The Beneficial and Debilitating Effects of Environmental and Microbial Toxins, Drugs, Organic Solvents and Heavy Metals on the Onset and Progression of Multiple Sclerosis



Abstract
:1. Introduction
2. Involvement of the Immune System in MS Disease
3. Toxic Effects of Environmental and Geographical Factors
4. Toxic Effects of Lifestyle Habits
5. Toxic Effects of Food, Diet and Gut Microbiota
6. Toxic Effects of Microbes
7. Toxicity of Chemicals, Organic Solvents and Heavy Metals
8. Efficacy of Various Drugs in MS Patients
9. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Mahad, D.H.; Trapp, B.D.; Lassmann, H. Pathological mechanisms in progressive multiple sclerosis. Lancet Neurol. 2015, 14, 183–193. [Google Scholar] [CrossRef]
- De Souza, J.M.; Goncalves, B.D.C.; Gomez, M.V.; Vieira, L.B.; Ribeiro, F.M. Animal toxins as therapeutic tools to treat neurodegenerative diseases. Front. Pharmacol. 2018, 9, 145. [Google Scholar] [CrossRef] [PubMed]
- Zeller, D.; Classen, J. Plasticity of the motor system in multiple sclerosis. Neuroscience 2014, 283, 222–230. [Google Scholar] [CrossRef] [PubMed]
- Browne, P.; Chandraratna, D.; Angood, C.; Tremlett, H.; Baker, C.; Taylor, B.V.; Thompson, A.J. Atlas of multiple sclerosis 2013: A growing global problem with widespread inequity. Neurology 2014, 83, 1022–1024. [Google Scholar] [CrossRef] [PubMed]
- Dilokthornsakul, P.; Valuck, R.J.; Nair, K.V.; Corboy, J.R.; Allen, R.R.; Campbell, J.D. Multiple sclerosis prevalence in the united states commercially insured population. Neurology 2016, 86, 1014–1021. [Google Scholar] [CrossRef] [PubMed]
- Dendrou, C.A.; Fugger, L.; Friese, M.A. Immunopathology of multiple sclerosis. Nat. Rev. Immunol. 2015, 15, 545. [Google Scholar] [CrossRef] [PubMed]
- Adamczyk, B.; Adamczyk-Sowa, M. New insights into the role of oxidative stress mechanisms in the pathophysiology and treatment of multiple sclerosis. Oxid. Med. Cell Longev. 2016, 2016, 18. [Google Scholar] [CrossRef] [PubMed]
- Lassmann, H.; Bradl, M. Multiple sclerosis: Experimental models and reality. Acta Neuropathol. 2017, 133, 223–244. [Google Scholar] [CrossRef] [PubMed]
- Comi, G.; Radaelli, M.; Soelberg Sørensen, P. Evolving concepts in the treatment of relapsing multiple sclerosis. Lancet 2017, 389, 1347–1356. [Google Scholar] [CrossRef]
- Murray, T.J. The history of multiple sclerosis: The changing frame of the disease over the centuries. J. Neurol. Sci. 2009, 277, S3–S8. [Google Scholar] [CrossRef]
- Gu, C. Kir4.1: K(+) channel illusion or reality in the autoimmune pathogenesis of multiple sclerosis. Front. Mol. Neurosci. 2016, 9, 90. [Google Scholar] [CrossRef] [PubMed]
- Høglund, R.A.; Maghazachi, A.A. Multiple sclerosis and the role of immune cells. World J. Exp. Med. 2014, 4, 27–37. [Google Scholar] [CrossRef] [PubMed]
- Maghazachi, A.A. On the role of natural killer cells in neurodegenerative diseases. Toxins (Basel) 2013, 5, 363–375. [Google Scholar] [CrossRef] [PubMed]
- Jones, A.P.; Kermode, A.G.; Lucas, R.M.; Carroll, W.M.; Nolan, D.; Hart, P.H. Circulating immune cells in multiple sclerosis. Clin. Exp. Immunol. 2016, 187, 193–203. [Google Scholar] [CrossRef] [PubMed]
- Jadidi-Niaragh, F.; Mirshafiey, A. Th17 cell, the new player of neuroinflammatory process in multiple sclerosis. Scand. J. Immunol. 2011, 74, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Volpe, E.; Sambucci, M.; Battistini, L.; Borsellino, G. Fas–fas ligand: Checkpoint of t cell functions in multiple sclerosis. Front. Immunol. 2016, 7, 382. [Google Scholar] [CrossRef] [PubMed]
- Elyaman, W.; Khoury, S.J. Th9 cells in the pathogenesis of eae and multiple sclerosis. Semin. Immunopathol. 2017, 39, 79–87. [Google Scholar] [CrossRef] [PubMed]
- Fard, N.A.; Azizi, G.; Mirshafiey, A. The potential role of t helper cell 22 and il-22 in immunopathogenesis of multiple sclerosis. Innov. Clin. Neurosci. 2016, 13, 30–36. [Google Scholar] [PubMed]
- Hestvik, A.L.K. The double-edged sword of autoimmunity: Lessons from multiple sclerosis. Toxins (Basel) 2010, 2, 856–877. [Google Scholar] [CrossRef] [PubMed]
- Garg, N.; Smith, T.W. An update on immunopathogenesis, diagnosis, and treatment of multiple sclerosis. Brain Behav. 2015, 5, e00362. [Google Scholar] [CrossRef] [PubMed]
- Huynh, J.L.; Casaccia, P. Epigenetic mechanisms in multiple sclerosis: Implications for pathogenesis and treatment. Lancet Neurol. 2013, 12, 195–206. [Google Scholar] [CrossRef]
- Olsson, T.; Barcellos, L.F.; Alfredsson, L. Interactions between genetic, lifestyle and environmental risk factors for multiple sclerosis. Nat. Rev. Neurol. 2016, 13, 25. [Google Scholar] [CrossRef] [PubMed]
- Riemann-Lorenz, K.; Eilers, M.; von Geldern, G.; Schulz, K.-H.; Köpke, S.; Heesen, C. Dietary interventions in multiple sclerosis: Development and pilot-testing of an evidence based patient education program. PLoS ONE 2016, 11, e0165246. [Google Scholar] [CrossRef] [PubMed]
- Peedicayil, J. Epigenetic drugs for multiple sclerosis. Curr. Neuropharmacol. 2016, 14, 3–9. [Google Scholar] [CrossRef] [PubMed]
- Babenko, O.; Kovalchuk, I.; Metz, G.A. Epigenetic programming of neurodegenerative diseases by an adverse environment. Brain Res. 2012, 1444, 96–111. [Google Scholar] [CrossRef] [PubMed]
- Koch, M.W.; Metz, L.M.; Kovalchuk, O. Epigenetic changes in patients with multiple sclerosis. Nat. Rev. Neurol. 2012, 9, 35. [Google Scholar] [CrossRef] [PubMed]
- Miller, F.W.; Alfredsson, L.; Costenbader, K.H.; Kamen, D.L.; Nelson, L.M.; Norris, J.M.; De Roos, A.J. Epidemiology of environmental exposures and human autoimmune diseases: Findings from a national institute of environmental health sciences expert panel workshop. J. Autoimmun. 2012, 39, 259–271. [Google Scholar] [CrossRef] [PubMed]
- Hedström, A.K.; Alfredsson, L.; Olsson, T. Environmental factors and their interactions with risk genotypes in ms susceptibility. Curr. Opin. Neurol. 2016, 29, 293–298. [Google Scholar] [CrossRef] [PubMed]
- Kakalacheva, K.; Lünemann, J.D. Environmental triggers of multiple sclerosis. FEBS Lett. 2011, 585, 3724–3729. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Loken-Amsrud, K.I.; Lossius, A.; Torkildsen, O.; Holmoy, T. Impact of the environment on multiple sclerosis. Tidsskr. Nor. Laegeforen. 2015, 135, 856–860. [Google Scholar] [PubMed]
- Milo, R.; Kahana, E. Multiple sclerosis: Geoepidemiology, genetics and the environment. Autoimmun. Rev. 2010, 9, A387–A394. [Google Scholar] [CrossRef] [PubMed]
- Turabelidze, G.; Schootman, M.; Zhu, B.P.; Malone, J.L.; Horowitz, S.; Weidinger, J.; Williamson, D.; Simoes, E. Multiple sclerosis prevalence and possible lead exposure. J. Neurol. Sci. 2008, 269, 158–162. [Google Scholar] [CrossRef] [PubMed]
- Helmick, C.G.; Wrigley, J.M.; Zack, M.M.; Bigler, W.J.; Lehman, J.L.; Janssen, R.S.; Hartwig, E.C.; Witte, J.J. Multiple sclerosis in key west, florida. Am. J. Epidemiol. 1989, 130, 935–949. [Google Scholar] [CrossRef] [PubMed]
- Forbes, R.B.; Wilson, S.V.; Swingler, R.J. The prevalence of multiple sclerosis in tayside, scotland: Do latitudinal gradients really exist? J. Neurol. 1999, 246, 1033–1040. [Google Scholar] [CrossRef] [PubMed]
- Gray, O.M.; McDonnell, G.V.; Hawkins, S.A. Factors in the rising prevalence of multiple sclerosis in the north-east of ireland. Mult. Scler. 2008, 14, 880–886. [Google Scholar] [CrossRef] [PubMed]
- Beck, C.A.; Metz, L.M.; Svenson, L.W.; Patten, S.B. Regional variation of multiple sclerosis prevalence in canada. Mult. Scler. 2005, 11, 516–519. [Google Scholar] [CrossRef] [PubMed]
- McGuigan, C.; McCarthy, A.; Quigley, C.; Bannan, L.; Hawkins, S.; Hutchinson, M. Latitudinal variation in the prevalence of multiple sclerosis in ireland, an effect of genetic diversity. J. Neurol. Neurosurg. Psychiatry 2004, 75, 572–576. [Google Scholar] [CrossRef] [PubMed]
- Lucas, R.M.; Byrne, S.N.; Correale, J.; Ilschner, S.; Hart, P.H. Ultraviolet radiation, vitamin d and multiple sclerosis. Neurodegener. Dis. Manag. 2015, 5, 413–424. [Google Scholar] [CrossRef] [PubMed]
- Alharbi, F.M. Update in vitamin d and multiple sclerosis. Neurosciences (Riyadh) 2015, 20, 329–335. [Google Scholar] [CrossRef] [PubMed]
- Niino, M.; Sato, S.; Fukazawa, T.; Masaki, K.; Miyazaki, Y.; Matsuse, D.; Yamasaki, R.; Takahashi, E.; Kikuchi, S.; Kira, J. Decreased serum vitamin d levels in japanese patients with multiple sclerosis. J. Neuroimmunol. 2015, 279, 40–45. [Google Scholar] [CrossRef] [PubMed]
- Munger, K.L.; Zhang, S.M.; O’Reilly, E.; Hernan, M.A.; Olek, M.J.; Willett, W.C.; Ascherio, A. Vitamin d intake and incidence of multiple sclerosis. Neurology 2004, 62, 60–65. [Google Scholar] [CrossRef] [PubMed]
- Al-Jaderi, Z.; Maghazachi, A.A. Vitamin d3 and monomethyl fumarate enhance natural killer cell lysis of dendritic cells and ameliorate the clinical score in mice suffering from experimental autoimmune encephalomyelitis. Toxins (Basel) 2015, 7, 4730–4744. [Google Scholar] [CrossRef] [PubMed]
- Nielsen, N.M.; Munger, K.L.; Koch-Henriksen, N.; Hougaard, D.M.; Magyari, M.; Jorgensen, K.T.; Lundqvist, M.; Simonsen, J.; Jess, T.; Cohen, A.; et al. Neonatal vitamin d status and risk of multiple sclerosis: A population-based case-control study. Neurology 2017, 88, 44–51. [Google Scholar] [CrossRef] [PubMed]
- Fernandes de Abreu, D.A.; Babron, M.C.; Rebeix, I.; Fontenille, C.; Yaouanq, J.; Brassat, D.; Fontaine, B.; Clerget-Darpoux, F.; Jehan, F.; Feron, F. Season of birth and not vitamin d receptor promoter polymorphisms is a risk factor for multiple sclerosis. Mult. Scler. 2009, 15, 1146–1152. [Google Scholar] [CrossRef] [PubMed]
- Saccone, D.; Asani, F.; Bornman, L. Regulation of the vitamin d receptor gene by environment, genetics and epigenetics. Gene 2015, 561, 171–180. [Google Scholar] [CrossRef] [PubMed]
- Rinaldi, A.O.; Sanseverino, I.; Purificato, C.; Cortese, A.; Mechelli, R.; Francisci, S.; Salvetti, M.; Millefiorini, E.; Gessani, S.; Gauzzi, M.C. Increased circulating levels of vitamin d binding protein in ms patients. Toxins (Basel) 2015, 7, 129–137. [Google Scholar] [CrossRef] [PubMed]
- Hutter, C.D.; Laing, P. Multiple sclerosis: Sunlight, diet, immunology and aetiology. Med. Hypotheses 1996, 46, 67–74. [Google Scholar] [PubMed]
- Hedstrom, A.K.; Akerstedt, T.; Hillert, J.; Olsson, T.; Alfredsson, L. Shift work at young age is associated with increased risk for multiple sclerosis. Ann. Neurol. 2011, 70, 733–741. [Google Scholar] [CrossRef] [PubMed]
- Ponsonby, A.-L.; Lucas, R.M. Shift work and multiple sclerosis. Ann. Neurol. 2011, 70, 680–683. [Google Scholar] [CrossRef] [PubMed]
- Gustavsen, S.; Sondergaard, H.B.; Oturai, D.B.; Laursen, B.; Laursen, J.H.; Magyari, M.; Ullum, H.; Larsen, M.H.; Sellebjerg, F.; Oturai, A.B. Shift work at young age is associated with increased risk of multiple sclerosis in a danish population. Mult. Scler. Relat. Disord. 2016, 9, 104–109. [Google Scholar] [CrossRef] [PubMed]
- Hedstrom, A.K.; Akerstedt, T.; Olsson, T.; Alfredsson, L. Shift work influences multiple sclerosis risk. Mult. Scler. 2015, 21, 1195–1199. [Google Scholar] [CrossRef] [PubMed]
- Zhang, P.; Wang, R.; Li, Z.; Wang, Y.; Gao, C.; Lv, X.; Song, Y.; Li, B. The risk of smoking on multiple sclerosis: A meta-analysis based on 20,626 cases from case-control and cohort studies. PeerJ 2016, 4, e1797. [Google Scholar] [CrossRef] [PubMed]
- Turner, A.P.; Hartoonian, N.; Maynard, C.; Leipertz, S.L.; Haselkorn, J.K. Smoking and physical activity: Examining health behaviors and 15-year mortality among individuals with multiple sclerosis. Arch. Phys. Med. Rehabil. 2015, 96, 402–409. [Google Scholar] [CrossRef] [PubMed]
- Ramanujam, R.; Hedström, A.; Manouchehrinia, A.; Alfredsson, L.; Olsson, T.; Bottai, M.; Hillert, J. Effect of smoking cessation on multiple sclerosis prognosis. JAMA Neurol. 2015, 72, 1117–1123. [Google Scholar] [CrossRef] [PubMed]
- Öckinger, J.; Hagemann-Jensen, M.; Kullberg, S.; Engvall, B.; Eklund, A.; Grunewald, J.; Piehl, F.; Olsson, T.; Wahlström, J. T-cell activation and hla-regulated response to smoking in the deep airways of patients with multiple sclerosis. Clin. Immunol. 2016, 169, 114–120. [Google Scholar] [CrossRef] [PubMed]
- Ascherio, A.; Munger, K.L. Environmental risk factors for multiple sclerosis. Part ii: Noninfectious factors. Ann. Neurol. 2007, 61, 504–513. [Google Scholar] [CrossRef] [PubMed]
- Hedström, A.K.; Bomfim, I.L.; Barcellos, L.F.; Briggs, F.; Schaefer, C.; Kockum, I.; Olsson, T.; Alfredsson, L. Interaction between passive smoking and two hla genes with regard to multiple sclerosis risk. Int. J. Epidemiol. 2014, 43, 1791–1798. [Google Scholar] [CrossRef] [PubMed]
- Timmermans, S.; Bogie, J.F.J.; Vanmierlo, T.; Lütjohann, D.; Stinissen, P.; Hellings, N.; Hendriks, J.J.A. High fat diet exacerbates neuroinflammation in an animal model of multiple sclerosis by activation of the renin angiotensin system. J. Neuroimmune Pharmacol. 2014, 9, 209–217. [Google Scholar] [CrossRef] [PubMed]
- Farinotti, M.; Vacchi, L.; Simi, S.; Di Pietrantonj, C.; Brait, L.; Filippini, G. Dietary interventions for multiple sclerosis. Cochrane Database Syst. Rev. 2012, 12, CD004192. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.Y.; Hao, J.; Liu, R.; Turner, G.; Shi, F.-D.; Rho, J.M. Inflammation-mediated memory dysfunction and effects of a ketogenic diet in a murine model of multiple sclerosis. PLoS ONE 2012, 7, e35476. [Google Scholar] [CrossRef] [PubMed]
- Hedström, A.K.; Mowry, E.M.; Gianfrancesco, M.A.; Shao, X.; Schaefer, C.A.; Shen, L.; Olsson, T.; Barcellos, L.F.; Alfredsson, L. High consumption of coffee is associated with decreased multiple sclerosis risk; results from two independent studies. J. Neurol. Neurosurg. Psychiatry 2016, 87, 454–460. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mowry, E.; Hedstrom, A.; Gianfrancesco, M.; Shao, X.; Schaefer, C.; Barcellos, L.; Olsson, T.; Alfredsson, L. Greater consumption of coffee is associated with reduced odds of multiple sclerosis (s45.004). Neurology 2015, 84, S45.004. [Google Scholar]
- D’hooghe, M.B.; Haentjens, P.; Nagels, G.; De Keyser, J. Alcohol, coffee, fish, smoking and disease progression in multiple sclerosis. Eur. J. Neurol. 2011, 19, 616–624. [Google Scholar] [CrossRef] [PubMed]
- Farez, M.F.; Fiol, M.P.; Gaitán, M.I.; Quintana, F.J.; Correale, J. Sodium intake is associated with increased disease activity in multiple sclerosis. J. Neurol. Neurosurg. Psychiatry 2015, 86, 26. [Google Scholar] [CrossRef] [PubMed]
- Riccio, P.; Rossano, R. Nutrition facts in multiple sclerosis. ASN Neuro 2015, 7. [Google Scholar] [CrossRef] [PubMed]
- Mielcarz, D.W.; Kasper, L.H. The gut microbiome in multiple sclerosis. Curr. Treat. Opt. Neurol. 2015, 17, 18. [Google Scholar] [CrossRef] [PubMed]
- Kirby, T.O.; Ochoa-Repáraz, J. The gut microbiome in multiple sclerosis: A potential therapeutic avenue. Med. Sci. (Basel, Switzerland) 2018, 6, 69. [Google Scholar] [CrossRef] [PubMed]
- Chu, F.; Shi, M.; Lang, Y.; Shen, D.; Jin, T.; Zhu, J.; Cui, L. Gut microbiota in multiple sclerosis and experimental autoimmune encephalomyelitis: Current applications and future perspectives. Med. Inflamm. 2018, 2018, 8168717. [Google Scholar] [CrossRef] [PubMed]
- Jangi, S.; Gandhi, R.; Cox, L.M.; Li, N.; von Glehn, F.; Yan, R.; Patel, B.; Mazzola, M.A.; Liu, S.; Glanz, B.L.; et al. Alterations of the human gut microbiome in multiple sclerosis. Nat. Commun. 2016, 7, 12015. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roy Sarkar, S.; Banerjee, S. Gut microbiota in neurodegenerative disorders. J. Neuroimmunol. 2019, 328, 98–104. [Google Scholar] [CrossRef] [PubMed]
- Yokote, H.; Miyake, S.; Croxford, J.L.; Oki, S.; Mizusawa, H.; Yamamura, T. Nkt cell-dependent amelioration of a mouse model of multiple sclerosis by altering gut flora. Am. J. Pathol. 2008, 173, 1714–1723. [Google Scholar] [CrossRef] [PubMed]
- Brewer, J.H.; Thrasher, J.D.; Straus, D.C.; Madison, R.A.; Hooper, D. Detection of mycotoxins in patients with chronic fatigue syndrome. Toxins (Basel) 2013, 5, 605–617. [Google Scholar] [CrossRef] [PubMed]
- Ordog, G. 476 multiple sclerosis cluster: Mycotoxic leukoencephalopathy. J. Investig. Med. 2005, 53, S161. [Google Scholar] [CrossRef]
- Venkatesan, A. Multiple sclerosis and infections. Neurodegener. Dis. Manag. 2015, 5, 11–14. [Google Scholar] [CrossRef] [PubMed]
- Oleszak, E.L.; Chang, J.R.; Friedman, H.; Katsetos, C.D.; Platsoucas, C.D. Theiler’s virus infection: A model for multiple sclerosis. Clin. Microbiol. Rev. 2004, 17, 174–207. [Google Scholar] [CrossRef] [PubMed]
- Fong, I.W. The Role of Microbes in Common Non-Infectious Diseases; Springer: New York, NY, USA, 2014. [Google Scholar]
- Burns, J.; Littlefield, K.; Gill, J.; Trotter, J.L. Bacterial toxin superantigens activate human t lymphocytes reactive with myelin autoantigens. Ann. Neurol. 1992, 32, 352–357. [Google Scholar] [CrossRef] [PubMed]
- Gay, F. Bacterial toxins and multiple sclerosis. J. Neurol. Sci. 2007, 262, 105–112. [Google Scholar] [CrossRef] [PubMed]
- Blanch, M.; Dorca-Arévalo, J.; Not, A.; Cases, M.; Gómez de Aranda, I.; Martínez-Yélamos, A.; Martínez-Yélamos, S.; Solsona, C.; Blasi, J. The cytotoxicity of epsilon toxin from clostridium perfringens on lymphocytes is mediated by mal protein expression. Mol. Cell. Biol. 2018, 38, e00086-18. [Google Scholar] [CrossRef] [PubMed]
- Cases, M.; Llobet, A.; Terni, B.; Gómez de Aranda, I.; Blanch, M.; Doohan, B.; Revill, A.; Brown, A.M.; Blasi, J.; Solsona, C. Acute effect of pore-forming clostridium perfringens ε-toxin on compound action potentials of optic nerve of mouse. eNeuro 2017, 4. [Google Scholar] [CrossRef] [PubMed]
- Uzal, F.A.; Navarro, M.A.; Li, J.; Freedman, J.C.; Shrestha, A.; McClane, B.A. Comparative pathogenesis of enteric clostridial infections in humans and animals. Anaerobe 2018, 53, 11–20. [Google Scholar] [CrossRef] [PubMed]
- Linden, J.R.; Ma, Y.; Zhao, B.; Harris, J.M.; Rumah, K.R.; Schaeren-Wiemers, N.; Vartanian, T. Clostridium perfringens epsilon toxin causes selective death of mature oligodendrocytes and central nervous system demyelination. mBio 2015, 6, e02513–e02514. [Google Scholar] [CrossRef] [PubMed]
- Wagley, S.; Bokori-Brown, M.; Morcrette, H.; Malaspina, A.; D’Arcy, C.; Gnanapavan, S.; Lewis, N.; Popoff, M.R.; Raciborska, D.; Nicholas, R.; et al. Evidence of clostridium perfringens epsilon toxin associated with multiple sclerosis. Mult. Scler. J. 2018, 1352458518767327. [Google Scholar] [CrossRef] [PubMed]
- Yin, J.-X.; Tang, Z.; Gan, Y.; Li, L.; Shi, F.; Coons, S.; Shi, J. Pertussis toxin modulates microglia and t cell profile to protect experimental autoimmune encephalomyelitis. Neuropharmacology 2014, 81, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Weber, M.S.; Benkhoucha, M.; Lehmann-Horn, K.; Hertzenberg, D.; Sellner, J.; Santiago-Raber, M.-L.; Chofflon, M.; Hemmer, B.; Zamvil, S.S.; Lalive, P.H. Repetitive pertussis toxin promotes development of regulatory t cells and prevents central nervous system autoimmune disease. PLoS ONE 2011, 5, e16009. [Google Scholar] [CrossRef] [PubMed]
- Steelman, A.J. Infection as an environmental trigger of multiple sclerosis disease exacerbation. Front. Immunol. 2015, 6, 520. [Google Scholar] [CrossRef] [PubMed]
- Dressler, D.; Bhidayasiri, R.; Bohlega, S.; Chahidi, A.; Chung, T.M.; Ebke, M.; Jacinto, L.J.; Kaji, R.; Koçer, S.; Kanovsky, P.; et al. Botulinum toxin therapy for treatment of spasticity in multiple sclerosis: Review and recommendations of the iab-interdisciplinary working group for movement disorders task force. J. Neurol. 2017, 264, 112–120. [Google Scholar] [CrossRef] [PubMed]
- Cameron, M.H.; Bethoux, F.; Davis, N.; Frederick, M. Botulinum toxin for symptomatic therapy in multiple sclerosis. Curr. Neurol. Neurosci. Rep. 2014, 14, 463. [Google Scholar] [CrossRef] [PubMed]
- Latino, P.; Castelli, L.; Prosperini, L.; Marchetti, M.R.; Pozzilli, C.; Giovannelli, M. Determinants of botulinum toxin discontinuation in multiple sclerosis: A retrospective study. Neurol. Sci. 2017, 38, 1841–1848. [Google Scholar] [CrossRef] [PubMed]
- Purzycki, C.B.; Shain, D.H. Fungal toxins and multiple sclerosis: A compelling connection. Brain Res. Bull. 2010, 82, 4–6. [Google Scholar] [CrossRef] [PubMed]
- Speth, C.; Rambach, G.; Lass-Flörl, C.; Würzner, R.; Gasque, P.; Mohsenipour, I.; Dierich, M.P. Culture supernatants of patient-derived aspergillus isolates have toxic and lytic activity towards neurons and glial cells. FEMS Immunol. Med. Microbiol. 2006, 29, 303–313. [Google Scholar] [CrossRef] [PubMed]
- Razafimanjato, H.; Garmy, N.; Guo, X.-J.; Varini, K.; Di Scala, C.; Di Pasquale, E.; Taïeb, N.; Maresca, M. The food-associated fungal neurotoxin ochratoxin a inhibits the absorption of glutamate by astrocytes through a decrease in cell surface expression of the excitatory amino-acid transporters glast and glt-1. Neurotoxicology 2010, 31, 475–484. [Google Scholar] [CrossRef] [PubMed]
- Benito-León, J.; Pisa, D.; Alonso, R.; Calleja, P.; Díaz-Sánchez, M.; Carrasco, L. Association between multiple sclerosis and candida species: Evidence from a case-control study. Eur. J. Clin. Microbiol. Infect. Dis. 2010, 29, 1139–1145. [Google Scholar] [CrossRef] [PubMed]
- Pisa, D.; Alonso, R.; Carrasco, L. Fungal infection in a patient with multiple sclerosis. Eur. J. Clin. Microbiol. Infect. Dis. 2011, 30, 1173–1180. [Google Scholar] [CrossRef] [PubMed]
- Pisa, D.; Alonso, R.; Jiménez-Jiménez, F.J.; Carrasco, L. Fungal infection in cerebrospinal fluid from some patients with multiple sclerosis. Eur. J. Clin. Microbiol. Infect. Dis. 2013, 32, 795–801. [Google Scholar] [CrossRef] [PubMed]
- Fraga-Silva, T.F.C.; Mimura, L.A.N.; Marchetti, C.M.; Chiuso-Minicucci, F.; França, T.G.D.; Zorzella-Pezavento, S.F.G.; Venturini, J.; Arruda, M.S.P.; et al. Experimental autoimmune encephalomyelitis development is aggravated by candida albicans infection. J. Immunol. Res. 2015, 2015, 11. [Google Scholar] [CrossRef] [PubMed]
- Sotgiu, S.; Musumeci, S.; Marconi, S.; Gini, B.; Bonetti, B. Different content of chitin-like polysaccharides in multiple sclerosis and alzheimer’s disease brains. J. Neuroimmunol. 2008, 197, 70–73. [Google Scholar] [CrossRef] [PubMed]
- Barone, R.; Sotgiu, S.; Musumeci, S. Plasma chitotriosidase in health and pathology. Clin. Lab. 2007, 53, 321–333. [Google Scholar] [PubMed]
- Comabella, M.; Domínguez, C.; Rio, J.; Martín-Gallán, P.; Vilches, A.; Vilarrasa, N.; Espejo, C.; Montalban, X. Plasma chitotriosidase activity in multiple sclerosis. Clin. Immunol. 2009, 131, 216–222. [Google Scholar] [CrossRef] [PubMed]
- Correale, J.; Fiol, M. Chitinase effects on immune cell response in neuromyelitis optica and multiple sclerosis. Mult. Scler. J. 2010, 17, 521–531. [Google Scholar] [CrossRef] [PubMed]
- Lindstedt, M. Multiple sclerosis—Is research on the wrong track? Med. Hypotheses 1991, 34, 69–72. [Google Scholar] [CrossRef]
- Ramos, M.; Pisa, D.; Molina, S.; Rábano, A.; Juarranz, A.; Carrasco, L. Fungal infection in patients with multiple sclerosis. Open Mycol. J. 2008, 2, 22–28. [Google Scholar] [CrossRef]
- Napier, M.D.; Poole, C.; Satten, G.A.; Ashley-Koch, A.; Marrie, R.A.; Williamson, D.M. Heavy metals, organic solvents and multiple sclerosis: An exploratory look at gene-environment interactions. Arch. Environ. Occup. Health 2016, 71, 26–34. [Google Scholar] [CrossRef] [PubMed]
- Compston, A.; Lassmann, H.; McDonald, I.; Miller, D.; Noseworthy, J.; Smith, K.; Wekerle, H.; Confavreux, C. The story of multiple sclerosis. In Mcalpine’s Multiple Sclerosis, 4th ed.; Churchill Livingstone: London, UK, 2005. [Google Scholar]
- Parron, T.; Requena, M.; Hernandez, A.F.; Alarcon, R. Association between environmental exposure to pesticides and neurodegenerative diseases. Toxicol. Appl. Pharmacol. 2011, 256, 379–385. [Google Scholar] [CrossRef] [PubMed]
- Savage, E.P.; Keefe, T.J.; Mounce, L.M.; Heaton, R.K.; Lewis, J.A.; Burcar, P.J. Chronic neurological sequelae of acute organophosphate pesticide poisoning. Arch. Environ. Health 1988, 43, 38–45. [Google Scholar] [CrossRef] [PubMed]
- Bove, J.; Prou, D.; Perier, C.; Przedborski, S. Toxin-induced models of parkinson’s disease. NeuroRx 2005, 2, 484–494. [Google Scholar] [CrossRef] [PubMed]
- Dubey, D.; Sharma, V.; Stuve, O. Multiple mothballs or multiple sclerosis: A diagnostic dilemma (p5.192). Neurology 2014, 82, 192. [Google Scholar]
- Oddone, E.; Crosignani, P.; Scaburri, A.; Bai, E.; Modonesi, C.; Imbriani, M.; Bergamaschi, R. Occupation and multiple sclerosis: An italian case-control study. Occup. Environ. Med. 2013, 70, A91. [Google Scholar] [CrossRef]
- Magyari, M.; Koch-Henriksen, N.; Pfleger, C.C.; Sorensen, P.S. Physical and social environment and the risk of multiple sclerosis. Mult. Scler. Relat. Disord. 2014, 3, 600–606. [Google Scholar] [CrossRef] [PubMed]
- Graves, J.; Chitnis, T.; Weinstock-Guttman, B.; Rubin, J.; Zelikovitch, A.; Nourbakhsh, B.; Simmons, T.; Casper, C.; Waubant, E. Maternal illness in pregnancy and perinatal exposure to pesticides are associated with risk for pediatric onset ms (s29.005). Neurology 2016, 86, S29.005. [Google Scholar]
- Landtblom, A.-M.; Flodin, U.; Söderfeldt, B.; Wolfson, C.; Axelson, O. Organic solvents and multiple sclerosis: A synthesis of the current evidence. Epidemiology 1996, 7, 429–433. [Google Scholar] [CrossRef] [PubMed]
- Iranmanesh, F.; Ebrahimi, H.a.; Iranmanesh, M.; Sedighi, B.; Gadari, F. Multiple sclerosis and mines: An epidemiologic study from kerman province, Iran. Int. Clin. Neurosci. J. 2015, 2, 133–136. [Google Scholar]
- Razavi, Z.; Jokar, M.; Allafchian, A.; Hossinpour, Z.; Berenjani, L.; Shayegan Nejad, V. The relationship between blood lead levels and clinical features among multiple sclerosis patients in Isfahan, Iran. Iran. J. Health, Saf. Environ. 2016, 3, 412–420. [Google Scholar]
- Monti, M.C.; Guido, D.; Montomoli, C.; Sardu, C.; Sanna, A.; Pretti, S.; Lorefice, L.; Marrosu, M.G.; Valera, P.; Cocco, E. Is geo-environmental exposure a risk factor for multiple sclerosis? A population-based cross-sectional study in south-western sardinia. PLoS ONE 2016, 11, e0163313. [Google Scholar] [CrossRef] [PubMed]
- Giacoppo, S.; Galuppo, M.; Calabro, R.S.; D’Aleo, G.; Marra, A.; Sessa, E.; Bua, D.G.; Potorti, A.G.; Dugo, G.; Bramanti, P.; et al. Heavy metals and neurodegenerative diseases: An observational study. Biol. Trace Elem. Res. 2014, 161, 151–160. [Google Scholar] [CrossRef] [PubMed]
- Fulgenzi, A.; Zanella, S.G.; Mariani, M.M.; Vietti, D.; Ferrero, M.E. A case of multiple sclerosis improvement following removal of heavy metal intoxication: Lessons learnt from matteo’s case. Biometals 2012, 25, 569–576. [Google Scholar] [CrossRef] [PubMed]
- Crowe, W.; Allsopp, P.J.; Watson, G.E.; Magee, P.J.; Strain, J.J.; Armstrong, D.J.; Ball, E.; McSorley, E.M. Mercury as an environmental stimulus in the development of autoimmunity—A systematic review. Autoimmun. Rev. 2017, 16, 72–80. [Google Scholar] [CrossRef] [PubMed]
- Pamphlett, R.; Kum Jew, S. Inorganic mercury in human astrocytes, oligodendrocytes, corticomotoneurons and the locus ceruleus: Implications for multiple sclerosis, neurodegenerative disorders and gliomas. Biometals 2018, 31, 807–819. [Google Scholar] [CrossRef] [PubMed]
- Pourahmad, J.; Kahrizi, F.; Naderi, N.; Salimi, A.; Noorbakhsh, F.; Faizi, M.; Naserzadeh, P. Repeated administration of mercury accelerates progression of multiple sclerosis through mitochondrial dysfunction. Iran. J. Pharm. Res. 2016, 15, 834–841. [Google Scholar]
- Guzzi, G.; Costa, A.; Pigatto, P. Serum nse and multiple sclerosis. J. Neurol. Sci. 2015, 358, 463. [Google Scholar] [CrossRef] [PubMed]
- Soni, R.; Bhatnagar, A.; Vivek, R.; Chaturvedi, T.; Singh, A. A systematic review on mercury toxicity from dental amalgam fillings and its management strategies. J. Sci. Res. 2012, 56, 81–92. [Google Scholar]
- Bjørklund, G.; Hilt, B.; Dadar, M.; Lindh, U.; Aaseth, J. Neurotoxic effects of mercury exposure in dental personnel. Basic Clin. Pharmacol. Toxicol. 2018. [Google Scholar] [CrossRef] [PubMed]
- Cariccio, V.L.; Samà, A.; Bramanti, P.; Mazzon, E. Mercury involvement in neuronal damage and in neurodegenerative diseases. Biol. Trace Elem. Res. 2019, 187, 341–356. [Google Scholar] [CrossRef] [PubMed]
- Tsai, C.-P.; Lee, C.T.-C. Multiple sclerosis incidence associated with the soil lead and arsenic concentrations in taiwan. PLoS ONE 2013, 8, e65911. [Google Scholar] [CrossRef] [PubMed]
- Dehghanifiroozabadi, M.; Noferesti, P.; Amirabadizadeh, A.; Nakhaee, S.; Aaseth, J.; Noorbakhsh, F.; Mehrpour, O. Blood lead levels and multiple sclerosis: A case-control study. Mult. Scler. Relat. Disord. 2019, 27, 151–155. [Google Scholar] [CrossRef] [PubMed]
- Alizadeh-Ghodsi, M.; Zavvari, A.; Ebrahimi-Kalan, A.; Shiri-Shahsavar, M.R.; Yousefi, B. The hypothetical roles of arsenic in multiple sclerosis by induction of inflammation and aggregation of tau protein: A commentary. Nutr. Neurosci. 2018, 21, 92–96. [Google Scholar] [CrossRef] [PubMed]
- Jaiser, S.R.; Winston, G.P. Copper deficiency myelopathy. J. Neurol. 2010, 257, 869–881. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Palm, R.; Hallmans, G. Zinc and copper in multiple sclerosis. J. Neurol. Neurosurg. Psychiatry 1982, 45, 691–698. [Google Scholar] [CrossRef] [PubMed]
- Bredholt, M.; Frederiksen, J.L. Zinc in multiple sclerosis: A systematic review and meta-analysis. ASN Neuro 2016, 8. [Google Scholar] [CrossRef] [PubMed]
- Zahid Rasul, N.; Naqab, K.; Samiullah, K.; Mehboob, A.; Mohammad Amjad, K. Potential application of venom proteins in designing of medicines for treating human neurodegenerative disorders. Protein Pept. Lett. 2018, 25, 633–642. [Google Scholar]
- Iwai, S.; Okazaki, M.; Kiuchi, Y.; Oguchi, K. Changes in mrna levels of fibrinogen subunit polypeptides in rats defibrinogenated with batroxobin. Thromb. Res. 1999, 96, 421–426. [Google Scholar] [CrossRef]
- Hinman, C.L.; Stevens-Truss, R.; Schwarz, C.; Hudson, R.A. Sequence determinants of modified cobra venom neurotoxin which induce immune resistance to experimental allergic encephalomyelitis: Molecular mechans for immunologic action. Immunopharmacol. Immunotoxicol. 1999, 21, 483–506. [Google Scholar] [CrossRef] [PubMed]
- Komegae, E.N.; Souza, T.A.M.; Grund, L.Z.; Lima, C.; Lopes-Ferreira, M. Multiple functional therapeutic effects of tnp: A small stable synthetic peptide derived from fish venom in a mouse model of multiple sclerosis. PLoS ONE 2017, 12, e0171796. [Google Scholar] [CrossRef] [PubMed]
- Ontaneda, D.; Hyland, M.; Cohen, J.A. Multiple sclerosis: New insights in pathogenesis and novel therapeutics. Annu. Rev. Med. 2012, 63, 389–404. [Google Scholar] [CrossRef] [PubMed]
- Kamm, C.P.; Uitdehaag, B.M.; Polman, C.H. Multiple sclerosis: Current knowledge and future outlook. Eur. Neurol. 2014, 72, 132–141. [Google Scholar] [CrossRef] [PubMed]
- Hernan, M.A.; Alonso, A.; Hernandez-Diaz, S. Tetanus vaccination and risk of multiple sclerosis: A systematic review. Neurology 2006, 67, 212–215. [Google Scholar] [CrossRef] [PubMed]
- Alonso, A.; Jick, S.S.; Jick, H.; Hernan, M.A. Antibiotic use and risk of multiple sclerosis. Am. J. Epidemiol. 2006, 163, 997–1002. [Google Scholar] [CrossRef] [PubMed]
- Alonso, A.; Jick, S.S.; Hernan, M.A. Allergy, histamine 1 receptor blockers, and the risk of multiple sclerosis. Neurology 2006, 66, 572–575. [Google Scholar] [CrossRef] [PubMed]
- Ifergan, I.; Wosik, K.; Cayrol, R.; Kébir, H.; Auger, C.; Bernard, M.; Bouthillier, A.; Moumdjian, R.; Duquette, P.; Prat, A. Statins reduce human blood-brain barrier permeability and restrict leukocyte migration: Relevance to multiple sclerosis. Ann. Neurol. 2006, 60, 45–55. [Google Scholar] [CrossRef] [PubMed]
- Mabunga, D.F.N.; Gonzales, E.L.T.; Kim, J.-w.; Kim, K.C.; Shin, C.Y. Exploring the validity of valproic acid animal model of autism. Exp. Neurobiol. 2015, 24, 285–300. [Google Scholar] [CrossRef] [PubMed]
- Pazhoohan, S.; Satarian, L.; Asghari, A.A.; Salimi, M.; Kiani, S.; Mani, A.R.; Javan, M. Valproic acid attenuates disease symptoms and increases endogenous myelin repair by recruiting neural stem cells and oligodendrocyte progenitors in experimental autoimmune encephalomyelitis. Neurodegener. Dis. 2014, 13, 45–52. [Google Scholar] [CrossRef] [PubMed]
- Lv, J.; Du, C.; Wei, W.; Wu, Z.; Zhao, G.; Li, Z.; Xie, X. The antiepileptic drug valproic acid restores t cell homeostasis and ameliorates pathogenesis of experimental autoimmune encephalomyelitis. J. Biol. Chem. 2012, 287, 28656–28665. [Google Scholar] [CrossRef] [PubMed]
- Long, J.; Chang, L.; Shen, Y.; Gao, W.H.; Wu, Y.N.; Dou, H.B.; Huang, M.M.; Wang, Y.; Fang, W.Y.; Shan, J.H.; et al. Valproic acid ameliorates graft-versus-host disease by downregulating th1 and th17 cells. J. Immunol. 2015, 195, 1849–1857. [Google Scholar] [CrossRef] [PubMed]
- Askari, V.R.; Fereydouni, N.; Baradaran Rahimi, V.; Askari, N.; Sahebkar, A.H.; Rahmanian-Devin, P.; Samzadeh-Kermani, A. B-amyrin, the cannabinoid receptors agonist, abrogates mice brain microglial cells inflammation induced by lipopolysaccharide/interferon-γ and regulates mφ1/mφ2 balances. Biomed. Pharmacother. 2018, 101, 438–446. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, M.; Moran, S.; Wen, J.; Affram, K.; Chen, T.; Symes, A.J.; Zhang, Y. Wwl70 attenuates pge(2) production derived from 2-arachidonoylglycerol in microglia by abhd6-independent mechanism. J. Neuroinflamm. 2017, 14, 7. [Google Scholar] [CrossRef] [PubMed]
- Guo, C.; Fulp, J.W.; Jiang, Y.; Li, X.; Chojnacki, J.E.; Wu, J.; Wang, X.-Y.; Zhang, S. Development and characterization of a hydroxyl-sulfonamide analogue, 5-chloro-n-[2-(4-hydroxysulfamoyl-phenyl)-ethyl]-2-methoxy-benzamide, as a novel nlrp3 inflammasome inhibitor for potential treatment of multiple sclerosis. ACS Chem. Neurosci. 2017, 8, 2194–2201. [Google Scholar] [CrossRef] [PubMed]
- Leonoudakis, D.; Rane, A.; Angeli, S.; Lithgow, G.J.; Andersen, J.K.; Chinta, S.J. Anti-inflammatory and neuroprotective role of natural product securinine in activated glial cells: Implications for parkinson’s disease. Med. Inflamm. 2017, 2017, 8302636. [Google Scholar] [CrossRef] [PubMed]
- Kappos, L.; Gold, R.; Miller, D.H.; MacManus, D.G.; Havrdova, E.; Limmroth, V.; Polman, C.H.; Schmierer, K.; Yousry, T.A.; Yang, M.; et al. Efficacy and safety of oral fumarate in patients with relapsing-remitting multiple sclerosis: A multicentre, randomised, double-blind, placebo-controlled phase iib study. Lancet 2008, 372, 1463–1472. [Google Scholar] [CrossRef]
- Stangel, M.; Linker, R.A. Dimethyl fumarate (bg-12) for the treatment of multiple sclerosis. Expert Rev. Clin. Pharmacol. 2013, 6, 355–362. [Google Scholar] [CrossRef] [PubMed]
- Linker, R.A.; Gold, R. Dimethyl fumarate for treatment of multiple sclerosis: Mechanism of action, effectiveness, and side effects. Curr. Neurol. Neurosci. Rep. 2013, 13, 394. [Google Scholar] [CrossRef] [PubMed]
- Bomprezzi, R. Dimethyl fumarate in the treatment of relapsing–remitting multiple sclerosis: An overview. Ther. Adv. Neurol. Disord. 2015, 8, 20–30. [Google Scholar] [CrossRef] [PubMed]
- Gopal, S.; Mikulskis, A.; Gold, R.; Fox, R.J.; Dawson, K.T.; Amaravadi, L. Evidence of activation of the nrf2 pathway in multiple sclerosis patients treated with delayed-release dimethyl fumarate in the phase 3 define and confirm studies. Mult. Scler. 2017, 23, 1875–1883. [Google Scholar] [CrossRef] [PubMed]
- Mills, E.A.; Ogrodnik, M.A.; Plave, A.; Mao-Draayer, Y. Emerging understanding of the mechanism of action for dimethyl fumarate in the treatment of multiple sclerosis. Front. Neurol. 2018, 9, 5. [Google Scholar] [CrossRef] [PubMed]
- Vego, H.; Sand, K.L.; Høglund, R.A.; Fallang, L.-E.; Gundersen, G.; Holmøy, T.; Maghazachi, A.A. Monomethyl fumarate augments nk cell lysis of tumor cells through degranulation and the upregulation of nkp46 and cd107a. Cell Mol. Immunol. 2014, 13, 57. [Google Scholar] [CrossRef] [PubMed]
- Maghazachi, A.A.; Sand, K.L.; Al-Jaderi, Z. Glatiramer acetate, dimethyl fumarate, and monomethyl fumarate upregulate the expression of ccr10 on the surface of natural killer cells and enhance their chemotaxis and cytotoxicity. Front. Immunol. 2016, 7, 437. [Google Scholar] [CrossRef] [PubMed]
- Al-Jaderi, Z.; Maghazachi, A.A. Utilization of dimethyl fumarate and related molecules for treatment of multiple sclerosis, cancer, and other diseases. Front. Immunol. 2016, 7, 278. [Google Scholar] [CrossRef] [PubMed]
- Strader, C.R.; Pearce, C.J.; Oberlies, N.H. Fingolimod (fty720): A recently approved multiple sclerosis drug based on a fungal secondary metabolite. J. Nat. Prod. 2011, 74, 900–907. [Google Scholar] [CrossRef] [PubMed]
- Al-Jaderi, Z.; Maghazachi, A.A. Effects of vitamin d3, calcipotriol and fty720 on the expression of surface molecules and cytolytic activities of human natural killer cells and dendritic cells. Toxins (Basel) 2013, 5, 1932–1947. [Google Scholar] [CrossRef] [PubMed]


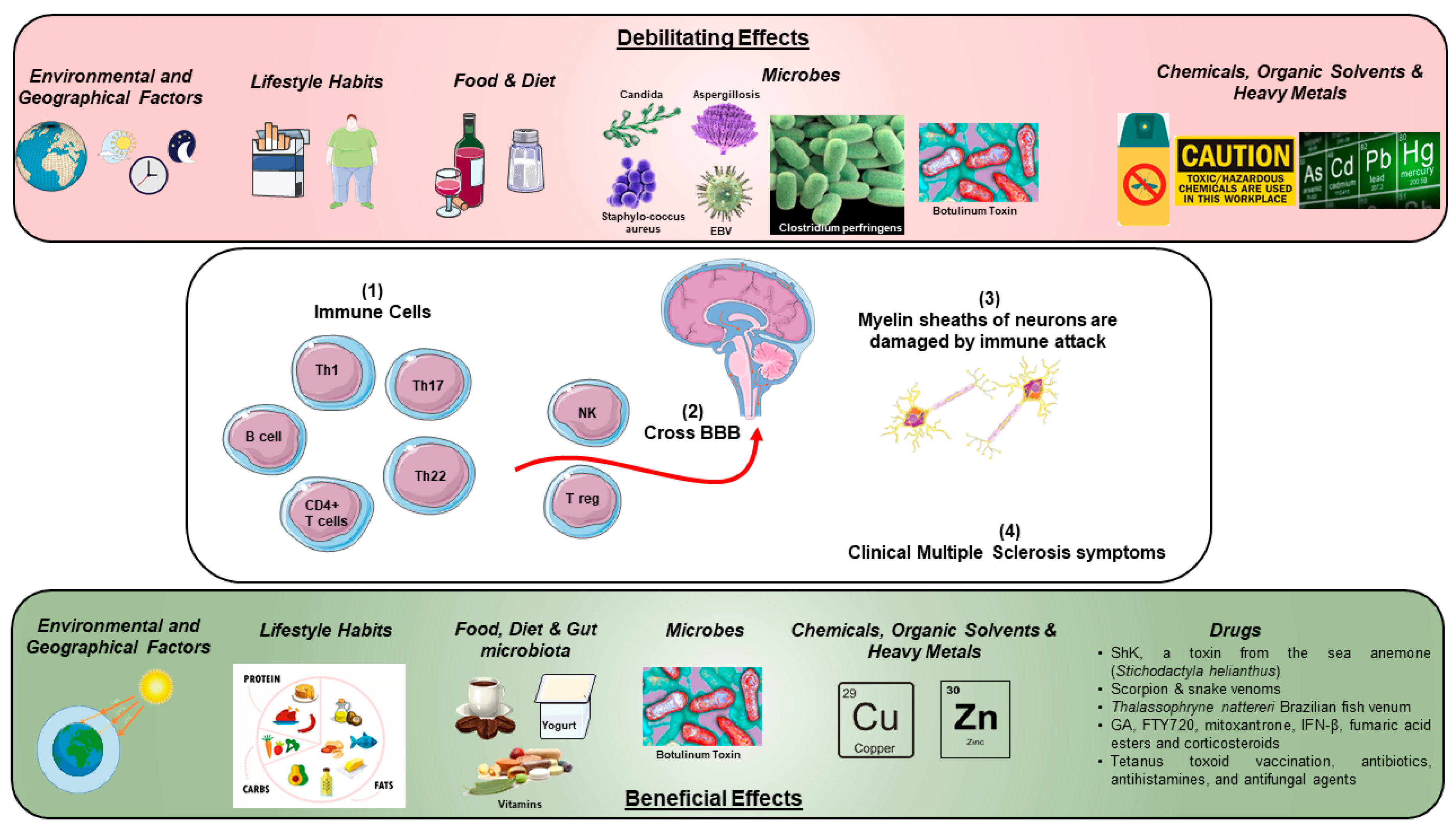{kind=link}
| Environmental and Microbial Toxins, Drugs, Organic Solvents and Heavy Metals | Beneficial Effect | Debilitating Effect |
|---|---|---|
| 1. Environmental and Geographical Factors | ||
| • Geographic latitude | Yes | |
| • Sun exposure as a vitamin D source | Yes | |
| • Circadian disruption and sleep restriction | Yes | |
| 2. Toxic Effects of Lifestyle Habits | ||
| • Smoking | Yes | |
| • Obesity and fatty acids intake | Yes | |
| • Ketogenic diet | Yes | |
| 3. Toxic Effects of Food, Diet and Gut Microbiota | ||
| • Coffee | Yes | |
| • Alcoholic beverages and fish | yes | |
| • High sodium intake | Yes | |
| • Vitamins | Yes | |
| • Probiotics | Yes | |
| 4. Toxic Effects of Microbes | ||
| • Epstein Barr virus (EBV) infection | Yes | |
| • Bacterial toxins include staphylococcal, nasopharyngeal normal flora | Yes | |
| • Clostridium perfringens epsilon toxin” ε-toxin,” | Yes | |
| • Pertussis toxin (PTX) and botulinum toxins | Yes | ? Yes |
| • Aspergillosis | Yes | |
| • Mycotoxin ochratoxin A | Yes | |
| • Candida species | Yes | |
| 5. Chemicals, Organic Solvents and Heavy Metals | ||
| • Pesticides and mothballs | Yes | |
| • Occupational chemical exposure | Yes | |
| • Heavy metals (mercury, lead, arsenic) | Yes | |
| • Copper | Yes | |
| • Zinc | Yes | Yes |
| 6. Drugs | ||
| • ShK, a toxin from the sea anemone (Stichodactyla helianthus) and scorpion venom | Yes | |
| • Snake venoms | Yes | |
| • Thalassophryne nattereri Brazilian fish venom | Yes | |
| • Glatiramer acetate, fingolimod (FTY720), mitoxantrone, IFN-β, fumaric acid esters and corticosteroids | Yes | |
| • Tetanus toxoid vaccination, antibiotics, antihistamines and antifungal agents | Yes |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hachim, M.Y.; Elemam, N.M.; Maghazachi, A.A. The Beneficial and Debilitating Effects of Environmental and Microbial Toxins, Drugs, Organic Solvents and Heavy Metals on the Onset and Progression of Multiple Sclerosis. Toxins 2019, 11, 147. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins11030147
Hachim MY, Elemam NM, Maghazachi AA. The Beneficial and Debilitating Effects of Environmental and Microbial Toxins, Drugs, Organic Solvents and Heavy Metals on the Onset and Progression of Multiple Sclerosis. Toxins. 2019; 11(3):147. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins11030147
Chicago/Turabian StyleHachim, Mahmood Y., Noha M. Elemam, and Azzam A. Maghazachi. 2019. "The Beneficial and Debilitating Effects of Environmental and Microbial Toxins, Drugs, Organic Solvents and Heavy Metals on the Onset and Progression of Multiple Sclerosis" Toxins 11, no. 3: 147. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins11030147