Identification and Characterization of ShSPI, a Kazal-Type Elastase Inhibitor from the Venom of Scolopendra Hainanum

 and
and
Abstract
:1. Introduction
2. Results
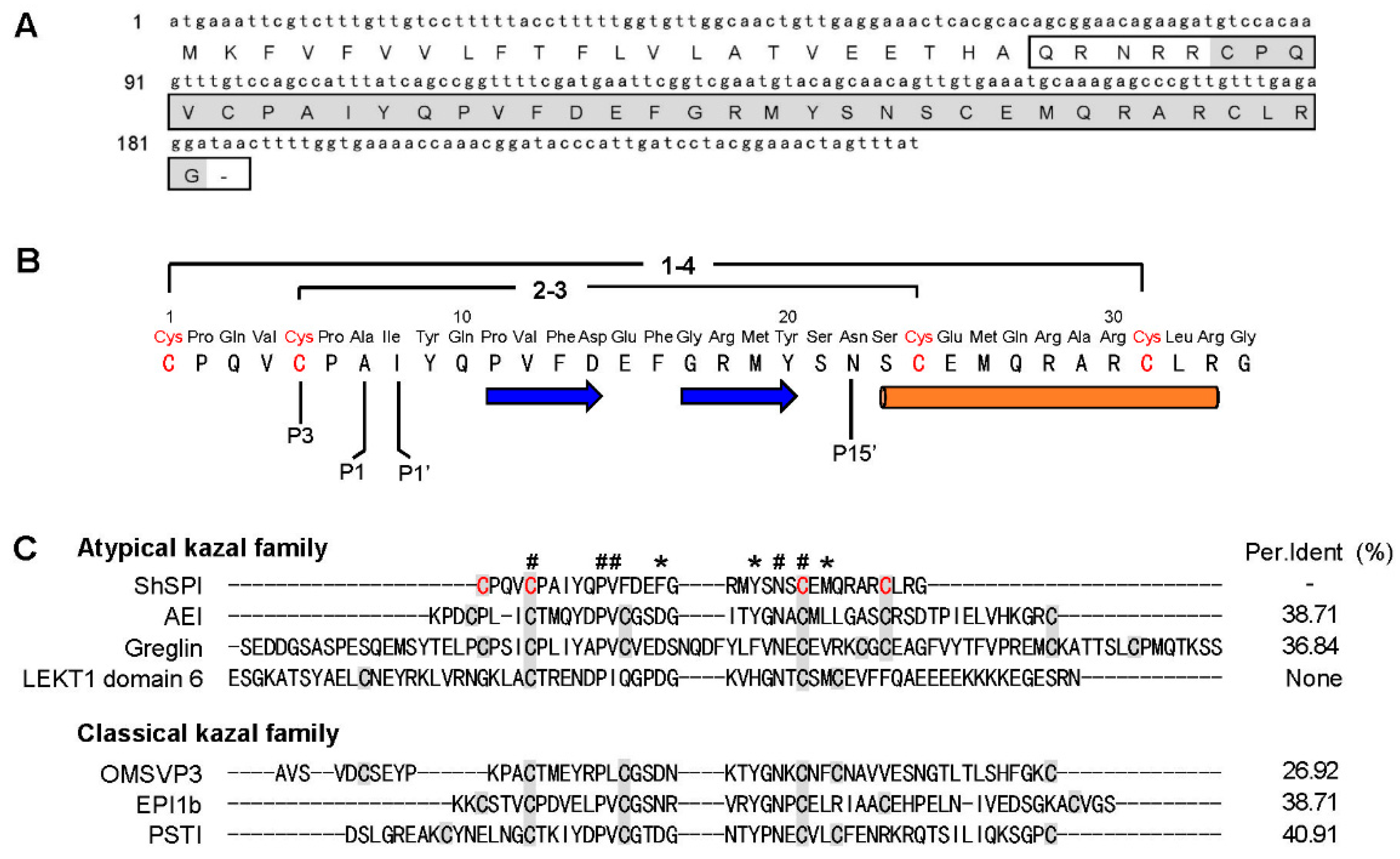
2.1. Determination of the Primary Structure of ShSPI
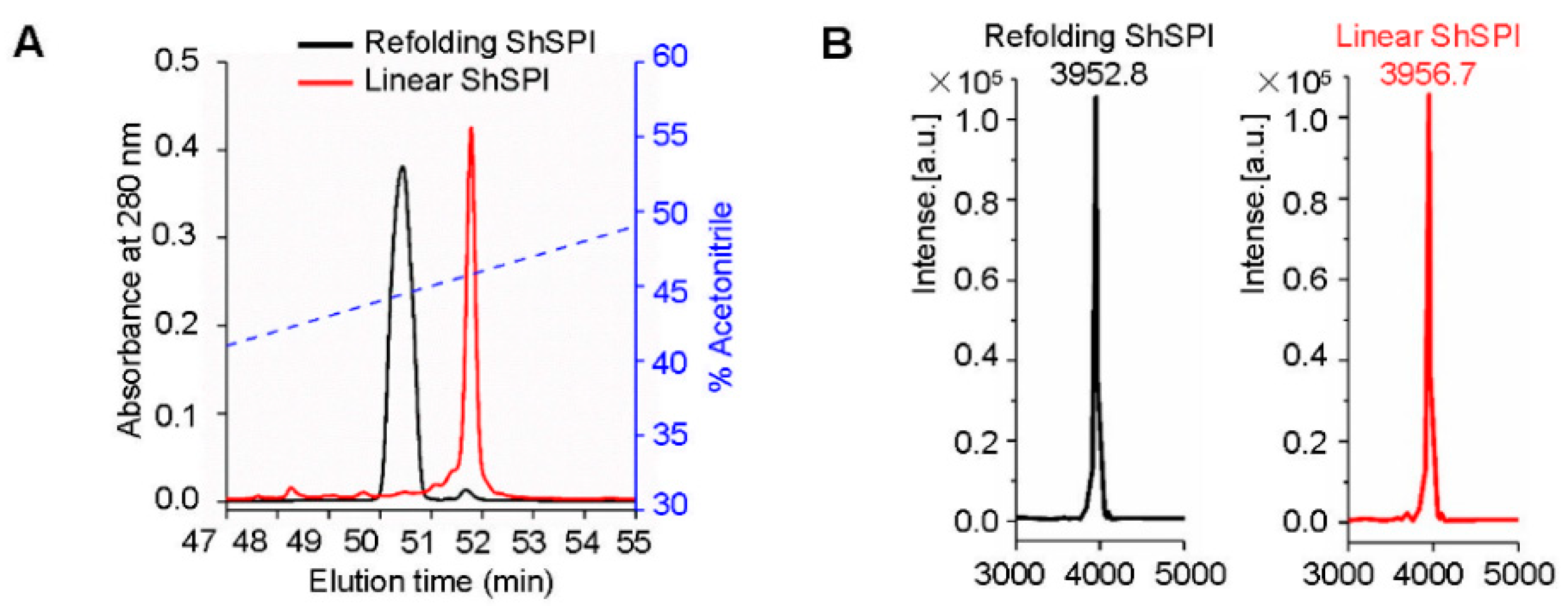
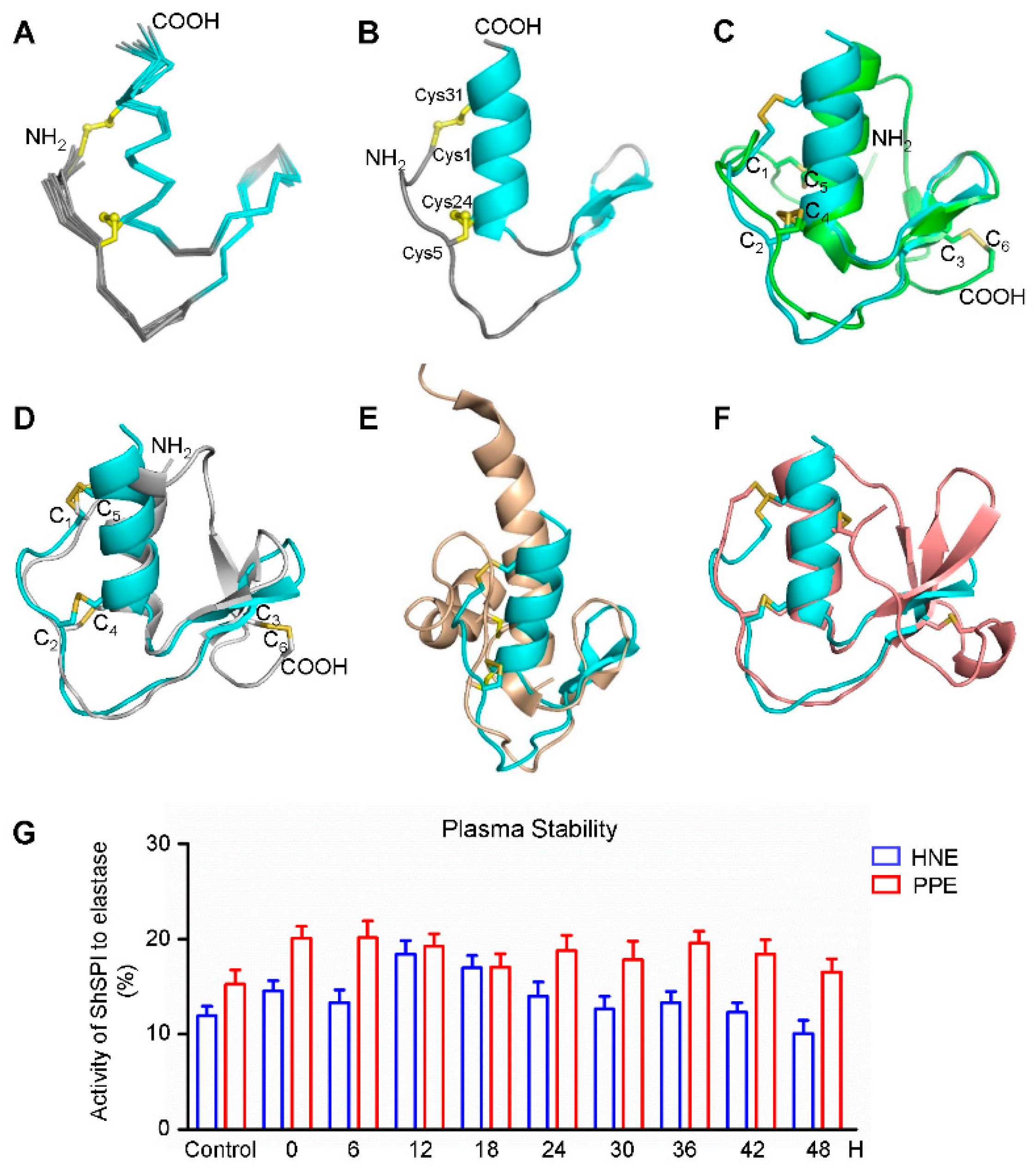
2.2. Refolding of ShSPI
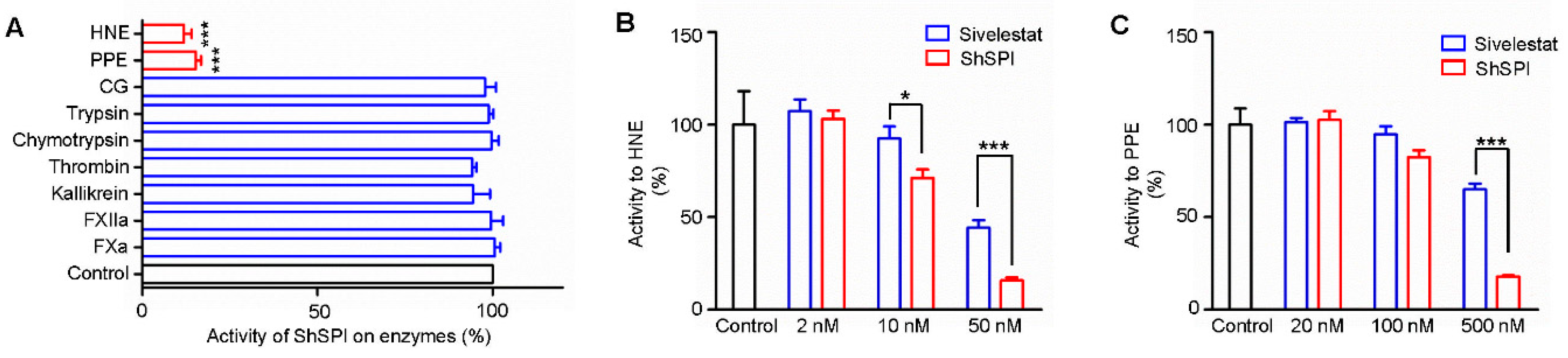
2.3. ShSPI Inhibits Elastase Potently and Selectively
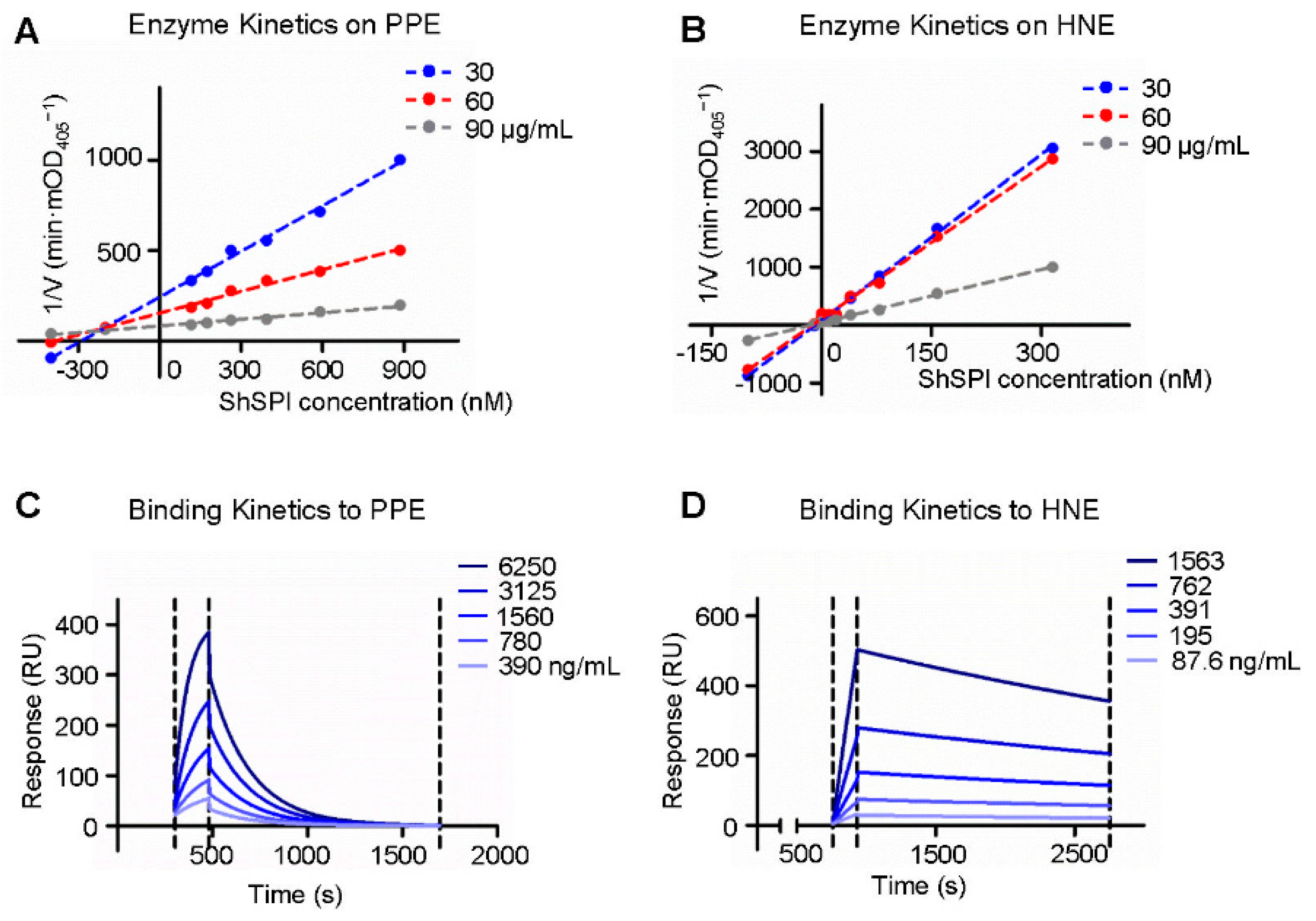
2.4. Enzymatic Kinetics of ShSPI on Elastase
2.5. Binding Kinetics of ShSPI on Elastase
2.6. The NMR Structure of ShSPI
2.7. ShSPI is an Atypical Kazal Domain and the Minimal Functional Unit
2.8. Point Mutations Confirm the Important Residues of ShSPI
2.9. ShSPI Showed Plasma Stability
3. Discussion
4. Materials and Methods
4.1. RNA Extraction and cDNA Synthesis
4.2. Peptides Synthesis and Refolding
4.3. Assignment of the Disulfide Bonds
4.4. Protease Assays
4.5. Surface Plasmon Resonance (SPR) Biosensor Analysis
4.6. NMR Experiments
4.7. Plasma Stability
4.8. Statistical Analysis
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Liu, Z.C.; Zhang, R.; Zhao, F.; Chen, Z.M.; Liu, H.W.; Wang, Y.J.; Jiang, P.; Zhang, Y.; Wu, Y.; Ding, J.P.; et al. Venomic and Transcriptomic Analysis of Centipede Scolopendra subspinipes dehaani. J. Proteome Res. 2012, 11, 6197–6212. [Google Scholar] [CrossRef]
- Kong, Y.; Shao, Y.; Chen, H.; Ming, X.; Wang, J.B.; Li, Z.Y.; Wei, J.F. A Novel Factor Xa-Inhibiting Peptide from Centipedes Venom. Int. J. Pept. Res. Ther. 2013, 19, 303–311. [Google Scholar] [CrossRef] [Green Version]
- Yang, S.; Liu, Z.; Xiao, Y.; Li, Y.; Rong, M.; Liang, S.; Zhang, Z.; Yu, H.; King, G.F.; Lai, R. Chemical punch packed in venoms makes centipedes excellent predators. Mol. Cell. Proteomics. 2012, 11, 640–650. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gonzalez-Morales, L.; Diego-Garcia, E.; Segovia, L.; Gutierrez Mdel, C.; Possani, L.D. Venom from the centipede Scolopendra viridis Say: Purification, gene cloning and phylogenetic analysis of a phospholipase A2. Toxicon 2009, 54, 8–15. [Google Scholar] [CrossRef] [PubMed]
- Luo, L.; Li, B.; Wang, S.; Wu, F.; Wang, X.; Liang, P.; Ombati, R.; Chen, J.; Lu, X.; Cui, J.; et al. Centipedes subdue giant prey by blocking KCNQ channels. Proc. Natl. Acad. Sci. USA 2018, 115, 1646–1651. [Google Scholar] [CrossRef] [Green Version]
- Yang, S.; Yang, F.; Wei, N.; Hong, J.; Li, B.; Luo, L.; Rong, M.; Yarov-Yarovoy, V.; Zheng, J.; Wang, K.; et al. A pain-inducing centipede toxin targets the heat activation machinery of nociceptor TRPV1. Nat. Commun. 2015, 6, 8297. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rong, M.; Yang, S.; Wen, B.; Mo, G.; Kang, D.; Liu, J.; Lin, Z.; Jiang, W.; Li, B.; Du, C.; et al. Peptidomics combined with cDNA library unravel the diversity of centipede venom. J. Proteomics 2015, 114, 28–37. [Google Scholar] [CrossRef] [PubMed]
- Kawabata, K.; Hagio, T.; Matsumoto, S.; Nakao, S.; Orita, S.; Aze, Y.; Ohno, H. Delayed neutrophil elastase inhibition prevents subsequent progression of acute lung injury induced by endotoxin inhalation in hamsters. Am. J. Resp. Crit. Care Med. 2000, 161, 2013–2018. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rosenbloom, J.; Macarak, E.; Piera-Velazquez, S.; Jimenez, S.A. Human Fibrotic Diseases: Current Challenges in Fibrosis Research. Methods Mol. Biol. 2017, 1627, 1–23. [Google Scholar] [CrossRef]
- Liu, H.; Chen, J.; Wang, X.; Yan, S.; Xu, Y.; San, M.; Tang, W.; Yang, F.; Cao, Z.; Li, W.; et al. Functional characterization of a new non-Kunitz serine protease inhibitor from the scorpion Lychas mucronatus. Int. J. Biol. Macromol. 2015, 72, 158–162. [Google Scholar] [CrossRef]
- Wan, H.; Lee, K.S.; Kim, B.Y.; Zou, F.M.; Yoon, H.J.; Je, Y.H.; Li, J.; Jin, B.R. A spider-derived Kunitz-type serine protease inhibitor that acts as a plasmin inhibitor and an elastase inhibitor. PLoS ONE 2013, 8, e53343. [Google Scholar] [CrossRef]
- Lee, K.S.; Kim, B.Y.; Yoon, H.J.; Choi, Y.S.; Jin, B.R. Secapin, a bee venom peptide, exhibits anti-fibrinolytic, anti-elastolytic, and anti-microbial activities. Dev. Comp. Immunol. 2016, 63, 27–35. [Google Scholar] [CrossRef] [PubMed]
- Ding, L.; Wang, X.; Liu, H.; San, M.; Xu, Y.; Li, J.; Li, S.; Cao, Z.; Li, W.; Wu, Y.; et al. A new Kunitz-type plasmin inhibitor from scorpion venom. Toxicon 2015, 106, 7–13. [Google Scholar] [CrossRef] [PubMed]
- Wan, H.; Lee, K.S.; Kim, B.Y.; Yuan, M.; Zhan, S.; You, H.; Li, J.; Jin, B.R. A spider (Araneus ventricosus) chymotrypsin inhibitor that acts as an elastase inhibitor and a microbial serine protease inhibitor. Comp. Biochem. Physiol. B Biochem. Mol. Biol. 2013, 165, 36–41. [Google Scholar] [CrossRef] [PubMed]
- Jung, H.I.; Kim, S.I.; Ha, K.S.; Joe, C.O.; Kang, K.W. Isolation and characterization of guamerin, a new human leukocyte elastase inhibitor from Hirudo nipponia. J. Biol. Chem. 1995, 270, 13879–13884. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takemasa, A.; Ishii, Y.; Fukuda, T. A neutrophil elastase inhibitor prevents bleomycin-induced pulmonary fibrosis in mice. Eur. Respir. J. 2012, 40, 1475–1482. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ishii, T.; Doi, K.; Okamoto, K.; Imamura, M.; Dohi, M.; Yamamoto, K.; Fujita, T.; Noiri, E. Neutrophil elastase contributes to acute lung injury induced by bilateral nephrectomy. Am. J. Pathol. 2010, 177, 1665–1673. [Google Scholar] [CrossRef]
- Schechter, I.; Berger, A. On the size of the active site in proteases. I. Papain. Biochem. Biophys. Res. Commun. 1967, 27, 157–162. [Google Scholar] [CrossRef]
- Koizumi, M.; Fujino, A.; Fukushima, K.; Kamimura, T.; Takimoto-Kamimura, M. Complex of human neutrophil elastase with 1/2SLPI. J. Synchrotron Radiat. 2008, 15, 308–311. [Google Scholar] [CrossRef] [Green Version]
- Aikawa, N.; Kawasaki, Y. Clinical utility of the neutrophil elastase inhibitor sivelestat for the treatment of acute respiratory distress syndrome. Ther. Clin. Risk Manag. 2014, 10, 621–629. [Google Scholar] [CrossRef] [Green Version]
- Kawabata, K.; Suzuki, M.; Sugitani, M.; Imaki, K.; Toda, M.; Miyamoto, T. ONO-5046, a novel inhibitor of human neutrophil elastase. Biochem. Biophys. Res. Commun. 1991, 177, 814–820. [Google Scholar] [CrossRef]
- Fry, B.G.; Roelants, K.; Champagne, D.E.; Scheib, H.; Tyndall, J.D.; King, G.F.; Nevalainen, T.J.; Norman, J.A.; Lewis, R.J.; Norton, R.S.; et al. The toxicogenomic multiverse: Convergent recruitment of proteins into animal venoms. Annu. Rev. Genomics. Hum. Genet. 2009, 10, 483–511. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tian, M.; Kamoun, S. A two disulfide bridge Kazal domain from Phytophthora exhibits stable inhibitory activity against serine proteases of the subtilisin family. BMC Biochem. 2005, 6, 15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cesar, P.H.S.; Cardoso Trento, M.V.; Sales, T.A.; Marques, T.R.; Braga, M.A.; Ramalho, T.C.; Marcussi, S. Molecular interactions between p-coumaric acid and snake venom toxins. J. Cell. Biochem. 2019, 120, 14594–14603. [Google Scholar] [CrossRef]
- Guarnieri, M.C.; de Albuquerque Modesto, J.C.; Perez, C.D.; Ottaiano, T.F.; Ferreira, R.D.S.; Batista, F.P.; de Brito, M.V.; Campos, I.; Oliva, M.L.V. Zoanthid mucus as new source of useful biologically active proteins. Toxicon 2018, 143, 96–107. [Google Scholar] [CrossRef]
- Kronmuller, C. Review of the subspecies of Scolopendra subspinipes Leach, 1815 with the new description of the South Chinese member of the genus Scolopendra Linnaeus, 1758 named Scolopendra hainanum spec. nov (Myriapoda, Chilopoda, Scolopendridae). Spixiana 2012, 35, 19–27. [Google Scholar]
- Luan, N.; Shen, W.; Liu, J.; Wen, B.; Lin, Z.; Yang, S.; Lai, R.; Liu, S.; Rong, M. A Combinational Strategy upon RNA Sequencing and Peptidomics Unravels a Set of Novel Toxin Peptides in Scorpion Mesobuthus martensii. Toxins 2016, 8, 286. [Google Scholar] [CrossRef] [Green Version]
- Kang, S.; Liu, Y.; Zeng, X.; Deng, H.; Luo, Y.; Chen, K.; Chen, S. Taxonomy and Identification of the Genus Scolopendra in China Using Integrated Methods of External Morphology and Molecular Phylogenetics. Sci. Rep. 2017, 7, 16032. [Google Scholar] [CrossRef] [Green Version]
- Luan, N.; Zhou, C.; Li, P.; Ombati, R.; Yan, X.; Mo, G.; Rong, M.; Lai, R.; Duan, Z.; Zheng, R. Joannsin, a novel Kunitz-type FXa inhibitor from the venom of Prospirobolus joannsi. Thromb. Haemost. 2017, 117, 1031–1039. [Google Scholar] [CrossRef] [Green Version]
- Morais, S.B.; Figueiredo, B.C.; Assis, N.R.G.; Alvarenga, D.M.; de Magalhaes, M.T.Q.; Ferreira, R.S.; Vieira, A.T.; Menezes, G.B.; Oliveira, S.C. Schistosoma mansoni SmKI-1 serine protease inhibitor binds to elastase and impairs neutrophil function and inflammation. PLoS Pathog. 2018, 14, e1006870. [Google Scholar] [CrossRef] [Green Version]
- Tang, X.; Chen, M.; Duan, Z.; Mwangi, J.; Li, P.; Lai, R. Isolation and Characterization of Poecistasin, an Anti-Thrombotic Antistasin-Type Serine Protease Inhibitor from Leech Poecilobdella manillensis. Toxins 2018, 10, 429. [Google Scholar] [CrossRef] [Green Version]
- He, X.; Shen, C.; Lu, Q.; Li, J.; Wei, Y.; He, L.; Bai, R.; Zheng, J.; Luan, N.; Zhang, Z.; et al. Prokineticin 2 Plays a Pivotal Role in Psoriasis. EBioMedicine 2016, 13, 248–261. [Google Scholar] [CrossRef] [Green Version]
- Hemmi, H.; Kumazaki, T.; Yoshizawa-Kumagaye, K.; Nishiuchi, Y.; Yoshida, T.; Ohkubo, T.; Kobayashi, Y. Structural and functional study of an Anemonia elastase inhibitor, a “nonclassical” Kazal-type inhibitor from Anemonia sulcata. Biochemistry 2005, 44, 9626–9636. [Google Scholar] [CrossRef]
- Ebata, R.; Yasukawa, K.; Nagai, K.; Saito, Y.; Higashi, K.; Homma, J.; Takada, N.; Takechi, F.; Saito, N.; Kobayashi, H.; et al. Sivelestat sodium hydrate treatment for refractory Kawasaki disease. Pediatr. Int. 2019, 61, 438–443. [Google Scholar] [CrossRef] [PubMed]
- Iwata, K.; Doi, A.; Ohji, G.; Oka, H.; Oba, Y.; Takimoto, K.; Igarashi, W.; Gremillion, D.H.; Shimada, T. Effect of neutrophil elastase inhibitor (sivelestat sodium) in the treatment of acute lung injury (ALI) and acute respiratory distress syndrome (ARDS): A systematic review and meta-analysis. Intern. Med. 2010, 49, 2423–2432. [Google Scholar] [CrossRef] [Green Version]
- Suda, K.; Takeuchi, H.; Hagiwara, T.; Miyasho, T.; Okamoto, M.; Kawasako, K.; Yamada, S.; Suganuma, K.; Wada, N.; Saikawa, Y.; et al. Neutrophil elastase inhibitor improves survival of rats with clinically relevant sepsis. Shock 2010, 33, 526–531. [Google Scholar] [CrossRef]
- Schortgen, F.; Asfar, P. Update in sepsis and acute kidney injury 2014. Am. J. Respir. Crit. Care Med. 2015, 191, 1226–1231. [Google Scholar] [CrossRef]
- Yan, J.; Li, S.; Li, S. The role of the liver in sepsis. Int. Rev. Immunol. 2014, 33, 498–510. [Google Scholar] [CrossRef] [Green Version]
- Drosatos, K.; Lymperopoulos, A.; Kennel, P.J.; Pollak, N.; Schulze, P.C.; Goldberg, I.J. Pathophysiology of sepsis-related cardiac dysfunction: Driven by inflammation, energy mismanagement, or both? Curr. Heart Fail. Rep. 2015, 12, 130–140. [Google Scholar] [CrossRef] [Green Version]
- Laskowski, M.; Qasim, M.A.; Yi, Z. Additivity-based prediction of equilibrium constants for some protein–protein associations. Curr. Opin. Struct. Biol. 2003, 13, 130–139. [Google Scholar] [CrossRef]
- He, W.; Wu, J.J.; Ning, J.; Hou, J.; Xin, H.; He, Y.Q.; Ge, G.B.; Xu, W. Inhibition of human cytochrome P450 enzymes by licochalcone A, a naturally occurring constituent of licorice. Toxicol. In Vitro 2015, 29, 1569–1576. [Google Scholar] [CrossRef] [PubMed]
- Stevens, T.; Ekholm, K.; Granse, M.; Lindahl, M.; Kozma, V.; Jungar, C.; Ottosson, T.; Falk-Hakansson, H.; Churg, A.; Wright, J.L.; et al. AZD9668: Pharmacological characterization of a novel oral inhibitor of neutrophil elastase. J. Pharmacol. Exp. Ther. 2011, 339, 313–320. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kover, K.E.; Hruby, V.J.; Uhrin, D. Sensitivity- and gradient-enhanced heteronuclear coupled/decoupled HSQC-TOCSY experiments for measuring long-range heteronuclear coupling constants. J. Magn. Reson. 1997, 129, 125–129. [Google Scholar] [CrossRef] [PubMed]
- Delaglio, F.; Grzesiek, S.; Vuister, G.W.; Zhu, G.; Pfeifer, J.; Bax, A. NMRPipe: A multidimensional spectral processing system based on UNIX pipes. J. Biomol. NMR 1995, 6, 277–293. [Google Scholar] [CrossRef] [PubMed]
- Schwieters, C.D.; Kuszewski, J.J.; Tjandra, N.; Clore, G.M. The Xplor-NIH NMR molecular structure determination package. J. Magn. Reson. 2003, 160, 65–73. [Google Scholar] [CrossRef] [Green Version]
- Williams, C.J.; Headd, J.J.; Moriarty, N.W.; Prisant, M.G.; Videau, L.L.; Deis, L.N.; Verma, V.; Keedy, D.A.; Hintze, B.J.; Chen, V.B.; et al. MolProbity: More and better reference data for improved all-atom structure validation. Protein Sci. 2018, 27, 293–315. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound | ShSPI | ||
|---|---|---|---|
| Enzyme Injected | PPE | HNE | |
| Enzyme kinetics | Ki (nM) | 225.83 ± 20 | 12.61 ± 2 |
| Binding Kinetics | ka (M−1s−1) | 4.5 × 104 | 4.5 × 103 |
| kd (s−1) | 4.7 × 10−3 | 1.9 × 10−4 | |
| KA (M−1) | 9.7 × 106 | 2.4 × 107 | |
| KD (M) | 1.0 × 10−7 | 4.2 × 10−8 | |
| Experimental Restraints of ShSPI (C1–G34) | |
|---|---|
| NOE distance restraints | 448 |
| Intra-residue | 78 |
| Sequential | 167 |
| Medium range (2 ≤ |i–j| ≤ 4) | 100 |
| Long range (|i–j| ≥ 5) | 103 |
| Hydrogen bonds constraints | 4 |
| Energies (kcal/mol) | |
| Bonds | 4.99 ± 0.39 |
| Angles | 39.88 ± 1.53 |
| Improper | 5.34 ± 0.35 |
| Van del Waals (repel) | 17.13 ± 2.00 |
| NOE | 30.16 ± 4.08 |
| r.m.s. deviations from idealized 1 | |
| Bonds (Å) | 0.003 ± 0.000 |
| Angles (deg) | 0.521 ± 0.010 |
| Improper (deg) | 0.347 ± 0.012 |
| NOE | 0.048 ± 0.003 |
| Ramachandran Plot | |
| Most favored regions | 93.8% |
| Additional allowed regions | 6.2% |
| Mean pairwise RMSD (ShSPI P2-R33) (Å) | |
| Backbone | 0.49 ± 0.10 |
| Heavy Atoms | 1.16 ± 0.20 |
| Ki Value of ShSPI to HNE: 12.61 nM | |||||
|---|---|---|---|---|---|
| Position 2 | Mutants | Ki (nM) | Position 2 | Mutants | Ki (nM) |
| P6 | P2A | 100.56 | P3 | C5A | insoluble |
| P5 | Q3A | 113.74 | P2′ | Y9A | >1 mM |
| P4 | V4A | 13.16 | P15′ | N22A | insoluble |
| P2 | P6A | 105.05 | P18′ | E25A | >1 mM |
| P3′ | Q10A | 13.55 | |||
| P14′ | S21A | 16.84 | |||
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Luan, N.; Zhao, Q.; Duan, Z.; Ji, M.; Xing, M.; Zhu, T.; Mwangi, J.; Rong, M.; Liu, J.; Lai, R. Identification and Characterization of ShSPI, a Kazal-Type Elastase Inhibitor from the Venom of Scolopendra Hainanum. Toxins 2019, 11, 708. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins11120708
Luan N, Zhao Q, Duan Z, Ji M, Xing M, Zhu T, Mwangi J, Rong M, Liu J, Lai R. Identification and Characterization of ShSPI, a Kazal-Type Elastase Inhibitor from the Venom of Scolopendra Hainanum. Toxins. 2019; 11(12):708. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins11120708
Chicago/Turabian StyleLuan, Ning, Qiyu Zhao, Zilei Duan, Mengyao Ji, Meichen Xing, Tengyu Zhu, James Mwangi, Mingqiang Rong, Jiangxin Liu, and Ren Lai. 2019. "Identification and Characterization of ShSPI, a Kazal-Type Elastase Inhibitor from the Venom of Scolopendra Hainanum" Toxins 11, no. 12: 708. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins11120708