Toxin–Antitoxin Systems in Bacillus subtilis

Friedrich-Schiller-Universität Jena, Matthias-Schleiden-Institut, AG Bakteriengenetik, Philosophenweg 12, D-07743 Jena, Germany
*
Author to whom correspondence should be addressed.
Toxins 2019, 11(5), 262; https://0-doi-org.brum.beds.ac.uk/10.3390/toxins11050262
Submission received: 10 April 2019
/
Revised: 30 April 2019
/
Accepted: 7 May 2019
/
Published: 9 May 2019
(This article belongs to the Special Issue Toxin-antitoxin (TA) systems)
Abstract
:Toxin–antitoxin (TA) systems were originally discovered as plasmid maintenance systems in a multitude of free-living bacteria, but were afterwards found to also be widespread in bacterial chromosomes. TA loci comprise two genes, one coding for a stable toxin whose overexpression kills the cell or causes growth stasis, and the other coding for an unstable antitoxin that counteracts toxin action. Of the currently known six types of TA systems, in Bacillus subtilis, so far only type I and type II TA systems were found, all encoded on the chromosome. Here, we review our present knowledge of these systems, the mechanisms of antitoxin and toxin action, and the regulation of their expression, and we discuss their evolution and possible physiological role.
Keywords:
toxin–antitoxin system; antisense RNA; small regulatory RNA (sRNA)/target RNA interaction; toxic peptide; RNA degradation; prophageKey Contribution: Toxin-antitoxin (TA) systems are ubiquitous in bacterial chromosomes and plasmids. This review provides a comprehensive overview of all currently characterized TA systems of Bacillus subtilis.
1. Introduction
Toxin–antitoxin (TA) systems are genetic modules that are widespread in bacterial genomes and plasmids. They consist of a gene pair encoding a stable toxin that impedes cell growth by interfering with fundamental processes, and an unstable antitoxin that counteracts the toxin activity. Currently, six types of TA systems are known. Whereas type I and III TA systems employ RNA antitoxins, the other TA systems use proteinaceous antitoxins. In type I TA systems, antitoxins interact with the corresponding toxin messenger RNAs (mRNAs) [1]. In type III systems, the RNA antitoxin binds the toxin protein directly. By contrast, type II antitoxins inhibit their toxins via a protein/protein interaction. Antitoxins in type IV TA systems interfere with binding of the toxin proteins to their cellular targets [2]. In type V systems, the antitoxin is an RNase which specifically cleaves the toxin mRNA, thus preventing toxin expression [3]. In the recently discovered type VI systems, the toxin binds directly to the β sliding clamp DnaN to block replication elongation, whereas the antitoxin promotes degradation by the ClpXP protease [4]. The toxin targets are diverse and comprise DNA replication, stability of mRNA or DNA, protein synthesis, cell-wall biosynthesis, membrane integrity, and ATP synthesis.
Although TA systems are ubiquitous in bacterial genomes and their molecular activities were investigated for many years, the biological function of the majority of them is still elusive. To date, the following major biological functions were demonstrated: Post-segregational killing (PSK), abortive infection, persister formation/antibiotic tolerance, and stress response (reviewed in Reference [5]).
In Bacillus subtilis, both type I and type II TA systems were discovered and characterized. In this review, we provide an overview of the known B. subtilis TA systems with special focus on their molecular mechanisms, the control of toxin and antitoxin expression, the modes of toxin action, and their biological role.
2. Type I Toxin–Antitoxin Systems
2.1. Overview of B. subtilis Type I TA Systems
In 2005, the first type I TA system was discovered in B. subtilis and designated txpA/RatA [6]. Subsequent transcriptomics [7] and bioinformatics studies [8] revealed 14 potential type I TA systems [9], five of which are located on prophage regions. Four of them—txpA/RatA [6,10], bsrG/SR4 [11], bsrE/SR5 [12,13] and yonT-yoyJ/SR6 [14]—were investigated in more detail (Table 1) and are reviewed here. The other nine potential systems, the putative toxins of which were grouped into the YhzE family [9], are yet to be confirmed as TA systems.
2.2. Mechanisms of Antitoxin Action
Two principal mechanisms of action of type I antitoxins are known: inhibition of toxin translation or promotion of degradation of the toxin mRNA. B. subtilis SR4 is the first bifunctional antitoxin; it both impedes translation of bsrG mRNA and facilitates its degradation [15] (Figure 1).
In cases of Escherichia coli type I TA systems, where the complementarity between antitoxin and toxin mRNA resides in their 5′ regions, the antitoxin either binds at a region overlapping the toxin Shine Dalgarno (SD) sequence to inhibit toxin translation directly (e.g., E. coli SymR), or it prevents translation of a leader peptide translationally coupled to the toxin (e.g., hok/Sok or ldrD/RdlD [16]). Alternatively, the antitoxin binds to a ribosome standby-site to block toxin translation (e.g., E. coli tisB/IstR1 [17]).
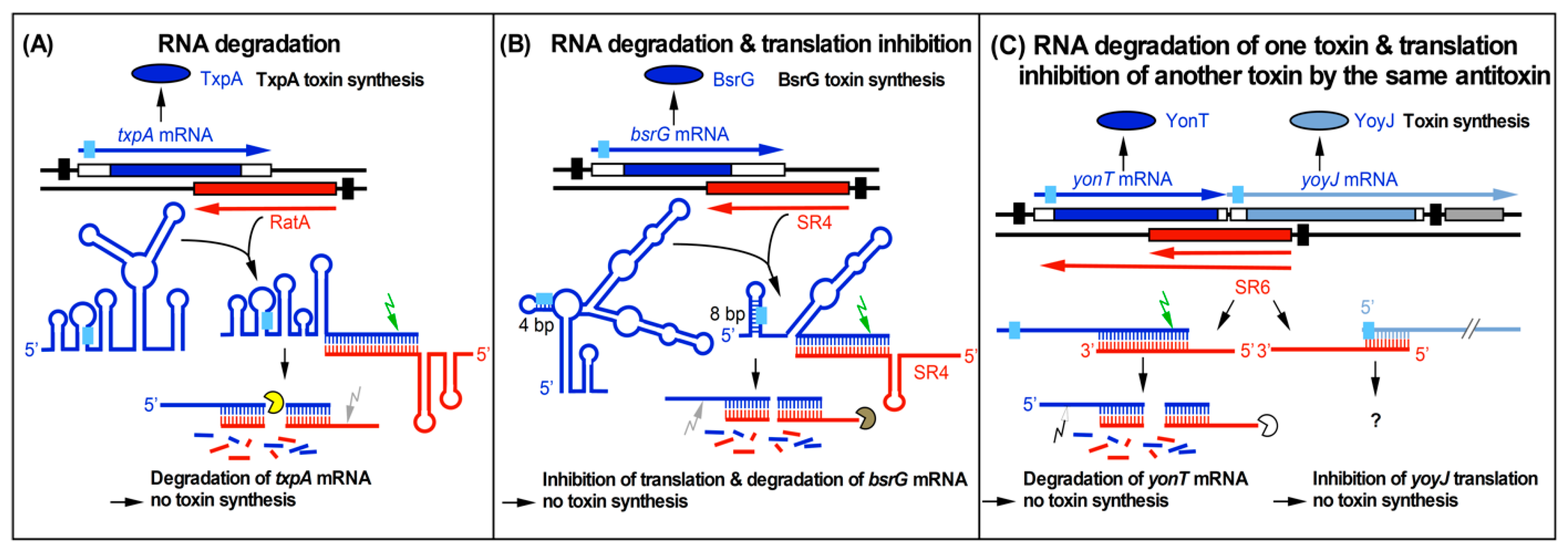
Figure 1.
Three currently known mechanisms of action used by Bacillus subtilis type I antitoxins. Black bars denote promoters. Toxins are drawn in blue or blue-gray and antitoxins in red. The toxin open reading frames (ORFs) are represented by blue and blue-gray bars. Light-blue boxes denote ribosome binding sites (RBS). Arrows symbolize endoribonucleases (RNase III, green; RNase Y, gray; white, unknown RNase) and circular segments symbolize 3′–5′ exoribonucleases (PNPase, yellow; RNase R, brown; unknown RNase, white). (A) Promotion of RNA degradation. The antitoxin RatA and its txpA toxin messenger RNA (mRNA) base-pair at their 3′ ends. (B) RNA degradation and translation inhibition. The antitoxin SR4 and the corresponding bsrG toxin mRNA interact at their 3′ ends. SR4 binding to bsrG mRNA induces a conformational alteration that extends the region sequestering the Shine Dalgarno (SD) sequence from 4 bp to 8 bp which inhibits bsrG translation. Additionally, the SR4/bsrG mRNA interaction facilitates toxin mRNA decay by an initial Rnase III cleavage followed by subsequent RNase R and RNase Y degradation. (C) One antitoxin inhibits two toxins via different mechanisms. Antitoxin SR6 and yonT toxin mRNA base-pair at their 3′ ends, which promotes yonT mRNA decay via an initial RNase III cleavage that is followed by degradation by so far unidentified RNases. Furthermore, SR6 interacts with yoyJ toxin mRNA by base-pairing at the 5′ ends, which does not promote yoyJ mRNA degradation, but prevents yoyJ overexpression, most likely via translational inhibition. (A,B) are based on Reference [18].
Figure 1.
Three currently known mechanisms of action used by Bacillus subtilis type I antitoxins. Black bars denote promoters. Toxins are drawn in blue or blue-gray and antitoxins in red. The toxin open reading frames (ORFs) are represented by blue and blue-gray bars. Light-blue boxes denote ribosome binding sites (RBS). Arrows symbolize endoribonucleases (RNase III, green; RNase Y, gray; white, unknown RNase) and circular segments symbolize 3′–5′ exoribonucleases (PNPase, yellow; RNase R, brown; unknown RNase, white). (A) Promotion of RNA degradation. The antitoxin RatA and its txpA toxin messenger RNA (mRNA) base-pair at their 3′ ends. (B) RNA degradation and translation inhibition. The antitoxin SR4 and the corresponding bsrG toxin mRNA interact at their 3′ ends. SR4 binding to bsrG mRNA induces a conformational alteration that extends the region sequestering the Shine Dalgarno (SD) sequence from 4 bp to 8 bp which inhibits bsrG translation. Additionally, the SR4/bsrG mRNA interaction facilitates toxin mRNA decay by an initial Rnase III cleavage followed by subsequent RNase R and RNase Y degradation. (C) One antitoxin inhibits two toxins via different mechanisms. Antitoxin SR6 and yonT toxin mRNA base-pair at their 3′ ends, which promotes yonT mRNA decay via an initial RNase III cleavage that is followed by degradation by so far unidentified RNases. Furthermore, SR6 interacts with yoyJ toxin mRNA by base-pairing at the 5′ ends, which does not promote yoyJ mRNA degradation, but prevents yoyJ overexpression, most likely via translational inhibition. (A,B) are based on Reference [18].

By contrast, in the B. subtilis type I TA systems studied so far (txpA/RatA [6], bsrG/SR4 [11], bsrE/SR5 [12], bsrH/as-bsrH [10], and yonT/SR6 [14]), the RNA antitoxins bind at the 3′ end of the toxin mRNAs and support their degradation. This was demonstrated in four instances by the longer half-lives of the toxin mRNAs in the absence of their corresponding antitoxins [10,11,12,14]. In these cases, RNA antitoxins and their toxin mRNAs share a rather long complementary stretch of 100 to 140 bp. For bsrG/SR4, it was found that SR4 not only promotes degradation of bsrG mRNA, but induces a conformational change around the bsrG RBS that further impairs ribosome binding, thus additionally impeding toxin translation [15]. Neither upon RatA binding to txpA mRNA [10] nor antitoxin SR5 binding to bsrE mRNA [13] was such a conformational change observed, suggesting that RatA and SR5 only cause toxin degradation.
For the yonT/SR6 TA system, no secondary structures were probed to investigate this issue. Interestingly, SR6 controls a second toxin gene, yoyJ, using its 5′ region to base-pair to the yoyJ ribosome binding site (RBS) [14]. Whereas SR6 promotes yonT mRNA degradation, it neither affects the amount nor half-life of yoyJ mRNA [14]. Therefore, it seems to inhibit yoyJ translation.
2.3. Binding Pathway of RNA Antitoxin and Toxin mRNA
Binding pathways were studied in detail for many cis-encoded sense/antisense systems, and binding kinetics of sense and antisense RNAs were measured (reviewed in Reference [19]). On average, kapp values (apparent pairing rate constants) are in the range of 1 × 105 to 1 × 106 M−1·s−1. High kapp values are indicative for efficient pairing between two complementary RNAs. For bsrG/SR4, a kapp value of 6.5 × 105 M−1·s−1 was determined [15] and, for bsrE/SR5, a kapp value of 4 × 106 M−1·s−1 [13]. For txpA/RatA and for yonT/SR6, kapp measurements are yet to be performed. To narrow down the minimal regions required for efficient complex formation with their toxin mRNAs, truncated SR4 or SR5 species were employed in interaction studies with their toxin mRNAs [13,15]. Both wild-type RNA antitoxins are composed of four stem-loops (SLs) (see Figure 2). An SR4 species composed of only SL 4 (terminator SL) and the 3′ arm of SL 3 was still perfectly able to bind its target [15]. By contrast, both SL 2 and SL 4 of SR5 were found to be essential for formation of a stable duplex with bsrE RNA, since replacement of either of them or of both loops 2 and 4 decreased binding 6–7-fold [13].
Initial contacts between a base-pairing small regulatory RNA (sRNA) and a target mRNA can either take place between two complementary loops (e.g., fst/RNAII [20]) or between a loop and a single-stranded region (e.g., hok/Sok [21]). In both pathways, the interaction always yields a complete duplex that is cleaved by the double-strand specific RNase III.
In the case of bsrG/SR4, binding starts between the so-called recognition loops, the terminator loop L4 of SR4 and L3 of bsrG RNA, followed by base-pairing between L3 of SR4 and a stretch within the main helix of bsrG RNA. Eventually, SR4 L2 and the bsrG L3 interact, although this interaction is not required for inhibition [15] (Figure 2A). A similar binding pathway including three consecutive interactions was elucidated for bsrE/SR5 ([13] Figure 2B). However, three differences to bsrG/SR4 can be pointed out. Firstly, whereas, in bsrG/SR4, an antitoxin comprising only SL 4 and the 3′ half of SL 3 is sufficient for inhibition, two loops of SR5 are necessary for successful interaction with bsrE RNA. Secondly, two U-turn motifs (see below) are involved in the first loop–loop contact, whereas only one U-turn motif participates in the bsrG RNA/SR4 interaction. Thirdly, in contrast to SR4, SR5 does not induce structural changes around the bsrE SD, i.e., the stem topped by L1 is not extended.
In the case of txpA/RatA, interaction most probably also initiates via a loop/loop contact, but the binding pathway is yet to be investigated in detail [10]. By contrast, a concurrent interaction of two loop pairs was observed for the TA system sib/Ibs of E. coli [22], as well as earlier for RNAII/III, the replication control system of the streptococcal plasmid pIP501 [23].
In sense/antisense RNA systems, a rapid interaction between both RNAs is crucial for efficient regulation. A 5′ YUNR motif (Y, pyrimidine; N, any base; R, purine) can potentially form a U-turn with a sharp bend in the RNA backbone that presents the next three or four bases in a stacked, solvent-exposed configuration. This provides a scaffold for rapid RNA/RNA interactions. Via one or two contacts across the bend, the backbone change is stabilized. U-turn motifs were first discovered in yeast transfer RNA (tRNA) and the hammerhead ribozyme—and confirmed by crystal structure analysis—and later in 45 sense or antisense RNAs (reviewed in Reference [24]). E. coli type I TA system hok/Sok was the first sense/antisense RNA system in which a 5′ YUNR motif present in the sense (hok) mRNA was demonstrated to be decisive for a rapid RNA interaction with the Sok antisense RNA [25]. Later, this was also shown for RNAII/RNAIII of pIP501 [26]. Only in these two systems was U-turn formation experimentally confirmed by N-ethyl, N-nitrosourea (ENU) footprinting. Surprisingly, in the case of bsrG/SR4, two potentially important 5′ YUNR motifs were detected. However, whereas the motif in L3 of bsrG RNA is implicated in the initial contact with L4 of SR4, the 5′ YUNR motif in L2 of SR4 that interacts with bsrG L4 was not required, as the entire SL2 was not essential for the inhibitory function of SR4 [15]. In bsrE/SR5, two loops (L4 of bsrE RNA and L4 of SR5) each containing a U-turn motif are involved in the initial rapid interaction ([13] Figure 2B). By contrast, in txpA/RatA, neither of the interacting loops contains a 5′ YUNR motif [10]. So far, secondary structures are yet to be determined experimentally for yonT-yoyJ/SR6 and bsrH/as-bsrH. Therefore, it is unknown if 5′ YUNR motifs are present in loop regions of either toxin mRNA or RNA antitoxin.
2.4. Additional Modes to Preclude Premature Toxin Expression
Additional strategies that utilize the high capacity of RNA to fold and refold are employed by the different TA systems to ensure tight regulation of toxin expression. Whereas in Gram-negative bacteria, processing events can convert long, inactive toxin mRNAs into short, translationally competent ones (reviewed in Reference [18]), alternative approaches were found in B. subtilis. The toxin genes txpA, bsrE, and bsrG sequester their ribosome binding sites (RBS) in four or five GC base-pairs [9,13,15], which suffice to prevent efficient binding of the 30S ribosomal subunit. For bsrG, in vitro translation products could be only obtained after mutations were introduced to open the double-stranded region in the mRNA [15]. An alternative strategy was found for txpA and yonT, whose RBSs display more than 11 bp complementarity to the anti-SD in 16S rRNA. Such perfect RBSs are predicted to allow efficient recruitment, but only slow release of ribosomes (reviewed in Reference [9]). B. subtilis yonT—as Enterococcus faecalis fst—contains the rare start codon GUG that is found instead of AUG in only 14% of all genes in B. subtilis. This is expected to reduce translation efficiency. In other type I TA systems from Gram-positive bacteria, compact secondary structures comprising RNA pseudoknots were proposed to impede internal ribosome binding (reviewed in Reference [18]).
2.5. Role of Hfq
Hfq is an abundant RNA chaperone that was found—among other things—to either stabilize trans-encoded base-pairing small regulatory RNAs (sRNAs) or to promote the interaction with their target mRNAs [27,28]. The latter function is particularly important when both interacting RNAs share only a few complementary base-pairs. Among the currently known type I TA systems, this is only the case for the E. coli TA system ralR/RalA with a short (16 bp) stretch of complementarity between ralR mRNA and its antitoxin RalA. Here, indeed, a requirement of Hfq was discovered [29]. Neither the other E. coli antitoxins nor the B. subtilis antitoxins like SR4, SR5, or SR6 [11,12,14], which display ~120-nt complementarity to their toxin mRNAs, need Hfq. Although Hfq stabilizes SR5 about seven-fold, it was not required for the inhibitory function of this antitoxin [12]. Interestingly, it was recently shown in B. subtilis that the widespread RNA chaperone CsrA whose recognition sequence comprises a 5′GGA motif in a single-stranded or loop region can also restructure an mRNA to facilitate the interaction with its regulatory sRNA [30]. Therefore, it is not excluded that CsrA or other, still unidentified RNA binding proteins might accomplish the function of Hfq for toxin mRNA/antitoxin pairs with very short base-pairing regions.
2.6. RNases Involved in Antitoxin/Toxin mRNA Degradation
In the majority of cis-encoded sRNAs, RNase III, the double-strand specific RNase present in all bacteria, cleaves the antisense/sense RNA duplex to initiate target RNA degradation resulting in inhibition of gene expression. This was discovered 30 years ago for antisense RNAs encoded on plasmids, transposons, and bacteriophages (reviewed in Reference [18]). In type I TA systems, RNase III was found to cleave the hok toxin mRNA/Sok antitoxin duplex from E. coli plasmid R1 [31], and >20 years later the txpA/RatA [10], bsrG/SR4 [11], and bsrE/SR5 [12] duplexes (Table 1). Therefore, in some cases, RNase III is essential to inhibit the toxin mRNA, e.g., for RatA, SR6 (formerly as-yonT [10]), and E. coli OrzO [32]. By contrast, it is not required by B. subtilis SR4 [11], as well as the E. coli antitoxins Sok (reviewed in Reference [18]), SymR (reviewed in Reference [16]), IstR1 (reviewed in Reference [33]), and Sib (reviewed in Reference [34]). Apparently, RNase III is only necessary in cases where target control is accomplished exclusively via mRNA degradation. However, in cases where RNA antitoxins regulate steps preceding degradation like translation (SymR, Sok, IstR1), RNase III is dispensable and only eventually cleaves the antitoxin/toxin mRNA duplex.
A comparison between txpA/RatA and bsrG/SR4 from B. subtilis corroborates this hypothesis. SR4 is a bifunctional antitoxin that not only facilitates bsrG RNA degradation, but also impedes bsrG translation [15]. Neither cell lysis nor mutations in the bsrG ORF were observed in an rnc knockout strain [11] demonstrating that RNase III is not essential for toxin inhibition. RNase III cleaves bsrG mRNA 14 nt downstream of the 3′ end of complementarity with SR4 at position 185. By contrast, RNase III cleavage of txpA RNA in its duplex with RatA is essential to prevent toxin expression [10]. RatA only decreases target mRNA stability by recruiting RNase III. However, as was previously shown by in vitro secondary structure probing [10], it does not induce a txpA mRNA conformation that further inhibits translation. In the absence of RNase III, txpA RNA levels were six-fold enhanced, whereas those of RatA were not affected. RNase III is also essential for yonT RNA degradation, which shows a seven-fold higher stability in the absence of RNase III [10]. For bsrE/SR5, two- to three-fold higher amounts of bsrE RNA were found in the absence of RNase III in the logarithmic and stationary phase, respectively [12]. Three distinct cleavage sites of RNase III were mapped near the bsrE stop codon [12]. However, as BsrE is a much weaker toxin than BsrG, and only bsrE overexpression inhibits cell growth and causes cell lysis, RNase III was not found to be essential [12].
The role of RNase III for antitoxin stability differs between the various B. subtilis type I TA systems. SR5 [12] and SR6 [10] behave like RatA in that their half-lives are almost not affected by RNase III, whereas SR4 displays a 2.5-fold higher stability in an rnc mutant strain [11]. Apparently, the relative amount of antitoxin and toxin mRNA is important. Both RatA [10] and SR5 [12] are in excess over their target RNAs. SR6 is in more than 10-fold excess over yonT mRNA in complex medium and minimal medium without glucose in all growth phases. Only in minimal medium with glucose, from late log phase toward stationary growth phase, were 3:1 to 1:1 SR6/yonT RNA ratios determined [14]. By contrast, SR4 is in excess over bsrG RNA only in complex medium from the transition phase until late stationary phase [35].
Surprisingly, in the case of B. subtilis bsrH/as-bsrH, the levels of bsrH toxin mRNA are neither affected by RNase III nor by RNase Y, but by RNase J1, an endonuclease and 5′–3′ exoribonuclease. Degradation was proposed to be rather exoribonucleolytic [10]. RNase J1 also participates in the degradation of antitoxin SR5 and bsrE mRNA [12], whereas its effect on bsrG RNA and SR4 was only 1.5-fold [11,35].
An influence of the main endoribonuclease RNase Y on the degradation of both SR4 and bsrG RNA was found. Whereas bsrG RNA was two-fold less stable in the absence of RNase Y, SR4 showed a two-fold higher stability [35]. The effect on bsrE RNA was similar to that on bsrG RNA, while RNase Y did not affect SR5 [12]. The higher stability of both bsrG and bsrE RNA in the absence of RNase Y might be due to indirect effects. RNase Y also initiates degradation of RatA. Afterwards, the 5′ part of RatA alone is degraded by 3′–5′ exoribonuclease PnpA and the 3′ part by 5′–3′ exoribonuclase J1 ([10] Figure 1).
RNase R is the main 3′–5′ exoribonuclease involved in the degradation of both bsrG mRNA and SR4. In addition, the 3′–5′ exonuclease PnpA trims both SR4 and SR5 precursors from their 3′ ends to the mature antitoxin, but only marginally contributes to degradation of both RNA antitoxins [11,12]. PnpA increases the stability of bsrE RNA [12], as, in its absence, two- to 10-fold lower amounts of bsrE RNA were observed in the stationary and logarithmic growth phase, respectively. Further processing of the RNase III cleavage products of txpA RNA is also performed by PnpA [10].
In summary, all B. subtilis type I toxin mRNAs—with the exception of bsrH mRNA—are cleaved by RNase III when they are base-paired to their RNA antitoxins. RNase III cleavage is, however, only required for inhibition when the corresponding antitoxins exclusively promote RNA degradation. Various other endo- and exoribonucleases act in concert to regulate antitoxin and toxin mRNA stability.
2.7. Mode of Action of Type I Toxins
The vast majority of the currently known type I toxins are small hydrophobic peptides of less than 60 amino acids (aa) that have a similar predicted secondary structure containing an α-helical transmembrane domain (see Figure 3), which in some cases was experimentally confirmed. They are localized in the membrane/inner membrane and are suggested to create pore-like structures destroying the membrane potential and causing cell death by energy starvation (summarized in Reference [18]). Examples are the E. coli toxins Hok and TisB (reviewed in Reference [36]). Exceptions are the E. coli toxins SymE (113 aa), an RNase [37], and RalR, a DNase [29], which are both located in the cytoplasm. However, little is known about the mechanism of action and the targets of the membrane-bound hydrophobic type I toxins, predominantly due to the fact that phenotypical effects are often only detectable upon toxin overexpression.
The B. subtilis type I toxins TxpA, BsrG, BsrE, and YonT reviewed here are smaller than 60 aa and contain transmembrane domains (Figure 3). Therefore, they are proposed to be inserted into the membrane. YoyJ is the only larger toxin (83 aa) and displays two transmembrane domains.
Whereas deletion of the antitoxins RatA or SR4 causes cell lysis by the cognate toxins TxpA or BsrG on agar plates after five days [6] or one day [11], respectively, this was not the case for deletion of antitoxin SR5 regulating bsrE. Only bsrE overexpression resulted in cell lysis and impaired growth [12]. In the case of yonT-yoyJ/SR6, it was not possible to delete the antitoxin gene or to overexpress the toxin gene, whether in E. coli or in B. subtilis, indicating a high level of toxicity. For YoyJ being a weak toxin, only indirect proof could be obtained. A yoyJ overexpression plasmid could only be established in B. subtilis in the presence of the sr6 gene, but not in B. subtilis strain 1A100 that lacks the SPβ prophage region encoding yonT-yoyJ/SR6 [14].
BsrG is the only B. subtilis type I toxin that was studied in more detail [38]. In contrast to E. coli type I toxins Hok, TisB, and LdrD that kill their hosts within 30 min of induction [39,40,41], BsrG unfolds its toxic effect in growing cells via a gradual, relatively slow process. Although it is inserted into the membrane, it neither induces pores nor disrupts the membrane potential. By contrast, it causes membrane invaginations which lead to delocalization of the MreB cytoskeleton and the associated cell-wall synthesis machinery. This results in altered cell morphology, aberrant cell division sites, and finally, cell lysis. Furthermore, compaction of the nucleoid and inhibition of protein biosynthesis were observed [38]. In two independent transposon mutagenesis screens, it was impossible to identify a BsrG target (Jahn, unpublished observations), suggesting that it is an essential protein.
Our data on the effects of BsrG question the simplistic assumption mentioned above that all membrane-targeting type I toxins act like phage holins and permeabilize the membrane. Interestingly, overproduction of the type I Fst toxin in Enterococcus faecalis yields misplaced septa, resulting in daughter cells containing little or no DNA, and the previously reported increase in membrane permeability was found to be a secondary effect, presumably caused by segregation defects [42]. This suggests that more type I toxins might act similarly to BsrG.
2.8. Regulation of Toxin and Antitoxin Expression and Biological Role
For a tight control of toxin expression, the balance between the antitoxin and toxin mRNA levels is critical. High antitoxin/toxin mRNA ratios successfully prevent toxin expression and ensure cell survival. By contrast, under conditions where the toxin mRNA is in significant excess over the antitoxin, the toxin can escape repression and cause growth arrest or cell death.
Toxin expression can be regulated transcriptionally or post-transcriptionally, e.g., depending on stress conditions or the nutritional state of the cell [43]. An example for transcriptional control is SOS control found for three E. coli type I toxin genes, symE, tisB, and dinQ, which contain Lex boxes overlapping their promoters (reviewed in Reference [18]).
By contrast, temperature sensitivity described for the three B. subtilis bsrG, bsrE, and yonT RNAs [11,12,14] depends on RNA degradation, i.e., it is based on post-transcriptional control. The amount of bsrG and bsrE RNA decreases after heat shock at 48 °C or 55 °C, whereas the amount of yonT RNA increases. Both antitoxins SR4 and SR5 are not affected by heat shock, but the amount of SR6 decreases concomitantly with the increase of its toxin mRNA [14]. Refolding at 55 °C makes bsrG RNA more accessible to single-strand-specific RNases J1 and Y [35] resulting in a three- to four-fold shorter half-life at 48 °C or 55 °C compared to 37 °C [11,35]. As bsrE RNA has a similar secondary structure to bsrG RNA (compare References [13] and [15], Figure 2), it can most probably refold in a similar manner at high temperatures. The secondary structure of yonT mRNA is yet to be determined. Therefore, it is unclear whether a structural alteration or the lower abundance of SR6 under heat-shock conditions is responsible for the higher abundance of yonT mRNA.
Both bsrE/SR5 and yonT/SR6 are multistress-responsive TA systems [12,14]. The bsrE and the yonT toxin RNAs are extremely unstable upon ethanol stress, whereas their RNA antitoxins are much less affected. The instability of bsrE RNA is due to rapid degradation by RNase Y [12]. In addition, bsrE RNA is also influenced by alkaline stress, while the amount of its antitoxin SR5 is affected by various stresses, among them oxygen deficiency, pH stress, and iron limitation. Oxygen stress is the only condition under which SR5 is not in excess over bsrE RNA anymore to prevent toxin expression [12]. For yonT RNA and SR6, strong effects of vancomycin (cell-wall stress) and manganese stress were observed (four- to five-fold decreased amounts). Lower effects were found for yonT RNA and SR6 upon salt stress, iron limitation, and oxidative stress [14]. Only the iron limitation effects in both TA systems and the oxygen stress effect on SR6 are sigB-dependent, suggesting a transcriptional regulation. Most probably, bsrH mRNA is also multistress-responsive, as riboprobes used for the detection of bsrE mRNA also allowed bsrH mRNA detection in Northern blots (Müller, unpublished observation).
No difference between antitoxin expression in rich or minimal medium was observed for B. subtilis SR4 [11] and SR5 [12]. By contrast, SR6 is much higher expressed in complex medium compared to minimal CSE medium with or without glucose [14].
Whereas plasmid-encoded type I TA systems play a role in post-segregational killing, the biological role of chromosome-encoded type I TA systems, such as the homologs of hok/Sok in E. coli (e.g., Reference [44]) or fst/RNAII in E. faecalis [45], is largely unknown.
Some chromosomally encoded type I TA systems are only induced under certain metabolic or environmental stress conditions. Therefore, they could play a role in metabolic or stress adaptation by, e.g., killing the cells when nutritional sources or oxygen are exhausted. For bsrG/SR4, a 6–8-fold excess of toxin mRNA over antitoxin in transition and early stationary growth phase in complex medium was calculated, suggesting a function in growth adaptation [35]. Furthermore, the bsrG toxin mRNA is temperature-sensitive and could play a role during temperature changes [11]. By contrast, BsrE might be important under anaerobic stress, as this is the only stress condition where bsrE toxin mRNA is in excess over its antitoxin SR5 [12], while, in unstressed cells and under heat shock, iron limitation and pH stress, SR5 is in >5-fold excess and can prevent toxin expression. For yonT RNA and SR6, strong effects of vancomycin (cell-wall stress) and manganese stress were observed. However, no relative ratios of SR6 and yonT RNA were calculated under stress conditions. In a transcriptomics study, the amounts of txpA and bsrH mRNA were found to increase after exhaustion of glucose [46], suggesting a role for TxpA and BsrH under nutritional stress.
2.9. Distribution of Type I TA Systems among Different Species and Evolutionary Considerations
It is difficult to analyze sequence conservation among type I toxins, because, in these systems, not the toxin protein, but the toxin mRNA and the antitoxin interact with each other. Mutations in the toxin mRNA will not affect base-pairing with the cognate antitoxin if the latter is transcribed from the complementary DNA strand. Such type I TA systems should be more prone to mutations than the type II TA systems. Conservation analysis (by Basic Local Alignment Search Tool (BLAST) search) of BsrG supported this hypothesis. Even closely related Bacillus subtilis strains display a variety of aa exchanges in BsrG, only maintaining the transmembrane domain. Thus, identification of homologous type I toxins is difficult.
Interestingly, BLAST search also identified conserved bsrG/SR4 loci in other Bacillus species, but some of them encoding BsrH/E instead of BsrG. This might be due to an exchange of the BsrG and the BsrE/H group toxin genes by mobile genetic elements via horizontal gene transfer. Moreover, the striking similarity of the secondary structures of bsrG mRNA and bsrE mRNA, as well as SR4 and SR5, implies a common ancestor of these two TA systems, although the toxicity and perhaps the mechanism of toxin action are different. BsrG encoded on prophage SPβ belongs to the same family as TxpA encoded on the skin element, and both cause lysis on agar plates in the absence of the antitoxin. By contrast, BsrE encoded by P6 is grouped with BsrH encoded on the skin element in one family, and only overexpression of bsrE—and most probably bsrH—is toxic.
The txpA/bsrG-like TA systems can only be found in a few Bacillus species, including B. subtilis, B. halotolerans, B. amyloliquefaciens, and also a Paenibacillus species. By contrast, bsrE/bsrH-like systems could be identified in a variety of Bacillus, Lactobacillus, Halobacillus, and Enterococcus species. Moreover, BsrE is also encoded on B. subtilis plasmid pBS72, but its effect on post-segregational killing is so far unknown. The genes encoding toxins YonT and YoyJ are also widespread in various Bacillus species, and sometimes truncated variants can be found, albeit mainly with conserved transmembrane (TM) domain regions.
Together, these findings provide evidence for an interspecies distribution of B. subtilis type I TA systems by mobile genetic elements such as phages or plasmids, for which they might have originally been addiction modules.
3. Type II Toxin-Antitoxin Systems
3.1. Overview of B. subtilis type II TA Systems
The first type II TA system discovered in Bacillus subtilis was SpoIISA (YkaC)/SpoIISB/SpoIISC in 2001 [47]. In 2005, the NdoAI/NdoA (YdcE/YdcD; MazF/MazE) system was initially described as a homolog of MazF/MazE from E. coli, although a high similarity to E. coli plasmid-encoded PemK/PemI was already shown in 1998 [48]. In 2012, a set of six new and highly similar potential type II TA systems (YqcG/YqcF, YokI/YokJ, YobL/yobK, Yxid/YxxD, YeeF/YezG, and YwqJ/YwqK) were described and some of them further investigated [49]. Except for YqcG/YqcF and YokI/YokJ encoded on the skin element and on prophage SPβ, respectively, all other type II TA systems are encoded in the core chromosome. Table 2 and Figure 4 provide an overview of these type II TA systems.
Bacillus subtilis encodes a variety of further toxins or antimicrobial substances and corresponding immunity genes, e.g., SunA/SunI [50], WapA/WapI [51], SkfA, and SdpC [52]. However, most of their toxins are secreted, involved in contact-dependent growth inhibition (CDI), or do not match the criteria for typical type II toxins. Therefore, these systems are excluded here.
3.2. Toxins and Their Mode of Action
The toxin SpoIISA is a 248-aa protein with three N-terminal transmembrane (TM) domains (aa 1–87) and a cytoplasmic C-terminal domain (aa 88–248) [47,53]. Truncation studies showed that neither the N- nor the C-terminal domain alone can display the toxicity of the full-length protein [54]. Moreover, the TM domains themselves are involved in the toxicity mechanism. Deletion of TM3 or TM2/3, as well as exchange of all three TM domains via an alternative set of TM domains from SpoIIE, resulted in a non-functional toxin [53]. Also, a mutation (R38Q) in the cytosolic loop between TM1 and TM2 rendered SpoIISA non-toxic, most probably by preventing oligomerization of the N-terminal TM domains [47,53]. The cytosolic domain, resolved by crystallography, displays a globular GAF domain fold and enables SpoIISA to dimerize [55,56]. Furthermore, additional expression of the inactive SpoIISAR38Q also inactivates the otherwise toxic SpoIISA, suggesting an oligomeric state for active SpoIISA [47]. Expression of spoIISA leads to disintegration of the cytoplasmic membrane, as well as detachment from and disruption of the cell wall [47].
Expression of spoIISA in E. coli showed a similar phenotype, suggesting a function directly in the plasma membrane and independent of further interaction partners [54]. It is assumed that SpoIISA forms holin-like pores in the plasma membrane [47], supported by the structural similarity of the N-terminal domain to class I holins and the observed phenotype. The effect of SpoIISA is bactericidal and irreversible [47,54].
The toxin NdoA is a 116-aa dimeric endoribonuclease that folds like the E. coli homolog MazF [57,58,59,60]. Upon dimerization, a groove is formed that binds extended single-stranded RNA (ssRNA) specifically at the sequence 5′–UACAU–3′ [60,61]. NdoA cleaves this sequence upstream of the 5′-U resulting in a 5′-hydroxyl and a 3′-phosphate group. Cleavage activity is independent of divalent cations [54]. Overexpression of ndoA in the absence of ndoAI inhibits growth of both E. coli and B. subtilis. Toxin action is rather bacteriostatic than bactericidal [57,61].
The toxins of the YqcG group are similar to each other. They consist of 531–669 aa with a well-conserved N-terminal and a more variable C-terminal domain (last 120–150 residues), both similar to contact-dependent growth inhibition (CDI) toxins in Gram-negative bacteria [49]. The C-terminal domain, most likely an HNH-fold, exerts endonuclease activity, which was shown for YobL, YxiD, and YqcG [49]. The differences in the C-terminal domains could cause slightly different nuclease specificities. While YobL, YxiD, and YqcG show RNase activity with a differing cleavage pattern in E. coli [49], in B. subtilis, YqcG displays exclusively DNase activity [62]. The N-terminal domain is supposed to be a secretion signal for type VII secretion systems, suggesting an extracellular function [49]. The C-terminal domains of YokI, YobL, YxiD, and YqcG exert toxicity in E. coli [49]. In B. subtilis, YqcG is bactericidal and rapidly kills the vast majority of cells.
3.3. Mechanisms of Antitoxin Action
SpoIISA is inactivated by two similar antitoxins: SpoIISB (56 aa) and SpoIISC (45 aa) [47,56]. SpoIISB is a natively disordered protein able to bind to the C-terminal domains of SpoIISA dimers, resulting in a SpoIISA2·SpoIISB2 heterotetramer [55] (see Figure 4). SpoIISB wraps around the SpoIISA dimer forming comprehensive interactions with both SpoIISA subunits and covering wide surface areas [55]. Nearly all residues of SpoIISB are involved in the interaction with SpoIISA, resulting in a fast assembling and very stable complex of sub-nanomolar affinity [55]. Experiments with truncated SpoIISB showed that the C-terminal part is most crucial for antitoxin action and only the 12 N-terminal residues could be deleted without loss of function [55]. SpoIISC displays high similarity to SpoIISB. It also binds to SpoIISA and prevents toxicity [56], most likely acting in a similar way as SpoIISB. The detailed mechanism of action of the SpoIISB/C antitoxins is still unknown. There is evidence for the C-terminal domains of SpoIISA not only to dimerize, but also further multimerize to activate toxicity [56]. Subsequent SpoIISB binding to SpoIISA could prevent its further oligomerization by masking interaction sites. However, this still needs to be investigated.
NdoA is neutralized by binding of its antitoxin NdoAI. Two NdoAI proteins form a homodimer via their N-terminal RHH motifs. Each subunit displays an extended C-terminal helix that binds an NdoA dimer and blocks the RNA binding groove, resulting in an inactive and stable hexameric (NdoA)2–(NdoAI)2–(NdoA)2 complex [60] (see Figure 4). The homologous E. coli antitoxin MazE is not able to inactivate NdoA [61].
The toxicity of YqcG, YokI, YobL, and YxiD (and likely also YeeF and YwqJ) is counteracted by their cognate antitoxins [49]. Although not verified, the antitoxins likely bind at and block the active nucleolytic site.
3.4. Expression and Physiological Role
Toxin action has to be tightly controlled by the cell to avoid undesired damage. Since toxin action is only aspired under very specific circumstances, most of the time, the antitoxin has to bind and neutralize the toxin. How this is achieved is detailed below.
In the spoIISABC system, three promoters could be identified (Figure 4). The first promoter PA in front of the spoIISA gene yields a transcript covering at least spoIISA and spoIISB. A second promoter PB, about four times stronger than the first, is situated within the ORF of spoIISA and allows a strong additional transcription of spoIISB [47]. Both PA and PB enable basal transcription during growth and are strongly induced during sporulation [48]. At least PA could be additionally regulated by the late sporulation sigma factor σK [63]. This expression pattern was further confirmed on protein level, showing a basal expression of spoIISA and a 10-fold induction at the onset of sporulation [64]. The third promoter PC is less well investigated, but seems to be also induced during sporulation. All three promoters show enhanced activity under specific conditions; PA is enhanced at ethanol stress, PB during swarming and at high cell density, and PC also during biofilm formation [46,56], likely to fine-tune toxin activation in response to environmental conditions. Whereas, for some type II TA systems, an autoregulatory feedback-loop was identified, there is no such evidence for spoIISABC. Furthermore, no data about SpoIISA/B/C protein stability are yet available. However, the unstructured fold of unbound antitoxin is an indication for a relative instability of SpoIISB (and most likely also SpoIISC).
The physiological role of the spoIISABC system remains unknown. Although toxin expression seems to be highest during sporulation, antitoxin expression is also elevated at the same time. One study used a combined ndoA/spoIISA deletion strain and observed 25% less programmed cell death and an altered morphology in biofilms [65]. Although this was a combined deletion of two toxin genes, this is the only effect found so far.
The ndoA/ndoAI system is transcribed as a bicistronic ndoAI-ndoA mRNA under control of one or two ndoAI promoter(s) which are yet to be further characterized. As the gene alr located upstream of ndoAI lacks a detectable terminator, read-through transcripts from the alr promoter cannot be excluded [46] (see Figure 4). While many homologous systems like E. coli mazE/mazF are autoregulated [66], this is not yet shown for ndoAI/ndoA in B. subtilis, although a palindromic sequence at the ndoAI promoter suggests autoregulation [57]. Nucleolytic activity of NdoA was initially detected in B. subtilis crude extracts [57], suggesting a faint constitutive activity and perhaps additional activation under specific conditions.
The physiological role of mazF/mazE systems in general is under debate. Notably, in E. coli, where the system is well characterized, a role in stress response, heterogeneity, and persistence is discussed (reviewed in Reference [67]). Since NdoA cleaves a specific pentad sequence, a role in expression regulation of specific mRNAs is conceivable, taking into account that NdoA is not directly toxic to the cell. The 5′–UACAU–3′ pentad sequence was found to be enriched in genes involved in secondary metabolite biosynthesis [61]. In the absence of ndoA, cells were more resistant against antimicrobials and some environmental stressors like hydrogen peroxide, but also more sensitive to heat and high ultraviolet (UV) irradiation [68], suggesting a role in programmed cell death or stress response regulation. An “extracellular death factor”, the hexapeptide RGQQNE, produced by B. subtilis cells as a quorum-sensing molecule, activates NdoA by blocking antitoxin, but not RNA binding [69], further supporting a role during stress. Moreover, ndoA deficiency suppresses sporulation [68]. E. coli MazF generates leaderless mRNAs, as well as ribosomes with cleaved 16S rRNAs, thus establishing a modified translational machinery to expedite expression of specific genes [70]. However, B. subtilis 16S rRNA lacks a recognition site for NdoA, rendering this mechanism unlikely.
Transcription of all six yqcG homologs is highly similar and identical to their cognate antitoxin genes. Expression of both toxins and antitoxins is elevated during sporulation, under starvation, and at high cell density [46]. Additionally, yqcG transcription during sporulation was shown to be dependent on Spo0A via the SinI/SinR pathway and further via DegU [71,72]. Considering genomic and translational similarities, this is likely also true for the other homologous systems. In the case of yobL/yobK, yeeF/yezG, and yqcG/yqcF, toxin and antitoxin genes are transcribed bicistronically. By contrast, yxiD/yxxD, yokI/yokJ, and ywqJ/ywqK are located in in larger operons and are, therefore, translated from multicistronic transcripts [46].
Most likely, these six highly similar systems fulfil comparable functions, although their C-terminal toxin domains differ. The shared putative N-terminal secretion signal implies a function outside of the cell. YqcG was shown to eliminate defective cells and to reduce the number of living cells in starved cultures and biofilms [72]. This could save resources and enable a part of the population to survive longer under harsh conditions. Alternatively, the systems could provide an advantage in competitive growth by inhibiting other strains and/or provide nutrients by degrading cellular debris [49]. Both functions could benefit from several slightly different copies of the toxins.
3.5. Distribution of Type II TA Systems among Different Species and Evolutionary Considerations
Like type I TA systems, type II TA systems are thought to originate from plasmids, phages, or other mobile genetic elements where they function in post-segregational killing. However, type II TA systems are also found in well-conserved regions of the core genome, where they were not eliminated during evolution and, most likely, adopted by the cells for own regulatory fine-tuning.
The spoIISABC system is part of the core genome in many Bacillus species and also found in some non-Bacillus species, likely adopted by horizontal gene transfer [73]. However, some relatives secondarily lost the system during evolution, whereas a few even acquired a second copy [73]. In Bacillus subtilis and close relatives, the gene locus is adjacent to the PBSX prophage, but does not seem to be involved in phage maintenance. A few other homologs were also found on plasmids [73]. However, at least in the heterologous E. coli, the system was not able to maintain plasmid stability by post-segregational killing [61]. All known spoIISABC systems display a high dissimilarity with only four invariant and eight conserved SpoIISA residues [73]. The second antitoxin is a result of a gene duplication that could even have happened twice and independently in two lineages [73].
The ndoAI/ndoA (mazE/mazF) system is widely distributed among Gram-positive and Gram-negative bacteria. However, B. subtilis ndoA shares much more similarity to mazF homologs of other Gram-positive bacteria than to those of Gram-negative bacteria [61]. In addition, the target specificity differs. While E. coli MazF cleaves 5′–ACA–3′ sequences [74], B. subtilis NdoA needs a more specific and less abundant 5′–UACAU–3′ sequence [61]. Despite huge differences in aa sequence, folding structure, and endonucleolytic activity of MazF, R1 plasmid-encoded toxin Kid, F plasmid-encoded CcdB from E. coli, and NdoA from B. subtilis are well conserved [59], suggesting the same evolutionary origin.
The six YqcG homologs are very similar and most likely have a common ancestor (see Figure 5). Comparable genes can be found in various Bacillus species. In addition, similarities to the widespread CDI genes from Gram-negative bacteria are apparent. Both toxin domains, the well-conserved putative N-terminal translocation signal and the toxic C-terminal domain harboring nuclease activity, are similar (reviewed in Reference [75]). However, the Bacillus subtilis genes do not share the central spacer domain, presumably required to span the distance to another cell to realize CDI action. Interestingly the wapA gene of Bacillus subtilis resembles such a full-length CDI [51], but the sequence identity to the yqcG family members is relatively low.
4. Concluding Remarks
To date, only type I and type II TA systems were discovered in B. subtilis and investigated to some degree. Although valuable information about toxin action could be obtained for BsrG, the targets of all type I toxins are still unknown. On the one hand, they might be encoded by essential genes as seems to be the case for BsrG. On the other hand, it is challenging to purify sufficient amounts of these membrane-bound toxins for biochemical studies, even with tightly controlled expression systems, as only a few molecules suffice to kill the host cells. Furthermore, experiments are needed to elucidate whether the additional nine potential systems, whose putative toxins were grouped into the YhzE family [9], are indeed type I TA systems.
For the type II systems, the picture is different. SpoIISA seems to act like a phage holin, although more data, e.g., on its effects on the membrane potential, are needed to confirm this assumption. Whereas NdoA that resembles E. coli MazF is known to act as interferase to degrade specific mRNAs, its physiological role is still unknown. YqcG and some of its homologs act as DNases and RNases, but their sequence specificity has to be analyzed and their location confirmed. Although the crystal structures of SpoIISA and NdoA were solved, information regarding regulatory and physiological function is sparse and hard to obtain. Currently, we cannot exclude that B. subtilis and other Bacillus species might also encode type III, IV, or V TA systems. Moreover, more type I and type II systems or even combined systems could exist. More bioinformatics and subsequent experimental work is required to answer these questions.
Author Contributions
S.B. wrote the section on type I TA systems; P.M. wrote the section on type II TA systems.
Funding
This work was funded by the Deutsche Forschungsgemeinschaft, grant BR1552/10-1 (to Sabine Brantl).
Acknowledgments
We acknowledge our former colleagues and students Natalie Jahn, Christin Meißner, Christiane Ring, Robert Neubert, Céline Reif, and Charlotte Löser who participated in our work on type I TA systems.
Conflicts of Interest
The authors declare no conflicts of interest.
Abbreviations
| aa | amino acid |
| bp | base-pair |
| CDI | contact-dependent growth inhibition |
| L | loop |
| nt | nucleotide |
| RBS | ribosome binding site |
| SD | Shine Dalgarno sequence |
| SL | stem-loop |
| sRNA | small regulatory RNA |
| ssRNA | single stranded RNA |
| TA | toxin–antitoxin |
References
- Brantl, S. Bacterial type I toxin-antitoxin systems. RNA Biol. 2012, 9, 1488–1490. [Google Scholar] [CrossRef] [Green Version]
- Masuda, H.; Tan, Q.; Awano, N.; Wu, K.P.; Inouye, M. YeeU enhances the bundling of cytoskeletal polymers of MreB and FtsU, antagonizing the CbtA (YeeV) toxicity in Escherichia coli. Mol. Microbiol. 2012, 84, 979–989. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Lord, D.M.; Cheng, H.Y.; Osbourne, D.O.; Hong, S.H.; Sanchez-Torrez, V.; Quiroga, C.; Zheng, K.; Herrmann, T.; Peti, W.; et al. A new type V toxin-antitoxin system where mRNA for toxin GhoT is cleaved by antitoxin GhoS. Nat. Chem. Biol. 2012, 8, 855–861. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aakre, C.D.; Phung, T.N.; Huang, D.; Laub, M.T. A bacterial toxin inhibits DNA replication elongation through a direct interaction with a β sliding clamp. Mol. Cell 2013, 52, 617–628. [Google Scholar] [CrossRef] [PubMed]
- Harms, A.; Brodersen, D.E.; Mitarai, N.; Gerdes, K. Toxins, targets, and triggers: An overview of toxin-antitoxin biology. Mol. Cell 2018, 70, 768–784. [Google Scholar] [CrossRef]
- Silvaggi, J.M.; Perkins, J.B.; Losick, R. Small untranslated RNA antitoxin in Bacillus subtilis. J. Bacteriol. 2005, 187, 6641–6650. [Google Scholar] [CrossRef]
- Irnov, I.; Sharma, C.M.; Vogel, J.; Winkler, W.C. Identification of regulatory RNAs in Bacillus subtilis. Nucleic Acids Res. 2010, 38, 6637–6651. [Google Scholar] [CrossRef]
- Fozo, E.M.; Makarova, K.S.; Shabalina, S.A.; Yutin, N.; Koonin, E.V.; Storz, G. Abundance of type I toxin-antitoxin systems in bacteria: Searches for new candidates and discovery of novel families. Nucleic Acids Res. 2010, 38, 3743–3759. [Google Scholar] [CrossRef] [PubMed]
- Durand, S.; Jahn, N.; Condon, C.; Brantl, S. Type I toxin-antitoxin systems in Bacillus subtilis. RNA Biol. 2012, 9, 1491–1497. [Google Scholar] [CrossRef]
- Durand, S.; Gilet, L.; Condon, C. The essential function of B. subtilis RNase III is to silence foreign toxic genes. PLoS Genet. 2012, 8, e1003181. [Google Scholar] [CrossRef] [PubMed]
- Jahn, N.; Preis, H.; Wiedemann, C.; Brantl, S. BsrG/SR4 from Bacillus subtilis—The first temperature-dependent type I toxin-antitoxin system. Mol. Microbiol. 2012, 83, 579–598. [Google Scholar] [CrossRef] [PubMed]
- Müller, P.; Jahn, N.; Ring, C.; Maiwald, C.; Neubert, R.; Meißner, C.; Brantl, S. A multistress responsive type I toxin-antitoxin system: bsrE/SR5 from the B. subtilis chromosome. RNA Biol. 2016, 13, 511–523. [Google Scholar] [CrossRef] [PubMed]
- Meißner, C.; Jahn, N.; Brantl, S. In vitro characterization of the type I toxin-antitoxin system bsrE/SR5 from Bacillus subtilis. J. Biol. Chem. 2016, 291, 560–571. [Google Scholar] [CrossRef] [PubMed]
- Reif, C.; Löser, C.; Brantl, S. Bacillus subtilis type I antitoxin SR6 promotes degradation of toxin yonT mRNA and is required to prevent toxic yoyJ overexpression. Toxins 2018, 10, 74. [Google Scholar] [CrossRef] [PubMed]
- Jahn, N.; Brantl, S. One antitoxin—two functions: SR4 controls toxin mRNA decay and translation. Nucleic Acids Res. 2013, 41, 9870–9880. [Google Scholar] [CrossRef] [PubMed]
- Kawano, M. Divergently overlapping cis-encoded antisense RNA regulating toxin-antitoxin systems from E. coli: Hok/sok, ldr/rdl, symE/symR. RNA Biol. 2012, 9, 1520–1527. [Google Scholar]
- Darfeuille, F.; Unoson, C.; Vogel, J.; Wagner, E.G.H. An antisense RNA inhibits translation by competing with standby ribosomes. Mol. Cell 2007, 26, 381–392. [Google Scholar] [CrossRef]
- Brantl, S.; Jahn, N. sRNAs in bacterial type I and type III toxin/antitoxin systems. FEMS Microbiol. Rev. 2015, 39, 413–427. [Google Scholar] [CrossRef]
- Brantl, S. Regulatory mechanisms employed by cis-encoded antisense RNAs. Curr. Opin. Microbiol. 2007, 10, 102–109. [Google Scholar] [CrossRef]
- Greenfield, T.J.; Franch, T.; Gerdes, K.; Weaver, K.E. Antisense RNA regulation of the par post-segregational killing system: Structural analysis and mechanism of binding of the antisense RNA, RNAII and its target, RNA I. Mol. Microbiol. 2001, 42, 527–537. [Google Scholar] [CrossRef]
- Thisted, T.; Gerdes, K. Mechanism of post-segregational killing by the hok/sok system of plasmid R1. Sok antisense RNA regulates hok gene expression indirectly through the overlapping mok gene. J. Mol. Biol. 1992, 223, 41–54. [Google Scholar] [CrossRef]
- Han, K.; Kim, K.S.; Bak, G.; Park, H.; Lee, Y. Recognition and discrimination of target mRNAs by Sib RNAs, a cis-encoded sRNA family. Nucleic Acids Res. 2010, 38, 5851–5866. [Google Scholar] [CrossRef]
- Heidrich, N.; Brantl, S. Antisense RNA-mediated transcriptional attenuation in plasmid pIP501: The simultaneous interaction between two complementary loop pairs is required for efficient inhibition by the antisense RNA. Microbiology 2007, 53, 420–427. [Google Scholar] [CrossRef]
- Franch, T.; Gerdes, K. U-turns and regulatory RNAs. Curr. Opin. Microbiol. 2000, 3, 159–164. [Google Scholar] [CrossRef]
- Franch, T.; Petersen, M.; Wagner, E.G.; Jacobsen, J.P.; Gerdes, K. Antisense RNA regulation in prokaryotes: Rapid RNA/RNA interaction facilitated by a general U-turn loop structure. J. Mol. Biol. 1999, 294, 1115–1125. [Google Scholar] [CrossRef]
- Heidrich, N.; Brantl, S. Antisense-RNA mediated transcriptional attenuation: Importance of a U-turn loop structure in the target RNA of plasmid pIP501 for efficient inhibition by the antisense RNA. J. Mol. Biol. 2003, 333, 917–929. [Google Scholar] [CrossRef]
- Vogel, J.; Luisi, B.F. Hfq and its constellation of RNA. Nat. Rev. Microbiol. 2011, 9, 578–589. [Google Scholar] [CrossRef] [Green Version]
- Kavita, K.; de Mets, F.; Gottesman, S. New aspects of RNA-based regulation by Hfq and its partner sRNAs. Curr. Opin. Microbiol. 2018, 42, 53–61. [Google Scholar] [CrossRef]
- Guo, Y.; Quiroga, C.; Chen, Q.; McAnulty, M.J.; Benedik, M.J.; Wood, T.K.; Wang, X. RalR (a DNase) and RalA (a small RNA) form a type I toxin-antitoxin system in Escherichia coli. Nucleic Acids Res. 2014, 42, 6448–6462. [Google Scholar] [CrossRef]
- Müller, P.; Gimpel, M.; Wildenhain, T.; Brantl, S. A new role for CsrA: Promotion of complex formation between an sRNA and its mRNA target in Bacillus subtilis. RNA Biol. 2019. [Google Scholar] [CrossRef]
- Gerdes, K.; Nielsen, A.; Thorsted, P.; Wagner, E.G.H. Mechanism of killer gene activation. Antisense RNA-dependent RNase III cleavage ensures rapid turn-over of the stable hok, srnB and pndA effector messenger RNAs. J. Mol. Biol. 1992, 226, 637–649. [Google Scholar] [CrossRef]
- Wen, J.; Won, D.; Fozo, E.M. The ZorO-OrzO type I toxin-antitoxin locus: Repression by the OrzO antitoxin. Nucleic Acids Res. 2014, 42, 1930–1946. [Google Scholar] [CrossRef]
- Wagner, E.G.H.; Unoson, C. The toxin-antitoxin system tisB-istR1: Expression, regulation, and biological role in persister phenotypes. RNA Biol. 2012, 9, 1513–1519. [Google Scholar] [CrossRef]
- Fozo, E.M. New type I toxin-antitoxin families from “wild” and laboratory strains of E. coli. Ibs-Sib, ShoB-OhsC and Zor-Orz. RNA Biol. 2012, 9, 1504–1512. [Google Scholar] [CrossRef]
- Jahn, N.; Brantl, S. Heat shock induced refolding entails rapid degradation of bsrG toxin mRNA by RNases Y and J1. Microbiology 2016, 162, 590–599. [Google Scholar] [CrossRef]
- Gerdes, K.; Wagner, E.G.H. RNA antitoxins. Curr. Opin. Microbiol. 2007, 10, 117–124. [Google Scholar] [CrossRef]
- Kawano, M.; Aravind, L.; Storz, G. An antisense RNA controls synthesis of an SOS-induced toxin evolved from an antitoxin. Mol. Microbiol. 2007, 64, 738–754. [Google Scholar] [CrossRef] [Green Version]
- Jahn, N.; Brantl, S.; Strahl, H. Against the mainstream: The membrane associated type I toxin BsrG from Bacillus subtilis interferes with cell envelope biosynthesis without increasing membrane permeability. Mol. Microbiol. 2015, 98, 651–666. [Google Scholar] [CrossRef]
- Gerdes, K.; Rasmussen, P.B.; Molin, S. Unique type of plasmid maintenance function: Postsegregational killing of plasmid-free cells. Proc. Natl. Acad. Sci. USA 1986, 83, 3116–3120. [Google Scholar] [CrossRef]
- Unoson, C.; Wagner, E.G.H. A small SOS induced toxin is targeted against the inner membrane in Escherichia coli. Mol. Microbiol. 2008, 70, 258–270. [Google Scholar] [CrossRef]
- Kawano, M.; Oshima, T.; Kasai, H.; Mori, H. Molecular characterization of long direct repeat (LDR) sequences expressing a stable mRNA encoding for a 35 amino-acid cell-killing peptide and a cis-encoded small antisense RNA in Echerichia coli. Mol. Microbiol. 2002, 45, 333–349. [Google Scholar] [CrossRef]
- Patel, S.; Weaver, K.E. Addiction toxin Fst has unique effects on chromosome segregation and cell division in Enterococcus faecalis and Bacillus subtilis. J. Bacteriol. 2006, 188, 5374–5384. [Google Scholar] [CrossRef]
- Gerdes, K.; Christensen, S.K.; Løbner-Olesen, A. Prokaryotic toxin-antitoxin stress response loci. Nat. Rev. Microbiol. 2005, 3, 371–382. [Google Scholar] [CrossRef]
- Pedersen, K.; Gerdes, K. Multiple hok genes on the chromosome of Escherichia coli. Mol. Microbiol. 1999, 32, 1090–1102. [Google Scholar] [CrossRef] [PubMed]
- Weaver, K.E.; Redy, S.G.; Brinkman, C.L.; Patel, S.; Bayles, K.W.; Endres, J.L. Identification and characterization of a family of toxin-antitoxin systems related to the Enterococcus faecalis plasmid pAD1 par addiction module. Microbiology 2009, 155, 2930–2940. [Google Scholar] [CrossRef]
- Nicolas, P.; Mäder, U.; Dervyn, E.; Rochat, T.; Leduc, A.; Pigeonneau, N.; Bidnenko, E.; Marchadier, E.; Hoebeke, M.; Aymerich, S.; et al. Condition dependent transcriptome reveals high-level regulatory architecture in Bacillus subtilis. Science 2012, 335, 1103–1106. [Google Scholar] [CrossRef]
- Adler, E.; Barák, I.; Stragier, P. Bacillus subtilis locus encoding a killer protein and its antidote. J. Bacteriol. 2001, 183, 3574–3581. [Google Scholar] [CrossRef]
- Kullik, I.; Jenni, R.; Berger-Bächi, B. Sequence of the putative alanine racemase operon in Staphylococcus aureus: Insertional interruption of this operon reduces D-alanine substitution of lipoteichoic acid and autolysis. Gene 1998, 219, 9–17. [Google Scholar] [CrossRef]
- Holberger, L.E.; Garza-Sánchez, F.; Lamoureux, J.; Low, D.A.; Hayes, C.S. A novel family of toxin/antitoxin proteins in Bacillus species. FEBS Lett. 2012, 586, 132–136. [Google Scholar] [CrossRef]
- Paik, S.H.; Chakicherla, A.; Hansen, J.N. Identification and characterization of the structural and transporter genes for, and the chemical and biological properties of, sublancin 168, a novel lantibiotic produced by Bacillus subtilis 168. J. Biol. Chem. 1998, 273, 23134–23142. [Google Scholar] [CrossRef]
- Koskiniemi, S.; Lamoureux, J.G.; Nikolakakis, K.C.; T’Kint de Roodenbeke, C.; Kaplan, M.D.; Low, D.A.; Hayes, C.S. Rhs proteins from diverse bacteria mediate intercellular competition. Proc. Natl. Acad. Sci. USA 2013, 110, 7032–7037. [Google Scholar] [CrossRef]
- González-Pastor, J.E.; Hobbs, E.C.; Losick, R. Cannibalism by sporulating bacteria. Science 2003, 301, 510–513. [Google Scholar] [CrossRef]
- Makroczyová, J.; Rešetárová, S.; Florek, P.; Barák, I. Topology of the Bacillus subtilis SpoIISA protein and its role in toxin–antitoxin function. FEMS Microbiol. Lett. 2014, 358, 180–187. [Google Scholar] [CrossRef]
- Florek, P.; Muchová, K.; Pavelčíková, P.; Barák, I. Expression of functional Bacillus SpoIISAB toxin–antitoxin modules in Escherichia coli. FEMS Microbiol. Lett. 2008, 278, 177–184. [Google Scholar] [CrossRef]
- Florek, P.; Levdikov, V.M.; Blagova, E.; Lebedev, A.A.; Škrabana, R.; Rešetárová, S.; Pavelčíková, P.; Barák, I.; Wilkinson, A.J. The structure and interactions of SpoIISA and SpoIISB, a toxin-antitoxin system in Bacillus subtilis. J. Biol. Chem. 2011, 286, 6808–6819. [Google Scholar] [CrossRef]
- Melničáková, J.; Bečárová, Z.; Makroczyová, J.; Barák, I. Analysis of the Bacillus cereus SpoIIS antitoxin-toxin system reveals its three-component nature. Front. Microbiol. 2015, 6, A808. [Google Scholar] [CrossRef]
- Pellegrini, O.; Mathy, N.; Gogos, A.; Shapiro, L.; Condon, C. The Bacillus subtilis ydcDE operon encodes an endoribonuclease of the MazF/PemK family and its inhibitor. Mol. Microbiol. 2005, 56, 1139–1148. [Google Scholar] [CrossRef]
- Kamada, K.; Hanaoka, F.; Burley, S.K. Crystal Structure of the MazE/MazF Complex: Molecular bases of antidote-toxin recognition. Mol. Cell 2003, 11, 875–884. [Google Scholar] [CrossRef]
- Gogos, A.; Mu, H.; Bahna, F.; Gomez, C.A.; Shapiro, L. Crystal structure of YdcE Protein from Bacillus subtilis. Proteins 2003, 53, 320–322. [Google Scholar] [CrossRef] [PubMed]
- Simanshu, D.K.; Yamaguchi, Y.; Park, J.H.; Inouye, M.; Patel, D.J. Structural basis of mRNA recognition and cleavage by toxin MazF and its regulation by antitoxin MazE in Bacillus subtilis. Mol. Cell 2013, 52, 447–458. [Google Scholar] [CrossRef]
- Park, J.H.; Yamaguchi, Y.; Inouye, M. Bacillus subtilis MazF-bs (EndoA) is a UACAU-specific mRNA interferase. FEBS Lett. 2011, 585, 2526–2532. [Google Scholar] [CrossRef]
- Elbaz, M.; Ben-Yehuda, S. Following the fate of bacterial cells experiencing sudden chromosome loss. MBio 2015, 6, e00092-15. [Google Scholar] [CrossRef]
- Steil, L.; Serrano, M.; Henriques, A.O.; Völker, U. Genome-wide analysis of temporally regulated and compartment-specific gene expression in sporulating cells of Bacillus subtilis. Microbiology 2005, 151, 399–420. [Google Scholar] [CrossRef]
- Rešetárová, S.; Florek, P.; Muchová, K.; Wilkinson, A.J.; Barák, I. Expression and localization of SpoIISA toxin during the life cycle of Bacillus subtilis. Res. Microbiol. 2010, 161, 750–756. [Google Scholar] [CrossRef]
- Asally, M.; Kittisopikul, M.; Rué, P.; Du, Y.; Hu, Z.; Çağatay, T.; Robinson, A.B.; Lu, H.; Garcia-Ojalvo, J.; Süel, G.M. Localized cell death focuses mechanical forces during 3D patterning in a biofilm. Proc. Natl. Acad. Sci. USA 2012, 109, 18891–18896. [Google Scholar] [CrossRef] [Green Version]
- Marianovsky, I.; Aizenman, E.; Engelberg-Kulka, H.; Glaser, G. The regulation of the Escherichia coli mazEF promoter involves an unusual alternating palindrome. J. Biol. Chem. 2001, 276, 5975–5984. [Google Scholar] [CrossRef] [PubMed]
- Nikolic, N. Autoregulation of bacterial gene expression: Lessons from the MazEF toxin-antitoxin system. Curr. Genet. 2019, 65, 133–138. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Wang, X.; Drlica, K.; Zhao, X. A toxin-antitoxin module in Bacillus subtilis can both mitigate and amplify effects of lethal stress. PLoS ONE 2011, 6, e23909. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Kolodkin-Gal, I.; Engelberg-Kulka, H. Novel quorum-sensing peptides mediating interspecies bacterial cell death. MBio 2013, 4, e00314-13. [Google Scholar] [CrossRef]
- Vesper, O.; Amitai, S.; Belitsky, M.; Byrgazov, K.; Kaberdina, A.C.; Engelberg-Kulka, H.; Moll, I. Selective translation of leaderless mRNAs by specialized ribosomes generated by MazF in Escherichia coli. Cell 2011, 147, 147–157. [Google Scholar] [CrossRef]
- Molle, V.; Fujita, M.; Jensen, S.T.; Eichenberger, P.; González-Pastor, J.E.; Liu, J.S.; Losick, R. The Spo0A regulon of Bacillus subtilis. Mol. Microbiol. 2003, 50, 1683–1701. [Google Scholar] [CrossRef] [PubMed]
- Bloom-Ackermann, Z.; Steinberg, N.; Rosenberg, G.; Oppenheimer-Shaanan, Y.; Pollack, D.; Ely, S.; Storzi, N.; Levy, A.; Kolodkin-Gal, I. Toxin-Antitoxin systems eliminate defective cells and preserve symmetry in Bacillus subtilis biofilms. Environ. Microbiol. 2016, 18, 5032–5047. [Google Scholar] [CrossRef]
- Gabriško, M.; Barák, I. Evolution of the SpoIISABC Toxin-Antitoxin-Antitoxin system in Bacilli. Toxins 2016, 8, 180. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhang, J.; Hara, H.; Kato, I.; Inouye, M. Insights into the mRNA cleavage mechanism by MazF, an mRNA interferase. J. Biol. Chem. 2005, 280, 3143–3150. [Google Scholar] [CrossRef] [PubMed]
- Willett, J.L.; Ruhe, Z.C.; Goulding, C.W.; Low, D.A.; Hayes, C.S. Contact-Dependent Growth Inhibition (CDI) and CdiB/CdiA two-partner secretion proteins. J. Mol. Biol. 2015, 427, 3754–3765. [Google Scholar] [CrossRef]
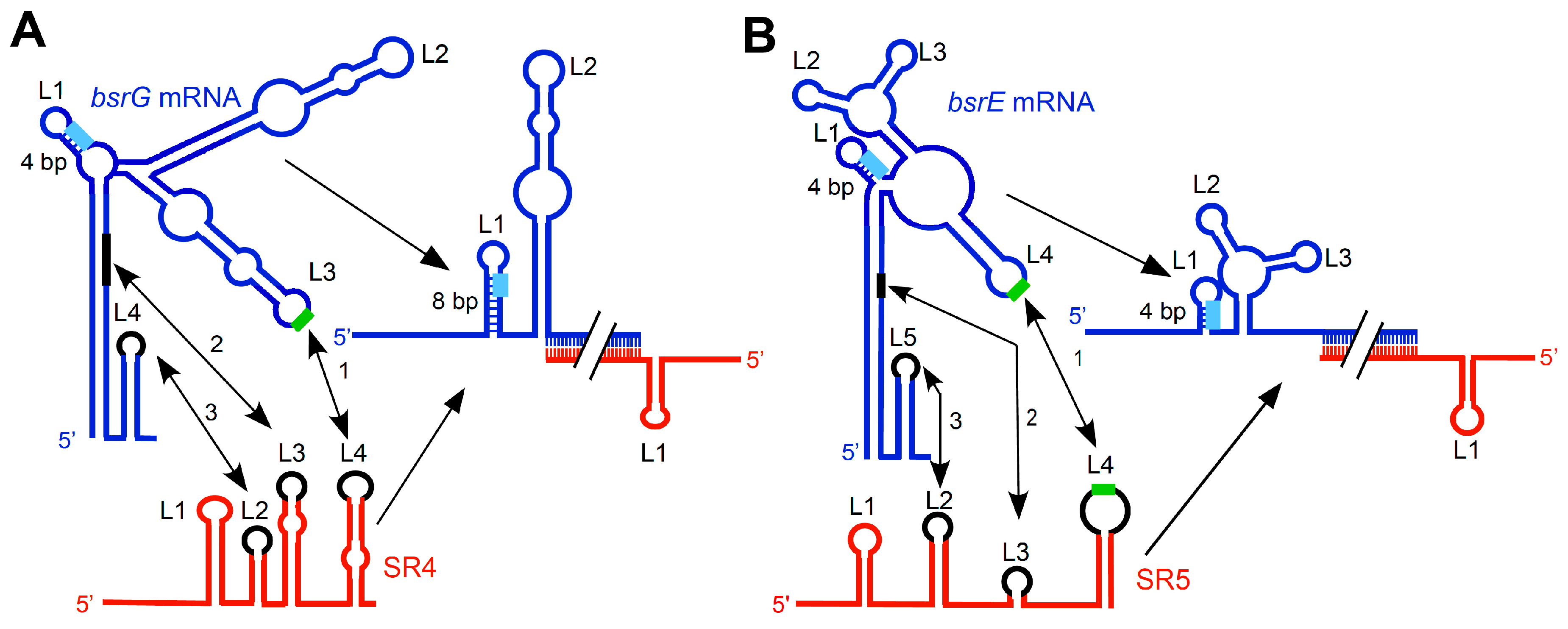
Figure 2.
Comparison of the SR4/bsrG RNA (A) and SR5/bsrE RNA (B) interaction pathways. Blue, toxin mRNAs; red, RNA antitoxins. U-turn motifs are indicated by green and SD sequences by light-blue boxes. The interaction chronology is designated by 1 to 3; L, loop. (A) The initial contact between SR4 and bsrG RNA takes place between L4 of SR4 and L3 of bsrG RNA (1). It is followed by helix progression to an interaction between SR4 loop L3 and the 3′ part of helix P1 of bsrG RNA (2), and finally reaches L2 of SR4 that binds terminator loop L4 of bsrG RNA (3). The latter interaction is not essential. (B) The binding pathway of SR5 and bsrE RNA comprises three similar subsequent interactions. The schematic secondary structures are based on the experimentally probed structures [13,15]. (A) is based on Reference [18].
Figure 2.
Comparison of the SR4/bsrG RNA (A) and SR5/bsrE RNA (B) interaction pathways. Blue, toxin mRNAs; red, RNA antitoxins. U-turn motifs are indicated by green and SD sequences by light-blue boxes. The interaction chronology is designated by 1 to 3; L, loop. (A) The initial contact between SR4 and bsrG RNA takes place between L4 of SR4 and L3 of bsrG RNA (1). It is followed by helix progression to an interaction between SR4 loop L3 and the 3′ part of helix P1 of bsrG RNA (2), and finally reaches L2 of SR4 that binds terminator loop L4 of bsrG RNA (3). The latter interaction is not essential. (B) The binding pathway of SR5 and bsrE RNA comprises three similar subsequent interactions. The schematic secondary structures are based on the experimentally probed structures [13,15]. (A) is based on Reference [18].

Figure 3.
Overview of B. subtilis type I toxins. The amino acid (aa) sequences are shown with the predicted secondary structures (Jpred4) below. Red helices indicate α-helical and blue arrows β-sheet structures. A black line indicates unstructured regions. Transmembrane domains (predicted by TMHMM v. 2.0, DTU Health Tech, Lynby, Denmark) are highlighted in gray; polar aa are shown in bold, and charged aa are red (−) and blue (+). Conserved aa are indicated with three dots, and similar aa with one dot (.) between similar sequences. Based on Reference [9].
Figure 3.
Overview of B. subtilis type I toxins. The amino acid (aa) sequences are shown with the predicted secondary structures (Jpred4) below. Red helices indicate α-helical and blue arrows β-sheet structures. A black line indicates unstructured regions. Transmembrane domains (predicted by TMHMM v. 2.0, DTU Health Tech, Lynby, Denmark) are highlighted in gray; polar aa are shown in bold, and charged aa are red (−) and blue (+). Conserved aa are indicated with three dots, and similar aa with one dot (.) between similar sequences. Based on Reference [9].

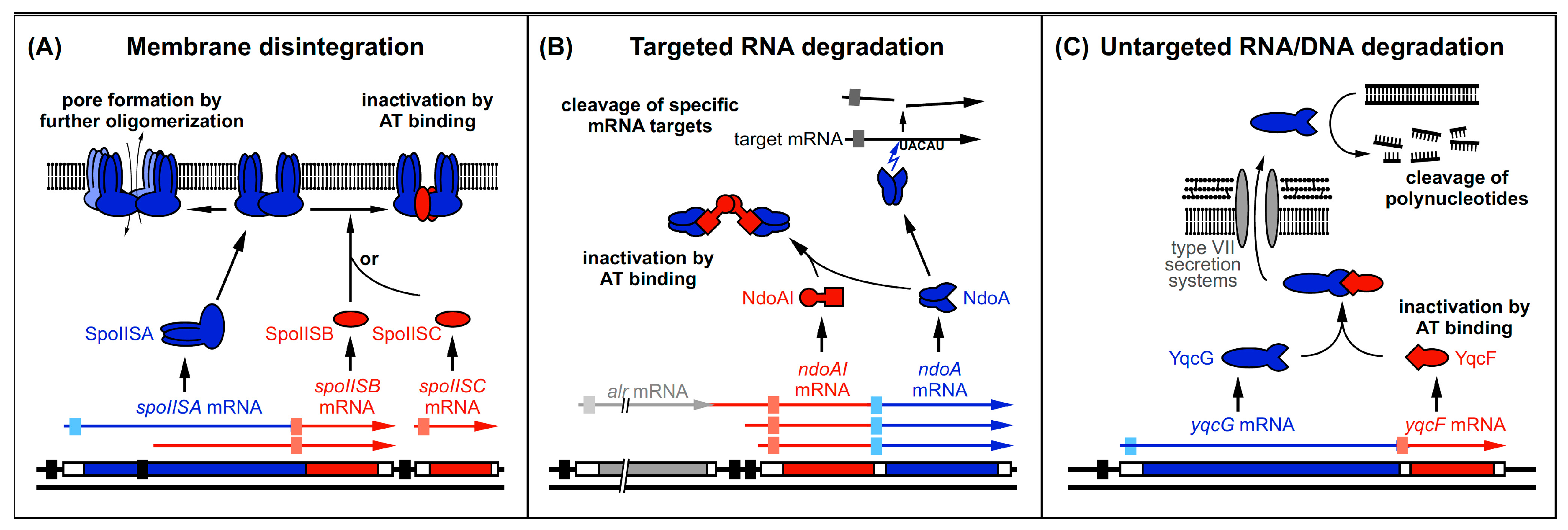
Figure 4.
Three mechanisms of toxin and antitoxin action employed by B. subtilis type II TA systems. Black bars denote promoters. Toxins are drawn in blue and antitoxins in red. The toxin ORFs are represented by blue bars. Light-blue boxes indicate SD sequences. Antitoxin ORFs are represented by red bars and light-red boxes denote SD sequences. Arrows symbolize endoribonucleolytic activity. (A) SpoIISA toxin dimerizes in the membrane and is either inactivated by SpoIISB or SpoIISC or forms pores and causes membrane disintegration. (B) NdoA dimerizes and is either inactivated by NdoAI binding or cleaves RNAs upstream of single-stranded 5′–UACAU–3′ sequences. (C) YqcG is intracellularly inactivated by its antitoxin YqcF, predicted to be secreted by type VII secretion systems to be active outside the cell. There, it is proposed to cleave DNA to allow resorption of the cleavage products. The homologous toxins are proposed to cleave RNA.
Figure 4.
Three mechanisms of toxin and antitoxin action employed by B. subtilis type II TA systems. Black bars denote promoters. Toxins are drawn in blue and antitoxins in red. The toxin ORFs are represented by blue bars. Light-blue boxes indicate SD sequences. Antitoxin ORFs are represented by red bars and light-red boxes denote SD sequences. Arrows symbolize endoribonucleolytic activity. (A) SpoIISA toxin dimerizes in the membrane and is either inactivated by SpoIISB or SpoIISC or forms pores and causes membrane disintegration. (B) NdoA dimerizes and is either inactivated by NdoAI binding or cleaves RNAs upstream of single-stranded 5′–UACAU–3′ sequences. (C) YqcG is intracellularly inactivated by its antitoxin YqcF, predicted to be secreted by type VII secretion systems to be active outside the cell. There, it is proposed to cleave DNA to allow resorption of the cleavage products. The homologous toxins are proposed to cleave RNA.

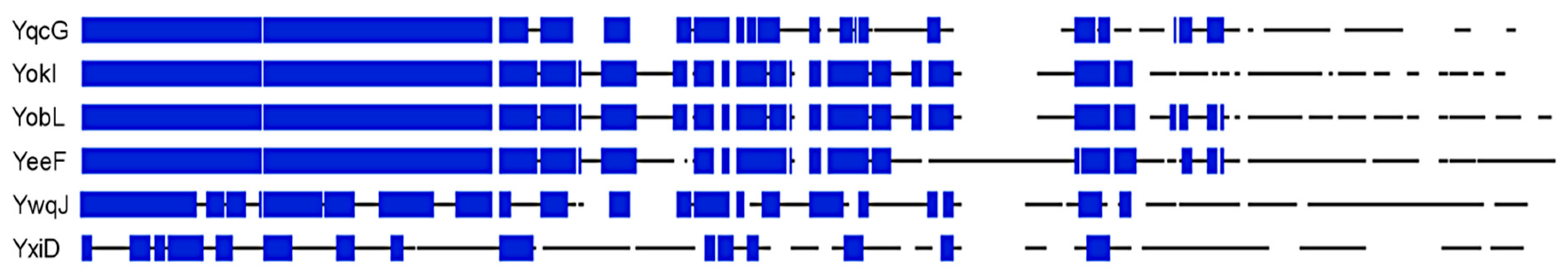
Figure 5.
Multiple alignment of aa sequences of YqcG homologs from B. subtilis. Alignment was performed using a BLOSUM 62 scoring matrix and full-length protein sequences. Sequence stretches are represented as black lines and white gaps are gaps in the alignment. Stretches of at least 5 aa with at least 60% similarity are depicted as blue boxes.
Figure 5.
Multiple alignment of aa sequences of YqcG homologs from B. subtilis. Alignment was performed using a BLOSUM 62 scoring matrix and full-length protein sequences. Sequence stretches are represented as black lines and white gaps are gaps in the alignment. Stretches of at least 5 aa with at least 60% similarity are depicted as blue boxes.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Overview of type I toxin-antitoxin (TA) systems from Bacillus subtilis: characteristics, mechanism of antitoxin action, RNases involved in degradation of toxin messenger RNA (mRNA) and RNA antitoxin, and regulation of expression.
Table 1.
Overview of type I toxin-antitoxin (TA) systems from Bacillus subtilis: characteristics, mechanism of antitoxin action, RNases involved in degradation of toxin messenger RNA (mRNA) and RNA antitoxin, and regulation of expression.
| TA System | Location on Prophage | AT (nt) | T RNA (nt) | Mode of AT Action | RNases Cleaving T/AT RNA | T (aa) | Regulation/Peculiarity |
|---|---|---|---|---|---|---|---|
| txpA/RatA | skin | 222 | 270 | RD | III *, P/III | 59 | glucose-dependent |
| bsrG/SR4 | SPβ | 180 | 294 | TI + RD | III, Y, R/III, R, Y, J1, P | 38 | temperature-dependent |
| bsrE/SR5 | P6 | 163 | 255–260 | RD | III *, Y, P/III, J1, P | 30 | multistress-responsive |
| yonT-yoyJ/SR6 | SPβ | 100/215 a | >450 | RD yonT TI yoyJ | III */III | 58 | multistress-responsive, SR6 unusually stable |
| bsrH/as-bsrH | skin | 200 | 285 | RD ? | J1/n.d. | 29 | multistress-responsive, glucose-dependent |
AT, antitoxin; T, toxin; TI, translational inhibition; RD, promotion of toxin mRNA degradation; aa, amino acid; ?, proposed but not experimentally shown. a, longer SR6 species due to read-through of SR6 terminator. III, RNase III; Y, RNase Y; R, RNase R; J1, RNase J1; P, PnpA. * Essential for inhibition by the antitoxin; n.d., not determined. For further details, see text.
Table 2.
Overview of type II TA systems from B. subtilis: toxin and antitoxin properties, toxin target, and antitoxin necessity.
Table 2.
Overview of type II TA systems from B. subtilis: toxin and antitoxin properties, toxin target, and antitoxin necessity.
| Toxin (T) | Antitoxin (AT) | T (aa) | AT (aa) | T Target | AT Essential |
|---|---|---|---|---|---|
| SpoIISA (YkaC) | SpoIISB | 248 | 56 | plasma | no |
| SpoIISC | 248 | 45 | membrane | no | |
| NdoA (YdcE, MazF) | NdoAI (YdcD, MazE) | 116 | 93 | 5′–UACAU RNA | not determined |
| YqcG | YqcF | 531 | 192 | DNA | yes |
| YokI | YokJ | 571 | 165 | DNA or RNA | no |
| YobL | YobK | 600 | 152 | RNA | no |
| YxiD | YxxD | 569 | 147 | RNA | yes |
| YeeF | YezG | 669 | 151 | DNA or RNA | yes |
| YwqJ | YwqK | 602 | 154 | DNA or RNA | no |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Brantl, S.; Müller, P. Toxin–Antitoxin Systems in Bacillus subtilis. Toxins 2019, 11, 262. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins11050262
AMA Style
Brantl S, Müller P. Toxin–Antitoxin Systems in Bacillus subtilis. Toxins. 2019; 11(5):262. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins11050262
Chicago/Turabian StyleBrantl, Sabine, and Peter Müller. 2019. "Toxin–Antitoxin Systems in Bacillus subtilis" Toxins 11, no. 5: 262. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins11050262
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.