Regulation of Microcystin-LR-Induced DNA Damage by miR-451a in HL7702 Cells

 ,
,
Abstract
:1. Introduction
2. Results and Discussion
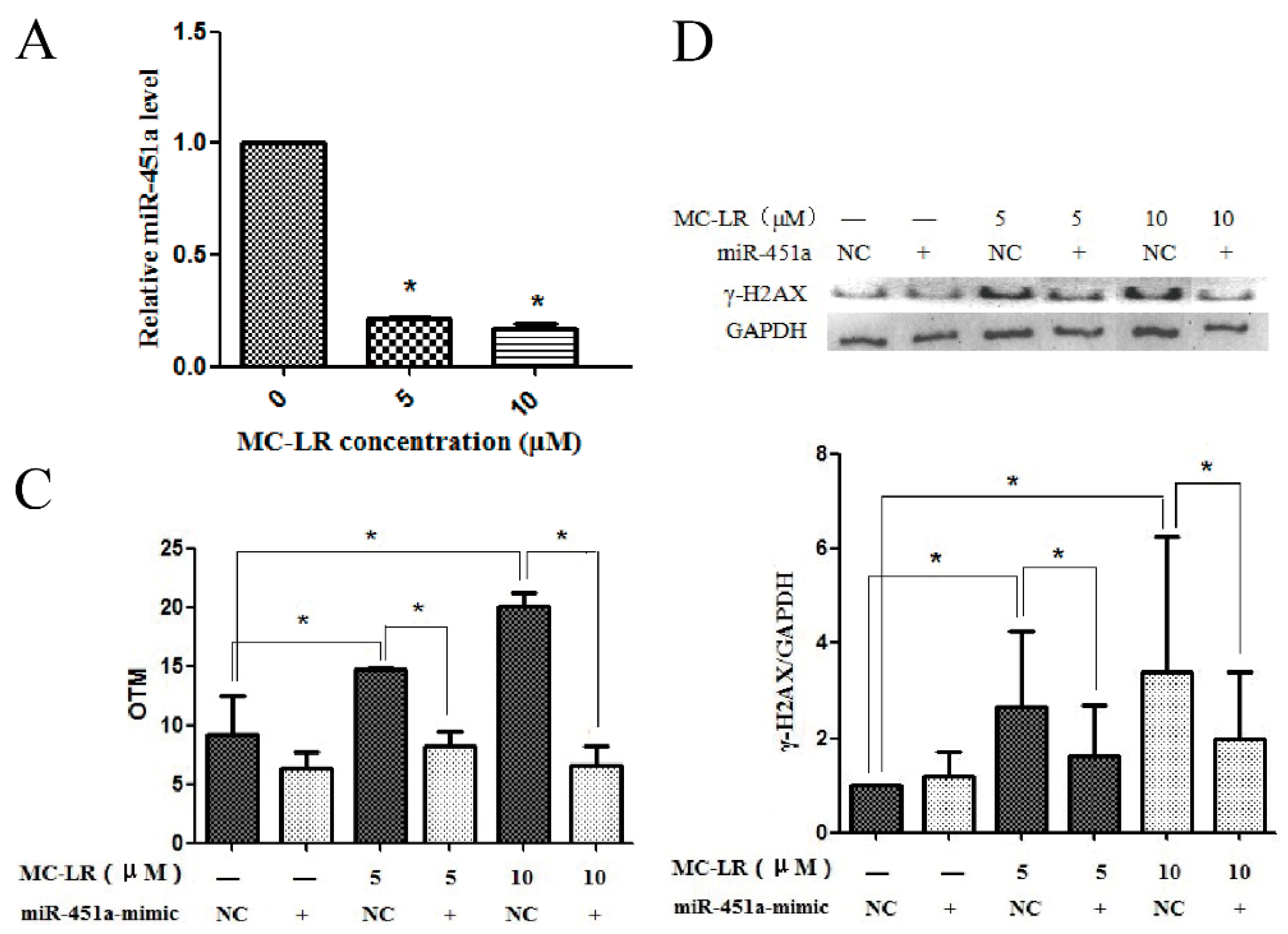
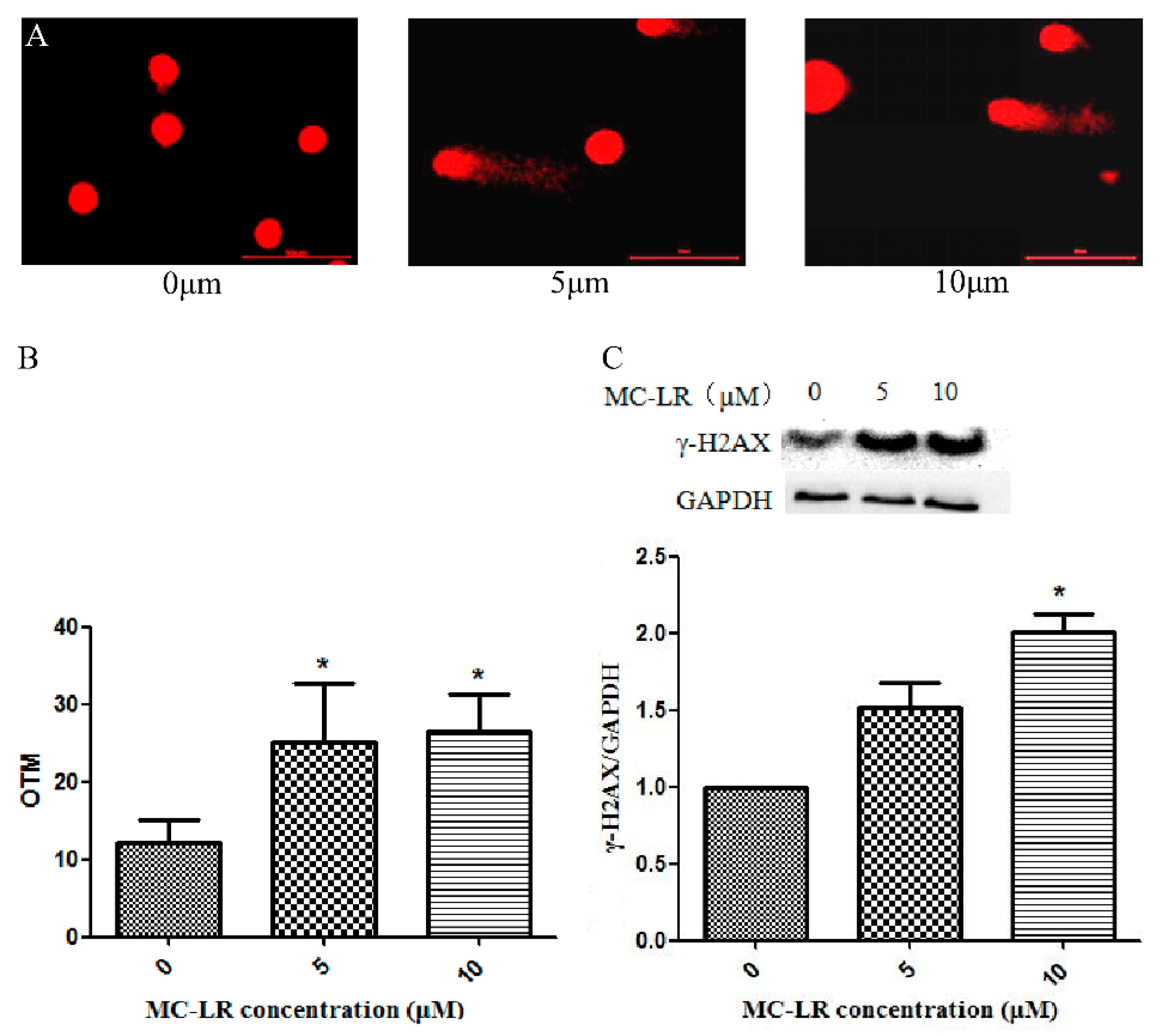
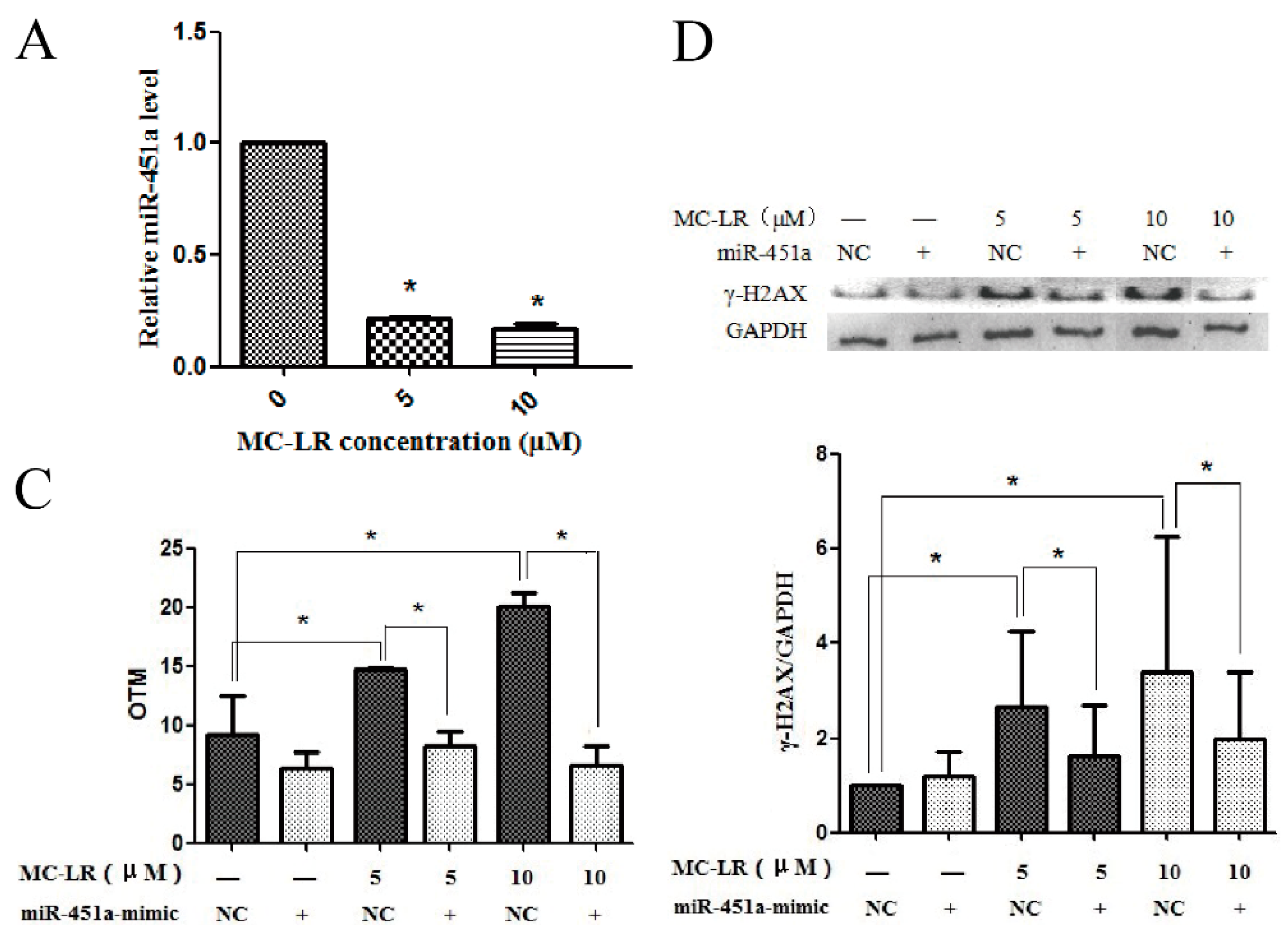
2.1. MC-LR Exerts DNA Damage on HL7702 Cells
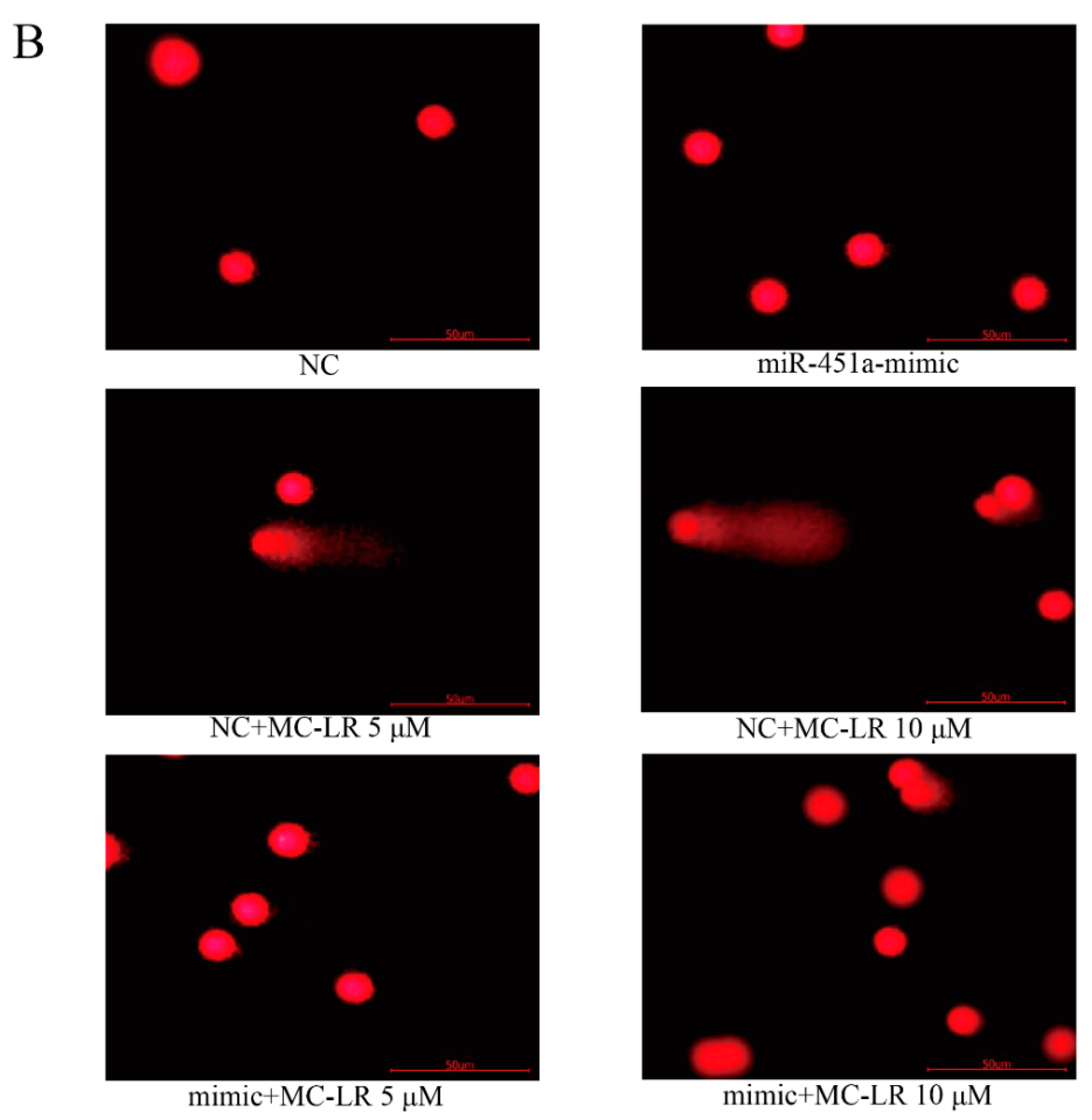
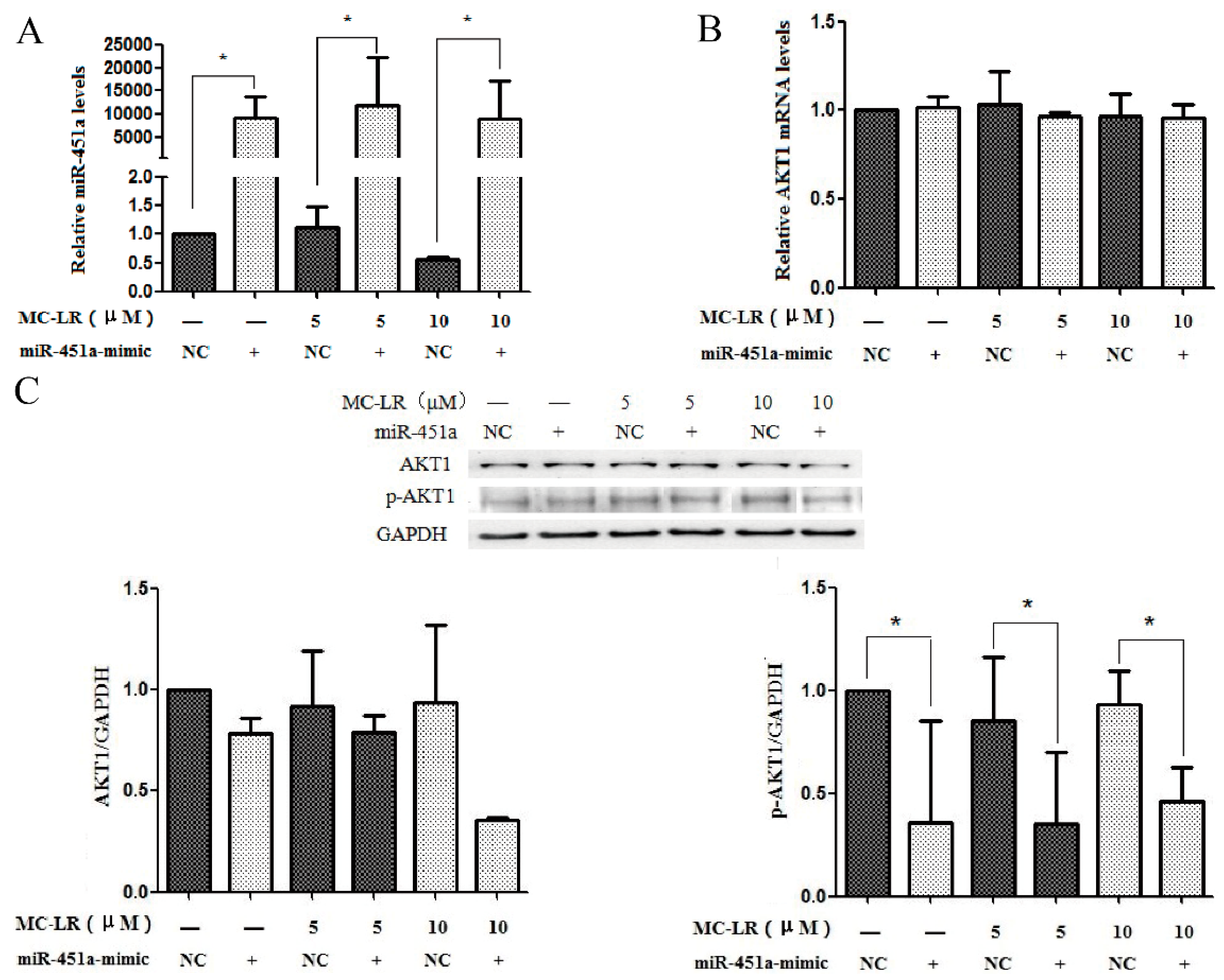
2.2. Over-Expression of miR-451a Protects HL7702 Cells from MC-LR-Induced DNA Damage
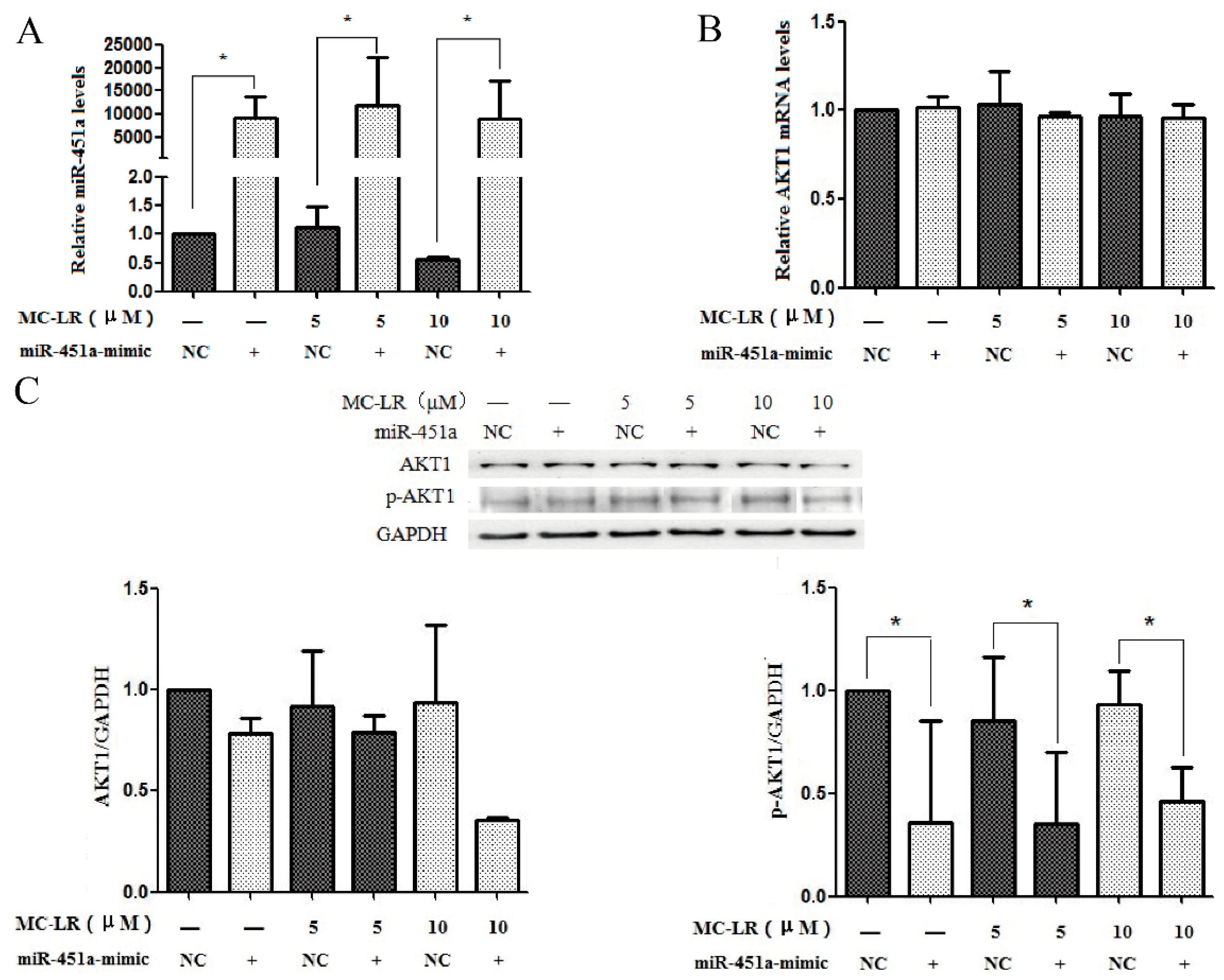
2.3. The Expression of AKT1 and p-AKT1 Is Regulated by miR-451a
3. Conclusions
4. Materials and Methods
4.1. Cell Culture and Treatment
4.2. The Comet Assay
4.3. Cell Transfection
4.4. Quantitative RT-PCR (qRT-PCR)
4.5. Western Blot
4.6. Statistical Analysis
Author Contributions
Funding
Conflicts of Interest
References
- Yang, F.; Guo, J.; Huang, F.; Massey, I.Y.; Huang, R.; Li, Y.; Wen, C.; Ding, P.; Zeng, W.; Liang, G. Removal of Microcystin-LR by a Novel Native Effective Bacterial Community Designated as YFMCD4 Isolated from Lake Taihu. Toxins 2018, 10, 363. [Google Scholar] [CrossRef] [PubMed]
- WHO. Cyanobacterial Toxins: Microcystin-lr. Guidelines for Drinking Water Quality; World Health Organization: Geneva, Switzerland, 1998. [Google Scholar]
- Massey, I.Y.; Yang, F.; Ding, Z.; Yang, S.; Guo, J.; Tezi, C.; Al-Osman, M.; Kamegni, R.B.; Zeng, W. Exposure routes and health effects of microcystins on animals and humans: A mini-review. Toxicon 2018, 151, 156–162. [Google Scholar] [CrossRef] [PubMed]
- Massey, I.Y.; Zhang, X.; Yang, F. Importance of bacterial biodegradation and detoxification processes of microcystins for environmental health. J. Toxicol. Environ. Health B 2018, 29, 1–13. [Google Scholar] [CrossRef] [PubMed]
- MacKintosh, C.; Beattie, K.A.; Klumpp, S.; Cohen, P.; Codd, G.A. Cyanobacterial microcystin-LR is a potent and specific inhibitor of protein phosphatases 1 and 2A from both mammals and higher plants. FEBS Lett. 1990, 264, 187–192. [Google Scholar] [CrossRef] [Green Version]
- Yang, F.; Wen, C.; Zheng, S.; Yang, S.; Chen, J.; Feng, X. Involvement of MAPK/ERK1/2 pathway in microcystin-induced microfilament reorganization in HL7702 hepatocytes. J. Toxicol. Environ. Health A 2018, 81, 1135–1141. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Zhang, S.; Liu, C.; Wu, J.; Wang, Y.; Yuan, L.; Du, X.; Wang, R.; Marwa, P.W.; Zhuang, D.; et al. Resveratrol Ameliorates Microcystin-LR-Induced Testis Germ Cell Apoptosis in Rats via SIRT1 Signaling Pathway Activation. Toxins 2018, 10, 235. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Liu, C.; Li, Y.; Imam, M.U.; Huang, H.; Liu, H.; Xin, Y.; Zhang, H. Novel Role of ER Stress and Autophagy in Microcystin-LR Induced Apoptosis in Chinese Hamster Ovary Cells. Front. Physiol. 2016, 7, 527. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Zhao, Q.; Zhou, W.; Xu, L.; Wang, Y. Effects of chronic exposure to microcystin-LR on hepatocyte mitochondrial DNA replication in mice. Environ. Sci. Technol. 2015, 49, 4665–4672. [Google Scholar] [CrossRef] [PubMed]
- Gaudin, J.; Huet, S.; Jarry, G.; Fessard, V. In vivo DNA damage induced by the cyanotoxin microcystin-LR: Comparison of intra-peritoneal and oral administrations by use of the comet assay. Mutat. Res. 2008, 652, 65–71. [Google Scholar] [CrossRef] [PubMed]
- Žegura, B.; Gajski, G.; Štraser, A.; Garaj-Vrhovac, V.; Filipič, M. Microcystin-LR induced DNA damage in human peripheral blood lymphocytes. Mutat. Res. Genet. Toxicol. Environ. Mutagen. 2011, 726, 116–122. [Google Scholar] [CrossRef] [PubMed]
- Ma, J.; Li, Y.; Yao, L.; Li, X. Analysis of MicroRNA Expression Profiling Involved in MC-LR-Induced Cytotoxicity by High-Throughput Sequencing. Toxins 2017, 9, 23. [Google Scholar] [CrossRef] [PubMed]
- Meng, X.; Zhang, L.; Chen, X.; Xiang, Z.; Li, D.; Han, X. miR-541 Contributes to Microcystin-LR-Induced Reproductive Toxicity through Regulating the Expression of p15 in Mice. Toxins 2016, 8, 260. [Google Scholar] [CrossRef] [PubMed]
- Bartel, D.P. MicroRNAs: Genomics, biogenesis, mechanism and function. Cell 2004, 116, 281–297. [Google Scholar] [CrossRef]
- Yang, S.; Chen, L.; Wen, C.; Zhang, X.; Feng, X.; Yang, F. MicroRNA expression profiling involved in MC-LR-induced hepatotoxicity using high-throughput sequencing analysis. J. Toxicol. Environ. Health A 2018, 81, 89–97. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Zeng, X.; Zhang, B.; Long, J.; Zhou, B.; Tan, G.; Zeng, W.; Chen, W.; Yang, J. MiR-451 inhibits cell proliferation in human hepatocellular carcinoma through direct suppression of IKK-β. Carcinogenesis 2013, 34, 2443–2451. [Google Scholar] [CrossRef] [PubMed]
- Minna, E.; Romeo, P.; Dugo, M.; Cecco, L.D.; Todoerti, K. MiR-451a is underexpressed and targets AKT/mTOR pathway in papillary thyroid carcinoma. Oncotarget 2016, 7, 12731–12747. [Google Scholar] [CrossRef] [PubMed]
- Hur, W.; Lee, J.H.; Kim, S.W.; Kim, J.H.; Bae, S.H.; Kim, M.; Hwang, D.; Kim, Y.S.; Park, T.; Um, S.J.; et al. Downregulation of microRNA-451 in non-alcoholic steatohepatitis inhibits fatty acid-induced proinflammatory cytokine production through the AMPK/AKT pathway. Int. J. Biochem. Cell Biol. 2015, 64, 265–276. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Szymonowicz, K.; Oeck, S.; Malewicz, N.; Jendrossek, V. New insights into protein kinase B/Akt signaling: Role of localized akt activation and Compartment-Specific target proteins for the cellular radiation response. Cancers 2018, 10, 78. [Google Scholar] [CrossRef] [PubMed]
- Xian, H.; Wang, F.; Teng, W.; Yang, D.; Zhang, M. Thyroid hormone induce a p53-dependent DNA damage through PI3K/Akt activation in sperm. Gene 2017, 615, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Habib, S.L.; Liang, S. Hyperactivation of Akt/mTOR and deficiency in tuberin increased the oxidative DNA damage in kidney cancer patients with diabetes. Oncotarget 2014, 5, 2542–2550. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, X.; He, X.; Yang, Z.; Jabbari, E. Mammalian PER2 regulates AKT activation and DNA damage response. Binchem. Cell Biol. 2012, 90, 675–682. [Google Scholar] [CrossRef] [PubMed]
- Xu, N.; Hegarat, N.; Black, E.J.; Scott, M.T.; Hochegger, H.; Gillespie, D.A. Akt/PKB suppresses DNA damage processing and checkpoint activation in late G2. J. Cell Biol. 2010, 190, 297–305. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, X.; Wu, X.; Shen, L.; Li, J.; Yu, R.; Liu, Y.; Qiu, G.; Zeng, W. Whole Genome Sequencing and Comparative Genomics Analyses of Pandoraea sp. XY-2, a New Species Capable of Biodegrade Tetracycline. Front. Microbiol. 2019, 10, 1–19. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| miRNA/mRNA | Forward Primer | Reverse Primer |
|---|---|---|
| miR-451a | 5′-CGGCGAAACCGTTACCATTAC-3′ | 5′-GTCGTATCCAGTGCAGGGTCCGAGGT-3′ |
| U6 | 5′-CTCGCTTCGGCAGCACATATACT-3′ | 5′-ACGCTTCACGAATTTGCGTGTC-3′ |
| AKT1 | 5′-CCATCACACCACCTGACCAA-3′ | 5′-TCCCTCCAAGCTATCGTCCA-3′ |
| GAPDH | 5′-TCGGAGTCAACGGATTTGGT-3′ | 5′-TGGAATTTGCCATGGGTGGA-3′ |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, L.; Yang, S.; Wen, C.; Zheng, S.; Yang, Y.; Feng, X.; Chen, J.; Luo, D.; Liu, R.; Yang, F. Regulation of Microcystin-LR-Induced DNA Damage by miR-451a in HL7702 Cells. Toxins 2019, 11, 164. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins11030164
Chen L, Yang S, Wen C, Zheng S, Yang Y, Feng X, Chen J, Luo D, Liu R, Yang F. Regulation of Microcystin-LR-Induced DNA Damage by miR-451a in HL7702 Cells. Toxins. 2019; 11(3):164. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins11030164
Chicago/Turabian StyleChen, Lv, Shu Yang, Cong Wen, Shuilin Zheng, Yue Yang, Xiangling Feng, Jihua Chen, Dan Luo, Ran Liu, and Fei Yang. 2019. "Regulation of Microcystin-LR-Induced DNA Damage by miR-451a in HL7702 Cells" Toxins 11, no. 3: 164. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins11030164