Dissecting Toxicity: The Venom Gland Transcriptome and the Venom Proteome of the Highly Venomous Scorpion Centruroides limpidus (Karsch, 1879)

 , and
, and 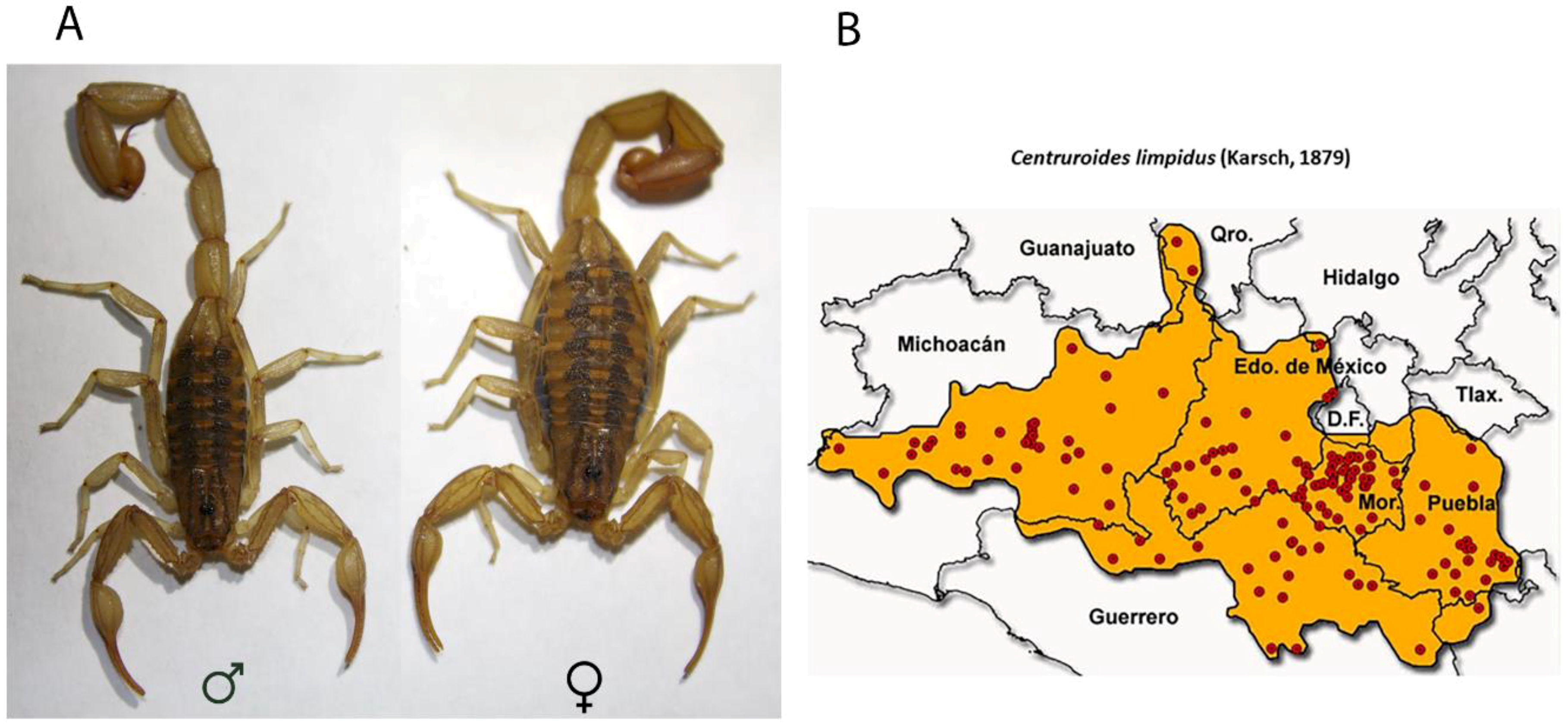{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
2.1. RNA Isolation, Sequencing, and Assembly
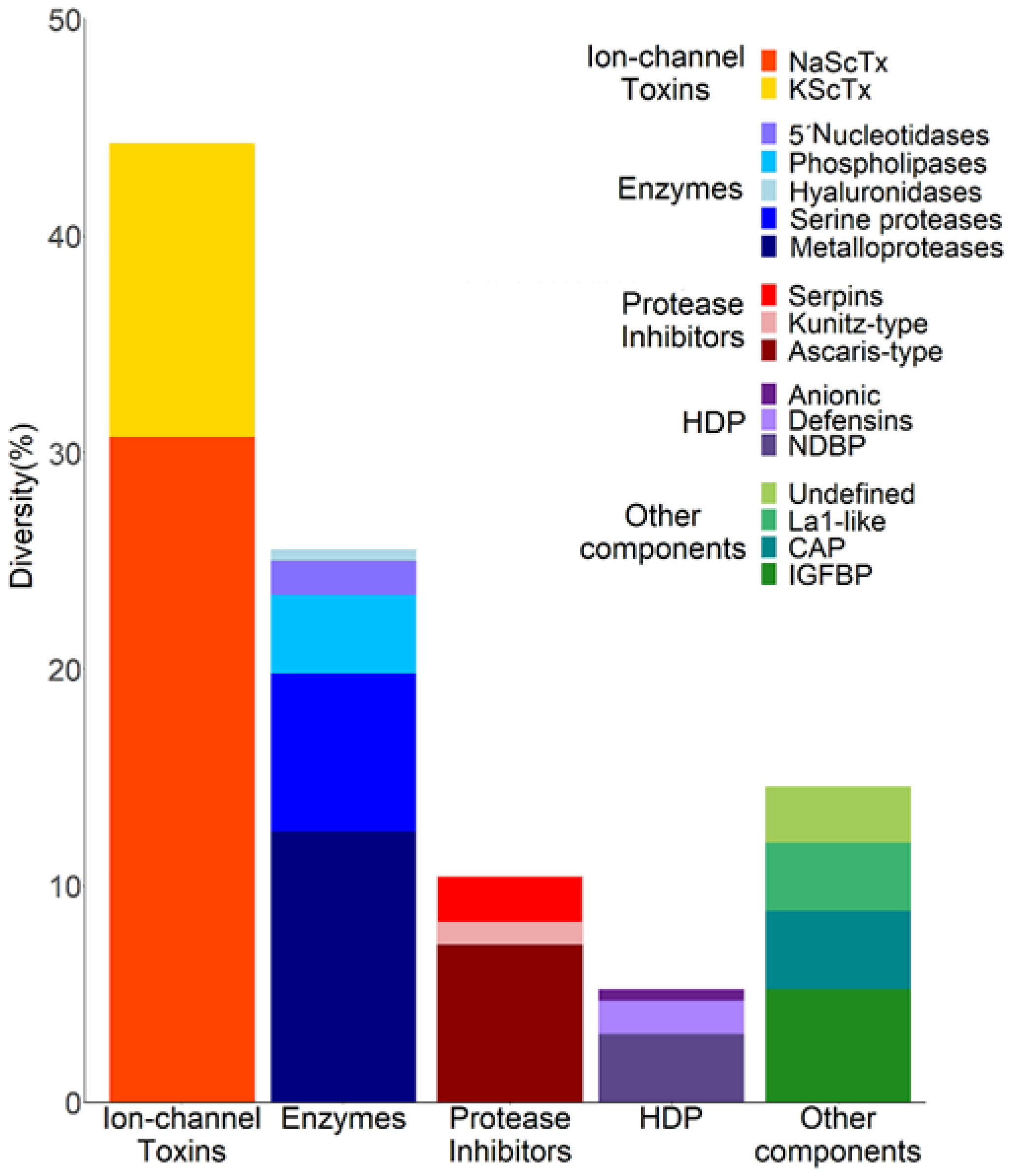
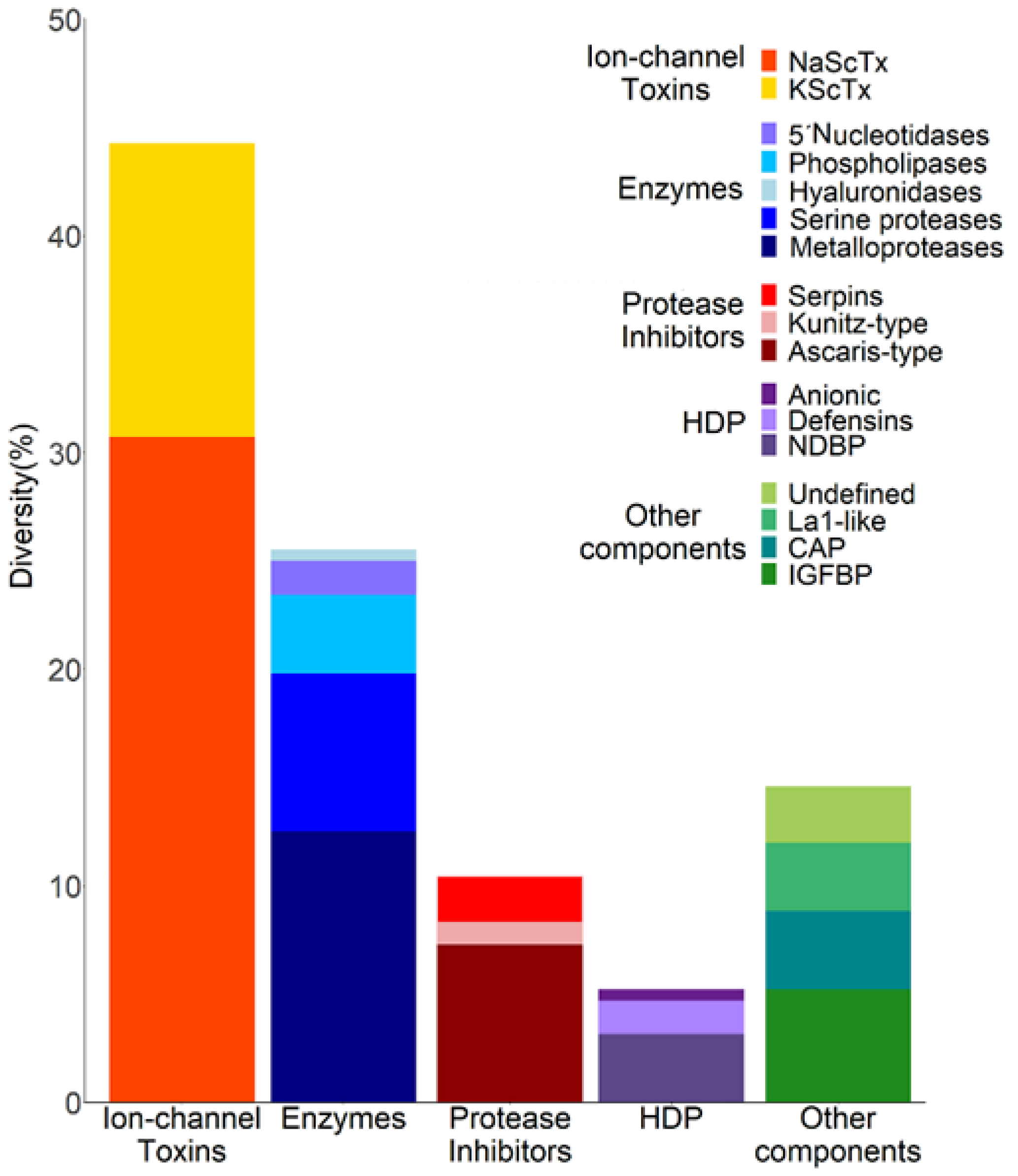
2.2. The Diversity of Transcripts Related to Venom Components in the Venom Gland of C. limpidus
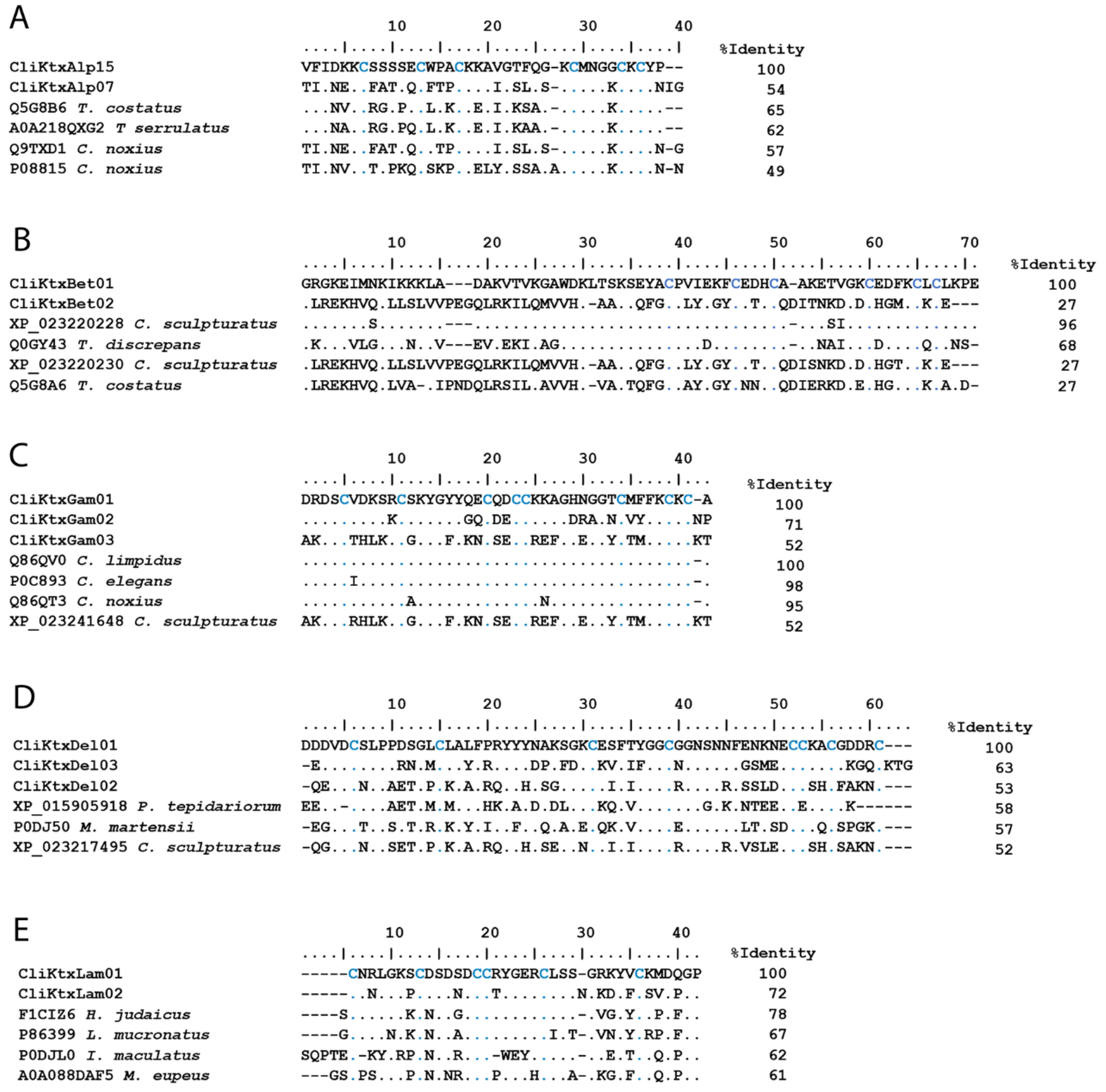
2.2.1. Ion Channel-Acting Toxins
Toxins Active on Voltage-Gated Sodium Channels (NaScTx)
Toxins Active on Potassium Channels (KScTx)
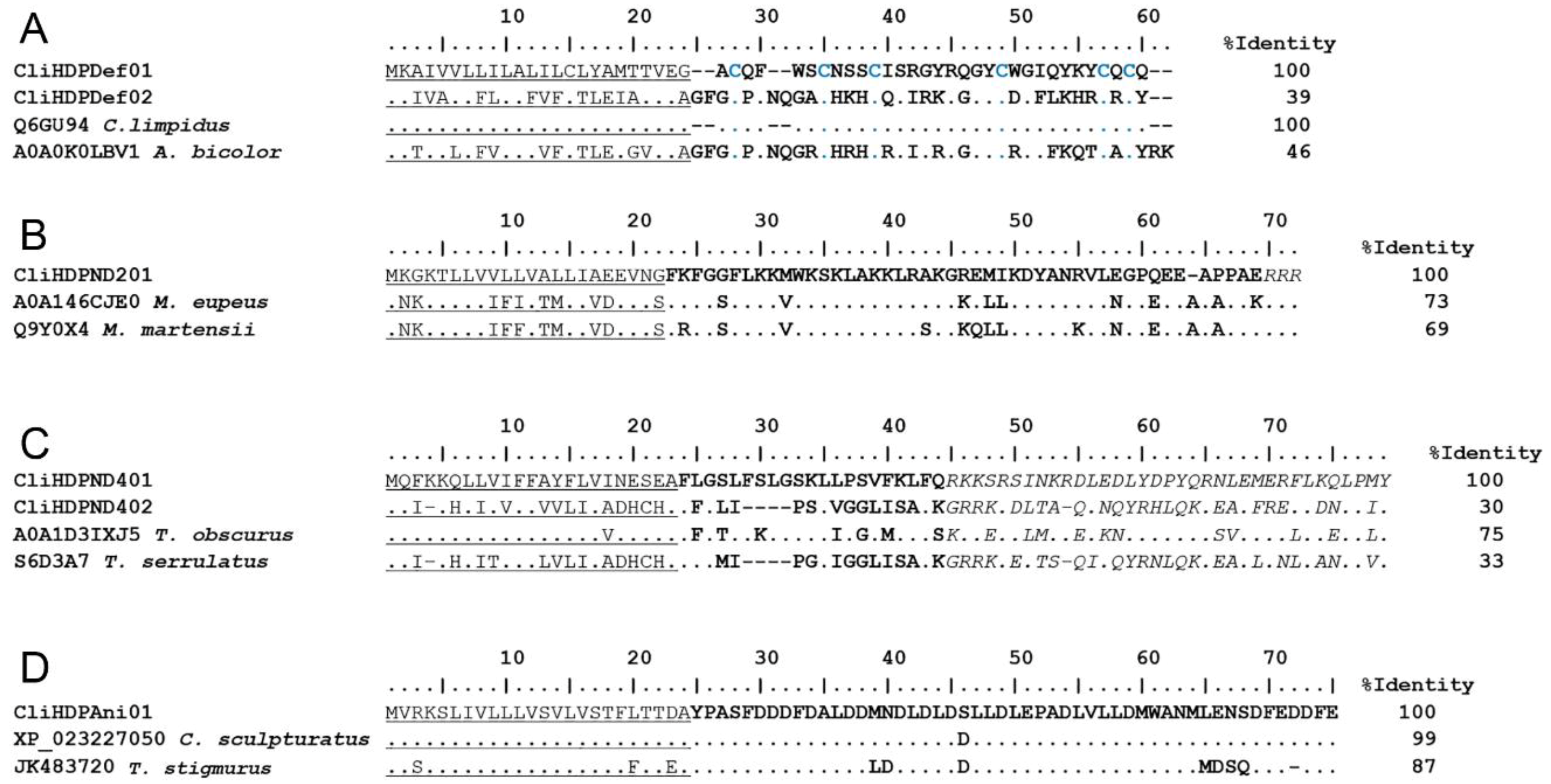
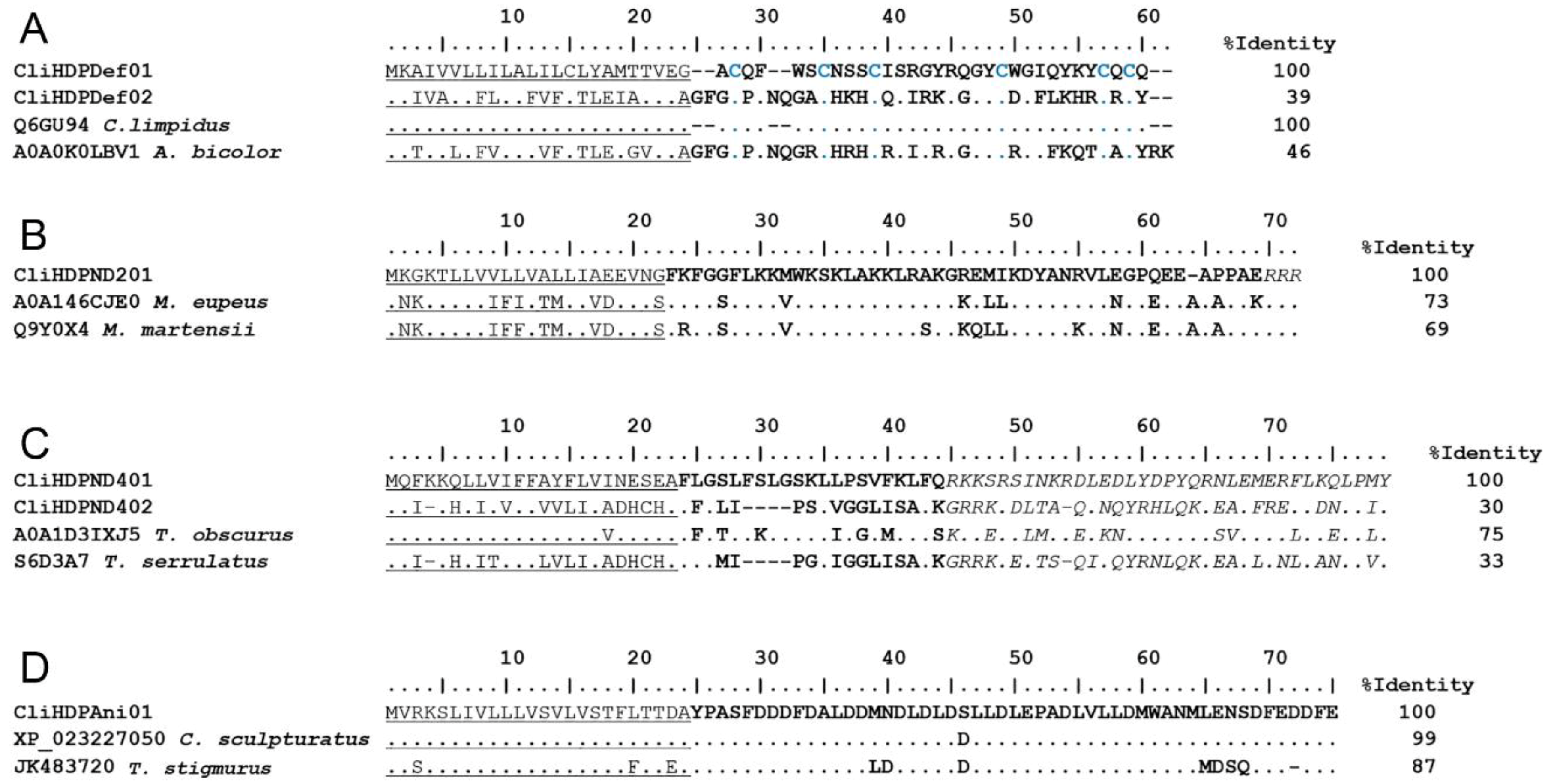
2.2.2. Host Defense Peptides (HDPs)
2.2.3. Enzymes
2.2.4. Other Venom Components
2.3. Proteomic Exploration of the Venom Components of C. limpidus
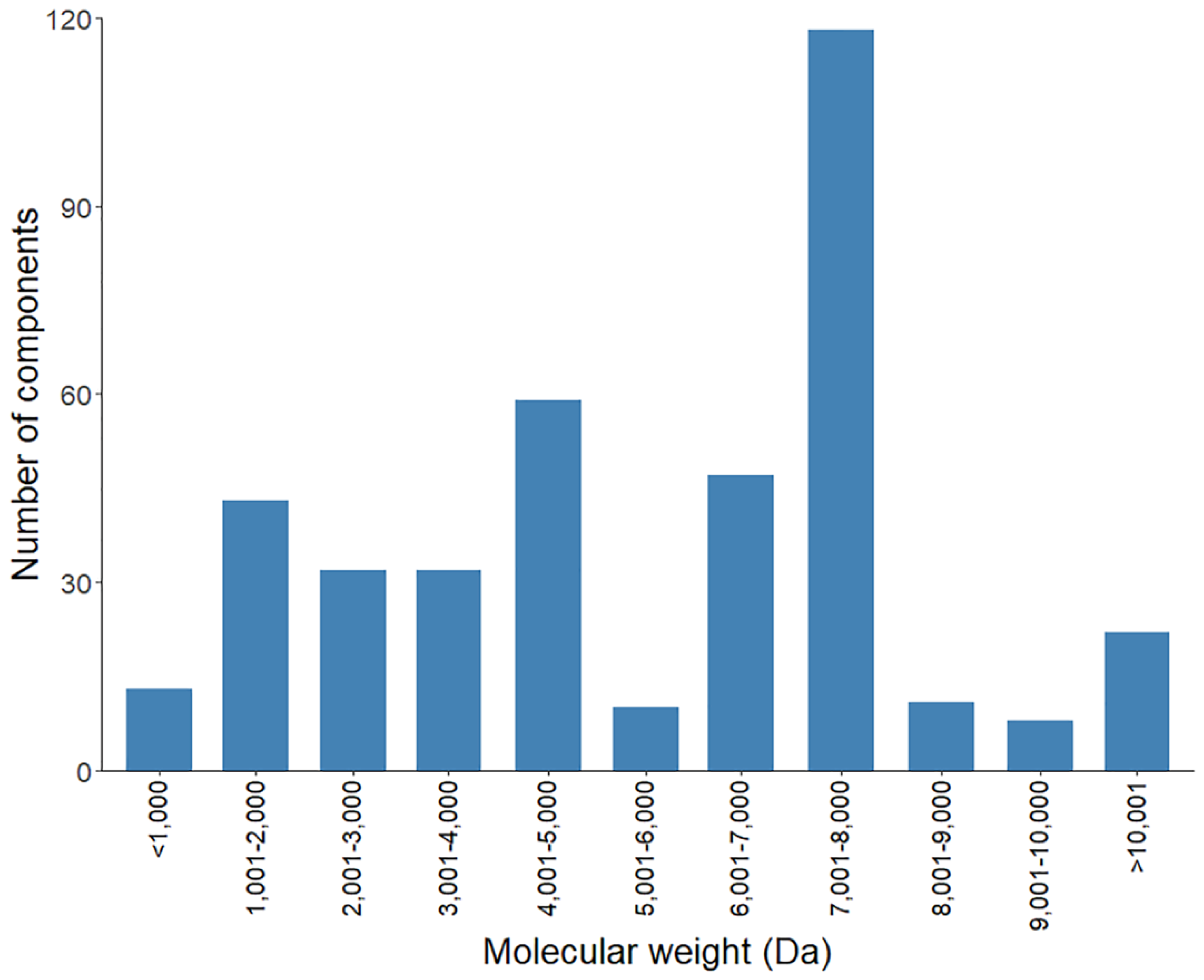
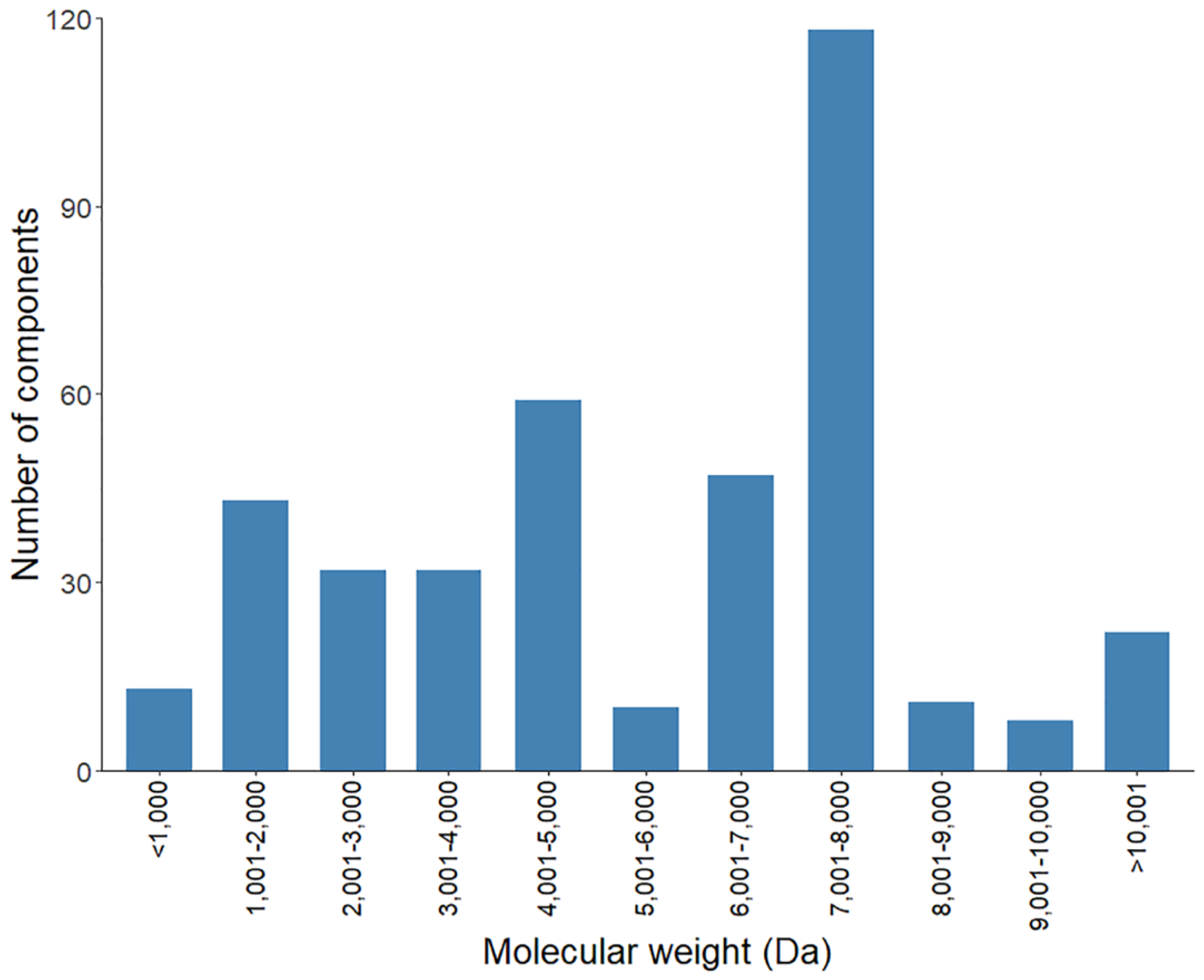
2.3.1. Mass Fingerprint of the Soluble Venom
2.3.2. Identification of Peptides by LC-MS/MS
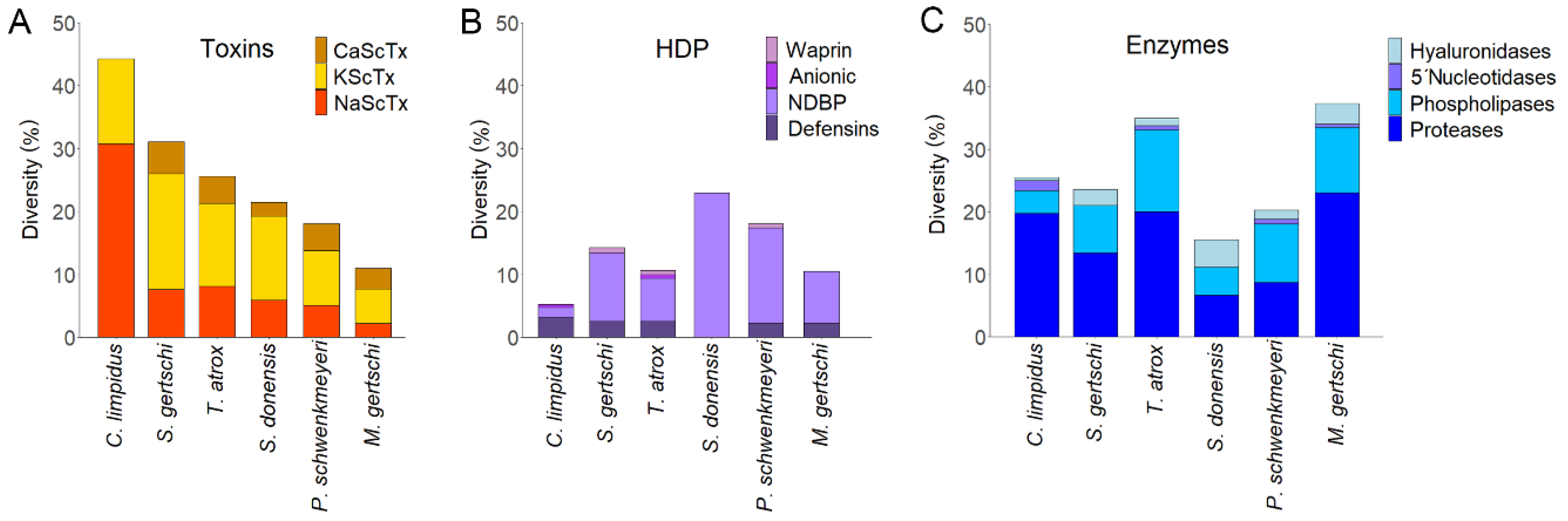
2.4. The Venom of the Highly Toxic C. limpidus versus the Venoms of Non-buthid Scorpions
3. Conclusions
4. Materials and Methods
4.1. Biological Material
4.2. RNA Isolation, Sequencing, and Assembly
4.3. Bioinformatics
4.4. Molecular Mass Fingerprint by LC-MS of the Venom
4.5. Identification of Venom Components by LC-MS/MS
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Possani, L.D.; Becerril, B.; Delepierre, M.; Tytgat, J. Scorpion toxins specific for Na+-channels. Eur. J. Biochem. 1999, 264, 287–300. [Google Scholar] [CrossRef]
- The Scorpion Files. Available online: https://www.ntnu.no/ub/scorpion-files/ (accessed on 29 April 2019).
- Chippaux, J.-P.; Goyffon, M. Epidemiology of scorpionism: A global appraisal. Acta Trop. 2008, 107, 71–79. [Google Scholar] [CrossRef] [PubMed]
- Santibáñez-López, C.E.; Francke, O.F.; Ureta, C.; Possani, L.D.; Lai, R. Scorpions from Mexico: From species diversity to venom complexity. Toxins 2016, 8, 2. [Google Scholar] [CrossRef]
- González-Santillán, E.; Possani, L.D. North American scorpion species of public health importance with a reappraisal of historical epidemiology. Acta Trop. 2018, 187, 264–274. [Google Scholar] [CrossRef]
- Rendón-Anaya, M.; Delaye, L.; Possani, L.D.; Herrera-Estrella, A. Global transcriptome analysis of the scorpion Centruroides noxius: New toxin families and evolutionary insights from an ancestral scorpion species. PLoS ONE 2012, 7, e43331. [Google Scholar] [CrossRef]
- Santibáñez-López, C.E.; Cid-Uribe, J.I.; Batista, C.V.F.; Ortiz, E.; Possani, L.D. Venom gland transcriptomic and proteomic analyses of the enigmatic scorpion Superstitionia donensis (Scorpiones: Superstitioniidae), with insights on the evolution of its venom components. Toxins 2016, 8, 367. [Google Scholar] [CrossRef]
- Ward, M.J.; Ellsworth, S.A.; Rokyta, D.R. Venom-gland transcriptomics and venom proteomics of the Hentz striped scorpion (Centruroides hentzi; Buthidae) reveal high toxin diversity in a harmless member of a lethal family. Toxicon 2018, 142, 14–29. [Google Scholar] [CrossRef] [PubMed]
- Quintero-Hernández, V.; Ramírez-Carreto, S.; Romero-Gutiérrez, M.T.; Valdez-Velázquez, L.L.; Becerril, B.; Possani, L.D.; Ortiz, E. Transcriptome analysis of scorpion species belonging to the Vaejovis Genus. PLoS ONE 2015, 10, e0117188. [Google Scholar] [CrossRef] [PubMed]
- Romero-Gutierrez, T.; Peguero-Sanchez, E.; Cevallos, M.A.; Batista, C.V.F.; Ortiz, E.; Possani, L.D. A deeper examination of Thorellius atrox Scorpion venom components with omic techonologies. Toxins 2017, 9, 399. [Google Scholar] [CrossRef]
- Zhong, J.; Zeng, X.-C.; Zeng, X.; Nie, Y.; Zhang, L.; Wu, S.; Bao, A. Transcriptomic analysis of the venom glands from the scorpion Hadogenes troglodytes revealed unique and extremely high diversity of the venom peptides. J. Proteom. 2017, 150, 40–62. [Google Scholar] [CrossRef]
- Santibáñez-López, C.E.; Cid-Uribe, J.I.; Zamudio, F.Z.; Batista, C.V.F.; Ortiz, E.; Possani, L.D. Venom gland transcriptomic and venom proteomic analyses of the scorpion Megacormus gertschi Díaz-Najera, 1966 (Scorpiones: Euscorpiidae: Megacorminae). Toxicon 2017, 133, 95–109. [Google Scholar] [CrossRef] [PubMed]
- Luna-Ramírez, K.; Quintero-Hernández, V.; Juárez-González, V.R.; Possani, L.D. Whole transcriptome of the venom gland from Urodacus yaschenkoi Scorpion. PLoS ONE 2015, 10, e0127883. [Google Scholar] [CrossRef]
- Ponce-Saavedra, J.; Francke, O.F.; Cano-Camacho, H.; Hernández-Calderón, E. Morphological and molecular evidence supporting specific status for Centruroides tecomanus (Scorpiones, Buthidae). Rev. Mex. Biodivers. 2009, 80, 71–84. [Google Scholar]
- Chavez-Haro, A.L.; Ortiz, E. Scorpionism and Dangerous Species of Mexico. In Scorpion Venoms; Springer: Dordrecht, The Netherlands, 2014; pp. 201–213. [Google Scholar]
- Riaño-Umbarila, L.; Rodríguez-Rodríguez, E.R.; Santibañez-López, C.E.; Güereca, L.; Uribe-Romero, S.J.; Gómez-Ramírez, I.V.; Cárcamo-Noriega, E.N.; Possani, L.D.; Becerril, B. Updating knowledge on new medically important scorpion species in Mexico. Toxicon 2017, 138, 130–137. [Google Scholar] [CrossRef]
- Available online: https://www.gob.mx/cms/uploads/attachment/file/425972/sem52.pdf (accessed on 29 April 2019).
- Riaño-Umbarila, L.; Olamendi-Portugal, T.; Morelos-Juárez, C.; Gurrola, G.B.; Possani, L.D.; Becerril, B. A novel human recombinant antibody fragment capable of neutralizing Mexican scorpion toxins. Toxicon 2013, 76, 370–376. [Google Scholar] [CrossRef]
- Dehesa-Dávila, M.; Ramfrez, A.N.; Zamudio, F.Z.; Gurrola-Briones, G.; Liévano, A.; Darszon, A.; Possani, L.D. Structural and functional comparison of toxins from the venom of the scorpions Centruroides infamatus infamatus, Centruroides limpidus limpidus and Centruroides noxius. Comp. Biochem. Physiol. Part B: Biochem. Mol. Boil. 1996, 113, 331–339. [Google Scholar] [CrossRef]
- Ramírz, A.N.; Martin, B.M.; Gurrola, G.B.; Possani, L.D. Isolation and characterization of a novel toxin from the venom of the scorpion Centruroides limpidus limpidus Karsch. Toxicon 1994, 32, 479–490. [Google Scholar] [CrossRef]
- Alagon, A.; Guzmán, H.; Martin, B.; Ramírez, A.; Carbone, E.; Possani, L. Isolation and characterization of two toxins from the mexican scorpion Centruroides limpidus limpidus Karsch. Comp. Biochem. Physiol. Part B: Comp. Biochem. 1988, 89, 153–161. [Google Scholar] [CrossRef]
- Lebreton, F.; Delepierre, M.; Ramírez, A.N.; Balderas, C.; Possani, L.D. Primary and NMR three-dimensional structure determination of a novel crustacean toxin from the venom of the scorpion Centruroides limpidus limpidus Karsch. Biochemistry 1994, 33, 11135–11149. [Google Scholar] [CrossRef]
- Olamendi-Portugal, T.; Restano-Cassulini, R.; Riaño-Umbarila, L.; Becerril, B.; Possani, L.D. Functional and immuno-reactive characterization of a previously undescribed peptide from the venom of the scorpion Centruroides limpidus. Peptides 2017, 87, 34–40. [Google Scholar] [CrossRef]
- Cid-Uribe, J.I.; Santibáñez-López, C.E.; Meneses, E.P.; Batista, C.V.; Jiménez-Vargas, J.M.; Ortiz, E.; Possani, L.D. The diversity of venom components of the scorpion species Paravaejovis schwenkmeyeri (Scorpiones: Vaejovidae) revealed by transcriptome and proteome analyses. Toxicon 2018, 151, 47–62. [Google Scholar] [CrossRef] [PubMed]
- Romero-Gutiérrez, M.T.; Santibáñez-López, C.E.; Jiménez-Vargas, J.M.; Batista, C.V.F.; Ortiz, E.; Possani, L.D. Transcriptomic and proteomic analyses reveal the diversity of venom components from the vaejovid scorpion Serradigitus gertschi. Toxins 2018, 10, 359. [Google Scholar] [CrossRef] [PubMed]
- Cid-Uribe, J.I.; Jiménez-Vargas, J.M.; Batista, C.V.F.; Zuñiga, F.Z.; Possani, L.D. Comparative proteomic analysis of female and male venoms from the Mexican scorpion Centruroides limpidus: Novel components found. Toxicon 2017, 125, 91–98. [Google Scholar] [CrossRef]
- Pfam. Available online: https://pfam.xfam.org/ (accessed on 29 April 2019).
- Venom Zone. Available online: https://venomzone.expasy.org/ (accessed on 29 April 2019).
- Gene Ontology. Available online: http://geneontology.org/ (accessed on 29 April 2019).
- Quintero-Hernandez, V.; Jimenez-Vargas, J.; Gurrola, G.; Valdivia, H.; Possani, L. Scorpion venom components that affect ion-channels function. Toxicon 2013, 76, 328–342. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gurevitz, M.; Froy, O.; Zilberberg, N.; Turkov, M.; Strugatsky, D.; Gershburg, E.; Lee, D.; Adams, M.E.; Tugarinov, V.; Anglister, J.; et al. Sodium channel modifiers from scorpion venom: Structure–activity relationship, mode of action and application. Toxicon 1998, 36, 1671–1682. [Google Scholar] [CrossRef]
- Escalona, M.P.; Possani, L.D. Scorpion beta-toxins and voltage-gated sodium channels: Interactions and effects. Front Biosci. 2013, 18, 572–587. [Google Scholar] [CrossRef]
- De Oliveira, U.C.; Candido, D.M.; Dorce, V.A.C.; Junqueira-De-Azevedo, I.D.L.M. The transcriptome recipe for the venom cocktail of Tityus bahiensis scorpion. Toxicon 2015, 95, 52–61. [Google Scholar] [CrossRef] [PubMed]
- Jablonsky, M.J.; Jackson, P.L.; Krishna, N.R. Solution structure of an insect-specific neurotoxin from the new world scorpion Centruroides sculpturatus Ewing†,‡. Biochemistry 2001, 40, 8273–8282. [Google Scholar] [CrossRef] [PubMed]
- Jablonsky, M.J.; Watt, D.D.; Krishna, N.R. Solution structure of an old world-like neurotoxin from the venom of the new world scorpion Centruroides sculpturatus Ewing. J. Mol. Boil. 1995, 248, 449–458. [Google Scholar] [CrossRef]
- Kuang, Q.; Purhonen, P.; Hebert, H. Structure of potassium channels. Cell. Mol. Life Sci. 2015, 72, 3677–3693. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiménez-Vargas, J.M.; Possani, L.D.; Luna-Ramírez, K. Arthropod toxins acting on neuronal potassium channels. Neuropharmacology 2017, 127, 139–160. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.M.; Nimigean, C.M. Voltage-gated potassium channels: A structural examination of selectivity and gating. Cold Spring Harbor Perspect. Boil. 2016, 8, a029231. [Google Scholar] [CrossRef]
- Tytgat, J.; Chandy, K.; Garcia, M.L.; A Gutman, G.; Martin-Eauclaire, M.-F.; Van Der Walt, J.J.; Possani, L.D. A unified nomenclature for short-chain peptides isolated from scorpion venoms: α-KTx molecular subfamilies. Trends Pharmacol. Sci. 1999, 20, 444–447. [Google Scholar] [CrossRef]
- Rodríguez De La Vega, R.C.; Possani, L.D. Current views on scorpion toxins specific for K+-channels. Toxicon 2004, 43, 865–875. [Google Scholar] [CrossRef]
- Cremonez, C.M.; Maiti, M.; Peigneur, S.; Cassoli, J.S.; Dutra, A.A.A.; Waelkens, E.; Lescrinier, E.; Herdewijn, P.; de Lima, M.E.; Pimenta, A.M.C.; et al. Structural and functional elucidation of peptide TS11 shows evidence of a novel subfamily of scorpion venom toxins. Toxins 2016, 8, 288. [Google Scholar] [CrossRef] [PubMed]
- Correnti, C.E.; Gewe, M.M.; Mehlin, C.; Bandaranayake, A.D.; Johnsen, W.A.; Rupert, P.B.; Brusniak, M.-Y.; Clarke, M.; Burke, S.E.; De Van Der Schueren, W.; et al. Screening, large-scale production, and structure-based classification for cystine-dense peptides. Nat. Struct. Mol. Boil. 2018, 25, 270–278. [Google Scholar] [CrossRef] [PubMed]
- Nieto, A.R.; Gurrola, G.B.; Vaca, L.; Possani, L.D. Noxiustoxin 2, a novel K+ channel blocking peptide from the venom of the scorpion Centruroides noxius Hoffmann. Toxicon 1996, 34, 913–922. [Google Scholar] [CrossRef]
- Valdivia, H.H.; Smith, J.S.; Martin, B.M.; Coronado, R.; Possani, L.D. Charybdotoxin and noxiustoxin, two homologous peptide inhibitors of the K+(Ca2+) channel. FEBS Lett. 1988, 226, 280–284. [Google Scholar] [CrossRef]
- Kuzmenkov, A.I.; Krylov, N.A.; Chugunov, A.O.; Grishin, E.V.; Vassilevski, A.A. Kalium: A database of potassium channel toxins from scorpion venom. Database 2016, 2016. [Google Scholar] [CrossRef]
- Restano-Cassulini, R.; Olamendi-Portugal, T.; Zamudio, F.; Becerril, B.; Possani, L.D. Two novel ergtoxins, blockers of K+-channels, purified from the Mexican scorpion Centruroides elegans elegans. Neurochem Res. 2008, 33, 1525–1533. [Google Scholar] [CrossRef]
- Hill, A.P.; Sunde, M.; Campbell, T.J.; Vandenberg, J.I. Mechanism of block of the hERG K+ channel by the scorpion toxin CnErg1. Biophys. J. 2007, 92, 3915–3929. [Google Scholar] [CrossRef] [PubMed]
- Torres, A.M.; Bansal, P.; Alewood, P.F.; A Bursill, J.; Kuchel, P.W.; I Vandenberg, J.; Vandenberg, J. Solution structure of CnErg1 (Ergtoxin), a HERG specific scorpion toxin. FEBS Lett. 2003, 539, 138–142. [Google Scholar] [CrossRef] [Green Version]
- Jimenez-Vargas, J.; Restano-Cassulini, R.; Quintero-Hernandez, V.; Gurrola, G.; Possani, L. Recombinant expression of the toxic peptide ErgTx1 and role of Met35 on its stability and function. Peptides 2011, 32, 560–567. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.-Y.; Hu, Y.-T.; Yang, W.-S.; He, Y.-W.; Feng, J.; Wang, B.; Zhao, R.-M.; Ding, J.-P.; Cao, Z.-J.; Li, W.-X.; et al. Hg1, Novel peptide inhibitor specific for Kv1.3 channels from first scorpion kunitz-type potassium channel toxin family*. J. Boil. Chem. 2012, 287, 13813–13821. [Google Scholar] [CrossRef]
- Santibáñez-López, C.E.; Kriebel, R.; Ballesteros, J.A.; Rush, N.; Witter, Z.; Williams, J.; Janies, D.A.; Sharma, P.P.; Crandall, K. Integration of phylogenomics and molecular modeling reveals lineage-specific diversification of toxins in scorpions. PeerJ 2018, 6, e5902. [Google Scholar] [CrossRef]
- Chen, Z.; Hu, Y.; Han, S.; Yin, S.; He, Y.; Wu, Y.; Cao, Z.; Li, W. ImKTx1, a new Kv1.3 channel blocker with a unique primary structure. J. Biochem. Mol. Toxicol. 2011, 25, 244–251. [Google Scholar] [CrossRef]
- Gao, B.; Harvey, P.J.; Craik, D.J.; Ronjat, M.; De Waard, M.; Zhu, S. Functional evolution of scorpion venom peptides with an inhibitor cystine knot fold. Biosci. Rep. 2013, 33, 513–527. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, X.; Wang, G. Insights into Antimicrobial Peptides from Spiders and Scorpions. Protein Pept. Lett. 2016, 23, 707–721. [Google Scholar] [CrossRef]
- Yan, L.; Adams, M.E. Lycotoxins, antimicrobial peptides from venom of the wolf spider Lycosa carolinensis. J. Boil. Chem. 1998, 273, 2059–2066. [Google Scholar] [CrossRef]
- Torres-Larios, A.; Gurrola, G.B.; Zamudio, F.Z.; Possani, L.D.; Torres-Larios, A. Hadrurin, a new antimicrobial peptide from the venom of the scorpion Hadrurus aztecus. JBIC J. Boil. Inorg. Chem. 2000, 267, 5023–5031. [Google Scholar] [CrossRef]
- Lorenzini, D.M.; Da Silva, P.I.; Fogaça, A.C.; Bulet, P.; Daffre, S. Acanthoscurrin: A novel glycine-rich antimicrobial peptide constitutively expressed in the hemocytes of the spider Acanthoscurria gomesiana. Dev. Comp. Immunol. 2003, 27, 781–791. [Google Scholar] [CrossRef]
- Luna-Ramirez, K.; Skaljac, M.; Grotmann, J.; Kirfel, P.; Vilcinskas, A.; Possani, L.D. Orally delivered scorpion antimicrobial peptides exhibit activity against pea aphid (Acyrthosiphon pisum) and Its Bacterial Symbionts. Toxins 2017, 9, 261. [Google Scholar] [CrossRef]
- Ferreira, L.; Alves, W.; Lucas, M.; Habermehl, G. Isolation and characterization of a bradykinin potentiating peptide (BPP-S) isolated from Scaptocosa raptoria venom. Toxicon 1996, 34, 599–603. [Google Scholar] [CrossRef]
- Liu, Z.; Deng, M.; Xiang, J.; Ma, H.; Hu, W.; Zhao, Y.; Li, D.-C.; Liang, S. A novel spider peptide toxin suppresses tumor growth through dual signaling pathways. Mol. Med. 2012, 12, 1350–1360. [Google Scholar] [CrossRef]
- Guo, X.; Ma, C.; Du, Q.; Wei, R.; Wang, L.; Zhou, M.; Chen, T.; Shaw, C. Two peptides, TsAP-1 and TsAP-2, from the venom of the Brazilian yellow scorpion, Tityus serrulatus: Evaluation of their antimicrobial and anticancer activities. Biochimie 2013, 95, 1784–1794. [Google Scholar] [CrossRef]
- Zeng, X.-C.; Zhou, L.; Shi, W.; Luo, X.; Zhang, L.; Nie, Y.; Wang, J.; Wu, S.; Cao, B.; Cao, H. Three new antimicrobial peptides from the scorpion Pandinus imperator. Peptides 2013, 45, 28–34. [Google Scholar] [CrossRef]
- De La Vega, R.R.; García, B.I.; D’Ambrosio, C.; Diego-García, E.; Scaloni, A.; Possani, L.D. Antimicrobial peptide induction in the haemolymph of the Mexican scorpion Centruroides limpidus limpidus in response to septic injury. Cell. Mol. Life Sci. 2004, 61, 1507–1519. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Shi, W.; Zeng, X.-C.; Ge, F.; Yang, M.; Nie, Y.; Bao, A.; Wu, S.; E, G. Unique diversity of the venom peptides from the scorpion Androctonus bicolor revealed by transcriptomic and proteomic analysis. J. Proteom. 2015, 128, 231–250. [Google Scholar] [CrossRef] [PubMed]
- Zeng, X.-C.; Wang, S.; Nie, Y.; Zhang, L.; Luo, X. Characterization of BmKbpp, a multifunctional peptide from the Chinese scorpion Mesobuthus martensii Karsch: Gaining insight into a new mechanism for the functional diversification of scorpion venom peptides. Peptides 2012, 33, 44–51. [Google Scholar] [CrossRef]
- Marques-Neto, L.M.; Trentini, M.M.; Das Neves, R.C.; Resende, D.P.; Procopio, V.O.; Da Costa, A.C.; Kipnis, A.; Mortari, M.R.; Schwartz, E.F.; Junqueira-Kipnis, A.P. Antimicrobial and chemotactic activity of scorpion-derived peptide, ToAP2, against Mycobacterium massiliensis. Toxins 2018, 10, 219. [Google Scholar] [CrossRef]
- Melo, M.M.; Daniele-Silva, A.; Teixeira, D.G.; Estrela, A.B.; Melo, K.R.; Oliveira, V.S.; Rocha, H.A.; Ferreira, L.D.S.; Pontes, D.L.; Lima, J.P.; et al. Structure and in vitro activities of a copper II-chelating anionic peptide from the venom of the scorpion Tityus stigmurus. Peptides 2017, 94, 91–98. [Google Scholar] [CrossRef]
- Ramos, O.; Selistre-De-Araújo, H. Snake venom metalloproteases—structure and function of catalytic and disintegrin domains. Comp. Biochem. Physiol. Part C: Toxicol. Pharmacol. 2006, 142, 328–346. [Google Scholar] [CrossRef]
- Lu, J.; Yang, H.; Yu, H.; Gao, W.; Lai, R.; Liu, J.; Liang, X. A novel serine protease inhibitor from Bungarus fasciatus venom. Peptides 2008, 29, 369–374. [Google Scholar] [CrossRef]
- Touchard, A.; Aili, S.R.; Fox, E.G.P.; Escoubas, P.; Orivel, J.; Nicholson, G.M.; Dejean, A.; King, G.F. The biochemical toxin arsenal from ant venoms. Toxins 2016, 8, 30. [Google Scholar] [CrossRef] [PubMed]
- Toom, P.M.; Chan, D.S. Enzymatic activities of venom from the jellyfish Stomolophus meleagris. Comp. Biochem. Physiol. Part B: Comp. Biochem. 1972, 43, 435–441. [Google Scholar] [CrossRef]
- De Graaf, D.C.; Aerts, M.; Brunain, M.; Desjardins, C.A.; Jacobs, F.J.; Werren, J.H.; Devreese, B. Insights into the venom composition of the ectoparasitoid wasp Nasonia vitripennis from bioinformatic and proteomic studies. Insect Mol. Boil. 2010, 19, 11–26. [Google Scholar]
- Trevisan-Silva, D.; Bednaski, A.V.; Gremski, L.H.; Chaim, O.M.; Veiga, S.S.; Senff-Ribeiro, A. Differential metalloprotease content and activity of three Loxosceles spider venoms revealed using two-dimensional electrophoresis approaches. Toxicon 2013, 76, 11–22. [Google Scholar] [CrossRef]
- Ortiz, E.; Rendón-Anaya, M.; Rego, S.C.; Schwartz, E.F.; Possani, L.D. Antarease-like Zn-metalloproteases are ubiquitous in the venom of different scorpion genera. Biochim. Biophys. Acta (BBA)—General Subj. 2014, 1840, 1738–1746. [Google Scholar] [CrossRef]
- Almeida, F.; Pimenta, A.; De Figueiredo, S.; Santoro, M.; Martin-Eauclaire, M.; Diniz, C.; De Lima, M.; Pimenta, A.M.D.C. Enzymes with gelatinolytic activity can be found in Tityus bahiensis and Tityus serrulatus venoms. Toxicon 2002, 40, 1041–1045. [Google Scholar] [CrossRef]
- Conde, R.; Zamudio, F.Z.; Becerril, B.; Possani, L.D. Phospholipin, a novel heterodimeric phospholipase A2 from Pandinus imperator scorpion venom. FEBS Lett. 1999, 460, 447–450. [Google Scholar] [CrossRef]
- Morey, S.S.; Kiran, K.; Gadag, J. Purification and properties of hyaluronidase from Palamneus gravimanus (Indian black scorpion) venom. Toxicon 2006, 47, 188–195. [Google Scholar] [CrossRef] [PubMed]
- Fletcher, P.L.; Fletcher, M.D.; Weninger, K.; Anderson, T.E.; Martin, B.M. Vesicle-associated membrane protein (VAMP) cleavage by a new metalloprotease from the Brazilian scorpion Tityus serrulatus. J. Biol. Chem. 2010, 285, 7405–7416. [Google Scholar] [CrossRef] [PubMed]
- Otero, R.; Navío, E.; Céspedes, F.; Núñez, M.; Lozano, L.; Moscoso, E.; Matallana, C.; Arsuza, N.; García, J.; Fernandez, D.; et al. Scorpion envenoming in two regions of Colombia: Clinical, epidemiological and therapeutic aspects. Trans. Soc. Trop. Med. Hyg. 2004, 98, 742–750. [Google Scholar] [CrossRef] [PubMed]
- Fletcher, P.L., Jr.; Fletcher, M.D.; Possani, L.D. Characteristics of pancreatic exocrine secretion produced by venom from the brazilian scorpion, Tityus serrulatus. Eur. J. Cell Biol. 1992, 58, 259–270. [Google Scholar] [PubMed]
- D’Suze, G.; Sevcik, C.; Ramos, M. Presence of curarizing polypeptides and a pancreatitis-inducing fraction without muscarinic effects in the venom of the Venezuelan scorpion Tityus discrepans (Karsch). Toxicon 1995, 33, 295. [Google Scholar] [CrossRef]
- Borges, A.; Trejo, E.; Vargas, A.M.; Céspedes, G.; Hernández, A.; Alfonzo, M.J. Pancreatic toxicity in mice elicited by Tityus zulianus and Tityus discrepans scorpion venoms. Investig. Clin. 2004, 45, 269–276. [Google Scholar]
- Bartholomew, C. Acute Scorpion Pancreatitis in Trinidad. BMJ 1970, 1, 666–668. [Google Scholar] [CrossRef] [Green Version]
- Possani, L.D.; Martin, B.M.; Fletcher, M.D.; Fletcher, P.L. Discharge effect on pancreatic exocrine secretion produced by toxins purified from Tityus serrulatus scorpion venom. J. Boil. Chem. 1991, 266, 3178–3185. [Google Scholar]
- Fletcher, M.D.; Possani, L.D.; Fletcher, P.L., Jr. Morphological studies by light and electron microscopy of pancreatic acinar cells under the effect of Tityus serrulatus venom. Cell Tissue Res. 1994, 278, 255–264. [Google Scholar] [CrossRef]
- Fletcher, P.L.; Fletcher, M.D.; Fainter, L.K.; Terrian, D.M. Action of new world scorpion venom and its neurotoxins in secretion. Toxicon 1996, 34, 1399–1411. [Google Scholar] [CrossRef]
- Jiménez-Ferrer, E.; Reynosa-Zapata, I.; Pérez-Torres, Y.; Tortoriello, J. The secretagogue effect of the poison from Centruroides limpidus limpidus on the pancreas of mice and the antagonistic action of the Bouvardia ternifolia extract. Phytomedicine 2005, 12, 65–71. [Google Scholar] [CrossRef]
- Habermann, E.; Hardt, K. A sensitive and specific plate test for the quantitation of phospholipases. Anal. Biochem. 1972, 50, 163–173. [Google Scholar] [CrossRef]
- Dhananjaya, B.L.; D’Souza, C.J.M. The pharmacological role of nucleotidases in snake venoms. Cell Biochem. 2010, 28, 171–177. [Google Scholar] [CrossRef]
- Khan, N.; Niazi, Z.R.; Rehman, F.U.; Akhtar, A.; Khan, M.M.; Khan, S.; Baloch, N.; Khan, S. Hyaluronidases: A Therapeutic Enzyme. Protein Pept. Lett. 2018, 25, 663–676. [Google Scholar] [CrossRef] [PubMed]
- Bordon, K.C.F.; Wiezel, G.A.; Amorim, F.G.; Arantes, E.C. Arthropod venom Hyaluronidases: Biochemical properties and potential applications in medicine and biotechnology. J. Venom. Anim. Toxins Incl. Trop. Dis. 2015, 21, 165. [Google Scholar] [CrossRef]
- Almonte, A.G.; Sweatt, J.D. Serine proteases, serine protease inhibitors, and protease-activated receptors: Roles in synaptic function and behavior. Brain Res. 2011, 1407, 107–122. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Wang, B.; Hu, J.; Yang, W.; Cao, Z.; Zhuo, R.; Li, W.; Wu, Y. SjAPI, the first functionally characterized ascaris-type protease inhibitor from animal venoms. PLoS ONE 2013, 8, e57529. [Google Scholar] [CrossRef] [PubMed]
- Zhao, R.; Dai, H.; Qiu, S.; Li, T.; He, Y.; Ma, Y.; Chen, Z.; Wu, Y.; Li, W.; Cao, Z. SdPI, the first functionally characterized kunitz-type trypsin inhibitor from scorpion venom. PLoS ONE 2011, 6, e27548. [Google Scholar] [CrossRef] [PubMed]
- Law, R.H.P.; Zhang, Q.; McGowan, S.; Buckle, A.M.; A Silverman, G.; Wong, W.; Rosado, C.J.; Langendorf, C.G.; Pike, R.N.; I Bird, P.; et al. An overview of the serpin superfamily. Genome Boil. 2006, 7, 216. [Google Scholar]
- Gronenborn, A.M.; Nilges, M.; Peanasky, R.J.; Clore, G.M. Sequential resonance assignment and secondary structure determination of the ascaris trypsin inhibitor, a member of a novel class of proteinase inhibitors. Biochemistry 1990, 29, 183–189. [Google Scholar] [CrossRef]
- Meekins, D.A.; Kanost, M.R.; Michel, K. Serpins in arthropod biology. Semin. Cell Dev. Boil. 2017, 62, 105–119. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gibbs, G.M.; Roelants, K.; O’Bryan, M.K. The CAP Superfamily: Cysteine-rich secretory proteins, antigen 5, and pathogenesis-related 1 proteins—Roles in reproduction, cancer, and immune defense. Endocr. Rev. 2008, 29, 865–897. [Google Scholar] [CrossRef]
- Abraham, A.; Chandler, D.E. Tracing the evolutionary history of the CAP superfamily of proteins using amino acid sequence homology and conservation of splice sites. J. Mol. Evol. 2017, 85, 137–157. [Google Scholar] [CrossRef]
- Fry, B.G.; Roelants, K.; Champagne, D.E.; Scheib, H.; Tyndall, J.D.; King, G.F.; Nevalainen, T.J.; Norman, J.A.; Lewis, R.J.; Norton, R.S.; et al. The toxicogenomic multiverse: Convergent recruitment of proteins into animal venoms. Annu. Genom. Hum. Genet. 2009, 10, 483–511. [Google Scholar] [CrossRef]
- Wang, J.; Duan, Y.; Guo, M.; Huang, Q.; Liu, Q.; Niu, L.; Teng, M.; Hao, Q.; Shen, B.; Cheng, X.P.; et al. Blocking effect and crystal structure of natrin toxin, a cysteine-rich secretory protein from Naja atra Venom that Targets the BK Ca Channel†,‡. Biochemistry 2005, 44, 10145–10152. [Google Scholar] [CrossRef] [PubMed]
- Gibbs, G.M.; Orta, G.; Reddy, T.; Koppers, A.J.; Martínez-López, P.; De La Vega-Beltràn, J.L.; Lo, J.C.Y.; Veldhuis, N.; Jamsai, D.; McIntyre, P.; et al. Cysteine-rich secretory protein 4 is an inhibitor of transient receptor potential M8 with a role in establishing sperm function. Proc. Natl. Acad. Sci. USA 2011, 108, 7034–7039. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ruiming, Z.; Yibao, M.; Yawen, H.; Zhiyong, D.; Yingliang, W.; Zhijian, C.; Wenxin, L. Comparative venom gland transcriptome analysis of the scorpion Lychas mucronatus reveals intraspecific toxic gene diversity and new venomous components. BMC Genom. 2010, 11, 452. [Google Scholar] [CrossRef]
- Rokyta, D.R.; Ward, M.J. Venom-gland transcriptomics and venom proteomics of the black-back scorpion (Hadrurus spadix) reveal detectability challenges and an unexplored realm of animal toxin diversity. Toxicon 2017, 128, 23–37. [Google Scholar] [CrossRef] [PubMed]
- Miyashita, M.; Otsuki, J.; Hanai, Y.; Nakagawa, Y.; Miyagawa, H. Characterization of peptide components in the venom of the scorpion Liocheles australasiae (Hemiscorpiidae). Toxicon 2007, 50, 428–437. [Google Scholar] [CrossRef]
- Possani, L.D.; Becerril, B.; Riaño-Umbarila, L.; Juárez-González, V.R.; Olamendi-Portugal, T.; Ortíz-León, M.; Riaño-Umbarila, L.; Juárez-González, V.R.; Olamendi-Portugal, T.; Ortiz-León, M. A strategy for the generation of specific human antibodies by directed evolution and phage display. FEBS J. 2005, 272, 2591–2601. [Google Scholar] [Green Version]
- Riaño-Umbarila, L.; Rudiño-Piñera, E.; Becerril, B.; Torres-Larios, A.; Canul-Tec, J.C.; Possani, L.D. Structural basis of neutralization of the major toxic component from the scorpion Centruroides noxius Hoffmann by a human-derived single-chain antibody fragment. J. Boil. Chem. 2011, 286, 20892–20900. [Google Scholar]
- Zamudio, F.; Saavedra, R.; Martin, B.M.; Gurrola-Briones, G.; Herion, P.; Possani, L.D. Amino acid sequence and immunological characterization with monoclonal antibodies of two toxins from the venom of the scorpion Centruroides noxius Hoffmann. JBIC J. Boil. Inorg. Chem. 1992, 204, 281–292. [Google Scholar] [CrossRef]
- Fastqc. Available online: https://www.bioinformatics.babraham.ac.uk/projects/fastqc/ (accessed on 29 April 2019).
- Grabherr, M.G.; Haas, B.J.; Yassour, M.; Levin, J.Z.; Thompson, D.A.; Amit, I.; Adiconis, X.; Fan, L.; Raychowdhury, R.; Zeng, Q.; et al. Full-length transcriptome assembly from RNA-Seq data without a reference genome. Nat. Biotechnol. 2011, 29, 644–652. [Google Scholar] [CrossRef] [PubMed]
- ProP. Available online: http://www.cbs.dtu.dk/services/ProP/ (accessed on 29 April 2019).
- SpiderP. Available online: http://www.arachnoserver.org/spiderP.html (accessed on 29 April 2019).
- Mafft. Available online: https://www.ebi.ac.uk/Tools/msa/mafft/ (accessed on 29 April 2019).
- Hall, T.A. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Rstudio. Available online: https://www.rstudio.com/ (accessed on 29 April 2019).
- Wego. Available online: http://wego.genomics.org.cn/ (accessed on 29 April 2019).




© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cid-Uribe, J.I.; Meneses, E.P.; Batista, C.V.F.; Ortiz, E.; Possani, L.D. Dissecting Toxicity: The Venom Gland Transcriptome and the Venom Proteome of the Highly Venomous Scorpion Centruroides limpidus (Karsch, 1879). Toxins 2019, 11, 247. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins11050247
Cid-Uribe JI, Meneses EP, Batista CVF, Ortiz E, Possani LD. Dissecting Toxicity: The Venom Gland Transcriptome and the Venom Proteome of the Highly Venomous Scorpion Centruroides limpidus (Karsch, 1879). Toxins. 2019; 11(5):247. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins11050247
Chicago/Turabian StyleCid-Uribe, Jimena I., Erika P. Meneses, Cesar V. F. Batista, Ernesto Ortiz, and Lourival D. Possani. 2019. "Dissecting Toxicity: The Venom Gland Transcriptome and the Venom Proteome of the Highly Venomous Scorpion Centruroides limpidus (Karsch, 1879)" Toxins 11, no. 5: 247. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins11050247