Deoxynivalenol Inhibits Porcine Intestinal Trefoil Factors Expression in Weanling Piglets and IPEC-J2 Cells

 ,
,
Abstract
:1. Introduction
2. Results
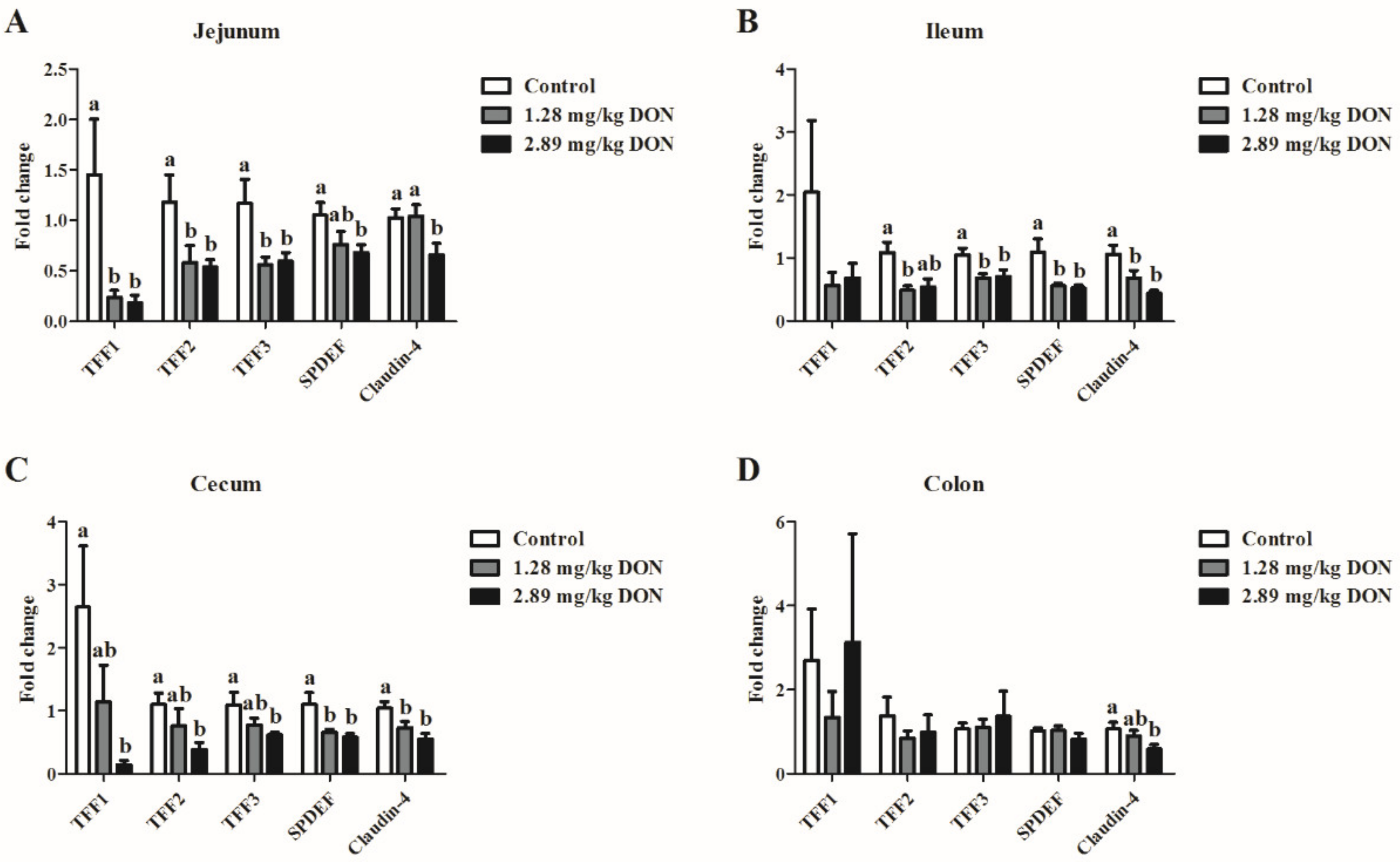
2.1. mRNA Expressions of TFFs Genes in the Intestine
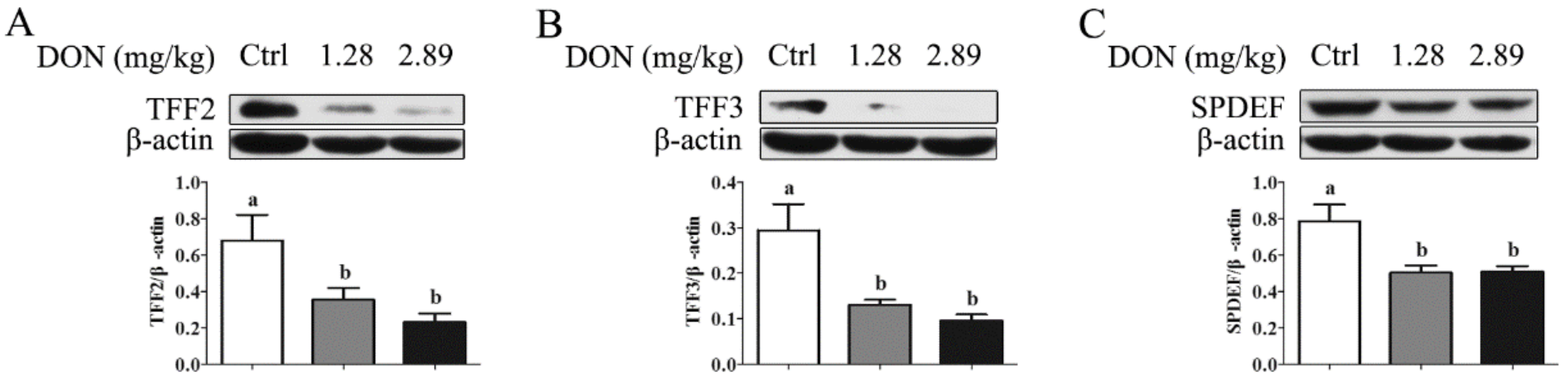
2.2. Depression of SPDEF in the Jejunum
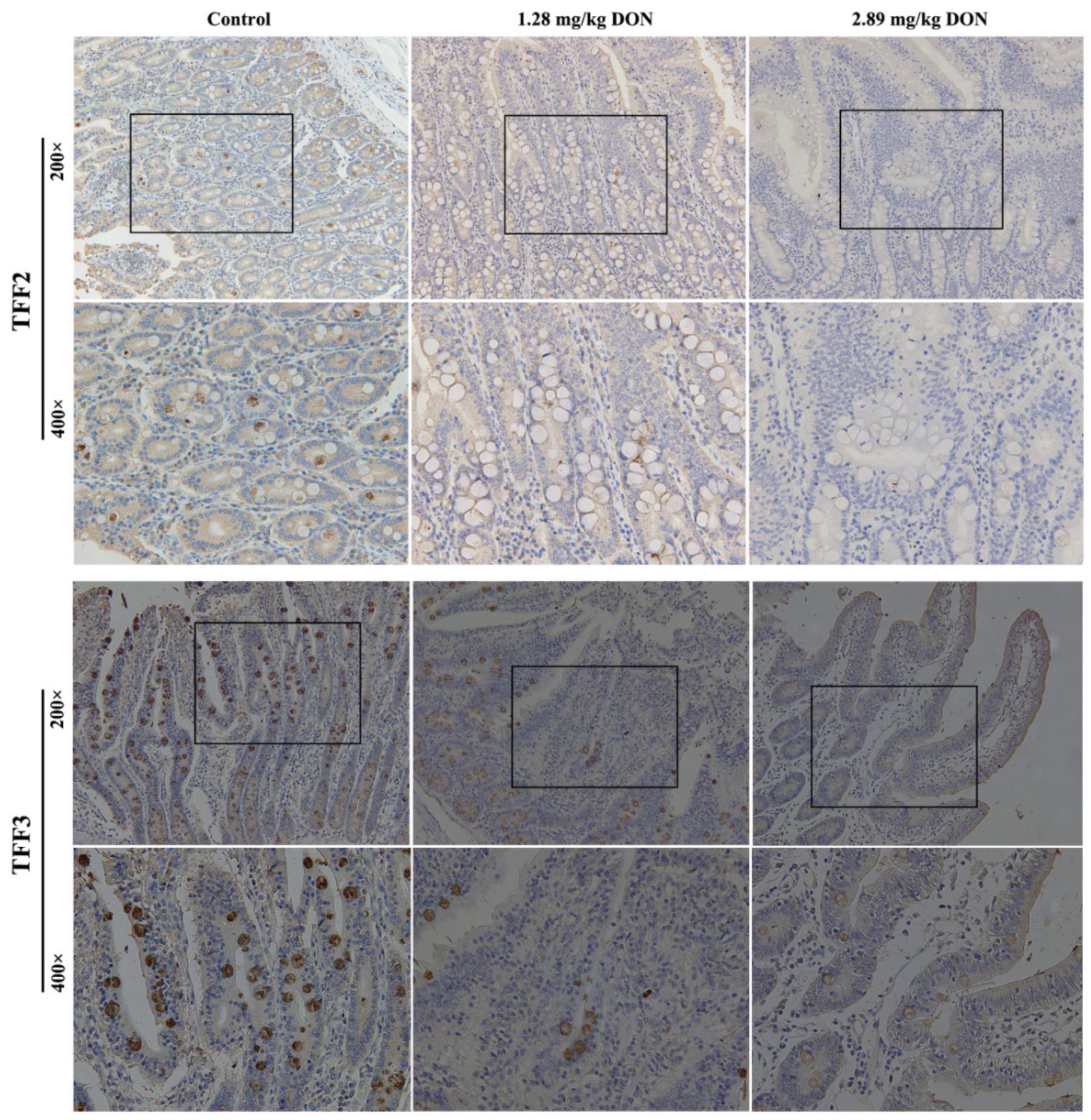
2.3. TFF Staining in the Jejunum
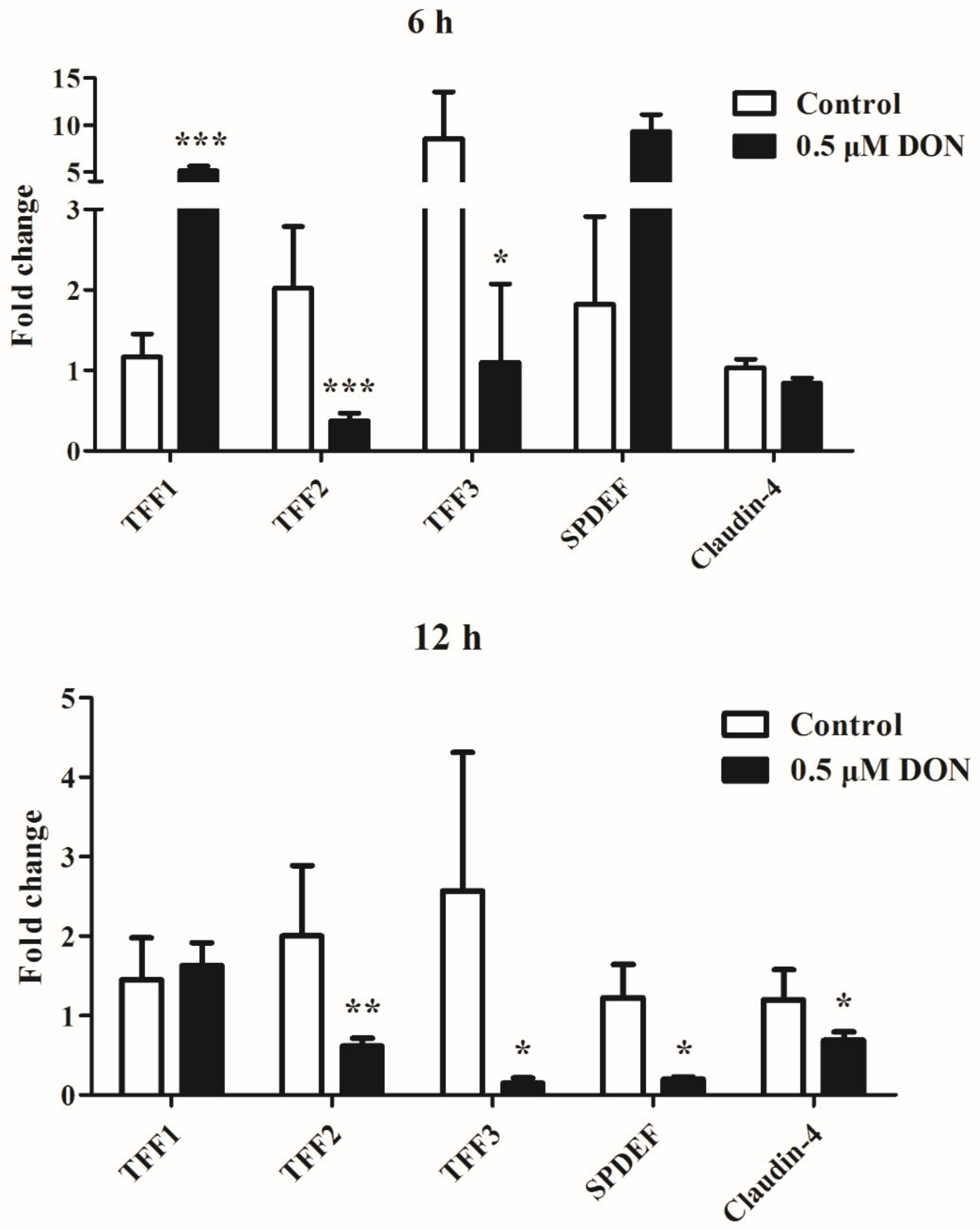
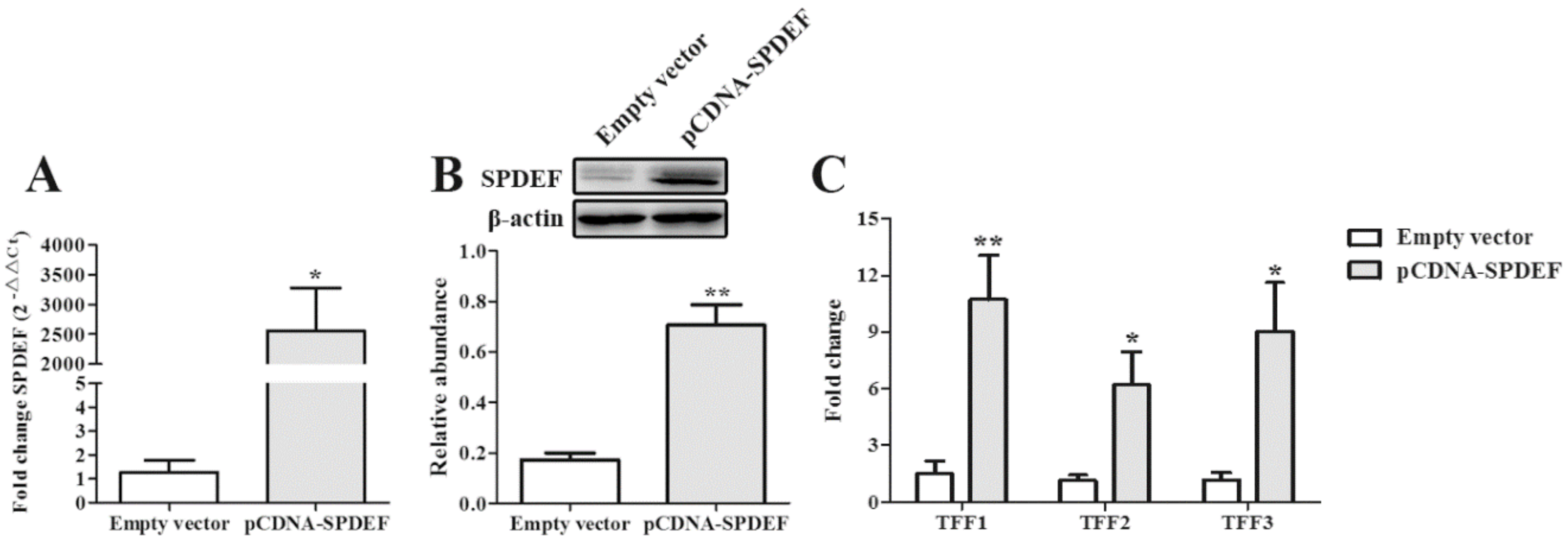
2.4. DON Inhibits the mRNA Expression of TFFs by IPEC-J2 cells
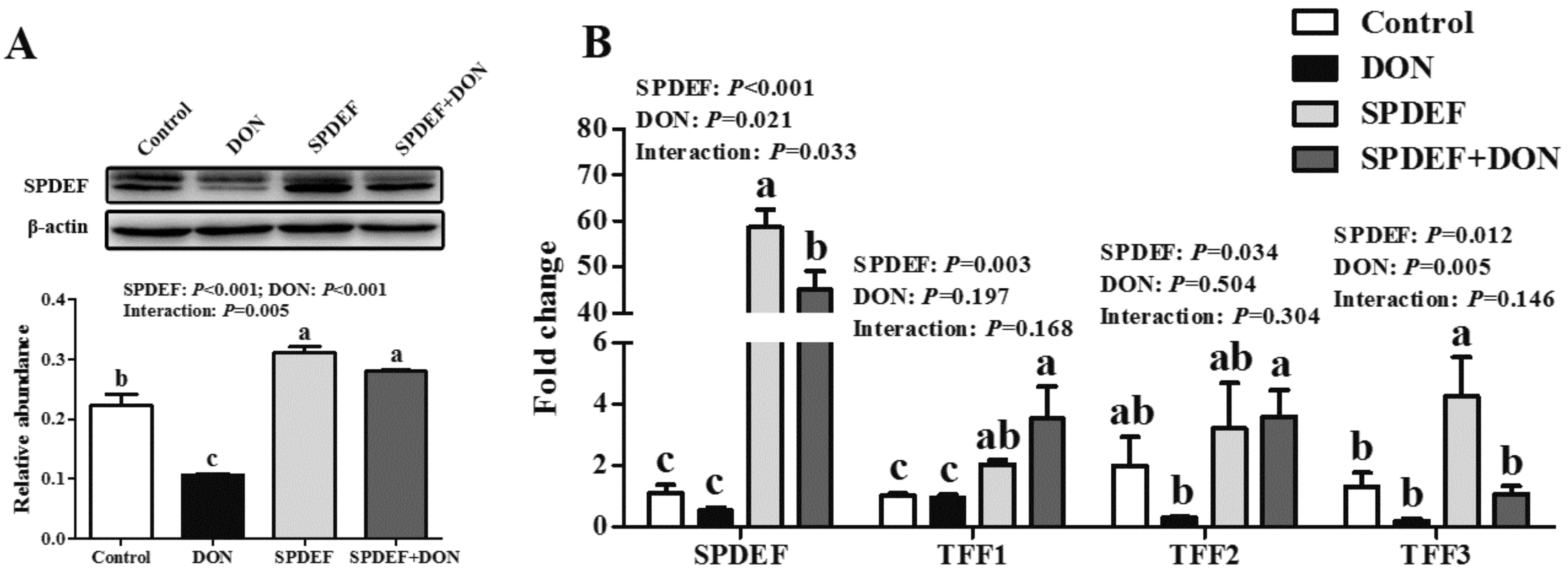
2.5. SPDEF was a Critical Regulator Regulating TFFs Expression in Response to DON Exposure
3. Discussion
4. Materials and Methods
4.1. Animals and Experimental Design
4.2. Immunohistochemistry
4.3. Cell Culture and Treatment
4.4. Plasmid Construct and Transfection
4.5. Quantitative PCR
4.6. Western Blotting
4.7. Statistical Analysis
Author Contributions
Funding
Conflicts of Interest
References
- Reddy, K.R.N.; Salleh, B.; Saad, B.; Abbas, H.K.; Abel, C.A.; Shier, W.T. An overview of mycotoxin contamination in foods and its implications for human health. Toxin Rev. 2010, 29, 3–26. [Google Scholar] [CrossRef]
- Pinton, P.; Oswald, I.P. Effect of deoxynivalenol and other Type B trichothecenes on the intestine: A review. Toxins 2014, 6, 1615–1643. [Google Scholar] [CrossRef]
- Pasternak, J.A.; Aiyer, V.; Hamonic, G.; Beaulieu, A.D.; Columbus, D.A.; Wilson, H.L. Molecular and physiological effects on the small intestine of weaner pigs following feeding with deoxynivalenol-contaminated feed. Toxins 2018, 10, 40. [Google Scholar] [CrossRef]
- Graziani, F.; Pinton, P.; Olleik, H.; Pujol, A.; Nicoletti, C.; Sicre, M.; Quinson, N.; Ajandouz, E.H.; Perrier, J.; Pasquale, E.D.; et al. Deoxynivalenol inhibits the expression of trefoil factors (TFF) by intestinal human and porcine goblet cells. Arch. Toxicol. 2019, 93, 1039–1049. [Google Scholar] [CrossRef]
- Taupin, D.; Podolsky, D.K. Trefoil factors: Initiators of mucosal healing. Nat. Rev. Mol. Cell Biol. 2003, 4, 721–732. [Google Scholar] [CrossRef]
- Aamann, L.; Vestergaard, E.M.; Gronbaek, H. Trefoil factors in inflammatory bowel disease. World J Gastroenterol. 2014, 20, 3223–3230. [Google Scholar] [CrossRef]
- Hensel, K.O.; Boland, V.; Postberg, J.; Zilbauer, M.; Heuschkel, R.; Vogel, S.; Godde, D.; Wirth, S.; Jenke, A.C. Differential expression of mucosal trefoil factors and mucins in pediatric inflammatory bowel diseases. Sci. Rep. 2014, 4, 7343. [Google Scholar] [CrossRef]
- Maresca, M.; Mahfoud, R.; Garmy, N.; Fantini, J. The mycotoxin deoxynivalenol affects nutrient absorption in human intestinal epithelial cells. J. Nutr. 2002, 132, 2723–2731. [Google Scholar] [CrossRef] [PubMed]
- Kolf-Clauw, M.; Castellote, J.; Joly, B.; Bourges-Abella, N.; Raymond-Letron, I.; Pinton, P.; Oswald, I.P. Development of a pig jejunal explant culture for studying the gastrointestinal toxicity of the mycotoxin deoxynivalenol: Histopathological analysis. Toxicol. In Vitro 2009, 23, 1580–1584. [Google Scholar] [CrossRef] [PubMed]
- Pinton, P.; Braicu, C.; Nougayrede, J.P.; Laffitte, J.; Taranu, I.; Oswald, I.P. Deoxynivalenol impairs porcine intestinal barrier function and decreases the protein expression of claudin-4 through a mitogen-activated protein kinase-dependent mechanism. J. Nutr. 2010, 140, 1956–1962. [Google Scholar] [CrossRef] [PubMed]
- Marko, C.K.; Menon, B.B.; Chen, G.; Whitsett, J.A.; Clevers, H.; Gipson, I.K. Spdef null mice lack conjunctival goblet cells and provide a model of dry eye. Am. J. Pathol. 2013, 183, 35–48. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.I.; Kim, I.H. Nucleotide-mediated SPDEF modulates TFF3-mediated wound healing and intestinal barrier function during the weaning process. Sci. Rep. 2018, 8, 4827. [Google Scholar] [CrossRef] [PubMed]
- Gregorieff, A.; Stange, D.E.; Kujala, P.; Begthel, H.; van den Born, M.; Korving, J.; Peters, P.J.; Clevers, H. The ets-domain transcription factor Spdef promotes maturation of goblet and paneth cells in the intestinal epithelium. Gastroenterology 2009, 137, 1333–1345. [Google Scholar] [CrossRef] [PubMed]
- Braga, E.N.; Hoffmann, W.; Brierley, S.M.; Muttenthaler, M. Trefoil factor family: Unresolved questions and clinical perspectives. Trends Biochem. Sci. 2019, 44, 387–390. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Yang, J.; Zhang, B.; Wu, K.; Yang, A.; Li, C.; Zhang, J.; Zhang, C.; Rajput, S.A.; Zhang, N.; et al. Deoxynivalenol impairs porcine intestinal host defense peptide expression in weaned piglets and IPEC-J2 cells. Toxins 2018, 10, 541. [Google Scholar] [CrossRef]
- Kjellev, S. The trefoil factor family-small peptides with multiple functionalities. Cell. Mol. Life Sci. 2009, 66, 1350–1369. [Google Scholar] [CrossRef]
- Wan, M.L.Y.; Turner, P.C.; Allen, K.J.; El-Nezami, H. Lactobacillus rhamnosus GG modulates intestinal mucosal barrier and inflammation in mice following combined dietary exposure to deoxynivalenol and zearalenone. J. Funct. Foods 2016, 22, 34–43. [Google Scholar] [CrossRef]
- Mashimo, H.; Wu, D.C.; Podolsky, D.K.; Fishman, M.C. Impaired defense of intestinal mucosa in mice lacking intestinal trefoil factor. Science 1996, 274, 262–265. [Google Scholar] [CrossRef]
- Beck, P.L.; Wong, J.F.; Li, Y.; Swaminathan, S.; Xavier, R.J.; Devaney, K.L.; Podolsky, D.K. Chemotherapy-and radiotherapy-induced intestinal damage is regulated by intestinal trefoil factor. Gastroenterology 2004, 126, 796–808. [Google Scholar] [CrossRef]
- Buda, A.; Jepson, M.A.; Pignatelli, M. Regulatory function of trefoil peptides (TFF) on intestinal cell junctional complexes. Cell Commun. Adhes. 2012, 19, 63–68. [Google Scholar] [CrossRef]
- National Research Council (NRC). Nutrient Requirements of Swine, 11th rev. ed.; The National Academies Press: Washington, DC, USA, 2012. [Google Scholar]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Luo, J.J.; Zhang, Y.; Sun, H.; Wei, J.T.; Khalil, M.M.; Wang, Y.W.; Dai, J.F.; Zhang, N.Y.; Qi, D.S.; Sun, L.H. The response of glandular gastric transcriptome to T-2 toxin in chicks. Food Chem. Toxicol. 2019, 132, 110658. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Forward Primer (5′–3′) | Reverse Primer (5′–3′) | Accession No. |
|---|---|---|---|
| β-actin | TACACCGCTACCAGTTCGC | GCTCGATGGGGTACTTGAGG | XM_021086047 |
| TFF1 | TGCCAGAGTGAACTGTGGTTTC | CAAAGCAGCAGCCTTTTTTTTC | AM283538.1 |
| TFF2 | ATCACCAGCGACCAGTGCTT | ATGACGCACTCCTCAGACTCTTG | XM_003358971.1 |
| TFF3 | CAGGATGTTCTGGCTGCTAGTG | GCAGTCCACCCTGTCCTTG | NM_001243483.1 |
| SPDEF | CATTCACCTGTGGCAGTTCC | TAGTTCATGGCAGGACGGTT | NM_001190254 |
| Claudin-4 | CTCTCGGACACCTTCCCAAG | GCAGTGGGAAGGTCAAAGG | NC_010445.4 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, S.; Zhang, C.; Wang, X.; Yang, J.; Wu, K.; Zhang, J.; Zhang, B.; Yang, A.; Qi, D. Deoxynivalenol Inhibits Porcine Intestinal Trefoil Factors Expression in Weanling Piglets and IPEC-J2 Cells. Toxins 2019, 11, 670. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins11110670
Wang S, Zhang C, Wang X, Yang J, Wu K, Zhang J, Zhang B, Yang A, Qi D. Deoxynivalenol Inhibits Porcine Intestinal Trefoil Factors Expression in Weanling Piglets and IPEC-J2 Cells. Toxins. 2019; 11(11):670. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins11110670
Chicago/Turabian StyleWang, Shuai, Cong Zhang, Xu Wang, Jiacheng Yang, Kuntan Wu, Jiacai Zhang, Beiyu Zhang, Ao Yang, and Desheng Qi. 2019. "Deoxynivalenol Inhibits Porcine Intestinal Trefoil Factors Expression in Weanling Piglets and IPEC-J2 Cells" Toxins 11, no. 11: 670. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins11110670