin and Its Major Metabolite HT-2 Toxin in BeWo Cells. Toxicol. Sci. 2013, 137, 168–178. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Niessen, C.M. Tight junctions/adherens junctions: Basic structure and function. J. Investig. Dermatol. 2007, 127, 2525–2532. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Akbari, P.; Braber, S.; Gremmels, H.; Koelink, P.J.; Verheijden, K.A.T.; Garssen, J.; Fink-Gremmels, J. Deoxynivalenol: A trigger for intestinal integrity breakdown. FASEB J. 2014, 28, 2414–2429. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Varasteh, S.; Braber, S.; Garssen, J.; Fink-Gremmels, J. Galacto-oligosaccharides exert a protective effect against heat stress in a Caco-2 cell model. J. Funct. Foods 2015, 16, 265–277. [Google Scholar] [CrossRef]
- Maiers, J.L.; Peng, X.; Fanning, A.S.; DeMali, K.A. ZO-1 recruitment to α-catenin--a novel mechanism for coupling the assembly of tight junctions to adherens junctions. J. Cell Sci. 2013, 126, 3904–3915. [Google Scholar] [CrossRef] [Green Version]
- Itoh, M.; Nagafuchi, A.; Moroi, S.; Tsukita, S. Involvement of ZO-1 in cadherin-based cell adhesion through its direct binding to alpha catenin and actin filaments. J. Cell Biol. 1997, 138, 181–192. [Google Scholar] [CrossRef] [Green Version]
- Wu, Q.H.; Wang, X.; Yang, W.; Nüssler, A.K.; Xiong, L.Y.; Kuča, K.; Dohnal, V.; Zhang, X.J.; Yuan, Z.H. Oxidative stress-mediated cytotoxicity and metabolism of T-2 toxin and deoxynivalenol in animals and humans: An update. Arch. Toxicol. 2014, 88, 1309–1326. [Google Scholar] [CrossRef]
- Bensassi, F.; Gallerne, C.; El Dein, O.S.; Lemaire, C.; Hajlaoui, M.R.; Bacha, H. Involvement of mitochondria-mediated apoptosis in deoxynivalenol cytotoxicity. Food Chem. Toxicol. 2012, 50, 1680–1689. [Google Scholar] [CrossRef]
- Braicu, C.; Laffitte, J.; Pinton, P.; Oswald, I.P.; Taranu, I.; Nougayrede, J.-P. Deoxynivalenol Impairs Porcine Intestinal Barrier Function and Decreases the Protein Expression of Claudin-4 through a Mitogen-Activated Protein Kinase-Dependent Mechanism. J. Nutr. 2010, 140, 1956–1962. [Google Scholar] [CrossRef] [Green Version]
- Abid-Essefi, S.; Ouanes, Z.; Hassen, W.; Baudrimont, I.; Creppy, E.; Bacha, H. Cytotoxicity, inhibition of DNA and protein syntheses and oxidative damage in cultured cells exposed to zearalenone. Toxicol. In Vitro 2004, 18, 467–474. [Google Scholar] [CrossRef] [PubMed]
- Egawa, M.; Kamata, H.; Kushiyama, A.; Sakoda, H.; Fujishiro, M.; Horike, N.; Yoneda, M.; Nakatsu, Y.; Ying, G.; Jun, Z.; et al. Long-term Forskolin Stimulation Induces AMPK Activation and Thereby Enhances Tight Junction Formation in Human Placental Trophoblast BeWo Cells. Placenta 2008, 29, 1003–1008. [Google Scholar] [CrossRef] [PubMed]
- Hauguel-de Mouzon, S.; Guerre-Millo, M. The placenta cytokine network and inflammatory signals. Placenta 2006, 27, 794–798. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, M.D.; Trautman, M.S.; Dudley, D.J. Cytokine networking in the placenta. Placenta 1993, 14, 249–275. [Google Scholar] [CrossRef]
- Fujisawa, N.K.; Nasu, K.; Arima, K.; Sugano, T.; Narahara, H.; Miyakawa, I. Production of interleukin (IL)-6 and IL-8 by a choriocarcinoma cell line, BeWo. Placenta 2000, 21, 354–360. [Google Scholar] [CrossRef] [PubMed]
- Capaldo, C.T.; Nusrat, A. Cytokine regulation of tight junctions. Biochim. Biophys. Acta (BBA) Biomembr. 2009, 1788, 864–871. [Google Scholar] [CrossRef] [Green Version]
- Desai, T.R.; Leeper, N.J.; Hynes, K.L.; Gewertz, B.L. Interleukin-6 Causes Endothelial Barrier Dysfunction via the Protein Kinase C Pathway. J. Surg. Res. 2002, 104, 118–123. [Google Scholar] [CrossRef]
- Suzuki, T.; Yoshinaga, N.; Tanabe, S. Interleukin-6 (IL-6) regulates claudin-2 expression and tight junction permeability in intestinal epithelium. J. Biol. Chem. 2011, 286, 31263–31271. [Google Scholar] [CrossRef] [Green Version]
- Tsukihara, S.; Harada, T.; Deura, I.; Mitsunari, M.; Yoshida, S.; Iwabe, T.; Terakawa, N. Interleukin-1β-induced Expression of IL-6 and Production of Human Chorionic Gonadotropin in Human Trophoblast Cells via Nuclear Factor-κB Activation. Am. J. Reprod. Immunol. 2004, 52, 218–223. [Google Scholar] [CrossRef]
- Lager, S.; Jansson, N.; Olsson, A.L.; Wennergren, M.; Jansson, T.; Powell, T.L. Effect of IL-6 and TNF-α on fatty acid uptake in cultured human primary trophoblast cells. Placenta 2011, 32, 121–127. [Google Scholar] [CrossRef]
- Dahlgren, J.; Samuelsson, A.M.; Jansson, T.; Holmäng, A. Interleukin-6 in the Maternal Circulation Reaches the Rat Fetus in Mid-gestation. Pediatric Res. 2006, 60, 147. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Samuelsson, A.M.; Öhrn, I.; Dahlgren, J.; Eriksson, E.; Angelin, B.; Folkow, B.r.; Holmäng, A. Prenatal Exposure to Interleukin-6 Results in Hypertension and Increased Hypothalamic-Pituitary-Adrenal Axis Activity in Adult Rats. Endocrinology 2004, 145, 4897–4911. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dahlgren, J.; Nilsson, C.; Jennische, E.; Ho, H.P.; Eriksson, E.; Niklasson, A.; Björntorp, P.; Wikland, K.A.; Holmäng, A. Prenatal cytokine exposure results in obesity and gender-specific programming. Am. J. Physiol. Endocrinol. Metab. 2001, 281, E326–E334. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stephanou, A.; Myatt, L.; Eis, A.; Sarlis, N.; Jikihara, H.; Handwerger, S. Ontogeny of the expression and regulation of interleukin-6 (IL-6) and IL-1 mRNAs by human trophoblast cells during differentiation in vitro. J. Endocrinol. 1995, 147, 487–496. [Google Scholar] [CrossRef]
- Guo, J.; Song, D.; Han, F.; Zhang, W.; Wang, Y.; Wang, Y.; Du, W. In vitro transport mechanism of psoralen in a human placental cell line (BeWo cells). Planta Med. 2015, 81, 138–144. [Google Scholar] [CrossRef]
- Correia Carreira, S.; Cartwright, L.; Mathiesen, L.; Knudsen, L.E.; Saunders, M. Studying placental transfer of highly purified non-dioxin-like PCBs in two models of the placental barrier. Placenta 2011, 32, 283–291. [Google Scholar] [CrossRef]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| TJ or AJ | mRNA | Protein | IF Staining |
|---|---|---|---|
| Mycotoxin DON | |||
| ZO-1 | ↑ | ↓ | Decreased and irregular assembly |
| OCLD | ↑ | − | Increased delocalization and intracellular accumulation |
| CLDNs | − | ↓ | Decreased and modified assembly and belt-like structure |
| E-cad | ↓ | ↓ | Decreased and irregular assembly |
| Mycotoxin ZEN | |||
| ZO-1 | ↓ | − | No obvious effect |
| OCLD | − | − | Increased delocalization and intracellular accumulation |
| CLDNs | ↓ | − | Modified assembly and belt-like structure |
| E-cad | ↓ | − | No obvious effect |
| Mycotoxin T-2 toxin | |||
| ZO-1 | ↑ | ↓ | Decreased and irregular assembly |
| OCLD | ↑ | Increased delocalization and intracellular accumulation | |
| CLDNs | − | ↓ | Decreased and modified assembly and belt-like structure |
| E-cad | ↓ | ↓ | Decreased and irregular assembly |
| Genes | Primer Sequence (5′–3′) | References | AT | |
|---|---|---|---|---|
| Forward | Reverse | |||
| β-actin | CTGGAACGGTGAAGGTGACA | AAGGGACTTCCTGTAACAATGCA | NM-001101 | 63 |
| Claudin-3 | CTGCTCTGCTGCTCGTGTC | CGTAGTCCTTGCGGTCGTAG | NM-001306 | 63 |
| Claudin-4 | GTCTGCCTGCATCTCCTCTGT | CCTCTAAACCCGTCCATCCA | NM-001305 | 62.5 |
| E-cadherin | TGGACCGAGAGAGTTTCCCT | CCCTTGTACGTGGTGGGATT | BC-144283.1 | 60 |
| Occludin | TTGGATAAAGAATTGGATGACT | ACTGCTTGCAATGATTCTTCT | NM-002538 | 57 |
| ZO-1 | GAATGATGGTTGGTATGGTGCG | TCAGAAGTGTGTCTACTGTCCG | NT-010194.17 | 55.8 |
| IL-6 | TACCCCCAGGAGAAGATTCC | TTTTCTGCCAGTGCCTCTTT | S56892.1 | 63 |
| IL-8 | CTCTTGGCAGCCTTCCTGATT | TATGCACTGACATCTAAGTTCTTTAGCA | NM-000584.3 | 60 |
| IL-1β | GCTGAGGAAGATGCTGGTTC | GTGATCGTACAGGTGCATCG | NM-000756 | 57 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Seyed Toutounchi, N.; Hogenkamp, A.; Varasteh, S.; van’t Land, B.; Garssen, J.; Kraneveld, A.D.; Folkerts, G.; Braber, S. Fusarium Mycotoxins Disrupt the Barrier and Induce IL-6 Release in a Human Placental Epithelium Cell Line. Toxins 2019, 11, 665. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins11110665
Seyed Toutounchi N, Hogenkamp A, Varasteh S, van’t Land B, Garssen J, Kraneveld AD, Folkerts G, Braber S. Fusarium Mycotoxins Disrupt the Barrier and Induce IL-6 Release in a Human Placental Epithelium Cell Line. Toxins. 2019; 11(11):665. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins11110665
Chicago/Turabian StyleSeyed Toutounchi, Negisa, Astrid Hogenkamp, Soheil Varasteh, Belinda van’t Land, Johan Garssen, Aletta D. Kraneveld, Gert Folkerts, and Saskia Braber. 2019. "Fusarium Mycotoxins Disrupt the Barrier and Induce IL-6 Release in a Human Placental Epithelium Cell Line" Toxins 11, no. 11: 665. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins11110665