Pink Bollworm Resistance to Bt Toxin Cry1Ac Associated with an Insertion in Cadherin Exon 20


 , , , ,
, , , ,
Abstract
:1. Introduction
2. Results
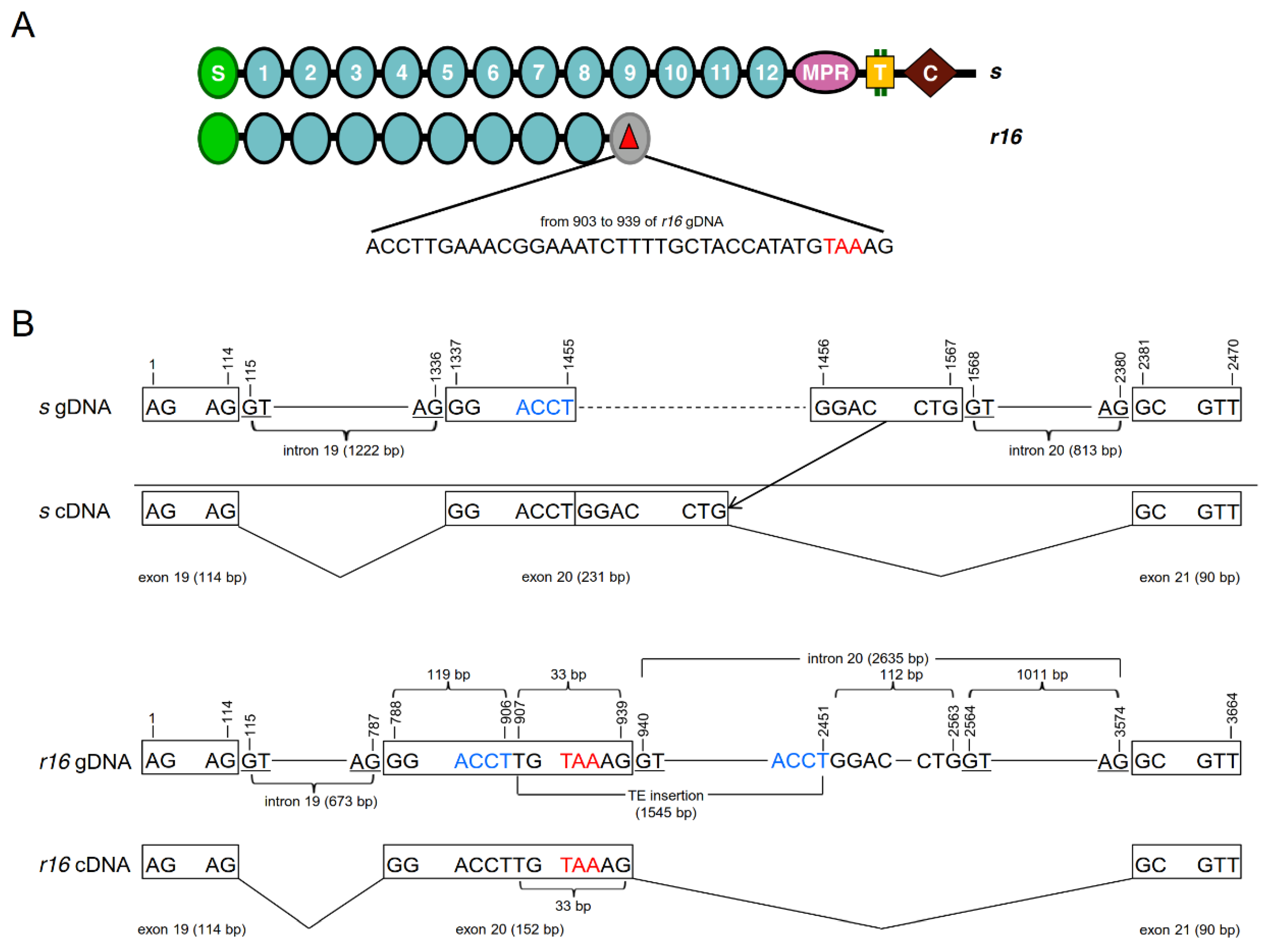
2.1. Isolation and Characterization of the Novel PgCad1 Allele r16
2.2. Inheritance of Cry1Ac Resistance and Cross-Resistance to Cry2Ab
2.3. Genetic Linkage between r16 Allele and Cry1Ac Resistance
2.4. Life History Traits of AQ65 on Bt and Non-Bt Cotton
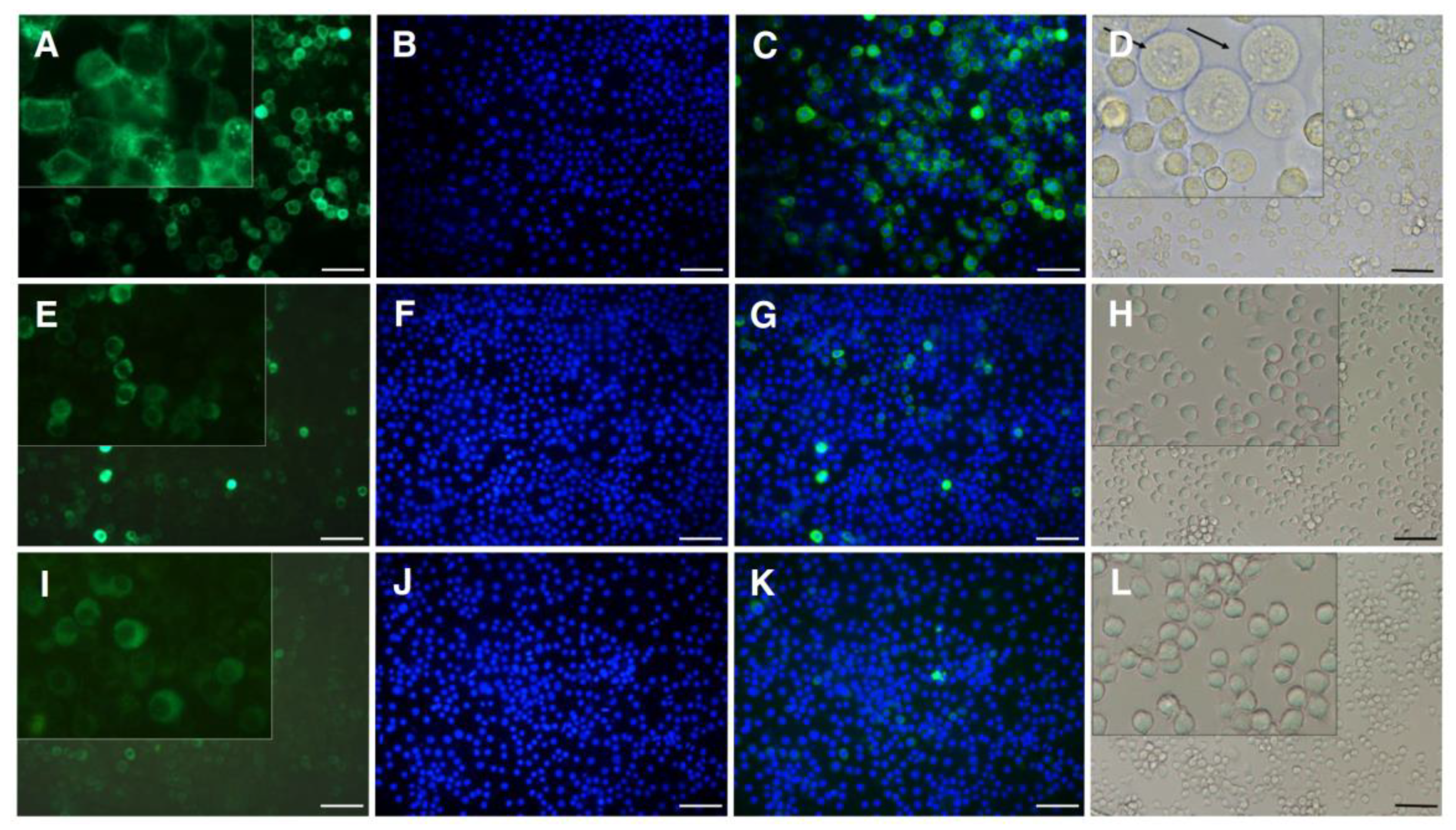
2.5. Transfected Cells: PgCad1 Localization and Susceptibility to Cry1Ac
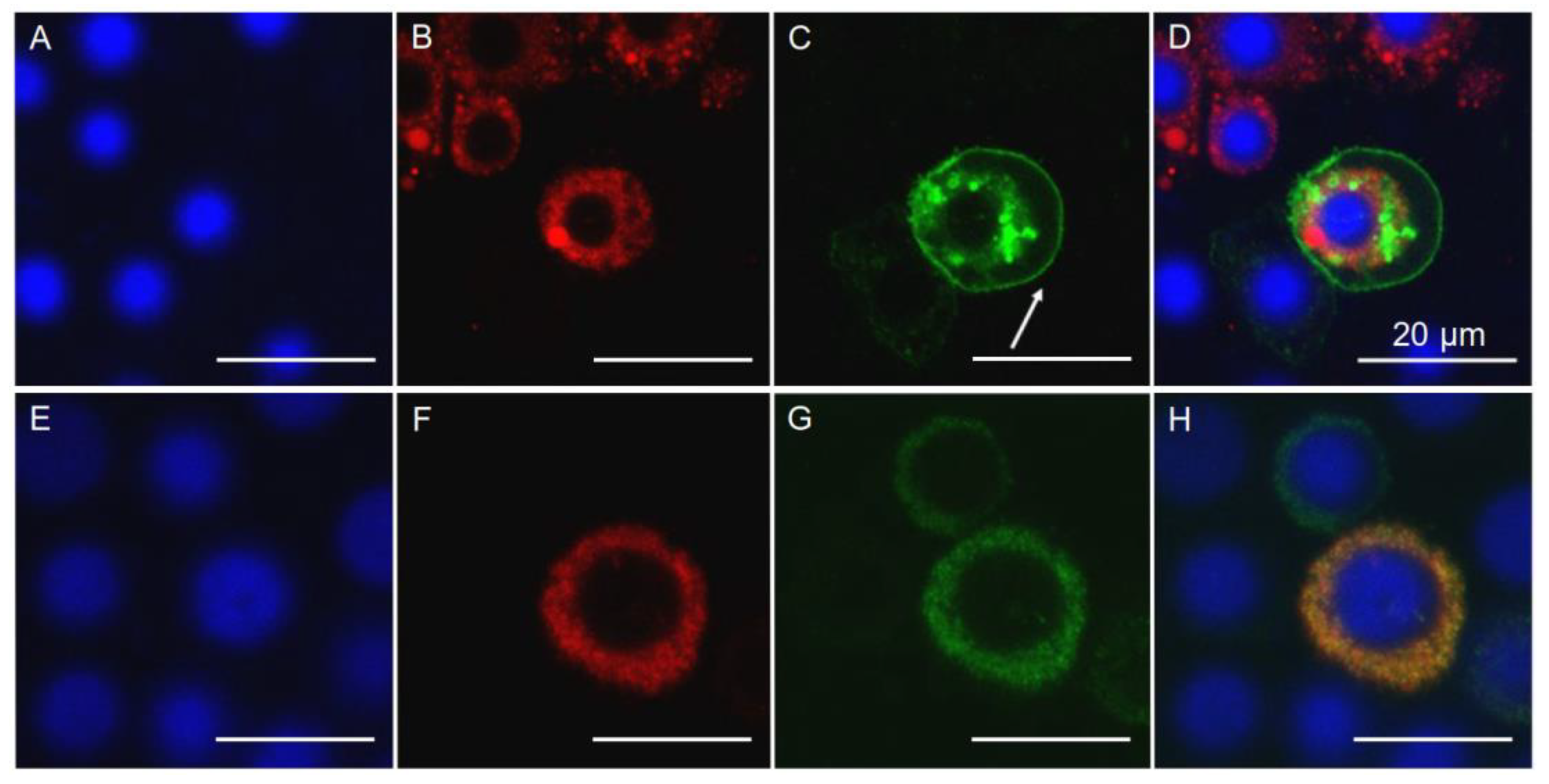
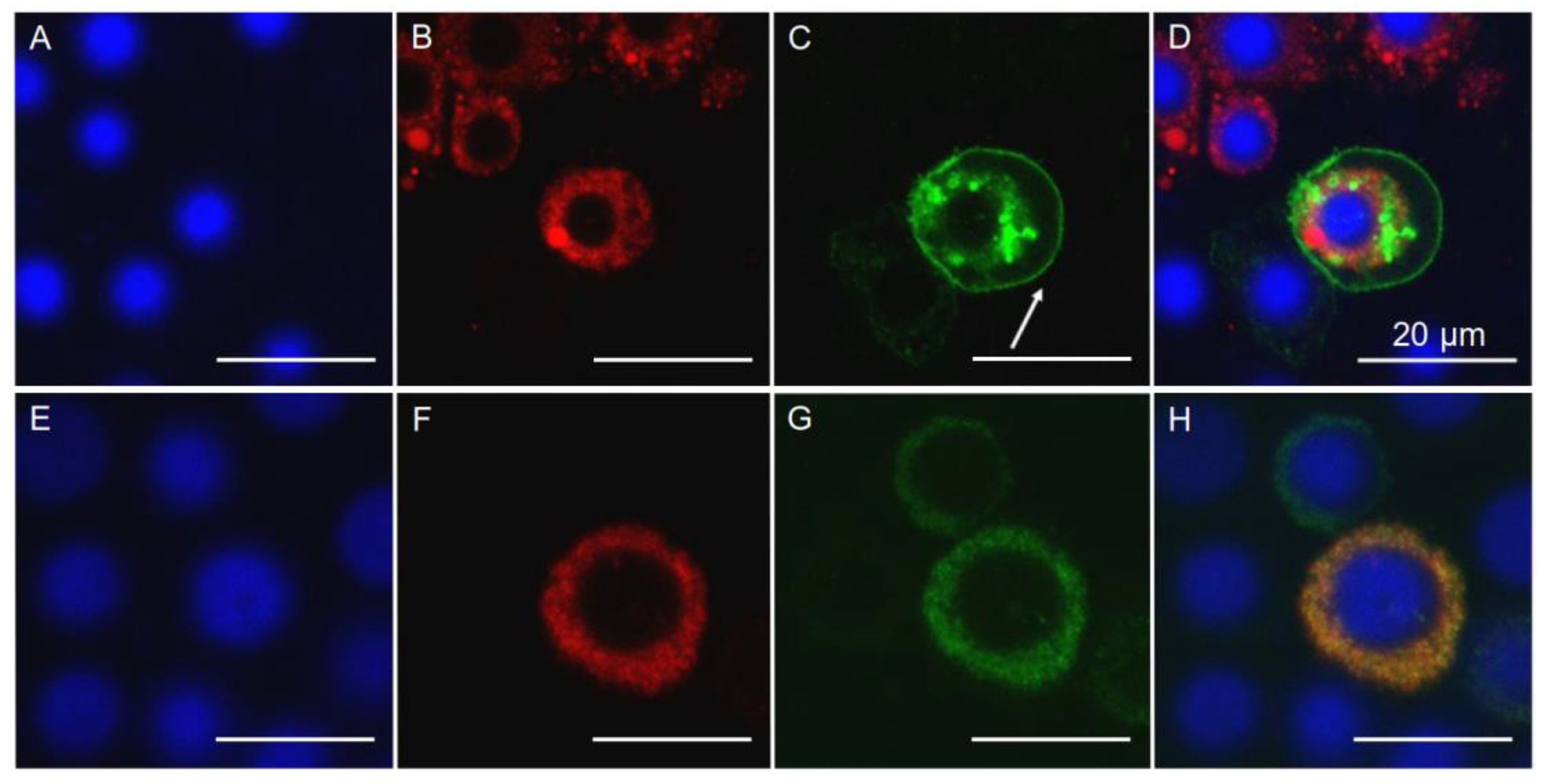
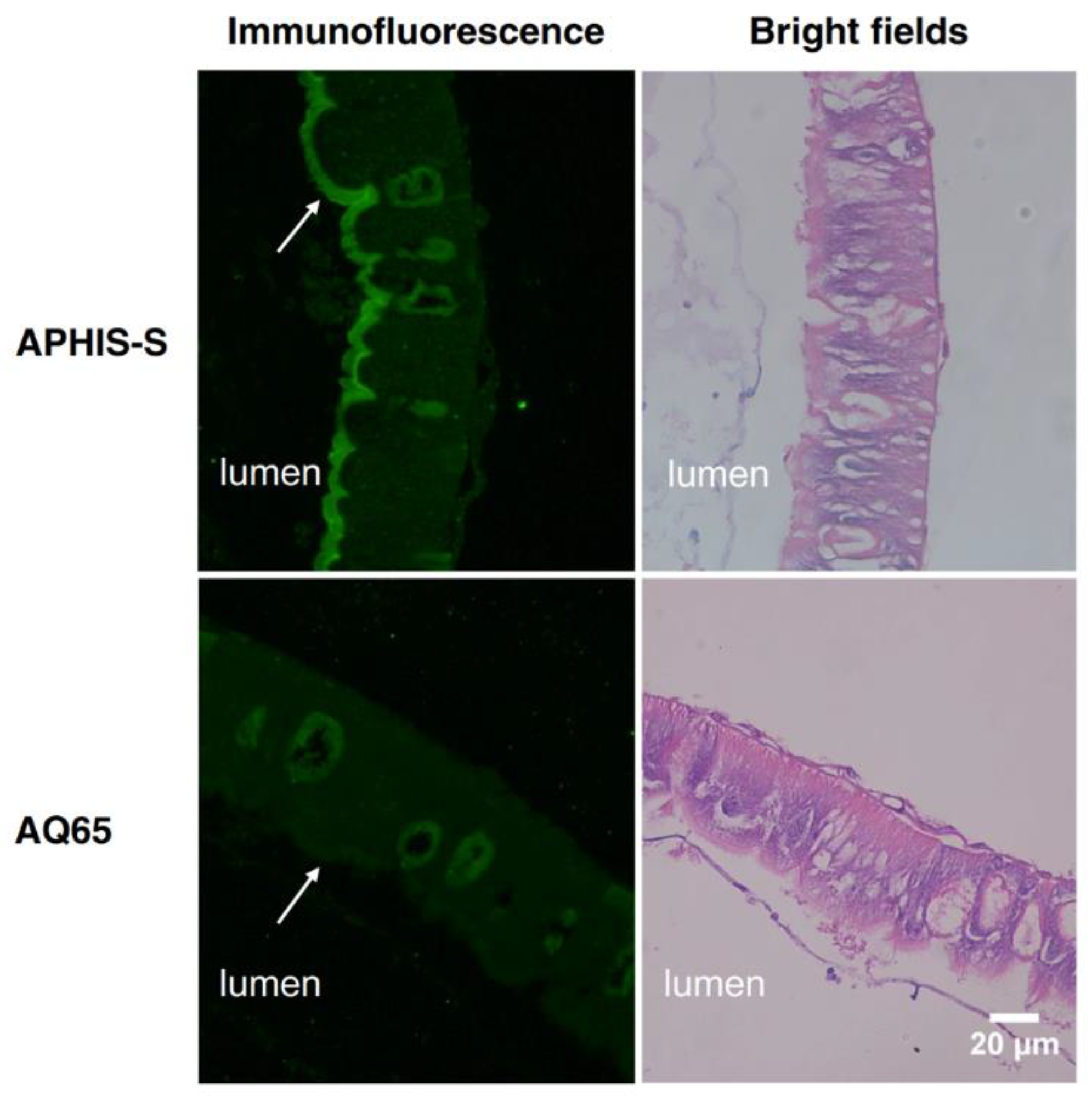
2.6. Localization of PgCad1 in APHIS-S and AQ65
3. Discussion
4. Materials and Methods
4.1. Insects and Bt Toxins
4.2. Cloning and Sequencing of PgCad1
4.3. Bioassays
4.4. Inheritance of Resistance
4.5. Genetic Linkage between r16 and Cry1Ac Resistance
4.6. Expression Vectors and Transfection of Hi5 Cells
4.7. Expression of Fusion Protein PgCad1-GFP in Hi5 Cells
4.8. Toxicity Assays on Hi5 Cells
4.9. Immunofluorescence Detection in Midgut Tissue Sections
4.10. Data Analysis
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- James, C. Global status of commercialized biotech/GM crops in 2017: Biotech crop adoption surges as economic benefits accumulate in 22 years; Brief No. 53; ISAAA: Ithaca, NY, USA, 2018. [Google Scholar]
- Romeis, J.; Naranjo, S.E.; Meissle, M.; Shelton, A.M. Genetically engineered crops help support conservation biological control. Biol. Control. 2019, 130, 136–154. [Google Scholar] [CrossRef]
- Lu, Y.; Wu, K.; Jiang, Y.; Guo, Y.; Desneux, N. Widespread adoption of Bt cotton and insecticide decrease promotes biocontrol services. Nature 2012, 487, 362–365. [Google Scholar] [CrossRef]
- Nicolia, A.; Manzo, A.; Veronesi, F.; Rosellini, D. An overview of the last 10 years of genetically engineered crop safety research. Crit. Rev. Biotechnol. 2014, 34, 77–88. [Google Scholar] [CrossRef] [PubMed]
- Dively, G.P.; Venugopal, P.D.; Bean, D.; Whalen, J.; Holmstrom, K.; Kuhar, T.P.; Doughty, H.B.; Patton, T.; Cissel, W.; Hutchison, W.D. Regional pest suppression associated with widespread Bt maize adoption benefits vegetable growers. Proc. Natl. Acad. Sci. USA 2018, 115, 3320–3325. [Google Scholar] [CrossRef] [PubMed]
- Tabashnik, B.E. ABCs of insect resistance to Bt. PLoS Genet. 2015, 11, e1005646. [Google Scholar] [CrossRef] [PubMed]
- Tay, W.T.; Mahon, R.J.; Heckel, D.G.; Walsh, T.K.; Downes, S.; James, W.J.; Lee, S.F.; Reineke, A.; Williams, A.K.; Gordon, K.H. Insect resistance to Bacillus thuringiensis toxin Cry2Ab is conferred by mutations in an ABC transporter subfamily A protein. PLoS Genet. 2015, 11, e1005534. [Google Scholar] [CrossRef] [PubMed]
- Tabashnik, B.E.; Carrière, Y. Surge in insect resistance to transgenic crops and prospects for sustainability. Nat. Biotechnol. 2017, 35, 926–935. [Google Scholar] [CrossRef] [PubMed]
- Yang, F.; Santiago González, J.C.; Williams, J.; Cook, D.C.; Gilreath, R.T.; Kerns, D.L. Occurrence and ear damage of Helicoverpa zea on transgenic Bacillus thuringiensis maize in the field in Texas, U.S. and its susceptibility to Vip3A protein. Toxins 2019, 11, 102. [Google Scholar] [CrossRef]
- Pardo-Lopez, L.; Soberón, M.; Bravo, A. Bacillus thuringiensis insecticidal three-domain Cry toxins: mode of action, insect resistance and consequences for crop protection. FEMS. Microbiol. Rev. 2013, 37, 3–22. [Google Scholar] [CrossRef]
- Wu, Y.D. Detection and mechanisms of resistance evolved in insects to Cry toxins from Bacillus thuringiensis. Adv. Insect Physiol. 2014, 47, 297–342. [Google Scholar]
- Jurat-Fuentes, J.L.; Crickmore, N. Specificity determinants for Cry insecticidal proteins: insights from their mode of action. J. Invertebr. Pathol. 2017, 142, 5–10. [Google Scholar] [CrossRef] [PubMed]
- Xiao, Y.T.; Wu, K.M. Recent progress on the interaction between insects and Bacillus thuringiensis crops. Philos. Trans. R. Soc. B 2019, 374, 20180316. [Google Scholar] [CrossRef]
- Centre for Agriculture and Bioscience International. Invasive Species Compendium. Datasheet report for Pectinophora gossypiella (pink bollworm). Available online: http://www.cabi.org/isc/datasheetreport?dsid=39417 (accessed on 31 July 2016).
- Dhurua, S.; Gujar, G.T. Field-evolved resistance to Bt toxin Cry1Ac in the pink bollworm, Pectinophora gossypiella (Saunders) (Lepidoptera: Gelechiidae), from India. Pest Manag. Sci. 2011, 67, 898–903. [Google Scholar] [CrossRef] [PubMed]
- Ojha, A.; Sree, K.S.; Sachdev, B.; Rashmi, M.A.; Ravi, K.C.; Suresh, P.J.; Mohan, K.S.; Bhatnagar, R.K. Analysis of resistance to Cry1Ac in field-collected pink bollworm, Pectinophora gossypiella (Lepidoptera: Gelechiidae), populations. GM Crops Food 2014, 5, 280–286. [Google Scholar] [CrossRef]
- Tabashnik, B.E.; Sisterson, M.S.; Ellsworth, P.C.; Dennehy, T.J.; Antilla, L.; Liesner, L.; Whitlow, M.; Staten, R.T.; Fabrick, J.A.; Unnithan, G.C.; et al. Suppressing resistance to Bt cotton with sterile insect releases. Nat. Biotechnol. 2010, 28, 1304–1307. [Google Scholar] [CrossRef] [PubMed]
- Wan, P.; Xu, D.; Cong, S.; Jiang, Y.; Huang, Y.; Wang, J.; Wu, H.; Wang, L.; Wu, K.; Carrière, Y.; et al. Hybridizing transgenic Bt cotton with non-Bt cotton counters resistance in pink bollworm. Proc. Natl. Acad. Sci. USA 2017, 114, 5413–5418. [Google Scholar] [CrossRef]
- Tabashnik, B.E.; Liu, Y.B.; Dennehy, T.J.; Sims, M.A.; Sisterson, M.S.; Biggs, R.W.; Carrière, Y. Inheritance of resistance to Bt toxin CrylAc in a field-derived strain of pink bollworm (Lepidoptera: Gelechiidae). J. Econ. Entomol. 2002, 95, 1018–1026. [Google Scholar] [CrossRef]
- Morin, S.; Biggs, R.W.; Sisterson, M.S.; Shriver, L.; Ellers-Kirk, C.; Higginson, D.; Holley, D.; Gahan, L.J.; Heckel, D.G.; Carrière, Y.; et al. Three cadherin alleles associated with resistance to Bacillus thuringiensis in pink bollworm. Proc. Natl. Acad. Sci. USA 2003, 100, 5004–5009. [Google Scholar] [CrossRef]
- Fabrick, J.A.; Tabashnik, B.E. Binding of Bacillus thuringiensis toxin Cry1Ac to multiple sites of cadherin in pink bollworm. Insect Biochem. Mol. Biol. 2007, 37, 97–106. [Google Scholar] [CrossRef]
- Fabrick, J.A.; Forlow Jech, L.; Henneberry, T.J. Novel pink bollworm resistance to the Bt toxin Cry1Ac: effects on mating, oviposition, larval development and survival. J. Insect Sci. 2009, 9, 24. [Google Scholar] [CrossRef]
- Fabrick, J.A.; Tabashnik, B.E. Similar genetic basis of resistance to Bt toxin Cry1Ac in boll-selected and diet-selected strains of pink bollworm. PloS ONE 2012, 7, e35658. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Ma, Y.; Wan, P.; Liu, K.; Xiao, Y.; Wang, J.; Cong, S.; Xu, D.; Wu, K.; Fabrick, J.A.; et al. Resistance to Bacillus thuringiensis linked with a cadherin transmembrane mutation affecting cellular trafficking in pink bollworm from China. Insect Biochem. Mol. Biol. 2018, 94, 28–35. [Google Scholar] [CrossRef]
- Fabrick, J.A.; Ponnuraj, J.; Singh, A.; Tanwar, R.K.; Unnithan, G.C.; Yelich, A.J.; Li, X.; Carrière, Y.; Tabashnik, B.E. Alternative splicing and highly variable cadherin transcripts associated with field-evolved resistance of pink bollworm to Bt cotton in India. PloS ONE 2014, 9, e97900. [Google Scholar] [CrossRef] [PubMed]
- Tabashnik, B.E.; Dennehy, T.J.; Carrière, Y. Delayed resistance to transgenic cotton in pink bollworm. Proc. Natl. Acad. Sci. USA 2005, 102, 15389–15393. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; Yu, L.; Wu, Y. Disruption of a cadherin gene associated with resistance to Cry1Ac delta-endotoxin of Bacillus thuringiensis in Helicoverpa armigera. Appl. Environ. Microb. 2005, 71, 948–954. [Google Scholar] [CrossRef] [PubMed]
- Xiao, Y.; Zhang, T.; Liu, C.; Heckel, D.G.; Li, X.; Tabashnik, B.E.; Wu, K. Mis-splicing of the ABCC2 gene linked with Bt toxin resistance in Helicoverpa armigera. Sci. Rep. 2014, 4, 6184. [Google Scholar] [CrossRef]
- Tabashnik, B.E.; Biggs, R.W.; Fabrick, J.A.; Gassmann, A.J.; Dennehy, T.J.; Carrière, Y.; Morin, S. High-level resistance to Bacillus thuringiensis toxin CrylAc and cadherin genotype in pink bollworm. J. Econ. Entomol. 2006, 99, 2125–2131. [Google Scholar] [CrossRef]
- Tabashnik, B.E.; Dennehy, T.J.; Sims, M.A.; Larkin, K.; Head, G.P.; Moar, W.J.; Carriere, Y. Control of resistant pink bollworm by transgenic cotton with Bacillus thuringiensis toxin Cry2Ab. Appl. Environ. Microbiol. 2002, 68, 3790–3794. [Google Scholar] [CrossRef]
- Gonzalez-Cabrera, J.; Escriche, B.; Tabashnik, B.E.; Ferré, J. Binding of Bacillus thuringiensis toxins in resistant and susceptible strains of pink bollworm (Pectinophora gossypiella). Insect Biochem. Mol. Biol. 2003, 33, 929–935. [Google Scholar] [CrossRef]
- Liu, Y.B.; Tabashnik, B.E.; Meyer, S.K.; Carrière, Y.; Bartlett, A.C. Genetics of pink bollworm resistance to Bacillus thuringiensis toxin Cry1Ac. J. Econ. Entomol. 2001, 94, 248–252. [Google Scholar] [CrossRef]
- Tabashnik, B.E.; Patin, A.L.; Dennehy, T.J.; Liu, Y.B.; Carrière, Y.; Sims, M.A.; Antilla, L. Frequency of resistance to Bacillus thuringiensis in field populations of pink bollworm. Proc. Natl. Acad. Sci. USA 2000, 97, 12980–12984. [Google Scholar] [CrossRef]
- Liu, Y.; Tabashnik, B.E. Inheritance of resistance to the Bacillus thuringiensis toxin Cry1C in the diamondback moth. Appl. Environ. Microbiol. 1997, 63, 2218–2223. [Google Scholar] [PubMed]
- Heckel, D.G.; Gahan, L.J.; Liu, Y.B.; Tabashnik, B.E. Genetic mapping of resistance to Bacillus thuringiensis toxins in diamondback moth using biphasic linkage analysis. Proc. Natl. Acad. Sci. USA 1999, 96, 8373–8377. [Google Scholar] [CrossRef] [PubMed]
- Xu, P.; Islam, M.; Xiao, Y.; He, F.; Li, Y.; Peng, J.; Hong, H.; Liu, C.; Liu, K. Expression of recombinant and mosaic Cry1Ac receptors from Helicoverpa armigera and their influences on the cytotoxicity of activated Cry1Ac to Spodoptera litura Sl-HP cells. Cytotechnology 2016, 68, 481–496. [Google Scholar] [CrossRef]
- Xiao, Y.; Dai, Q.; Hu, R.; Pacheco, S.; Yang, Y.; Liang, G.; Soberón, M.; Bravo, A.; Liu, K.; Wu, K. A single point mutation resulting in cadherin mis-localization underpins resistance against Bacillus thuringiensis toxin in cotton bollworm. J. Biol. Chem. 2017, 292, 2933–2943. [Google Scholar] [CrossRef] [PubMed]
- Brown, R.E.; Jarvis, K.L.; Hyland, K.J. Protein measurement using bicinchoninic acid: elimination of interfering substances. Anal Biochem. 1989, 180, 136–139. [Google Scholar] [CrossRef]
- Rabiau, N.; Dechelotte, P.; Guy, L.; Satih, S.; Bosviel, R.; Fontana, L.; Kemeny, J.L.; Boiteux, J.P.; Bignon, Y.J.; Bernard-Gallon, D. Immunohistochemical staining of mucin 1 in prostate tissues. In Vivo. 2009, 23, 203–207. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain or Cross | Slope (SE) a | LC50 (95% FL) b | RR c |
|---|---|---|---|
| APHIS-S | 3.78 (0.336) | 0.097 (0.048–0.132) | |
| AQ65 | 1.79 (0.446) | 29.5 (22.4–48.0) | 300 |
| AQ65♀ × APHIS-S♂ | 2.38 (0.203) | 0.454 (0.494–0.623) | 4.7 |
| AQ65♂ × APHIS-S♀ | 2.94 (0.278) | 0.559 (0.397–0.514) | 5.8 |
| Strain | Cotton Type | Number of Pupae | Time to Pupation (days) | Pupal wt. (mg) |
|---|---|---|---|---|
| APHIS-S | Non-Bt | 70 | 15.0 ± 0.2 a | 13.7 ± 0.4 a |
| AQ65 | Non-Bt | 85 | 17.1 ± 0.3 b | 13.8 ± 0.4 a |
| AQ65 | Bt | 47 | 20.8 ± 0.4 c | 11.6 ± 0.5 b |
| Trait | N | Bt | Non-Bt | Bt/non-Bt | |
|---|---|---|---|---|---|
| Bt | Non-Bt | ||||
| Neonate to adult survival | 270 | 230 | 0.16 | 0.35 | 0.46 |
| Proportion of females | 43 | 80 | 0.37 | 0.45 | 0.82 |
| Eggs per female | 16 | 36 | 126 ± 26 | 171 ± 22 | 0.74 |
| Hatch rate | 1345 | 1607 | 0.82 ± 0.02 | 0.79 ± 0.03 | 1.04 |
| Net reproductive rate a | 6.1 | 21.3 | 0.29 | ||
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, L.; Ma, Y.; Guo, X.; Wan, P.; Liu, K.; Cong, S.; Wang, J.; Xu, D.; Xiao, Y.; Li, X.; et al. Pink Bollworm Resistance to Bt Toxin Cry1Ac Associated with an Insertion in Cadherin Exon 20. Toxins 2019, 11, 186. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins11040186
Wang L, Ma Y, Guo X, Wan P, Liu K, Cong S, Wang J, Xu D, Xiao Y, Li X, et al. Pink Bollworm Resistance to Bt Toxin Cry1Ac Associated with an Insertion in Cadherin Exon 20. Toxins. 2019; 11(4):186. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins11040186
Chicago/Turabian StyleWang, Ling, Yuemin Ma, Xueqin Guo, Peng Wan, Kaiyu Liu, Shengbo Cong, Jintao Wang, Dong Xu, Yutao Xiao, Xianchun Li, and et al. 2019. "Pink Bollworm Resistance to Bt Toxin Cry1Ac Associated with an Insertion in Cadherin Exon 20" Toxins 11, no. 4: 186. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins11040186