Monitoring of Fusarium Species and Trichothecene Genotypes Associated with Fusarium Head Blight on Wheat in Hebei Province, China

 ,
,
Abstract
:1. Introduction
2. Results
2.1. Fusarium Species Determination
2.2. Prediction of Trichothecene Genotype
3. Discussion
4. Materials and Methods
4.1. Sampling
4.2. Isolation and Morphological Identification
4.3. DNA Extraction
4.4. Species Determination and Trichothecene Genotype Prediction
Author Contributions
Funding
Conflicts of Interest
References
- McMullen, M.P.; Jones, R.; Gallenberg, D. Scab of wheat and barley: A re-emerging disease of devastating impact. Plant Dis. 1997, 81, 1340–1348. [Google Scholar] [CrossRef] [PubMed]
- Seitz, L.M.; Eustace, W.D.; Mohr, H.E.; Shogren, M.D.; Yamazaki, W.T. Cleaning, milling, and baking tests with hard red winter wheat containing deoxynivalenol. Cereal Chem. 1986, 63, 146–150. [Google Scholar]
- Yao, J.B.; Lu, W.Z. Research advances in wheat breeding for scab resistance in China. Jiangsu J. Agric. Sci. 2000, 16, 242–248. [Google Scholar]
- Burgess, L.W.; Klein, T.A.; Bryden, W.L.; Tobin, N.F. Head blight of wheat caused by Fusarium graminearum Group1 in New South Wales in 1983. Australas. Plant Path. 1987, 16, 72–78. [Google Scholar] [CrossRef]
- Boshof, W.H.P.; Swart, W.J.; Pretorius, Z.A. Isozyme characterisation of Fusarium graminearum isolates associated with head blight of irrigated wheat in South Africa. S. Afr. J. Bot. 1999, 65, 281–286. [Google Scholar] [CrossRef]
- Stack, R.W. Return of an Old Problem: Fusarium Head Blight of Small Grains. APSnet Feature. APSnet Plant Pathology Online. 1999. Available online: https://www.apsnet.org/publications/apsnetfeatures/Pages/headblight.aspx (accessed on 28 January 2019).
- Windels, C.E. Economic and social impacts of Fusarium head blight: Changing farms and rural communities in the Northern Great Plains. Phytopathology 2000, 90, 17–21. [Google Scholar] [CrossRef]
- Song, Y.L.; He, W.L.; Yang, G.Q.; Liu, H.Y. The prevalence of Fusarium head blight in Henan Province and the counter measures. J. Henan Agric. Sci. 2000, 4, 14. [Google Scholar]
- Wang, H.J.; Gou, J.J.; Xue, Y.; Lu, S.Z.; Li, J.X.; Hu, X.Z.; Wu, C.L.; Chen, H.Y. Discuss the integrated control strategy of wheat pests according to the occurrence of Fusarium head blight in Hebei province in 2003. Res. Plant Pathol. Hebei 2003, 1, 7–9. [Google Scholar]
- Goswami, R.S.; Kistler, H.C. Heading for disaster: Fusarium graminearum on cereal crops. Mol. Plant Pathol. 2004, 5, 515–525. [Google Scholar] [CrossRef]
- Osborne, L.E.; Stein, J.M. Epidemiology of Fusarium head blight on small-grain cereals. Int. J. Food Microbiol. 2007, 119, 103–108. [Google Scholar] [CrossRef] [PubMed]
- Parry, D.W.; Jenkinson, P.; McLeod, L. Fusarium ear blight (scab) in small grain cereals-a review. Plant Pathol. 1995, 44, 207–238. [Google Scholar] [CrossRef]
- Jennings, P.; Coates, M.E.; Walsh, K.; Turner, J.A.; Nicholson, P. Determination of deoxynivalenol-and nivalenol-producing chemotypes of Fusarium graminearum isolated from wheat crops in England and Wales. Plant Pathol. 2004, 53, 643–652. [Google Scholar] [CrossRef]
- Sarver, B.A.; Ward, T.J.; Gale, L.R.; Broz, K.; Corby Kistler, H.; Aoki, T.; Nicholson, P.; Carter, J.; O’Donnell, K. Novel Fusarium head blight pathogens from Nepal and Louisiana revealed by multilocus genealogical concordance. Fungal Genet. Biol. 2011, 48, 1096–1107. [Google Scholar] [CrossRef] [PubMed]
- O’Donnell, K.; Ward, T.J.; Aberra, D.; Kistler, H.C.; Aoki, T.; Orwig, N.; Kimura, M.; Bjornstad, S.; Klemsdal, S.S. Multilocus genotyping and molecular phylogenetics resolve a novel head blight pathogen within the Fusarium graminearum species complex from Ethiopia. Fungal Genet. Biol. 2008, 45, 1514–1522. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Van der Lee, T.; Waalwijk, C.; Chen, W.Q.; Xu, J.; Xu, J.S.; Zhang, Y.; Feng, J. Population analysis of the Fusarium graminearum species complex from wheat in China show a shift to more aggressive isolates. PLoS ONE 2012, 7, e31722. [Google Scholar] [CrossRef]
- Suga, H.; Karugia, G.W.; Ward, T.; Gale, L.R.; Tomimura, K.; Nakajima, T.; Miyasaka, A.; Koizumi, S.; Kageyama, K.; Hyakumachi, M. Molecular characterization of the Fusarium graminearum species complex in Japan. Phytopathology 2008, 98, 159–166. [Google Scholar] [CrossRef]
- Karugia, G.W.; Suga, H.; Gzle, L.R.; Nakajima, T.; Ueda, A.; Hyakumachi, M. Population structure of Fusarium asiaticum from two Japanese regions and eastern China. J. Gen. Plant Pathol. 2009, 75, 110–118. [Google Scholar] [CrossRef]
- Ueda, A.; Nishimoto, H.; Kato, N.; Hirano, T.; Fukaya, M. Lineages and trichothecene mycotoxin types of Fusarium head blight pathogens of wheat and barley in Todai district. Res. Bull. Aichi. Agric. Res. Ctr. 2007, 39, 17–23. [Google Scholar]
- Lu, W.Z.; Cheng, S.H.; Wang, Y.Z. Fusarium Head Blight Research; Science Press: Beijing, China, 2001. [Google Scholar]
- Zhang, J.B.; Li, H.P.; Dang, F.J.; Qu, B.; Xu, Y.B.; Zhao, C.S.; Liao, Y.C. Determination of the trichothecene mycotoxin chemotypes and associated geographical distribution and phylogenetic species of the Fusarium graminearum clade from China. Mycol. Res. 2007, 111, 967–975. [Google Scholar] [CrossRef]
- Qu, B.; Li, H.P.; Zhang, J.B.; Xu, Y.B.; Huang, T.; Wu, A.B.; Zhao, C.S.; Carter, J.; Nicholson, P.; Liao, Y.C. Geographic distribution and genetic diversity of Fusarium graminearum and F. asiaticum on wheat spikes throughout China. Plant Pathol. 2008, 57, 15–24. [Google Scholar] [CrossRef]
- Hao, J.J.; Xie, S.N.; Sun, J.; Yang, G.Q.; Liu, J.Z.; Xu, F.; Ru, Y.Y.; Song, Y.L. Analysis of Fusarium graminearum species complex from wheat-maize rotation regions in Henan (China). Plant Dis. 2017, 101, 720–725. [Google Scholar] [CrossRef]
- Hu, L.F.; Zhang, L.F. Analysis and comparison of livestock breeding in the north with straws from the south with straw return to field. J. Agric. Univ. Hebei 2003, 26, 12–14. [Google Scholar]
- Ji, L.J.; Kong, L.X.; Li, Q.S.; Wang, L.S.; Chen, D.; Ma, P. First report of Fusarium pseudograminearum causing Fusarium head blight of wheat in Hebei Province, China. Plant Dis. 2016, 100, 220. [Google Scholar] [CrossRef]
- Nicholson, P.; Simpson, D.R.; Wilson, A.H.; Chandler, E.; Thomsett, M. Detection and differentiation of trichothecene and enniatin-producing Fusarium species on small-grain cereals. Eur. J. Plant Pathol. 2004, 110, 503–514. [Google Scholar] [CrossRef]
- Langseth, W.; Bernhoft, A.; Rundberget, T.; Kosiak, B.; Gareis, M. Mycotoxin production and cytotoxicity of Fusarium strains isolated from Norwegian cereals. Mycopathologia 1999, 144, 103–113. [Google Scholar] [CrossRef]
- Doohan, F.M.; Brennan, J.; Cooke, B.M. Influence of climatic factors on Fusarium species pathogenic to cereals. Eur. J. Plant Pathol. 2003, 109, 755–768. [Google Scholar] [CrossRef]
- Obst, A.; Günther, B.; Beck, R.; Lepschy, J.; Tischner, H. Weather conditions conductive to Gibberella zeae and Fusarium graminearum head blight of wheat. J. Appl. Genet. A 2002, 43, 185–192. [Google Scholar]
- Maldonado-Ramirez, S.L.; Schmale III, D.G.; Shields, E.J.; Bergstrom, G.C. The relative abundance of viable spores of Gibberella zeae in the planetary boundary layer suggests the role of long-distance transport in regional epidemics of Fusarium head blight. Agric. For. Meteorol. 2005, 132, 20–27. [Google Scholar] [CrossRef]
- Schmale, D.G.; Leslie, J.F.; Zeller, K.A.; Saleh, A.A.; Shields, E.J.; Bergstrom, G.C. Genetic structure of atmospheric populations of Gibberella zeae. Phytopathology 2006, 96, 1021–1026. [Google Scholar] [CrossRef]
- De Galich, M.T.V. Fusarium head blight in Argentina. In Fusarium Head Scab: Global Status and Future Prospects, Proceedings of a Workshop Held at CIMMYT, EI Batan, Mexico, 13–17 October 1996; Dubin, H.J., Gilchrist, L., Reeves, J., McNab, A., Eds.; D.F.: EI Batan, Mexico, 1997; pp. 19–28. [Google Scholar]
- Muthomi, J.W.; Ndung’u, J.K.; Gathumbi, J.K.; Mutitu, E.W.; Wagacha, J.M. The occurrence of Fusarium species and mycotoxins in Kenyan wheat. Crop Prot. 2008, 27, 1215–1219. [Google Scholar] [CrossRef]
- Lee, J.; Chang, I.Y.; Kim, H.; Yun, S.H.; Leslie, J.F.; Lee, Y.W. Genetic diversity and fitness of Fusarium graminearum populations from rice in Korea. Appl. Environ. Microbiol. 2009, 75, 3289–3295. [Google Scholar] [CrossRef] [PubMed]
- Del Ponte, E.M.; Spolti, P.; Ward, T.J.; Gomes, L.B.; Nicolli, C.P.; Kuhnem, P.R.; Silva, C.N.; Tessmann, D.J. Regional and field-specific factors affect the composition of Fusarium head blight pathogens in subtropical no-till wheat agro ecosystem of Brazil. Phytopathology 2015, 105, 246–254. [Google Scholar] [CrossRef]
- Gomes, L.B.; Ward, T.J.; Badiale-Furlong, E.; Del Ponte, E.M. Species composition, toxigenic potential and pathogenicity of Fusarium graminearum species complex isolates from southern Brazilian rice. Plant Pathol. 2015, 64, 980–987. [Google Scholar] [CrossRef]
- Ji, L.J.; Kong, L.X.; Wang, Y.J.; Li, Q.S. Occurrence and development of wheat crown rot in Hebei province, China. In Paddock to Plates. In Proceedings of the 10th Australasian Soilborne Disease Symposium, Adelaide, Australia, 4–8 September 2018; p. 00136. [Google Scholar]
- Southwell, R.J.; Moore, K.J.; Manning, W.; Hayman, P.T. An outbreak of Fusarium head blight of durum wheat on the Liverpool Plains in northern New South Wales in 1999. Australas. Plant Path. 2003, 32, 465–471. [Google Scholar] [CrossRef]
- Akinsanmi, O.A.; Mitter, V.; Simpfendorfer, S.; Backhouse, D.; Chakraborty, S. Identity and pathogenicity of Fusarium spp. isolated from wheat fields in Queensland and northern New South Wales. Aust. J. Agric. Res. 2004, 55, 97–107. [Google Scholar] [CrossRef]
- Clement, J.A.; Parry, D.W. Stem-base disease and fungal colonization of winter wheat grown in compost inoculated with Fusarium culmorum, F. graminearum and Microdochium nivale. Eur. J. Plant Pathol. 1998, 104, 323–330. [Google Scholar] [CrossRef]
- Tan, M.K.; Simpfendorfer, S.; Backhouse, D.; Murray, G.M. Occurrence of Fusarium head blight (FHB) in southern NSW in 2000: Identification of causal fungi and determination of putative chemotype of Fusarium graminearum isolates by PCR. Australas. Plant Path. 2004, 33, 385–392. [Google Scholar] [CrossRef]
- Xu, X.M.; Nicholson, P. Community ecology of fungal pathogens causing wheat head blight. Annu. Rev. Phytopathol. 2009, 47, 83–103. [Google Scholar] [CrossRef]
- Champeil, A.; Dore, T.; Fourbet, J.F. Fusarium head blight: Epidemiological origin of the effects of cultural practices on head blight attacks and the production of mycotoxins by Fusarium in wheat grains. Plant Sci. 2004, 166, 1389–1415. [Google Scholar] [CrossRef]
- Dill-Macky, R.; Jones, R.K. The effect of previous crop residues and tillage in Fusarium head blight of wheat. Plant Dis. 2000, 84, 71–76. [Google Scholar] [CrossRef]
- Jennings, P.; Coates, M.E.; Turner, J.A.; Chandler, E.A.; Nicholson, P. Determination of deoxynivalenol and nivalenol chemotypes of Fusarium culmorum isolates from England and Wales by PCR assays. Plant Pathol. 2004, 53, 182–190. [Google Scholar] [CrossRef]
- Waalwijk, C.; Kastelein, P.; Vries, I.; Kerényi, Z.; van der Lee, T.; Hesselink, T.; Köhl, J.; Kema, G. Major changes in Fusarium spp. in wheat in the Netherlands. Eur. J. Plant Pathol. 2003, 109, 743–754. [Google Scholar] [CrossRef]
- Nielsen, L.K.; Jensen, J.D.; Nielsen, G.C.; Jensen, J.E.; Spliid, N.H.; Thomsen, I.K.; Justesen, A.F.; Collinge, D.B.; Jørgensen, L.N. Fusarium head blight of cereals in Denmark: Species complex and related mycotoxins. Phytopathology 2011, 101, 960–969. [Google Scholar] [CrossRef]
- Stępień, L.; Chełkowski, J. Fusarium head blight of wheat: Pathogenic species and their mycotoxins. World Mycotoxin J. 2010, 3, 107–119. [Google Scholar] [CrossRef]
- Nash, S.M.; Snyder, W.C. Quantitative estimations by plate counts of propagules of the bean root rot Fusarium in field soils. Phytopathology 1962, 52, 567–572. [Google Scholar]
- Burgess, L.W.; Summerell, B.A.; Bullock, S.; Gott, K.P.; Backhouse, D. Laboratory manual for Fusarium research, 3rd ed.; University of Sydney/Royal Botanic Gardens: Sydney, Australia, 1994. [Google Scholar]
- Fisher, N.L.; Burgess, L.W.; Toussoun, T.A.; Nelson, P.E. Carnation leaves as a substrate and for preserving cultures of Fusarium species. Phytopathology 1982, 72, 151–153. [Google Scholar] [CrossRef]
- Leslie, J.F.; Summerell, B.A. The Fusarium Laboratory Manual; Blackwell Publishing Ltd.: Oxford, UK, 2006. [Google Scholar]
- Carter, J.P.; Rezanoor, H.N.; Holden, D.; Desjardins, A.E.; Plattner, R.D.; Nicholson, P. Variation in pathogenicity associated with the genetic diversity of Fusarium graminearum. Eur. J. Plant Pathol. 2002, 108, 573–583. [Google Scholar] [CrossRef]
- O’Donnell, K.; Ward, T.J.; Geiser, D.M.; Kistler, H.C.; Aoki, T. Genealogical concordance between the mating type locus and seven other nuclear genes supports formal recognition of nine phylogenetically distinct species within the Fusarium graminearum clade. Fungal Genet. Biol. 2004, 41, 600–623. [Google Scholar] [CrossRef]
- Geiser, D.M.; del Mar Jiménez-Gasco, M.; Kang, S.; Makalowska, I.; Veeraraghavan, N.; Ward, T.J.; Zhang, N.; Kuldau, G.A.; O’Donnell, K. FUSARIUM-ID v. 1.0: A DNA sequence database for identifying Fusarium. Eur. J. Plant Pathol. 2004, 110, 473–479. [Google Scholar] [CrossRef]
- Nicholson, P.; Simpson, D.R.; Weston, G.; Rezanoor, H.N.; Lees, A.K.; Parry, D.W.; Joyce, D. Detection and quantification of Fusarium culmorum and Fusarium graminearum in cereals using PCR assays. Physiol. Mol. Plant Pathol. 1998, 53, 17–37. [Google Scholar] [CrossRef]
- Demeke, T.; Clear, R.M.; Patrick, S.K.; Gaba, D. Species-specific PCR-based assays for the detection of Fusarium species and a comparison with the whole seed agar plate method and trichothecene analysis. Int. J. Food Microbiol. 2005, 103, 271–284. [Google Scholar] [CrossRef]
- O’Donnell, K.; Kistler, H.C.; Cigelnik, E.; Ploetz, R.C. Multiple evolutionary origins of the fungus causing Panama disease of banana: Concordant evidence from nuclear and mitochondrial gene genealogies. Proc. Natl. Acad. Sci. USA 1998, 95, 2044–2049. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ward, T.J.; Clear, R.M.; Rooney, A.P.; O’Donnell, K.; Gaba, D.; Patrick, S.; Starkey, D.E.; Gilbert, J.; Geiser, D.M.; Nowicki, T.W. An adaptive evolutionary shift in Fusarium head blight pathogen populations is driving the rapid spread of more toxigenic Fusarium graminearum in North America. Fungal Genet. Biol. 2008, 45, 473–484. [Google Scholar] [CrossRef] [PubMed]
- Audenaert, K.; Van Broeck, R.; Bekaert, B.; De Witte, F.; Heremans, B.; Messens, K.; Höfte, M.; Haesaert, G. Fusarium head blight (FHB) in Flanders: Population diversity, inter-species associations and DON contamination in commercial winter wheat varieties. Eur. J. Plant Pathol. 2009, 125, 445–458. [Google Scholar] [CrossRef]


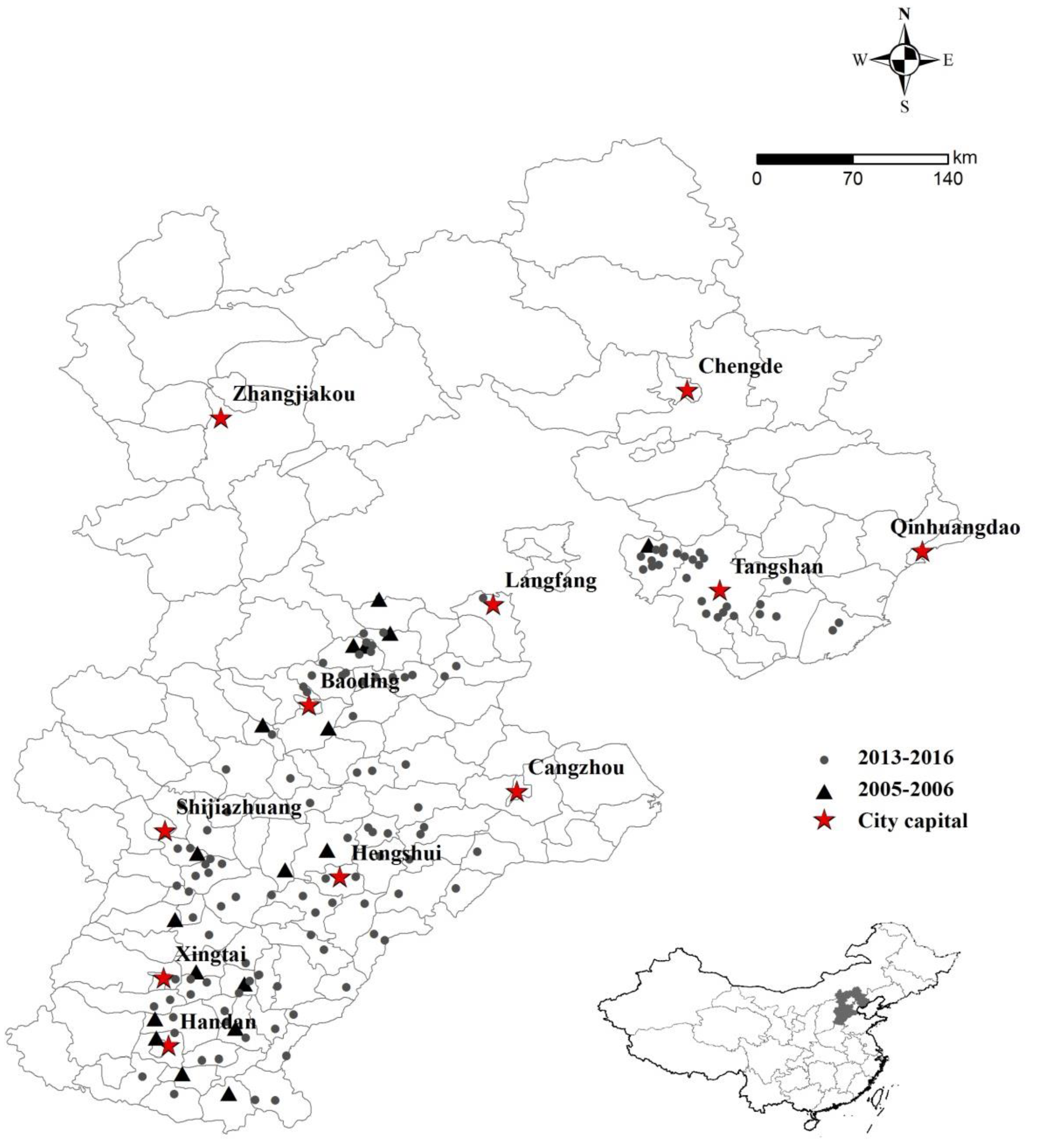{kind=link}
| Sampling Sites | Isolates Obtained | Fg a | Fc a | Fe a | Fv a | Fp a | Fs a | Fch a |
|---|---|---|---|---|---|---|---|---|
| Handan | 280 | 274(97.9%) b | 2(0.7%) | 2(0.7%) | 0 | 0 | 0 | 2(0.7%) |
| Xingtai | 87 | 75(86.2%) | 0 | 0 | 5(5.7%) | 3(3.4%) | 0 | 4(4.6%) |
| Hengshui | 15 | 15(100.0%) | 0 | 0 | 0 | 0 | 0 | 0 |
| Shijiazhuang | 36 | 35(97.2%) | 0 | 1(2.8%) | 0 | 0 | 0 | 0 |
| Baoding | 349 | 342(98.0%) | 0 | 0 | 1(0.3%) | 3(0.9%) | 1(0.3%) | 2(0.6%) |
| Tangshan | 11 | 11(100.0%) | 0 | 0 | 0 | 0 | 0 | 0 |
| Total | 778 | 752(96.7%) | 2(0.3%) | 3(0.4%) | 6(0.8%) | 6(0.8%) | 1(0.1%) | 8(1.0%) |
| Sampling Sites | Isolates Obtained | Fg a | Fa a | Fp a | Fc a | Fn a |
|---|---|---|---|---|---|---|
| Handan | 175 | 172(98.3%) b | 0 | 1(0.6%) | 2(1.1%) | 0 |
| Xingtai | 156 | 153(98.1%) | 1(0.6%) | 2(1.3%) | 0 | 0 |
| Hengshui | 132 | 129(97.7%) | 0 | 3(2.3%) | 0 | 0 |
| Cangzhou | 18 | 16(88.9%) | 0 | 2(11.1%) | 0 | 0 |
| Shijiazhuang | 153 | 148(96.7%) | 0 | 5(3.3%) | 0 | 0 |
| Baoding | 157 | 157(100.0%) | 0 | 0 | 0 | 0 |
| Langfang | 77 | 77(100.0%) | 0 | 0 | 0 | 0 |
| Tangshan | 134 | 126(94.0%) | 6(4.5%) | 1(0.7%) | 0 | 1(0.7%) |
| Total | 1002 | 978(97.6%) | 7(0.7%) | 14(1.4%) | 2(0.2%) | 1(0.1%) |
| Sampling Sites | 2005–2006 | 2013–2016 | |||||
|---|---|---|---|---|---|---|---|
| Fg a | Total | Fg a | Fg a | Fa a | Fp a | Total | |
| 15-ADON | 15-ADON | 3-ADON | NIV | 3-ADON | |||
| Handan | 274 | 274 | 171 | 1 | 0 | 1 | 173 |
| Xingtai | 75 | 75 | 150 | 3 | 1 | 2 | 156 |
| Hengshui | 15 | 15 | 129 | 0 | 0 | 3 | 132 |
| Cangzhou | - | - | 16 | 0 | 0 | 2 | 18 |
| Shijiazhuang | 35 | 35 | 145 | 3 | 0 | 5 | 153 |
| Baoding | 342 | 342 | 157 | 0 | 0 | 0 | 157 |
| Langfang | - | - | 77 | 0 | 0 | 0 | 77 |
| Tangshan | 11 | 11 | 126 | 0 | 6 | 1 | 133 |
| Total | 752 | 752 | 971 | 7 | 7 | 14 | 999 |
| Primers | Sequence | Size (bp) | Annealing Temperature °C | Reference |
|---|---|---|---|---|
| Fg16F | CTCCGGATATGTTGCGTCAA | 400–500 | 57 | [56] |
| Fg16R | GGTAGGTATCCGACATGGCAA | |||
| Fp1-1 | CGGGGTAGTTTCACATTTCYG | 523 | 57 | [57] |
| Fp1-2 | GAGAATGTGATGASGACAATA | |||
| Fc01F | ATGGTGAACTCGTCGTGGC | 570 | 62 | [56] |
| Fc01R | CCCTTCTTACGCCAATCTCG | |||
| EF-1α F | ATGGGTAAGGA(AG)GACAAGAC | 700 | 56 | [58] |
| EF-1α R | GGA(GA)GTACCAGT(GC)ATCATGTT | |||
| 3CON | TGGCAAAGACTGGTTCAC | 840 or 610 or 243 | 52 | [59] |
| 3NA | GTGCACAGAATATACGAGC | |||
| 3D15A | ACTGACCCAAGCTGCCATC | |||
| 3D3A | CGCATTGGCTAACACATG | |||
| Tri13F | TACGTGAAACATTGTTGGC | 234 or 415 | 57 | [60] |
| Tri13R | GGTGTCCCAGGATCTGCG | |||
| Tri303F | GATGGCCGCAAGTGGA | 583 | 52 | [60] |
| Tri303R | GCCGGACTGCCCTATTG | |||
| Tri315F | CTCGCTGAAGTTGGACGTAA | 863 | 61 | [60] |
| Tri315R | GTCTATGCTCTCAACGGACAAC |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ji, L.; Li, Q.; Wang, Y.; Burgess, L.W.; Sun, M.; Cao, K.; Kong, L. Monitoring of Fusarium Species and Trichothecene Genotypes Associated with Fusarium Head Blight on Wheat in Hebei Province, China. Toxins 2019, 11, 243. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins11050243
Ji L, Li Q, Wang Y, Burgess LW, Sun M, Cao K, Kong L. Monitoring of Fusarium Species and Trichothecene Genotypes Associated with Fusarium Head Blight on Wheat in Hebei Province, China. Toxins. 2019; 11(5):243. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins11050243
Chicago/Turabian StyleJi, Lijing, Qiusheng Li, Yajiao Wang, Lester W Burgess, Mengwei Sun, Keqiang Cao, and Lingxiao Kong. 2019. "Monitoring of Fusarium Species and Trichothecene Genotypes Associated with Fusarium Head Blight on Wheat in Hebei Province, China" Toxins 11, no. 5: 243. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins11050243