Physiological Effects on Coexisting Microalgae of the Allelochemicals Produced by the Bloom-Forming Cyanobacteria Synechococcus sp. and Nodularia Spumigena

 , and
, and
Abstract
:1. Introduction
2. Results
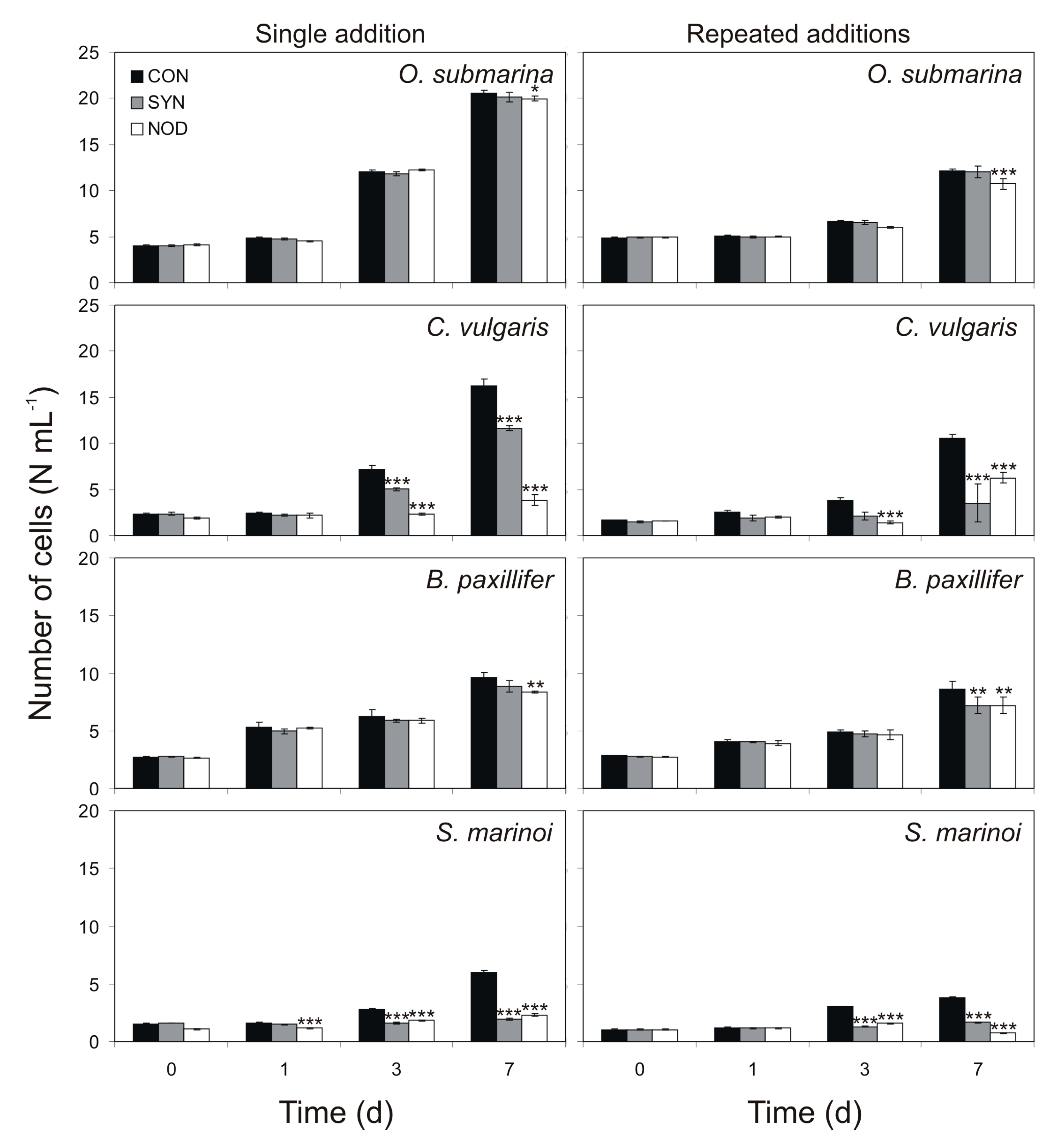
2.1. Effect of Cyanobacterial Exudates on Growth
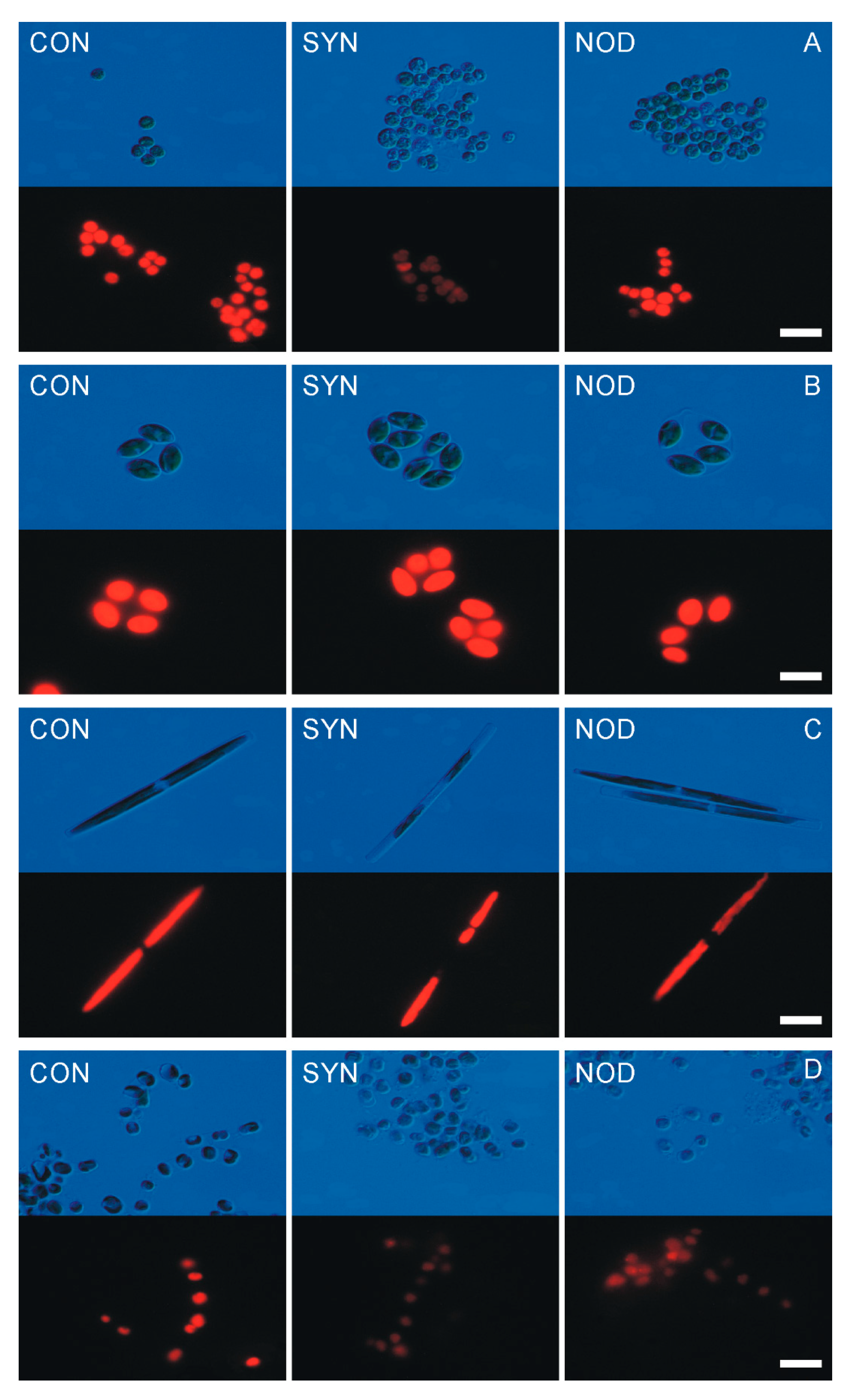
2.2. Allelopathic Effects of Cyanobacterial Exudates on the Cell Morphology
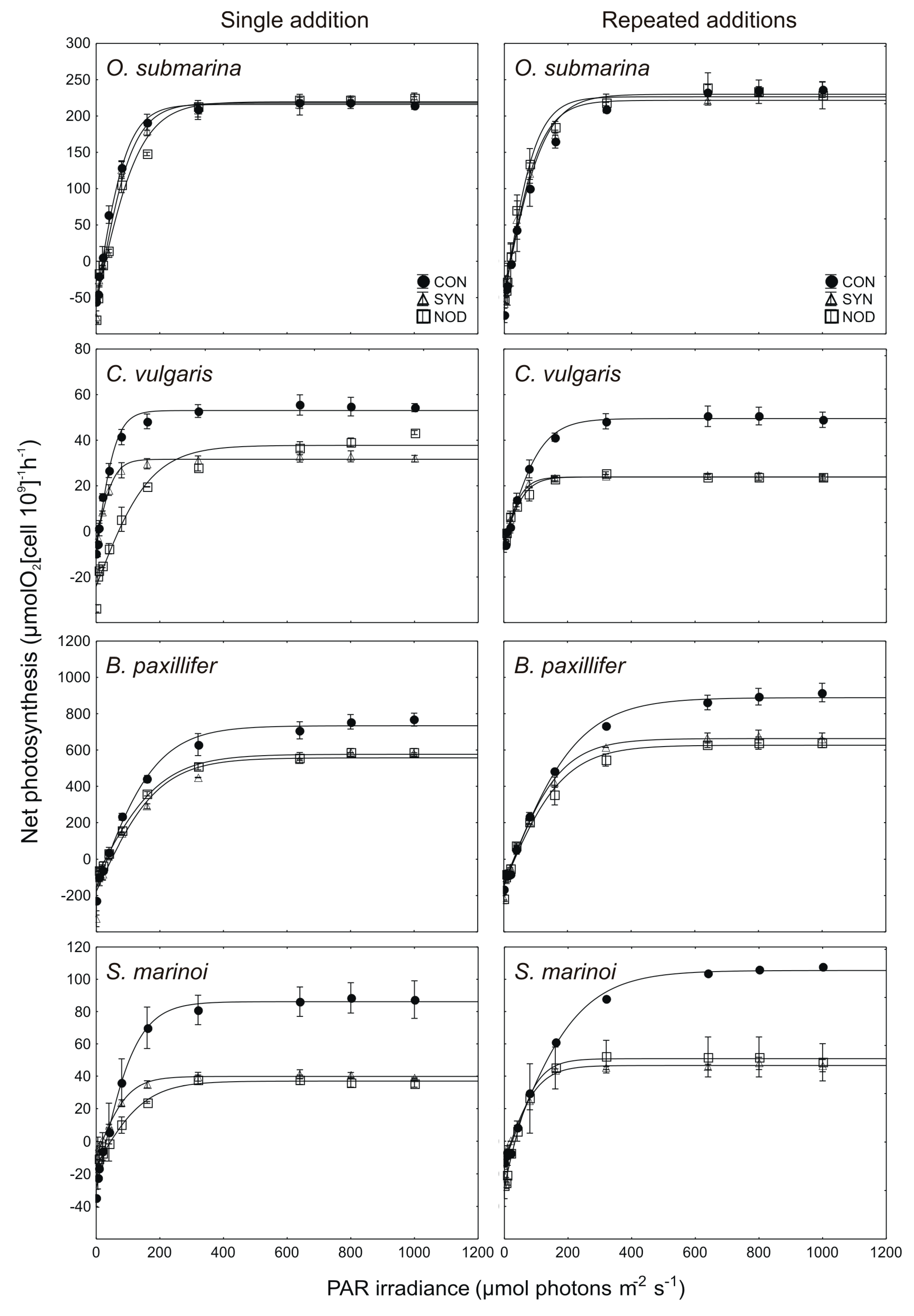
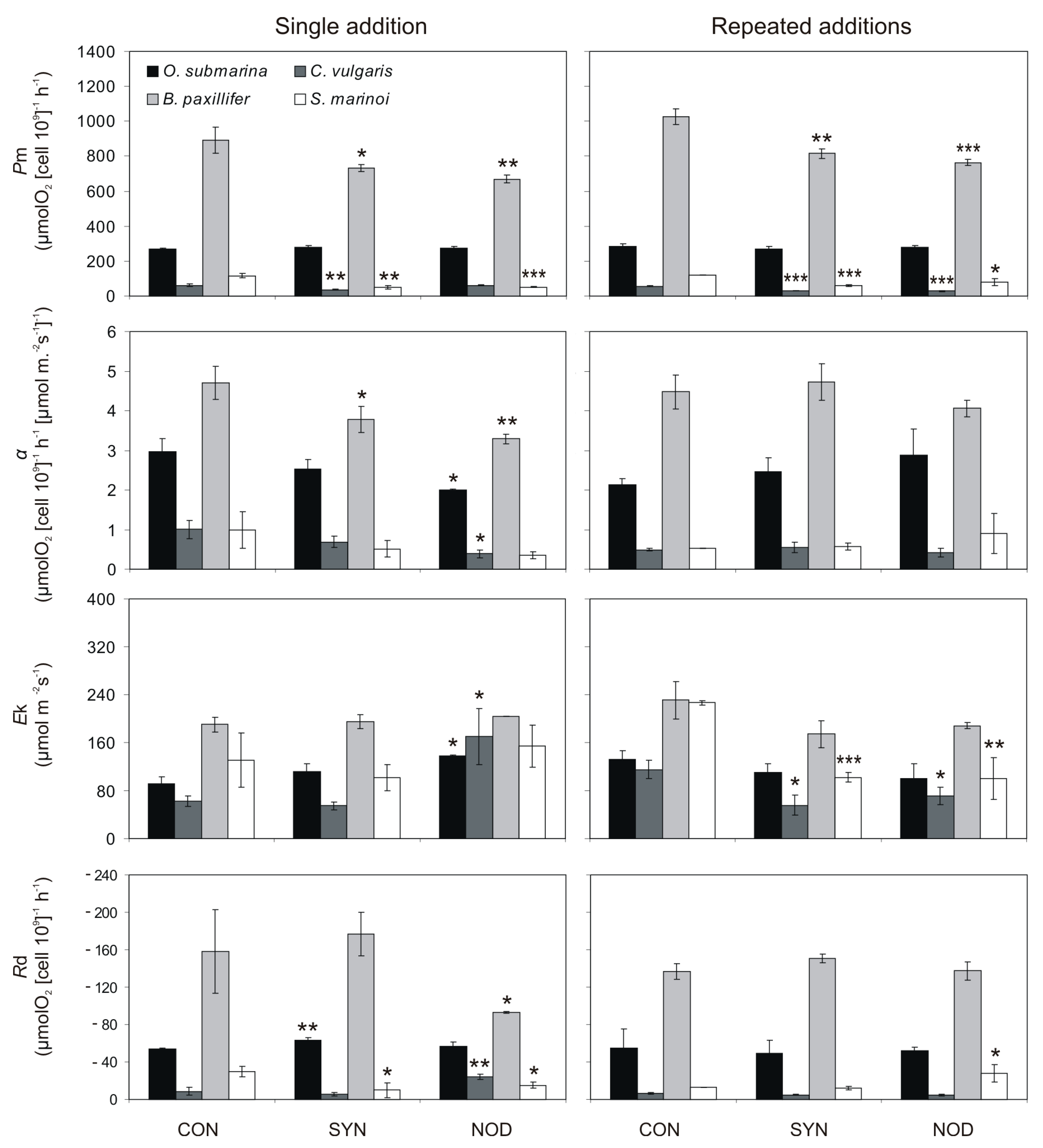
2.3. Allelopathic Effects of Cyanobacterial Exudates on P–E Curves and Photosynthetic Parameters
3. Discussion
3.1. Effect of the Selection of the Donor and Target Organisms
3.2. Effect of Single and Repeated Addition of Cyanobacterial Exudates
3.3. Comparison of Cyanobacterial Cell-Free Filtrate in Exponential and Stationary Growth Phases
3.4. Mode of Action of Allelochemicals
4. Materials and Methods
4.1. Culture Conditions
4.2. Cell-Free Filtrate Experiments
4.3. Examination of Cell Abundances
4.4. Examination of Cell Morphology
4.5. Measurements of the Rate of Photosynthesis
4.6. Statistical Analyses
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Pearson, L.; Mihali, T.; Moffitt, M.; Kellmann, R.; Neilan, B. On the chemistry, toxicology and genetics of the cyanobacterial toxins, microcystin, nodularin, saxitoxin and cylindrospermopsin. Mar. Drugs 2010, 8, 1650–1680. [Google Scholar] [CrossRef] [Green Version]
- Leão, P.N.; Engene, N.; Antunes, A.; Gerwick, W.H.; Vasconcelos, V. The chemical ecology of cyanobacteria. Nat. Prod. Rep. 2012, 29, 372–391. [Google Scholar] [CrossRef] [Green Version]
- Almeida, J.R.; Freitas, M.; Cruz, S.; Vasconcelos, V.; Cunha, I. Acetylcholinesterase in Biofouling Species: Characterization and Mode of Action of Cyanobacteria-Derived Antifouling Agents. Toxins 2015, 7, 2739–2756. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Poniedziałek, B.; Rzymski, P.; Kokociński, M. Cylindrospermopsin: Water-linked potential threat to human health in Europe. Environ. Toxicol. Pharmacol. 2012, 34, 651–660. [Google Scholar] [CrossRef] [PubMed]
- Poniedziałek, B.; Rzymski, P.; Kokociński, M.; Karczewski, J. Toxic potencies of metabolite(s) of non-cylindrospermopsin producing Cylindrospermopsis raciborskii isolated from temperate zone in human white cells. Chemosphere 2015, 120, 608–614. [Google Scholar] [CrossRef] [PubMed]
- Testai, E.; Scardala, S.; Vichi, S.; Buratti, F.M.; Funari, E. Risk to human health associated with the environmental occurrence of cyanobacterial neurotoxic alkaloids anatoxins and saxitoxins. Crit. Rev. Toxicol. 2016, 46, 385–419. [Google Scholar] [CrossRef]
- Kubanek, J.; Hicks, M.K.; Naar, J.; Villareal, T.A. Does the red tide dinoflagellate Karenia brevis use allelopathy to outcompete other phytoplankton? Limnol. Oceanogr. 2005, 50, 883–895. [Google Scholar] [CrossRef]
- Figueredo, C.C.; Giani, A.; Bird, D.F. Does allelopathy contribute to Cylindrospermopsis raciborskii (cyanobacteria) bloom occurrence and geographic expansion? J. Phycol. 2007, 43, 256–265. [Google Scholar] [CrossRef]
- Prince, E.K.; Myers, T.L.; Kubanek, J. Effects of harmful algal blooms on competitors: Allelopathic mechanisms of the red tide dinoflagellate Karenia brevis. Limnol. Oceanogr. 2008, 53, 531–541. [Google Scholar] [CrossRef]
- Hattenrath-Lehmann, T.K.; Gobler, C.J. Allelopathic inhibition of competing phytoplankton by North American strains of the toxic dinoflagellate, Alexandrium fundyense: Evidence from field experiments, laboratory experiments, and bloom events. Harmful Algae 2011, 11, 106–116. [Google Scholar] [CrossRef]
- Franzè, G.; Pierson, J.J.; Stoecker, D.K.; Lavrentyev, P. Diatom-produced allelochemicals trigger trophic cascades in the planktonic food web. Limnol. Oceanogr. 2017, 63, 1093–1108. [Google Scholar] [CrossRef] [Green Version]
- Rodríguez-Ramos, T.; Lorenzo, P.; González, L. Marine Allelopathy: Principles and perspectives. Thalassas 2007, 23, 39–49. [Google Scholar]
- Granéli, E.; Hansen, P.J. Allelopathy in Harmful Algae: A mechanism to compete for resources? In Ecology of Harmful Algae; Granéli, E., Turner, J., Eds.; Series: Ecological Studies 189; Springer: Berlin, Germany, 2006; pp. 189–201. [Google Scholar]
- Von Elert, E.; Jüttner, F. Phosphorus limitation and not light controls the extracellular release of allelopathic compounds by Trichormus doliolum (Cyanobacteria). Limnol. Oceanogr. 1997, 42, 1796–1802. [Google Scholar] [CrossRef] [Green Version]
- Smith, G.D.; Doan, D.T. Cyanobacterial metabolites with bioactivity against photosynthesis in cyanobacteria, algae and higher plants. J. Appl. Phycol. 1999, 11, 337–344. [Google Scholar] [CrossRef]
- Ma, Z.L.; Fang, T.X.; Thring, R.W.; Li, Y.B.; Yu, H.G.; Zhou, Q.; Zhao, M. Toxic and non-toxic strains of Microcystis aeruginosa induce temperature dependent allelopathy toward growth and photosynthesis of Chlorella vulgaris. Harmful Algae 2015, 48, 21–29. [Google Scholar] [CrossRef]
- Śliwińska-Wilczewska, S.; Pniewski, F.; Latała, A. Allelopathic activity of the picocyanobacterium Synechococcus sp. under varied light, temperature and salinity conditions. Int. Rev. Hydrobiol. 2016, 101, 69–77. [Google Scholar] [CrossRef]
- Gross, E.M. Allelopathy of aquatic autotrophs. Crit. Rev. Plant Sci. 2003, 22, 313–339. [Google Scholar] [CrossRef] [Green Version]
- Barreiro Felpeto, A.; Śliwińska-Wilczewska, S.; Złoch, I.; Vasconcelos, V. Light-dependent cytolysis in the allelopathic interaction between picoplanktic and filamentous cyanobacteria. J. Plankton Res. 2018, 40, 165–177. [Google Scholar] [CrossRef] [Green Version]
- Suikkanen, S.; Fistarol, G.O.; Granéli, E. Allelopathic effects of the Baltic cyanobacteria Nodularia spumigena, Aphanizomenon flos-aquae and Anabaena lemmermannii on algal monocultures. J. Exp. Mar. Biol. Ecol. 2004, 308, 85–101. [Google Scholar] [CrossRef]
- Barreiro, A.; Hairston, N.G., Jr. The influence of resource limitation on the allelopathic effect of Chlamydomonas reinhardtii on other unicellular freshwater planktonic organisms. J. Plankton Res. 2013, 35, 1339–1344. [Google Scholar] [CrossRef] [Green Version]
- Barreiro Felpeto, A.; Vasconcelos, V.M. Interactions between allelopathic properties and growth kynetics in four freshwater phytoplankton species studied by model simulations. Aquat. Ecol. 2014, 48, 191–205. [Google Scholar] [CrossRef]
- Barreiro Felpeto, A.; Roy, S.; Vasconcelos, V.M. Allelopathy prevents competitive exclusion and promotes phytoplankton biodiversity. Oikos 2017, 127, 85–98. [Google Scholar] [CrossRef] [Green Version]
- Antunes, J.T.; Leão, P.N.; Vasconcelos, V.M. Influence of Biotic and Abiotic Factors on the Allelopathic Activity of the Cyanobacterium Cylindrospermopsis raciborskii Strain LEGE 99043. Microb. Ecol. 2012, 64, 584–592. [Google Scholar] [CrossRef] [PubMed]
- Bittencourt-Oliveira, M.D.C.; Chia, M.A.; Oliveira, H.S.B.D.; Araújo, M.K.C.; Molica, R.J.R.; Dias, C.T.S. Allelopathic interactions between microcystin-producing and non-microcystin-producing cyanobacteria and green microalgae: Implications for microcystins production. J. Appl. Phycol. 2014, 27, 275–284. [Google Scholar] [CrossRef]
- Żak, A.; Kosakowska, A. The influence of extracellular compounds produced by selected Baltic cyanobacteria, diatoms and dinoflagellates on growth of green algae Chlorella vulgaris. Estuar. Coast. Shelf Sci. 2015, 167, 113–118. [Google Scholar] [CrossRef]
- Śliwińska-Wilczewska, S.; Latała, A. Allelopathic activity of the bloom-forming picocyanobacterium Synechococcus sp. on the coexisting microalgae: The role of eutrophication. Int. Rev. Hydrobiol. 2018, 103, 37–47. [Google Scholar] [CrossRef]
- Wang, L.; Zi, J.; Xu, R.; Hilt, S.; Hou, X.; Chang, X. Allelopathic effects of Microcystis aeruginosa on green algae and a diatom: Evidence from exudates addition and co-culturing. Harmful Algae 2017, 61, 56–62. [Google Scholar] [CrossRef]
- Suikkanen, S.; Laamanen, M.; Huttunen, M. Long-term changes in summer phytoplankton communities of the open northern Baltic Sea. Estuar. Coast. Shelf Sci. 2007, 71, 580–592. [Google Scholar] [CrossRef]
- Suikkanen, S.; Pulina, S.; Engström-Öst, J.; Lehtiniemi, M.; Lehtinen, S.; Brutemark, A. Climate Change and Eutrophication Induced Shifts in Northern Summer Plankton Communities. PLoS ONE 2013, 8, e66475. [Google Scholar] [CrossRef]
- Śliwińska-Wilczewska, S.; Maculewicz, J.; Tuszer, J.; Dobosz, K.; Kalusa, D.; Latała, A. First record of allelopathic activity of the picocyanobacterium Synechococcus sp. on a natural plankton community. Ecohydrol. Hydrobiol. 2017, 17, 227–234. [Google Scholar] [CrossRef]
- Lyczkowski, E.R.; Karp-Boss, L. Allelopathic effects of Alexandrium fundyense (Dinophyceae) on Thalassiosira cf. gravida (Bacillariophyceae): A matter of size. J. Phycol. 2014, 50, 376–387. [Google Scholar] [CrossRef] [PubMed]
- Fistarol, G.O.; Legrand, C.; Granéli, E. Allelopathic effect on a nutrient-limited phytoplankton species. Aquat. Microb. Ecol. 2005, 41, 153–161. [Google Scholar] [CrossRef] [Green Version]
- Śliwińska-Wilczewska, S.; Maculewicz, J.; Barreiro Felpeto, A.; Vasconcelos, V.; Latała, A. Allelopathic activity of the picocyanobacterium Synechococcus sp. on filamentous cyanobacteria. J. Exp. Mar. Biol. Ecol. 2017, 496, 16–21. [Google Scholar] [CrossRef]
- Christoffersen, K.; Lyck, S.; Winding, A. Microbial activity and bacterial community structure during degradation of microcystins. Aquat. Microb. Ecol. 2002, 27, 125–136. [Google Scholar] [CrossRef] [Green Version]
- Cole, J.J. Interactions between bacteria and algae in aquatic ecosystems. Annu. Rev. Ecol. Evol. Syst. 1982, 13, 291–314. [Google Scholar] [CrossRef]
- Schmidt, L.E.; Hansen, P.J. Allelopathy in the prymnesiophyte Chrysochromulina polylepis: Effect of cell concentration, growth phase and pH. Mar. Ecol. Prog. Ser. 2001, 216, 67–81. [Google Scholar] [CrossRef]
- Issa, A.A. Antibiotic production by the cyanobacteria Oscillatoria angustissima and Calothrix parietina. Environ. Toxicol. Pharmacol. 1999, 8, 33–37. [Google Scholar] [CrossRef]
- Arzul, G.; Seguel, M.; Guzman, L.; Denn, E.E. Comparison of allelopathic properties in three toxic Alexandrium species. J. Exp. Mar. Biol. Ecol. 1999, 232, 285–295. [Google Scholar] [CrossRef]
- Valdor, R.; Aboal, M. Effects of living cyanobacteria, cyanobacterial extracts and pure microcystins on growth and ultrastructure of microalgae and bacteria. Toxicon 2007, 49, 769–779. [Google Scholar] [CrossRef]
- Gantar, M.; Berry, J.P.; Thomas, S.; Wang, M.; Perez, R.; Rein, K.S. Allelopathic activity among cyanobacteria and microalgae isolated from Florida freshwater habitats. FEMS Microbiol. Ecol. 2008, 64, 55–64. [Google Scholar] [CrossRef] [Green Version]
- Pichierri, S.; Accoroni, S.; Pezzolesi, L.; Guerrini, F.; Romagnoli, T.; Pistocchi, R.; Totti, C. Allelopathic effects of diatom filtrates on the toxic benthic dinoflagellate Ostreopsis cf. ovata. Mar. Environ. Res. 2017, 131, 116–122. [Google Scholar] [CrossRef] [PubMed]
- Tillmann, U. Kill and eat your predator: A winning strategy of the marine flagellate Prymnesium parvum. Aquat. Microb. Ecol. 2003, 32, 73–84. [Google Scholar] [CrossRef] [Green Version]
- Tillmann, U.; John, U.; Cembella, A. On the allelochemical potency of the marine dinoflagellate Alexandrium ostenfeldi agains heterotrophic and autotrophic protists. J. Plankton Res. 2007, 29, 527–543. [Google Scholar] [CrossRef]
- Prézelin, B.B. Light reactions in photosynthesis. In Physiological Bases of Phytoplankton Ecology; Platt, T., Ed.; Canadian Journal of Fisheries and Aquatic Sciences, 210; Canadian Government Publishing: Ottawa, QC, Canada, 1981; pp. 1–46. [Google Scholar]
- Light, B.R.; Beardall, J. Photosynthetic characteristics of sub-tidal benthic microalgal populations from a temperate, shallow water marine ecosystem. Aquat. Bot. 2001, 70, 9–27. [Google Scholar] [CrossRef]
- Zhou, Y.H.; Yu, J.Q. Allelochemicals and photosynthesis. In Allelopathy: A Physiological Process with Ecological Implications; Reigosa, M.J., Pedrol, N., González, L., Eds.; Springer: Dordrecht, The Netherlands, 2006; pp. 127–139. [Google Scholar]
- Duke, S.O.; Dayan, F.E. Modes of action of phytotoxins from plants. In Allelopathy: A Physiological Process with Ecological Implications; Reigosa, M.J., Pedrol, N., González, L., Eds.; Springer: Dordrecht, The Netherlands, 2006; pp. 511–536. [Google Scholar]
- Guillard, R.R.L. Culture of phytoplankton for feeding marine invertebrates. In Culture of Marine Invertebrate Animals; Smith, W.L., Chanley, M.H., Eds.; Plenum Press: New York, NY, USA, 1975. [Google Scholar]
- Grasshoff, K. Methods of Seawater Analysis; Verlag Chemie Weinheim: New York, NY, USA, 1976. [Google Scholar]
- Guillard, R.R.; Sieracki, M.S. Counting cells in cultures with the light microscope. In Algal Culturing Techniques; Andersen, R.A., Ed.; Elsevier Academic Press: Burlington, MA, USA, 2005; pp. 239–252. [Google Scholar]
- Jassby, A.D.; Platt, T. Mathematical formulation of the relationship between photosynthesis and light for phytoplankton. Limnol. Oceanogr. 1976, 21, 540–547. [Google Scholar] [CrossRef] [Green Version]
- Sakshaug, E.; Bricaud, A.; Dandonneau, Y.; Falkowski, P.G.; Kiefer, D.A.; Legendre, L.L.; Morel, A.; Parslow, J.; Takahashi, M. Parameters of photosynthesis: Definitions, theory and interpretation of results. J. Plankton Res. 1997, 19, 1637–1670. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Response Variable (Number of Cells) | Source of Variation | Df | Ss | Mss | F | Ss | Mss | F |
|---|---|---|---|---|---|---|---|---|
| Synechococcus sp. | N. spumigena | |||||||
| Single Additions of Cell-Free Filtrate | ||||||||
| O. submarina | filtrate | 1 | 0.184 | 0.184 | 2.896 | 0.163 | 0.163 | 4.595 * |
| time | 3 | 1039.170 | 346.390 | 5459.262 *** | 1037.309 | 345.770 | 9726.294 *** | |
| filtrate time * | 3 | 0.135 | 0.045 | 0.708 | 0.543 | 0.181 | 5.090 * | |
| error | 16 | 1.015 | 0.063 | 0.569 | 0.036 | |||
| C. vulgaris | filtrate | 1 | 17.839 | 17.839 | 138.156 *** | 119.410 | 119.410 | 721.685 *** |
| time | 3 | 535.876 | 178.625 | 1383.420 *** | 242.597 | 80.866 | 488.730 *** | |
| filtrate time * | 3 | 20.064 | 6.688 | 51.798 *** | 144.634 | 48.211 | 291.376 *** | |
| error | 16 | 2.066 | 0.129 | 2.647 | 0.165 | |||
| B. paxillifer | filtrate | 1 | 0.813 | 0.813 | 6.688 * | 1.216 | 1.216 | 13.770 ** |
| time | 3 | 129.403 | 43.134 | 355.008 *** | 119.790 | 39.930 | 452.322 *** | |
| filtrate time * | 3 | 0.409 | 0.136 | 1.123 | 1.346 | 0.449 | 5.083 * | |
| error | 16 | 1.944 | 0.122 | 1.412 | 0.088 | |||
| S. marinoi | filtrate | 1 | 10.588 | 10.588 | 1341.763 *** | 11.724 | 11.724 | 1517.087 *** |
| time | 3 | 22.632 | 7.544 | 955.952 *** | 31.078 | 10.359 | 1340.444 *** | |
| filtrate time * | 3 | 15.964 | 5.321 | 674.305 *** | 10.315 | 3.438 | 444.896 *** | |
| error | 16 | 0.126 | 0.008 | 0.124 | 0.008 | |||
| Repeated Additions of Cell-Free Filtrate | ||||||||
| O. submarina | filtrate | 1 | 0.029 | 0.029 | 0.387 | 1.500 | 1.500 | 26.882 *** |
| time | 3 | 204.989 | 68.330 | 898.485 *** | 170.116 | 56.705 | 1016.222 *** | |
| filtrate time * | 3 | 0.025 | 0.008 | 0.108 | 1.844 | 0.615 | 11.018 *** | |
| error | 16 | 1.217 | 0.076 | 0.893 | 0.056 | |||
| C. vulgaris | filtrate | 1 | 34.016 | 34.016 | 56.588 *** | 19.960 | 19.960 | 238.473 *** |
| time | 3 | 109.672 | 36.557 | 60.816 *** | 178.845 | 59.615 | 712.265 *** | |
| filtrate time * | 3 | 45.661 | 15.220 | 25.320 *** | 17.262 | 5.754 | 68.748 *** | |
| error | 16 | 9.618 | 0.601 | 1.339 | 0.084 | |||
| B. paxillifer | filtrate | 1 | 1.096 | 1.096 | 8.723 ** | 1.456 | 1.456 | 10.204 ** |
| time | 3 | 85.712 | 28.571 | 227.319 *** | 86.871 | 28.957 | 202.981 *** | |
| filtrate time * | 3 | 2.009 | 0.670 | 5.328 * | 1.764 | 0.588 | 4.121 * | |
| error | 16 | 2.011 | 0.126 | 2.283 | 0.143 | |||
| S. marinoi | filtrate | 1 | 5.750 | 5.750 | 2175.294 *** | 7.915 | 7.915 | 3161.742 *** |
| time | 3 | 11.730 | 3.910 | 1479.173 *** | 8.253 | 2.751 | 1098.877 *** | |
| filtrate time * | 3 | 5.618 | 1.873 | 708.431*** | 9.551 | 3.184 | 1271.713 *** | |
| error | 16 | 0.042 | 0.003 | 0.040 | 0.003 | |||
| Target Species | Day | Number of Cells (N) | % of Control | Source of Variation | Df | Ss | Mss | F |
|---|---|---|---|---|---|---|---|---|
| Synechococcus sp. | ||||||||
| O. submarina | 0 | 0.619 ± 0.007 | 100.3 ± 1.5 | filtrate | 1 | 0.004 | 0.004 | 5.045 * |
| 1 | 0.649 ± 0.015 | 98.2 ± 1.6 | time | 3 | 1.571 | 0.524 | 651.147 *** | |
| 3 | 0.815 ± 0.032 | 97.1 ± 2.5 | filtrate time * | 3 | 0.004 | 0.001 | 1.816 | |
| 7 | 1.227 ± 0.054 | 94.6 ± 1.9 | error | 16 | 0.013 | 0.001 | ||
| C. vulgaris | 0 | 2.635 ± 0.098 | 99.9 ± 9.6 | filtrate | 1 | 2.885 | 2.885 | 15.230 ** |
| 1 | 2.545 ± 0.061 | 96.4 ± 12.3 | time | 3 | 78.566 | 26.189 | 138.273 *** | |
| 3 | 3.692 ± 0.205 | 88.4 ± 5.4 | filtrate time * | 3 | 4.282 | 1.427 | 7.536 ** | |
| 7 | 5.994 ± 0.247 | 74.7 ± 11.3 | error | 16 | 3.030 | 0.189 | ||
| B. paxillifer | 0 | 0.267 ± 0.012 | 99.1 ± 2.7 | filtrate | 1 | 0.001 | 0.001 | 1.809 |
| 1 | 0.254 ± 0.002 | 87.9 ± 2.3 | time | 3 | 0.312 | 0.104 | 359.793 *** | |
| 3 | 0.405 ± 0.022 | 101.1 ± 3.6 | filtrate time * | 3 | 0.001 | 0.000 | 1.605 | |
| 7 | 0.543 ± 0.036 | 99.1 ± 5.0 | error | 16 | 0.005 | 0.000 | ||
| S. marinoi | 0 | 1.379 ± 0.063 | 99.8 ± 2.4 | filtrate | 1 | 0.081 | 0.081 | 4.915 * |
| 1 | 1.757 ± 0.099 | 97.4 ± 4.2 | time | 3 | 68.187 | 22.729 | 1380.932 *** | |
| 3 | 2.822 ± 0.062 | 98.6 ± 4.6 | filtrate time * | 3 | 0.129 | 0.043 | 2.621 | |
| 7 | 5.508 ± 0.205 | 93.7 ± 2.4 | error | 16 | 0.263 | 0.016 | ||
| N. spumigena | ||||||||
| O. submarina | 0 | 0.621 ± 0.009 | 100.7 ± 1.1 | filtrate | 1 | 0.002 | 0.002 | 2.496 |
| 1 | 0.661 ± 0.009 | 100.0 ± 0.9 | time | 3 | 1.623 | 0.541 | 562.660 *** | |
| 3 | 0.799 ± 0.030 | 95.2 ± 2.3 | filtrate time * | 3 | 0.003 | 0.001 | 1.015 | |
| 7 | 1.253 ± 0.067 | 96.6 ± 3.5 | error | 16 | 0.015 | 0.001 | ||
| C. vulgaris | 0 | 1.903 ± 0.073 | 94.6 ± 8.5 | filtrate | 1 | 1.157 | 1.157 | 10.760 ** |
| 1 | 2.179 ± 0.028 | 90.3 ± 13.9 | time | 3 | 32.580 | 10.860 | 100.966 *** | |
| 3 | 2.391 ± 0.112 | 91.9 ± 4.4 | filtrate time * | 3 | 1.041 | 0.347 | 3.226 | |
| 7 | 4.335 ± 0.609 | 79.8 ± 16.2 | error | 16 | 1.721 | 0.108 | ||
| B. paxillifer | 0 | 0.273 ± 0.012 | 101.2 ± 2.5 | filtrate | 1 | 0.003 | 0.003 | 13.622 ** |
| 1 | 0.254 ± 0.005 | 87.9 ± 1.5 | time | 3 | 0.262 | 0.087 | 350.093 *** | |
| 3 | 0.386 ± 0.026 | 96.3 ± 4.0 | filtrate time * | 3 | 0.002 | 0.001 | 3.151 | |
| 7 | 0.499 ± 0.027 | 91.1 ± 3.6 | error | 16 | 0.004 | 0.000 | ||
| S. marinoi | 0 | 1.083 ± 0.016 | 104.0 ± 1.4 | filtrate | 1 | 0.086 | 0.086 | 9.304 ** |
| 1 | 1.192 ± 0.016 | 100.0 ± 0.0 | time | 3 | 8.020 | 2.673 | 290.170 *** | |
| 3 | 1.825 ± 0.008 | 95.0 ± 0.0 | filtrate time * | 3 | 0.199 | 0.066 | 7.218 ** | |
| 7 | 2.294 ± 0.083 | 84.7 ± 5.4 | error | 16 | 0.147 | 0.009 | ||
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Śliwińska-Wilczewska, S.; Barreiro Felpeto, A.; Możdżeń, K.; Vasconcelos, V.; Latała, A. Physiological Effects on Coexisting Microalgae of the Allelochemicals Produced by the Bloom-Forming Cyanobacteria Synechococcus sp. and Nodularia Spumigena. Toxins 2019, 11, 712. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins11120712
Śliwińska-Wilczewska S, Barreiro Felpeto A, Możdżeń K, Vasconcelos V, Latała A. Physiological Effects on Coexisting Microalgae of the Allelochemicals Produced by the Bloom-Forming Cyanobacteria Synechococcus sp. and Nodularia Spumigena. Toxins. 2019; 11(12):712. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins11120712
Chicago/Turabian StyleŚliwińska-Wilczewska, Sylwia, Aldo Barreiro Felpeto, Katarzyna Możdżeń, Vitor Vasconcelos, and Adam Latała. 2019. "Physiological Effects on Coexisting Microalgae of the Allelochemicals Produced by the Bloom-Forming Cyanobacteria Synechococcus sp. and Nodularia Spumigena" Toxins 11, no. 12: 712. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins11120712