Effects of Different Carbon Sources on Fumonisin Production and FUM Gene Expression by Fusarium proliferatum


1
Key Laboratory of Plant Resource Conservation and Sustainable Utilization, Guangdong Provincial Key Laboratory of Applied Botany, Key Laboratory of Post-harvest Handling of Fruits, Ministry of Agriculture, South China Botanical Garden, Chinese Academy of Sciences, Guangzhou 510650, China
2
School of Life Sciences, University of Chinese Academy of Sciences, Beijing 100039, China
3
Zhongshan Entry-Exit Inspection and Quarantine Bureau, Zhongshan 528403, China
*
Author to whom correspondence should be addressed.
Toxins 2019, 11(5), 289; https://0-doi-org.brum.beds.ac.uk/10.3390/toxins11050289
Submission received: 14 April 2019
/
Revised: 9 May 2019
/
Accepted: 14 May 2019
/
Published: 22 May 2019
(This article belongs to the Special Issue Molecular Genetics of Secondary Metabolite Biosynthesis in Fungal that Interact with Plants and Animals)
Abstract
:Fusarium proliferatum can infect many crops and then produce fumonisins that are very harmful to humans and animals. Previous study indicates that carbon sources play important roles in regulating the fumonisin biosynthesis. Unfortunately, there is limited information on the effects of carbon starvation in comparison with the carbon sources present in the host of fumonisin production in F. proliferatum. Our results indicated that F. proliferatum cultivated in the Czapek’s broth (CB) medium in the absence of sucrose could greatly induce production of fumonisin, while an additional supplementation of sucrose to the culture medium significantly reduced the fumonisin production. Furthermore, cellulose and hemicellulose, and polysaccharide extracted from banana peel, which replaced sucrose as the carbon source, can reduce the production of fumonisin by F. proliferatum. Further work showed that these genes related to the synthesis of fumonisin, such as FUM1 and FUM8, were significantly up-regulated in the culture medium in the absence of sucrose. Consistent with fumonisin production, the expressions of FUM gene cluster and ZFR1 gene decreased after the addition of sucrose. Moreover, these genes were also significantly down-regulated in the presence of cellulose, hemicellulose or polysaccharide extracted from peel. Altogether, our results suggested that fumonisin production was regulated in F. proliferatum in response to different carbon source conditions, and this regulation might be mainly via the transcriptional level. Future work on these expressions of the fumonisin biosynthesis-related genes is needed to further clarify the response under different carbon conditions during the infection of F. proliferatum on banana fruit hosts. The findings in this study will provide a new clue regarding the biological effect of the fumonisin production in response to environmental stress.
Keywords:
Fusarium proliferatum; fumonisin biosynthesis; carbon source; environmental stress; gene expressionKey Contribution: F. proliferatum can produce fumonisins which are harmful to human health. This study aimed to explore the effect in the absence; followed by addition of carbon sources on fumonisin production; which will be beneficial for understanding the infection of F. proliferatum into banana fruit.
1. Introduction
Mycotoxins as secondary metabolites produced by some fungi are capable of causing disease and even death in humans and other animals [1]. Fumonisins (FBs) were first discovered in the late 1980s and received worldwide attention due to their toxicity for humans or animals [2]. In humans, FB has been shown to be well correlated to a high incidence of esophageal cancer in South Africa [3]. The FBs consists of FB1, FB2, FB3 and FB4, of which FB1 is the major component [4]. On the basis of available toxicological evidence, the World Health Organization’s International Program known as the International Cancer Research Institute released a reference of carcinogen list in 2017 and classified fumonisin B1 (FB1), fumonisin B2 (FB2) and fusarium C as class 2B carcinogens. FB1 have the largest proportion of the total fumonisins accounting for up to 70% while FB2 and FB3 usually make up 10%–20% of the total fumonisin content [5]. FB1 mainly contaminated maize and its products [5], leading to two diseases [6] which occur in domestic animals: equine leukoencephalomalacia and porcine pulmonary edema syndrome. The two diseases involve disordered sphingolipid metabolism and cardiovascular disease [7,8]. The products from Fusarium verticillioides and Fusarium proliferatum are their main sources. Fumonisins B (FBs) are major mycotoxins synthesized by F. proliferatum [9]. F. proliferatum colonizes a broad range of host plants, such as maize, wheat, asparagus, banana, and various conventional crops or profitable crops [10,11]. In these hosts, root rot disease [12,13] and black point symptoms [14] have been caused by infection of F. proliferatum. Moreover, F. proliferatum can produce various toxic secondary metabolites, such as fumonisins [15], beauvericin [16], fusaric acid [17], and nontoxic secondary metabolites like bikaverin [18].
The biosynthetic pathway of FBs has been well documented. Fumonisins consist of a 19 to 20 carbon aminopolyhydroxyalkyl chain that generally undergoes four steps: reductions of carbonyl compounds, hydroxylation, alanine condensation, and esterification of two tricarboxylic acids [19]. Moreover, the gene cluster (FUM) of the fumonisin biosynthesis has been identified in F. proliferatum [20]. However, the FUM does not contain a pathway-specific regulatory gene, unlike other fungal secondary metabolite gene clusters [21]. In addition, several genes appear to regulate fumonisin biosynthesis such as a Zn(II)2Cys6 DNA binding protein (ZFR1), which is not linked to the FUM but rather encodes a polypeptide with significant homology to fungal proteins that contain a DNA binding motif consisting of a Zn(II)2Cys6 binuclear cluster [22,23]. Additionally, the possible fumonisin biosynthesis mechanism was also reported [24]. Carbon sources including sucrose and glucose have been proven to affect greatly fumonisin production [25]. Meanwhile, sugar is one of the nutrition components in edible fruits and can provide the main carbon source for fungi during their infection process. Stepien et al. [26] reported that F. proliferatum strains from different hosts are genetically diverse while host plant extracts can change the expression patterns of FUM and fumonisin production.
Banana, as one of the most economically important fruit crops worldwide, deteriorates easily due to rot development caused by postharvest pathogens, including F. proliferatum [25]. Banana fruit as a hosts for these types of pathogens can provide nutrition for F. proliferatum infection, especially the banana peel is rich in sugar ingredients like cellulose, hemicellulose and pectin as carbon sources when the fruit becomes edible. F. proliferatum might face these polysaccharides while infecting banana fruit. Additionally, banana peel is the first natural infection place for F. proliferatum, and, thus, F. proliferatum mainly grows on the banana peel after infection. Hence, investigation of the effect of the polysaccharides from banana peel on the fumonisin production from F. proliferatum might enable better understanding of the interaction between banana fruit and F. proliferatum.
Recent research suggests that Colletotrichum spp., Alternaria alternate and Fusarium oxysporum could alkalize the host plant for better infection. In contrast, a Penicillium spp. can acidify the host environment for attack [27,28,29]. In addition, carbon availability in the environment is a key factor for triggering the host pH change [30]. As is known, one of the most significant changes is a rapid increase of sugar content during fruit ripening. Unfortunately, there is currently a lack of research on the response of mycotoxin biosynthesis in a pathogen with the carbon status of the host plant and carbon stress. In this study, we investigated the production of fumonisin in the absence of sucrose and an addition of sucrose under the lack carbon condition in F. proliferatum. Additionally, the fumonisin production in F. proliferatum by cellulose, hemicellulose or polysaccharide extracted from banana peel instead of sucrose was also investigated. The expressions of these genes involved in fumonisin biosynthesis were conducted to understand the underlying mechanism. This study might provide new information on the regulation of fumonisin biosynthesis in response to different nutrition environments of F. proliferatum.
2. Results
2.1. Effect of Different Sucrose Conditions on the Growth, Sporulation and FB1 Content of F. Proliferatum
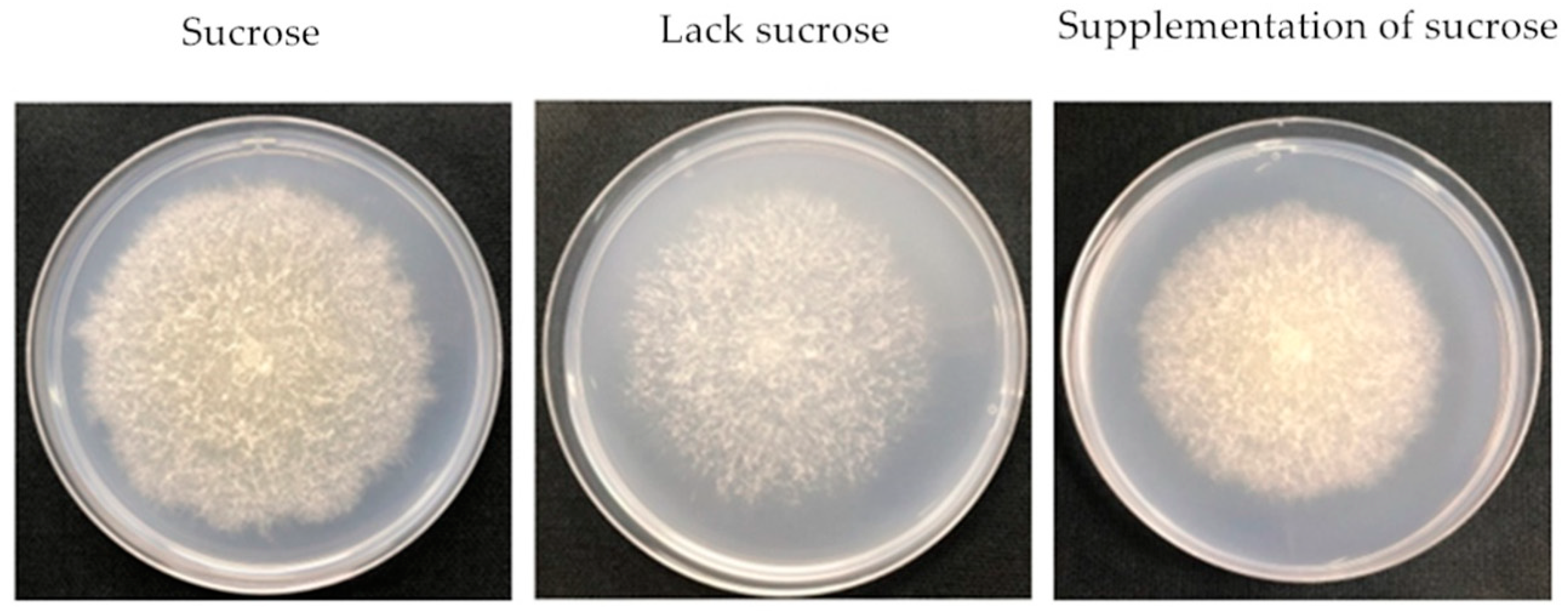
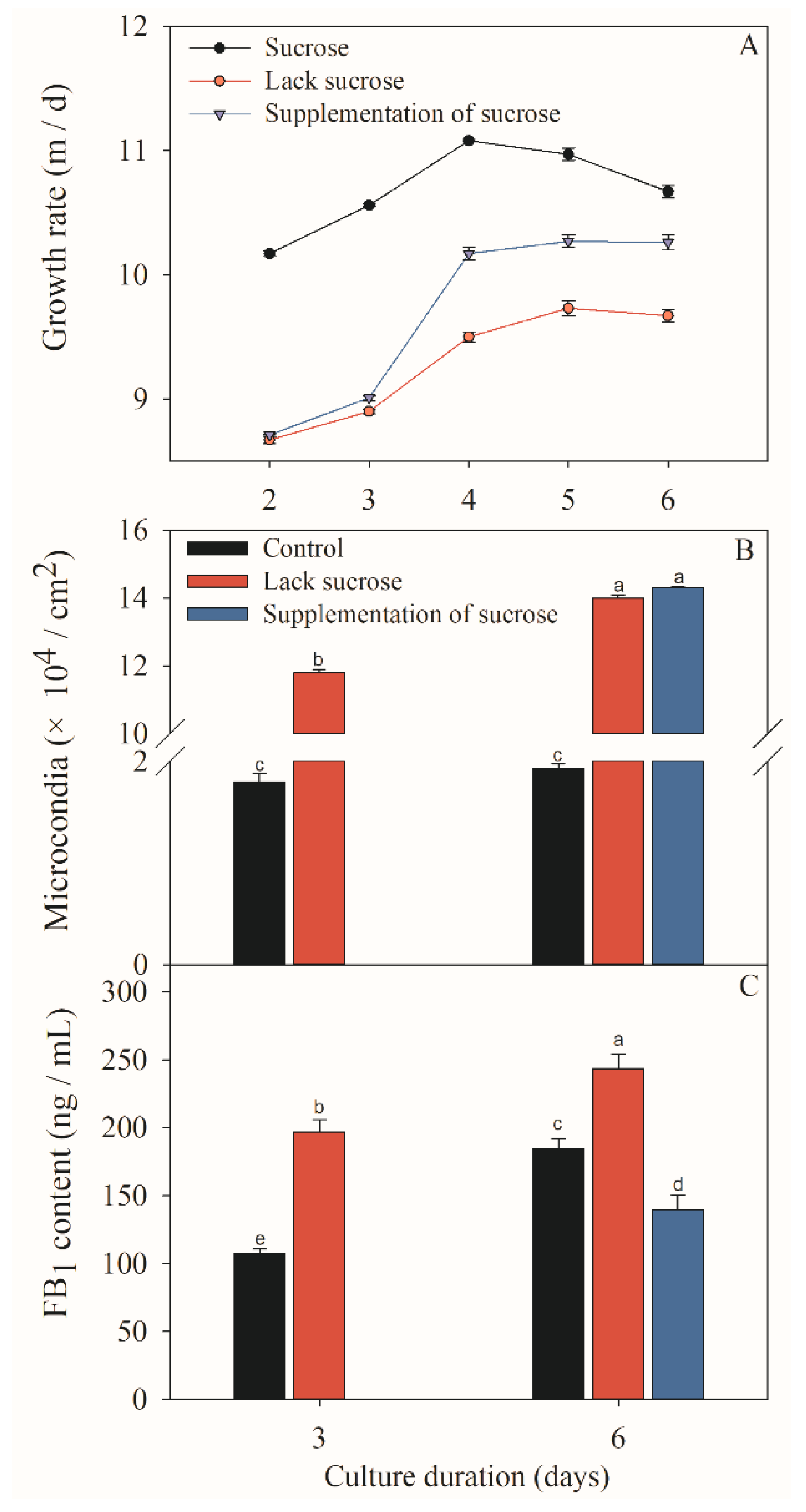
The F. proliferatum strain was cultivated in different culture media while mycelial growth rate, sporulation and FB1 production were checked after 3 and 6 days. Colony morphology varied when F. proliferatum was cultured in different media (Figure 1). In short, the F. proliferatum showed a better mycelial growth rate and more sporulation in the medium in the presence of sucrose than the medium in the absence of sucrose. Furthermore, an additional sucrose supplementation recovered the growth of F. proliferatum, which was almost the same as the medium in the presence of sucrose (Figure 2A,B). For FB1 content, the study indicated that F. proliferatum can produce FB1 in the medium with or without sucrose, but FB1 content in the medium without sucrose was significantly (p < 0.05) higher than the culture medium with sucrose, while an additional supplementation of sucrose after 3 days of culture in the medium without sucrose significantly (p < 0.05) inhibited the FB1 production (Figure 2C) according to the Duncan’s multiple comparison in ANOVA analysis. Thus, supplementation of sucrose recovered the growth but also inhibited the FB1 production by F. proliferatum, suggesting that the sucrose starvation increased FB1 production.
2.2. Effect of Different Carbon Sources on the Growth, Sporulation and FB Content of F. Proliferatum
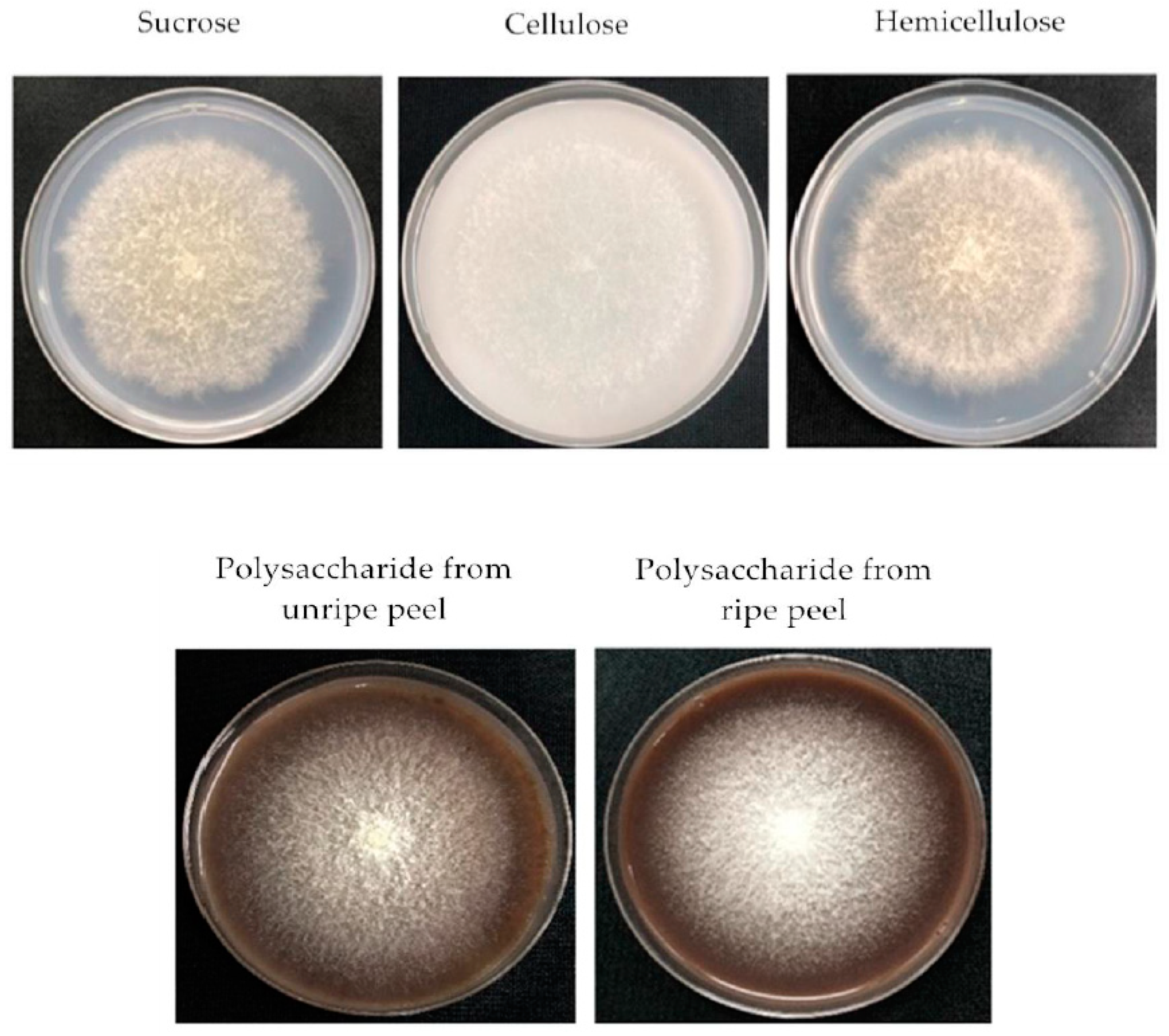
After the F. proliferatum strain was cultivated in the culture media containing five different carbon sources, mycelial growth rate, sporulation and FB content were investigated after 3 and 6 days. A different colony morphology of F. proliferatum was observed when five carbon sources were used (Figure 3) while the growth rate was almost the same as that on the 6th day (Figure 4A), but the sporulation of F. proliferatum was induced in the media containing cellulose, hemicellulose, and polysaccharide extracted from banana peel (Figure 4B). Furthermore, the contents of FB1 and FB2 produced by F. proliferatum in the culture media with cellulose, hemicellulose and polysaccharide extracted from banana peel were significantly (p < 0.05) lower than that in the culture medium with sucrose according to the Duncan’s multiple comparison in ANOVA analysis (Figure 4C,D).
2.3. Effect of Different Carbon Sources on the Expressions of FB-Related Genes
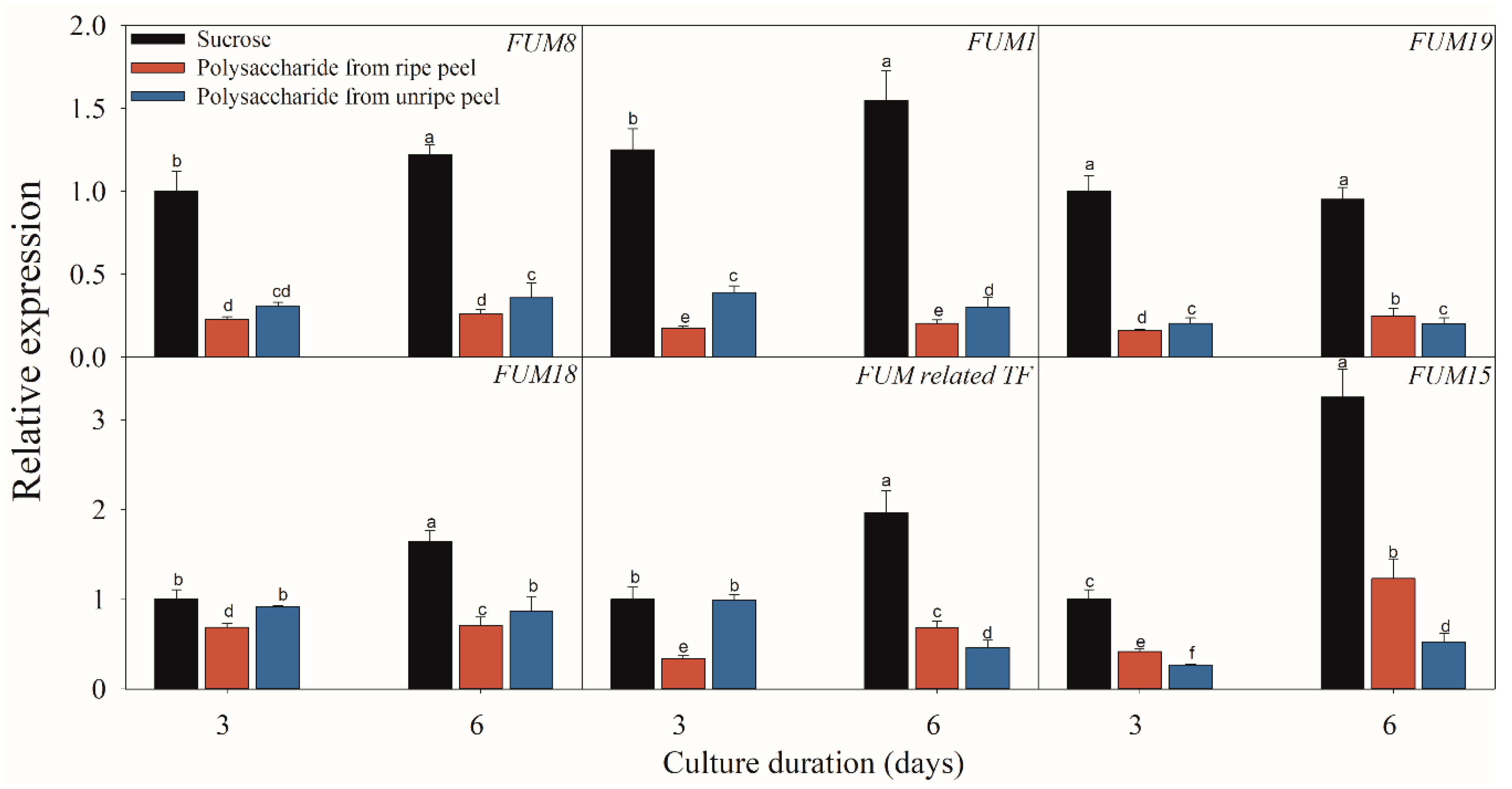
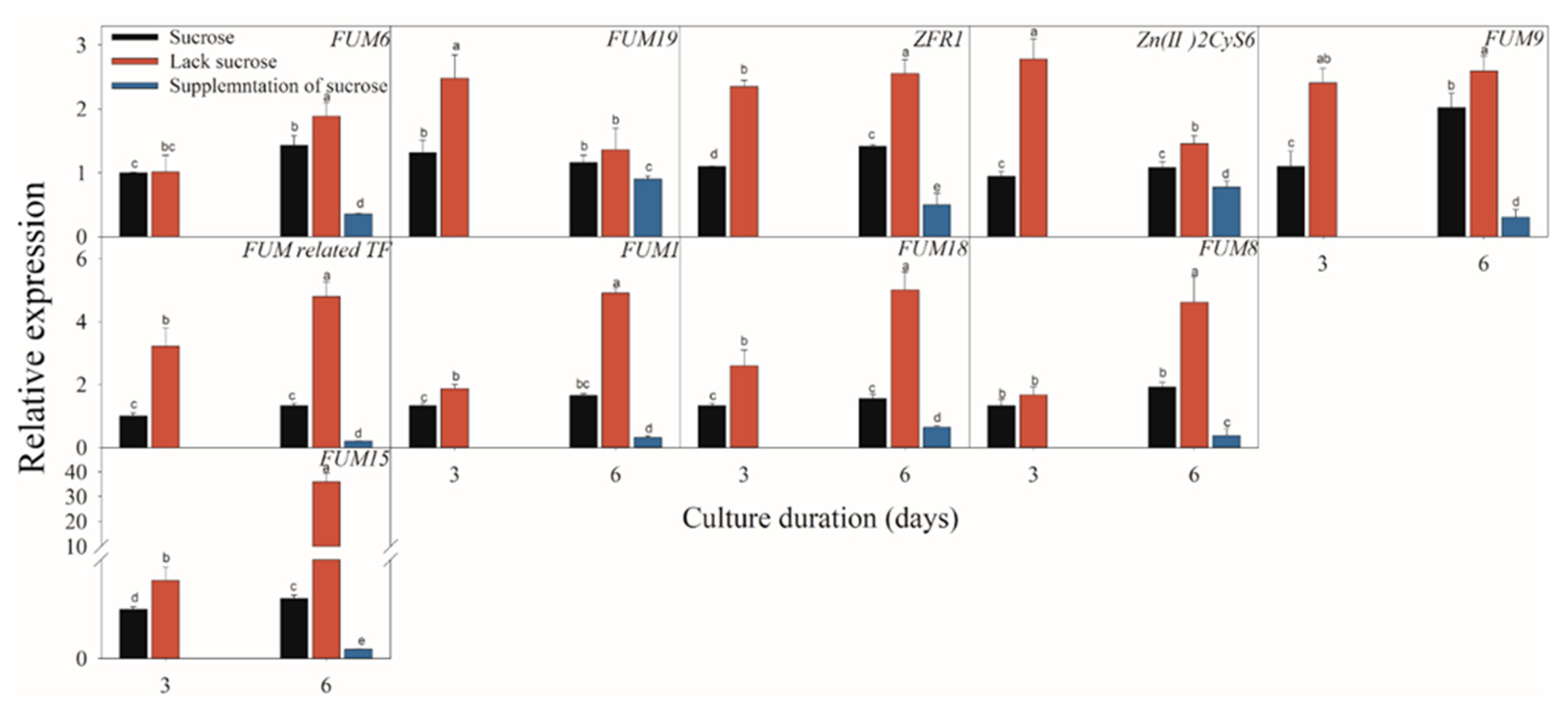
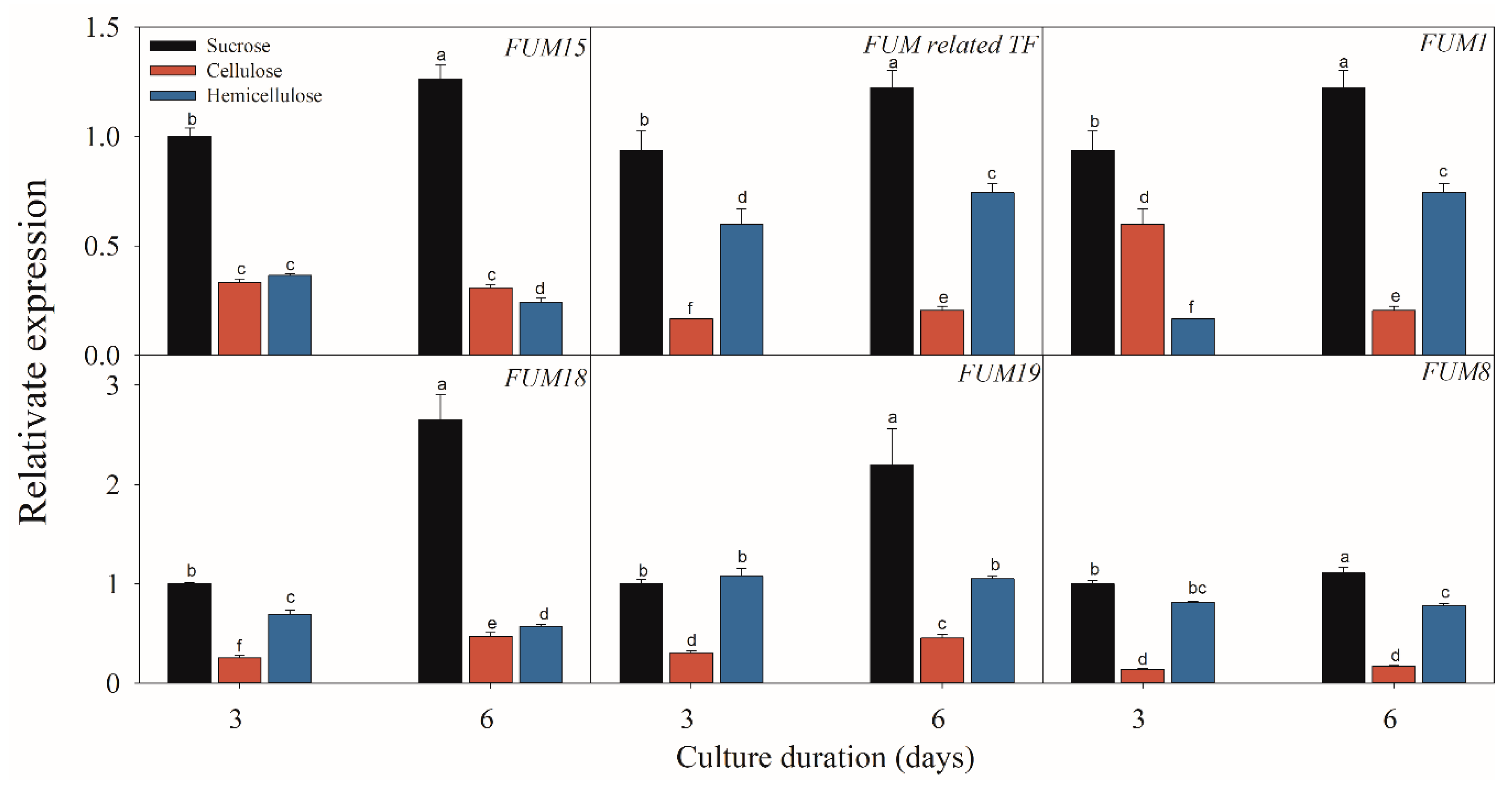
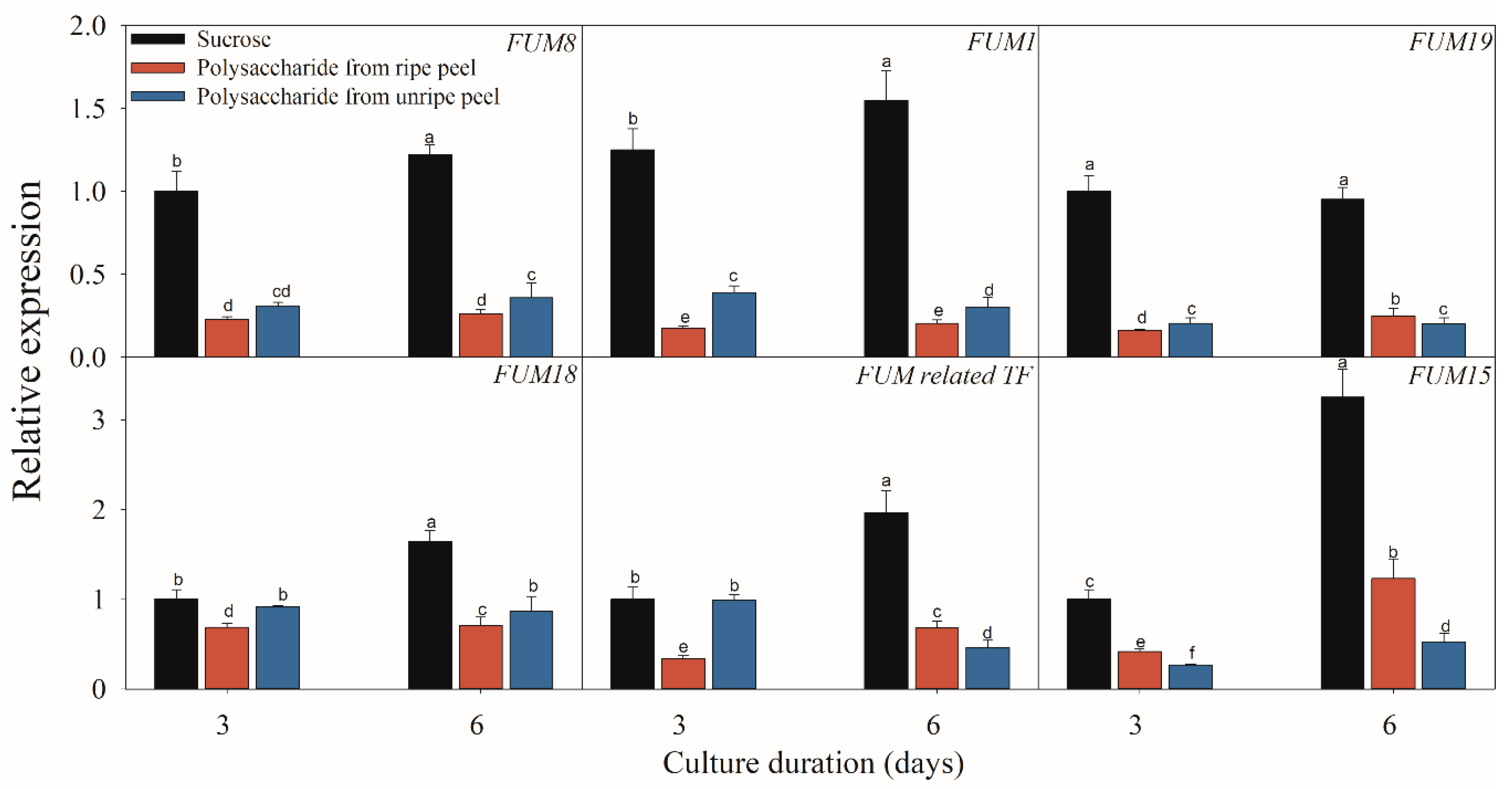
Figure 5, Figure 6 and Figure 7 present the results of the expressions of FUM and ZFR1 of the F. proliferatum cultured with different carbon media. The expressions of these genes were clearly induced in the medium in the absence of sucrose, with 1–5 folds higher than those in the medium with sucrose. After supplementation of sucrose to the medium in the lack sucrose, the expression levels of these genes were reduced significantly (Figure 5). Furthermore, these genes were significantly (p < 0.05) reduced in the culture media containing cellulose, hemicellulose (Figure 6), or polysaccharide from banana peel according to the Duncan’s multiple comparison in ANOVA analysis (Figure 7). These results further confirmed that the lack of sucrose greatly induced the expressions of FUM and ZFR1.
3. Discussion
Contamination by fumonisins is an important issue that affects crop quality and human health. To control FB production, attention has been paid to these key factors that greatly affect the synthesis and its mechanisms. Previous studies have exhibited that environmental and abiotic factors, such as carbon source, nitrogen source and pH, greatly influenced the FB biosynthesis of F. proliferatum [31,32]. In the present study, we comparatively evaluated the effects of lack carbon source and additional carbon sources such as sucrose, cellulose, hemicellulose and polysaccharide from banana peel on the growth and FB biosynthesis of F. proliferatum. The Supplementary Figures S1 and S2 present polysaccharide information obtained from banana peel. Our results exhibited that the microconidia growth generally increased in the presence of cellulose, hemicellulose or polysaccharide extracted from banana peel (Figure 4B) but decreased in the absence of sucrose (Figure 2B). The previous study also demonstrated that a sufficient carbon source is beneficial for mycelial and conidia growth [33]. In contrast, FB1 production of F. proliferatum was induced in response to lack carbon stress but it was inhibited in the culture medium containing carbon source, which was in agreement with the result of Kohut et al. [34], who reported that nitrogen starvation stress induced FUM expression and increased fumonisin production in F. proliferatum. It is interesting to note that FB1 or FB2 production from F. proliferatum cultivated in the culture medium containing the polysaccharide extracted from ripe banana peel was lower than unripe banana peel. The result may be due to the difference in the degradation of polysaccharide during banana fruit ripening. Thus, carbon sources played a key role in growth and fumonisin biosynthesis of F. proliferatum. Importantly, carbon starvation encouraged the FB production of F. proliferatum, but its mechanism needs to be elucidated further.
Fungal pathogens are able to modulate environment pH to increase their infective potential [35]. In the case of carbon excess pathogens can induce acidification while, in contrast, alkalization occurs under carbon deprivation condition [28]. During banana fruit ripening, starch is degraded gradually into sucrose [36], and sugar can be oxidized into carbon dioxide through tricarboxylic acid, whereas an enhanced glycolysis rate and production of organic acids help to secrete metabolites that decrease the host pH value, which could result in activation of some genes to enable fungi to use a specific set of pathogenicity factors to infect the host [30]. Moreover, the host pH environment could affect the production of FBs in F. proliferatum. These findings exhibit a high biological relevance because F. proliferatum infection may undergo a transition from alkalization to acidification as the sugar contents gradually increase during the ripening of banana fruit. In addition, the production of FBs produced by F. proliferatum was lower in the culture medium containing polysaccharides extracted from ripe peel than from an unripe peel (Figure 4C,D). These results were consistent with the report of Li et al. [37], who found that the production of FBs produced by F. proliferatum was significantly inhibited under the acidification condition.
Previous research indicated that mycotoxin biosynthesis could be mainly regulated at the transcriptional level [38]. To further investigate the possible mechanism of different carbon sources involving in FB biosynthesis, we examined the expression profiles of these related genes related to the FB biosynthesis pathways. The expressions of the crucial FB biosynthesis-related genes were affected greatly by various environmental factors [39]. Considering the FB pathway in F. proliferatum, real-time reverse transcription PCR (RT-PCR) assays were used. FUM1, FUM3, FUM6, FUM8, FUM15, FUM18 and FUM19 belonging to the member of FUM cluster includes 17 genes, as designated to be FUM1, FUM2, FUM3, FUM6, FUM7, FUM8, FUM10, FUM11, FUM13, FUM15, FUM18 and FUM21, respectively [40,41,42]. In this study, the expressions of these genes in F. proliferatum demonstrated a positive relationship with the FB production under different carbon conditions. For example, when F. proliferatum was cultured in the medium without sucrose, FB1 content was significantly induced with significantly up-regulated expressions of these genes. In particular, FUM1 encodes a polyketide synthase in the early step of participating the assembly of the FB backbone, while FUM8 is an aminotransferase gene which catalyzes the formation for a biologically active FB1 molecule. A previous study confirmed the positive relationship between the expression of these two genes and FB production [25]. In addition, ZFR1 encodes DNA-binding proteins containing a zinc binuclear cluster (Zn(II)2Cys6) belonging to the Gal4p family of transcriptional factor, regulates diverse pathways and acts as a positive regulator of FB1 biosynthesis in F. proliferatum [43,44]. FUM15 and FUM18 encoding cytochrome P450 monooxygenases and longevity assurance factor, respectively, were reported to be correlated with the FB production [41]. In the present study, the induced expressions of all these genes were in agreement with increased FB production when F. proliferatum was cultured in the medium in the absence of sucrose, while, in contrast, when additional supplementation of sucrose to the medium occurred, these gene expressions were significantly decreased in association with the reduced FB1 content. Jayashree and Subramanyam (2000) reported that some stress factors greatly affected mycotoxin production from fungi [45]. For example, FB1 production from F. proliferatum was enhanced by nitrogen starvation stress [34]. Therefore, carbon starvation stress can mediate the regulation of FB biosynthesis in F. proliferatum. Moreover, when F. proliferatum was cultured in the medium containing polysaccharide from ripe banana peel, the FB1 and FB2 contents and the expression levels of FUM cluster were significantly lower than the medium with unripe banana peel, which was in agreement with the previous report which indicated that a sufficient carbon source was only beneficial for fungal growth [33].
In general, our results showed that a carbon resource greatly influenced fungal growth and secondary metabolites. Based on the present results, we hypothesized that the changed fumonisin production might be a response of F. proliferatum to nutrition environmental stress to help to infect banana fruit host, which needs to be investigated further.
4. Conclusions
In this study, the different carbon sources significantly affected the FB biosynthesis in F. proliferatum. Results exhibited that F. proliferatum can regulate the FB production in response to different nutrition conditions while the regulation was performed via the transcriptional level. Importantly, F. proliferatum enhanced the FB biosynthesis with increased expression levels of FUM cluster and ZFR1 in the absence of sucrose. In addition, when F. proliferatum was cultured in the medium containing the polysaccharide extracted from unripe banana peel, the higher contents of FB1 and FB2 in association with the increased expression levels of FUM gene cluster were obtained and compared with the medium containing the polysaccharide extracted from ripe banana peel. Future work on the expressions of the FB biosynthesis-related genes is needed to further clarify the infection ability of F. proliferatum on banana fruit as a host.
5. Materials and Methods
5.1. Fungal Strain and Growth Condition
The strain of F. proliferatum was originally isolated from decayed banana fruit and was routinely maintained in the laboratory on potato dextrose agar (PDA) (Oxoid, Basingstoke, Hampshire, England) at 28 °C. The spores were washed from the PDA plate with sterile water, and then conidia were counted with a hemocytometer and then diluted to a concentration of 1 × 107 conidia/mL before 2 mL of the diluted spore suspension was inoculated to the Czapek’s broth (CB) medium. The medium was prepared for culture of F. proliferatum according to the method of Li et al. [46]. Conical flasks (250 mL) were prepared, containing 100 mL of the CB medium (3.0 g/L NaNO3, 1.0 g/L K2HPO4, 0.5 g/L MgSO4·7H2O, 0.5 g/L KCl and 0.01 g/L FeSO4) supplemented with 30 g/L sucrose (Aladdin, Shanghai, China), cellulose, hemicellulose (FeiBo, Guangzhou, China), or polysaccharide extracted from ripe or unripe banana peel, as shown in following Section 5.2, and then sterilized for 20 min at 121 °C. The conical flasks were incubated at 28 °C with 200 rpm and were shaken in the dark for sporulation, fumonisin and molecular analyses. 5 μL of conidia suspension was inoculated on CB plates with supplemented with 1.5% agar and then used for the morphological and growth assessments. Three biological replicates were conducted.
5.2. Polysaccharide Extraction
Ripe fruit of banana (Musa acuminate L. AAA group, cv. Brazilian) with a fully yellow skin and unripe fruit at harvested were obtained from a commercial orchard in Guangzhou, China. Banana peel tissues were collected, frozen with liquid nitrogen and smashed into powder with a pulverizer, respectively. Polysaccharides were extracted by the method of John et al. [47] with some modification. Briefly, 100 g of power from twelve banana fingers was homogenized with 1 L of distilled water and then incubated at 105 °C for 2 h. The extract was filtered through gauze and then concentrated by a vacuum rotary evaporator (Eyela N1100 V-W, Tokyo Rikakikai Co. Ltd., Tokyo, Japan). Anhydrate ethanol was added into the extract to obtain a final concentration of 60% (v/v) and then maintained for 12 h at 4 °C. The obtained precipitate was dissolved in distilled water and the solution was dialyzed against running tap water for 24 h. Finally, the solution was lyophilized to obtain the crude polysaccharides [48].
5.3. Fumonisin Analysis
Fumonisin was extracted from 20 mL of the liquid culture filtrate according to the method of Jian et al. [49]. The ABSCIEX triple quadTM 5500 UPLC–MS/MS system (AB SCIEX, Framingham, MA, USA) accompanied by a Ekspert 100 UPLC column (C18 column, 100 × 2.1 mm, 3 µm particle size, Thermo, USA) was used to analyze the FB1 and FB2. The FBs analyses were conducted according to the method described by Li et al. [37]. Briefly, 10 µL of sample was injected for fumonisin analysis. An optimized gradient of mobile phase (A: acetonitrile and B: 5 mM ammonium acetate) were applied as follows: the initial composition of the mobile phase 10% of A/90% of B was kept constant for 0.5 min, then the A solvent was linearly increased to 50% in 7.5 min. Finally, the A solvent was linearly decreased to 10% in 0.5 min and kept constant for 1 min. The flow rate of the mobile was 0.4 mL/min. Positive ionization was selected for mass spectrometric (MS) detection. A multiple reaction monitoring (MRM) function was employed for quantification, with the fragment ions at m/z 722.5 for FB1 and 706.4 for FB2, respectively. Three biological replicates were conducted.
5.4. RNA Isolation and cDNA Synthesis
For RNA isolation, the mycelia of F. proliferatum grown in the CB media with different carbon sources were filtered by Buchner funnel and then liquid nitrogen was added immediately and ground into powder. The powder (100 mg) was weighed and then used for RNA extraction. The RNA was extracted using the Hipure Fungal RNA Mini Kit (Magen, Guangzhou, China). The cDNA was synthesized using the cDNA PrimeScriptTM RT Master Mix Takara Kit (TAKARA-RR036A, Dalian, China).
5.5. Expression Analysis by Real-Time Quantitative PCR
ABI7500 fast real-time fluorescence quantitative PCR instrument (Applied Biosystems, Foster City, CA, USA) was used for RT-qPCR assay using our previous method [50]. SYBR Premix Ex TaqTM mix (TaKaRa, Dalian, China) was used in this study with 20 µL of reaction system, including 10.0 µL of SYBR Premix Ex TaqTM, 0.4µL of PCR forward primer (10 µM), 0.4 µL of PCR reverse primer (10 µM), 0.4 µL of ROX reference dyeⅡ and 2 µL (20 ng) of cDNA. After amplification (40 cycles at 95 °C for 30 s, 95 °C for 5 s and 60 °C for 34 s), the relative expression levels of target genes were calculated using the formula 2−ΔΔCT with Histone H3 as the reference gene. All these genes were selected due to their important roles in fumonisin production and their responses to different carbon sources [25]. The following prime pairs designed by PrimerPremier 5 (PREMIER Biosoft International, Palo Alto, CA, USA), were used for RT-qPCR, as shown in Table 1. Three biological replications were conducted.
5.6. Statistical Analysis
All experiments were performed in triplicate. Data for each sample were statistically analyzed using SPSS software (Version 16.0, SPSS Inc., Chicago, IL, USA). One-way analysis of variance (ANOVA) followed by the Duncan’s multiple comparison was used for statistical significance analysis. Differences were considered to be significant at p < 0.05.
Supplementary Materials
The following are available online at https://0-www-mdpi-com.brum.beds.ac.uk/2072-6651/11/5/289/s1. Figure S1: NMR spectra of unripe banana peel polysaccharides. (A) 1H spectra; (B) HSQC Spectra. Figure S2: NMR spectra of ripe banana peel polysaccharides. (A) 1H spectra; (B) HSQC Spectra.
Author Contributions
Y.J. and T.L. designed the experiments; Y.W., T.L., and Y.W. performed the experiments; L.G. and Y.W. helped data handling; Y.W. drafted the paper; Y.J. and T.L. revised the paper.
Funding
This research was supported by the National Natural Science Foundation of China (grant No. 31701657), Young Elite Scientists Sponsorship Program by China Association for Science and Technology (2017QNRC001) and Pearl River ST Nova Program of Guangzhou (No. 201906010005).
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Berthiller, F.; Crews, C.; Dall’Asta, C.; Saeger, S.D.; Haesaert, G.; Karlovsky, P.; Oswald, I.P.; Seefelder, W.; Speijers, G.; Stroka, J. Masked mycotoxins: A review. Mol. Nutr. Food Res. 2013, 57, 165–186. [Google Scholar] [CrossRef] [PubMed]
- Bennett, J.W.; Klich, M. Mycotoxins. Clin. Microbiol. Rev. 2003, 16, 497–516. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gorna, K.; Pawlowicz, I.; Waskiewicz, A.; Stepien, L. Fusarium proliferatum strains change fumonisin biosynthesis and accumulation when exposed to host plant extracts. Fungal Biol. 2016, 120, 884–893. [Google Scholar] [CrossRef] [PubMed]
- Rheeder, J.P.; Marasas, W.F.O.; Vismer, H.F. Production of fumonisin analogs by Fusarium species. Appl. Environ. Microbiol. 2002, 68, 2101–2105. [Google Scholar] [CrossRef] [PubMed]
- Li, R.; Tao, B.; Pang, M.; Liu, Y.; Dong, J. Natural occurrence of fumonisins B1 and B2 in maize from three main maize-producing provinces in China. Food Control 2015, 50, 838–842. [Google Scholar] [CrossRef]
- Hove, M.; Van Pouckea, C.; Njumbe-EdiageaL, E.; Nyanga, L.K.; De Saegera, S. Review on the natural co-occurrence of AFB1 and FB1 in maize and the combined toxicity of AFB1 and FB1. Food Control 2016, 59, 675–682. [Google Scholar] [CrossRef]
- Voss, K.A.; Smith, G.W.; Haschek, W.M. Fumonisins: Toxicokinetics, mechanism of action and toxicity. Anim. Feed Sci. Technol. 2007, 137, 299–325. [Google Scholar] [CrossRef]
- Abdallah, M.F.; Girgin, G.; Baydar, T. Occurrence, prevention and limitation of mycotoxins in feeds. Anim. Nutr. Feed Technol. 2015, 15, 471–490. [Google Scholar] [CrossRef]
- Liu, C.; Xu, W.; Liu, F.; Jiang, S. Fumonisins production by Fusarium proliferatum strains isolated from asparagus crown. Mycopathologia 2007, 164, 127–134. [Google Scholar] [CrossRef]
- Summerell, B.A.; Laurence, M.H.; Liew, E.C.Y.; Leslie, J.F. Biogeography and phylogeography of Fusarium: A review. Fungal Divers. 2010, 44, 3–13. [Google Scholar] [CrossRef]
- Li, J.; Jiang, G.; Yang, B.; Dong, X.; Feng, L.; Lin, S.; Chen, F.; Ashraf, M.; Jiang, Y. A luminescent bacterium assay of fusaric acid produced by Fusarium proliferatum from banana. Anal. Bioanal. Chem. 2012, 402, 1347–1354. [Google Scholar] [CrossRef]
- Arias, M.M.D.; Munkvold, G.P.; Ellis, M.L.; Leandro, L.F.S. Distribution and frequency of Fusarium species associated with soybean roots in Iowa. Plant Dis. 2013, 97, 1557–1562. [Google Scholar] [CrossRef] [PubMed]
- Chang, K.F.; Hwang, S.F.; Conner, R.L.; Ahmed, H.U.; Zhou, Q.; Turnbull, G.D.; Strelkov, S.E.; McLaren, D.L.; Gossen, B.D. First report of Fusarium proliferatum causing root rot in soybean (Glycine max L.) in Canada. Crop. Prot. 2015, 67, 52–58. [Google Scholar] [CrossRef]
- Busman, M.; Desjardins, A.E.; Proctor, R.H. Analysis of fumonisin contamination and the presence of Fusarium in wheat with kernel black point disease in the United States. Food Addit. Contam. 2012, 29, 1092–1100. [Google Scholar] [CrossRef] [PubMed]
- Fadl-Allah, E.; Stack, M.; Goth, R.; Bean, G. Production of fumonisins B1, B2 and B3 by Fusarium proliferatum isolated from rye grains. Mycotoxins Res. 1997, 13, 43–48. [Google Scholar] [CrossRef] [PubMed]
- Logrieco, A.; Moretti, A.; Castella, G.; Kostecki, M.; Golinski, P.; Ritieni, A.; Chelkowski, J. Beauvericin production by Fusarium species. Appl. Environ. Microbiol. 1998, 64, 3084–3088. [Google Scholar] [CrossRef]
- Han, Z.; Tangni, E.K.; Huybrechts, B.; Munaut, F.; Scauflaire, J.; Wu, A.; Callebaut, A. Screening survey of co-production of fusaric acid, fusarin C, and fumonisins B1, B2 and B3 by Fusarium strains grown in maize grains. Mycotoxin Res. 2014, 30, 231–240. [Google Scholar] [CrossRef]
- Kohut, G.; Olah, B.; Adam, A.L.; Garcia-Martinez, J.; Hornok, L. Adenylyl cyclase regulates heavy metal sensitivity, bikaverin production and plant tissue colonization in Fusarium proliferatum. J. Basic Microbiol. 2010, 50, 59–71. [Google Scholar] [CrossRef]
- Proctor, R.H.; Desjardins, A.E.; Mccormick, S.P.; Planttner, R.D. Genetic analysis of the role of trichothecene and fumonisin mycotoxins in the virulence of Fusarium. Eur. J. Plant Pathol. 2002, 108, 691–698. [Google Scholar] [CrossRef]
- Stepien, L.; Koczyk, G.; Waskiewicz, A. FUM cluster divergence in fumonisins-producing Fusarium species. Fungal Biol. 2011, 115, 112–123. [Google Scholar] [CrossRef]
- Rosler, S.M.; Sieber, C.M.; Humpf, H.U.; Tudzynski, B. Interplay between pathway-specific and global regulation of the fumonisin gene cluster in the rice pathogen Fusarium fujikuroi. Appl. Microbiol. Biotechnol. 2016, 100, 5869–5882. [Google Scholar] [CrossRef]
- Flaherty, J.E.; Woloshuk, C.P. Regulation of fumonisin biosynthesis in Fusarium verticillioides by a zinc binuclear cluster-type gene, ZFR1. Appl. Environ. Microbiol. 2004, 70, 2653–2659. [Google Scholar] [CrossRef]
- Bluhm, B.H.; Kim, H.; Butchko, R.A.E.; Woloshuk, C.P. Involvement of ZFR1 of Fusarium verticillioides in kernel colonization and the regulation of FST1, a putative sugar transporter gene required for fumonisin biosynthesis on maize kernels. Mol. Plant Pathol. 2008, 9, 203–211. [Google Scholar] [CrossRef]
- Sagaram, U.S.; Kolomiets, M.; Shim, W.B. Regulation of fumonisin biosynthesis in Fusarium verticillioides-maize system. Plant Pathol. J. 2006, 22, 203–210. [Google Scholar] [CrossRef]
- Li, T.; Gong, L.; Jiang, G.; Wang, Y.; Gupta, V.K.; Qu, H.; Duan, X.; Wang, J.; Jiang, Y. Carbon Sources Influence Fumonisin Production in Fusarium proliferatum. Proteomics 2017, 17, 33–37. [Google Scholar] [CrossRef]
- Stępien, L.; Waskiewicz, A.; Wilman, K. Host extract modulates metabolism and fumonisin biosynthesis by the plant-pathogenic fungus Fusarium proliferatum. Int. J. Food Microbiol. 2015, 193, 74–81. [Google Scholar] [CrossRef]
- Miyara, I.; Shafran, H.; Davidzon, M.; Sherman, A.; Prusky, D. pH regulation of ammonia secretion by Colletotrichum gloeosporioides and its effect on appressorium formation and pathogenicity. Mol. Plant-Microbe Interact. 2010, 23, 304–316. [Google Scholar] [CrossRef] [PubMed]
- Bi, F.; Barad, S.; Ment, D.; Luria, N.; Casado, V.; Galam, N.; Dubay, A. Carbon regulation of environmental pH by secreted small molecule effectors modulates pathogenicity in fungi. Mol. Plant Pathol. 2016, 17, 1178–1195. [Google Scholar] [CrossRef]
- Barad, S.; Horowitz, S.B.; Kobiler, I.; Sherman, A.; Prusky, D. Accumulation of the mycotoxin patulin in the presence of gluconic acid contributes to pathogenicity of penicillium expansum. Mol. Plant-Microbe Interact. 2014, 27, 66–77. [Google Scholar] [CrossRef]
- Prusky, D.; Bi, F.; Moral, J.; Barad, S. How does host carbon concentration modulate the lifestyle of postharvest pathogens during colonization? Front. Plant Sci. 2016, 7, 1306. [Google Scholar] [CrossRef]
- Cruz, A.; Marin, P.; Magan, N.; Gonzalez, M.T. Combined effects of benomyl and environmental factors on growth and expression of the fumonisin biosynthetic genes FUM1 and FUM19 by Fusarium verticillioides. Int. J. Food Microbiol. 2014, 191, 17–23. [Google Scholar] [CrossRef] [PubMed]
- Melcion, D.; Cahagnier, B.; Bakan, B.; Richard, D. Influence of temperature on fumonisin B1 production on maize grain by Fusarium proliferatum. Sci. Aliment. 1998, 18, 301–311. [Google Scholar]
- Kawakami, A.; Nakajima, T.; Hirayae, K. Effects of carbon sources and amines on induction of trichothecene production by Fusarium asiaticum in liquid culture. FEMS Microbiol. Lett. 2014, 352, 204–212. [Google Scholar] [CrossRef] [PubMed]
- Kohut, G.; Adam, A.L.; Fazekas, B.; Hornok, L. N-starvation stress induced FUM gene expression and fumonisin production is mediated via the HOG-type MAPK pathway in Fusarium proliferatum. Int. J. Food Microbiol. 2009, 130, 65–69. [Google Scholar] [CrossRef]
- Prusky, D.; Wilson, R. Does increased nutritional carbon availability in fruit and foliar hosts contribute to modulation of pathogen colonization? Postharvest Biol. Technol. 2018, 145, 27–32. [Google Scholar] [CrossRef]
- Prusky, D. Pathogen quiescence in postharvest diseases. Annu. Rev. Phytopathol. 1996, 34, 413–434. [Google Scholar] [CrossRef] [PubMed]
- Li, T.; Gong, L.; Wang, Y.; Chen, F.; Gupta, V.K.; Jian, Q.; Duan, X.; Jiang, Y. Proteomics analysis of Fusarium proliferatum under various initial pH during fumonisin production. J. Proteom. 2017, 164, 59–72. [Google Scholar] [CrossRef] [PubMed]
- Zong, Y.; Li, B.; Tian, S. Effects of carbon, nitrogen and ambient pH on patulin production and related gene expression in Penicillium expansum. Int. J. Food Microbiol. 2015, 206, 102–108. [Google Scholar] [CrossRef]
- Fanelli, F.; Schmidt, M.; Haidukowski, M.; Geisen, R.; Logrieco, A.; Mule, G. Influence of light on growth, fumonisin biosynthesis and FUM1 gene expression by Fusarium proliferatum. Int. J. Food Microbiol. 2012, 153, 148–153. [Google Scholar] [CrossRef]
- Rocha, L.O.; Reis, G.M.; Fontes, L.C.; Piacentini, K.C. Association between FUM expression and fumonisin contamination in maize from silking to harvest. Crop Prot. 2017, 94, 77–82. [Google Scholar] [CrossRef]
- Alexander, N.J.; Proctor, R.H.; McCormick, S.P. Genes, gene clusters, and biosynthesis of trichothecenes and fumonisins in Fusarium. Toxin Rev. 2009, 28, 198–215. [Google Scholar] [CrossRef]
- Proctor, R.H.; Brown, D.W.; Plattner, R.D.; Desjardins, A.E. Co-expression of 15 contiguous genes delineates a fumonisin biosynthetic gene cluster in Gibberella moniliformis. Fungal Genet. Biol. 2003, 38, 237–249. [Google Scholar] [CrossRef]
- Stepien, L. The use of Fusarium secondary metabolite biosynthetic genes in chemotypic and phylogenetic studies. Crit. Rev. Microbiol. 2014, 40, 176–185. [Google Scholar] [CrossRef]
- Visentin, I.; Montis, V.; Doll, K.; Alabouvette, C.; Tamietti, G.; Karlovsky, P.; Cardinale, F. Transcription of genes in the biosynthetic pathway for Fumonisin mycotoxins is epigenetically and differentially regulated in the fungal maize pathogen Fusarium verticillioides. Eukaryot. Cell 2012, 11, 252–259. [Google Scholar] [CrossRef]
- Jayashree, T.; Subramanyam, C. Oxidative stress as a prerequisite for aflatoxin production by Aspergillus parasiticus. Free Radic. Biol. Med. 2000, 29, 981–985. [Google Scholar] [CrossRef]
- Li, T.; Jian, Q.; Chen, F.; Wang, Y.; Gong, L.; Duan, X.; Yang, B.; Jiang, Y. Influence of butylated hydroxyanisole on the growth, hyphal morphology, and the biosynthesis of fumonisins in Fusarium proliferatum. Front. Microbiol. 2016, 7, 1038. [Google Scholar] [CrossRef]
- John, A.; Yang, J.; Liu, J.; Jiang, Y.; Yang, B. The structure changes of water-soluble polysaccharides in papaya during ripening. Int. J. Biol. Macromol. 2018, 115, 152–156. [Google Scholar] [CrossRef]
- Zhang, Z.; Wang, X.; Han, Z.; Zhao, M.; Yin, L. Purification, antioxidant and moisture-preserving activities of polysaccharides from papaya. Carbohydr. Polym. 2012, 87, 2332–2337. [Google Scholar] [CrossRef]
- Jian, Q.; Gong, L.; Li, T.; Wang, Y. Rapid Assessment of the toxicity of fungal compounds using luminescent Vibrio qinghaiensis sp. Q67. Toxins 2017, 9, 335. [Google Scholar] [CrossRef]
- Li, T.; Jiang, G.; Qu, H.; Wang, Y. Comparative transcriptome analysis of Penicillium citrinum cultured with different carbon sources identifies genes involved in citrinin biosynthesis. Toxins 2017, 9, 69. [Google Scholar] [CrossRef]
Figure 1.
The morphology of F. proliferatum on CB media with different carbon sources. A.B.C.

Figure 2.
Effects of different sucrose conditions on fugal growth (A, B) and FB1 production (C) of F. prolifeatum. Sucrose: 6 days in the presence of sucrose; lack sucrose: 6 days in the absence of sucrose; and supplementation of sucrose: 3 days in the absence of sucrose, followed by 3-day culture after the supplementation of sucrose. The vertical bars indicate standard errors of three replicates. Different letters represent significant differences (p < 0.05).
Figure 2.
Effects of different sucrose conditions on fugal growth (A, B) and FB1 production (C) of F. prolifeatum. Sucrose: 6 days in the presence of sucrose; lack sucrose: 6 days in the absence of sucrose; and supplementation of sucrose: 3 days in the absence of sucrose, followed by 3-day culture after the supplementation of sucrose. The vertical bars indicate standard errors of three replicates. Different letters represent significant differences (p < 0.05).

Figure 3.
The morphology of F. proliferatum on the CB media with different carbon sources.
Figure 4.
Effect of different sucrose conditions on fugal growth (A, B) and FB production (C, D) of F. prolifeatum. F. proliferatum were cultured in the culture media containing sucrose, cellulose, hemicellulose, and polysaccharide extracted from unripe or ripe banana peel for 6 days at 28 °C. The vertical bars indicate standard errors of three replicates. Different letters represent significant differences (p < 0.05).
Figure 4.
Effect of different sucrose conditions on fugal growth (A, B) and FB production (C, D) of F. prolifeatum. F. proliferatum were cultured in the culture media containing sucrose, cellulose, hemicellulose, and polysaccharide extracted from unripe or ripe banana peel for 6 days at 28 °C. The vertical bars indicate standard errors of three replicates. Different letters represent significant differences (p < 0.05).
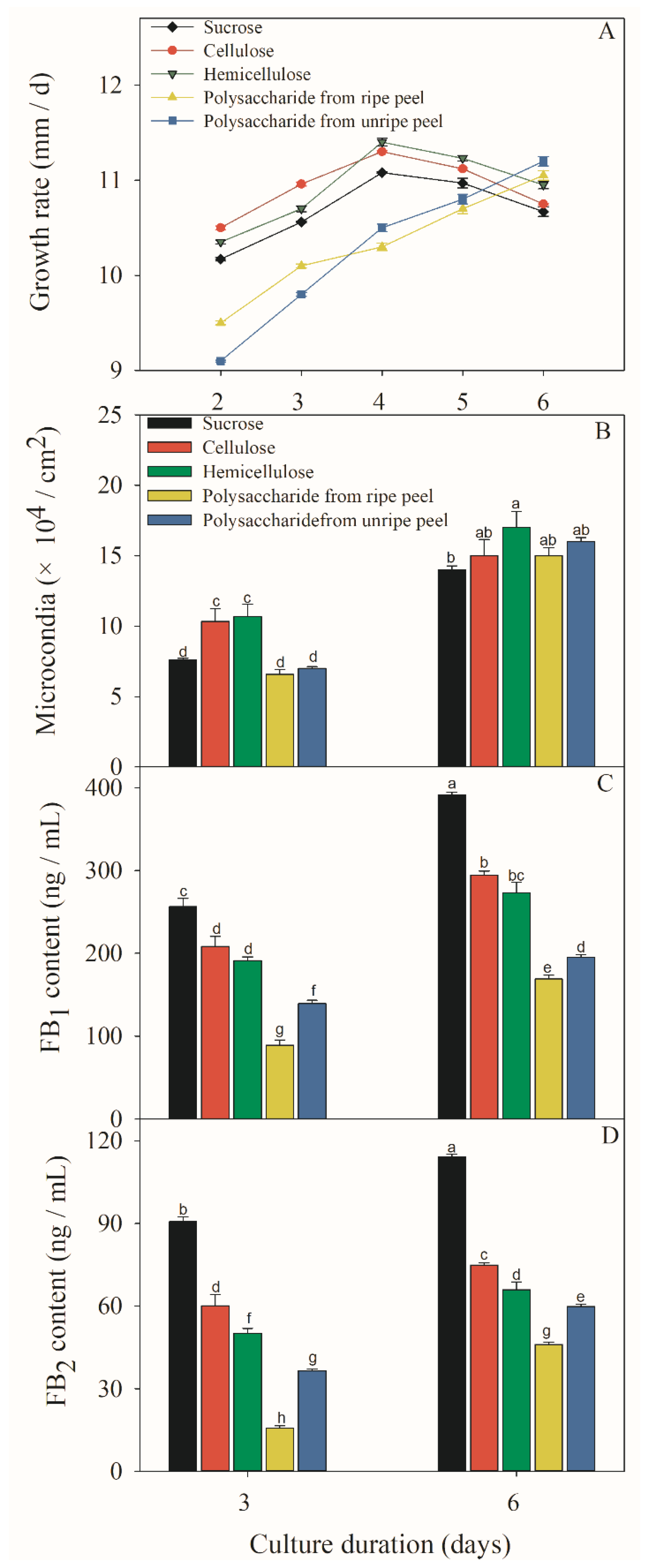
Figure 5.
Effect of sucrose, lack sucrose and supplementation of sucrose on the expressions of the fumonisin-related genes of F. proliferatum. The detailed information of these genes is shown in Table 1. The data are presented as means of three independent replicates. The vertical bars indicate standard errors of three replicates. Different letters represent significant differences (p < 0.05).
Figure 5.
Effect of sucrose, lack sucrose and supplementation of sucrose on the expressions of the fumonisin-related genes of F. proliferatum. The detailed information of these genes is shown in Table 1. The data are presented as means of three independent replicates. The vertical bars indicate standard errors of three replicates. Different letters represent significant differences (p < 0.05).

Figure 6.
Effect of sucrose, cellulose and hemicellulose on the expressions of the fumonisin-related genes of F. proliferatum. The detailed information of these genes is shown in Table 1. The data are presented as means of three independent replicates. The vertical bars indicate standard errors of three replicates. Different letters represent significant differences (p < 0.05).
Figure 6.
Effect of sucrose, cellulose and hemicellulose on the expressions of the fumonisin-related genes of F. proliferatum. The detailed information of these genes is shown in Table 1. The data are presented as means of three independent replicates. The vertical bars indicate standard errors of three replicates. Different letters represent significant differences (p < 0.05).
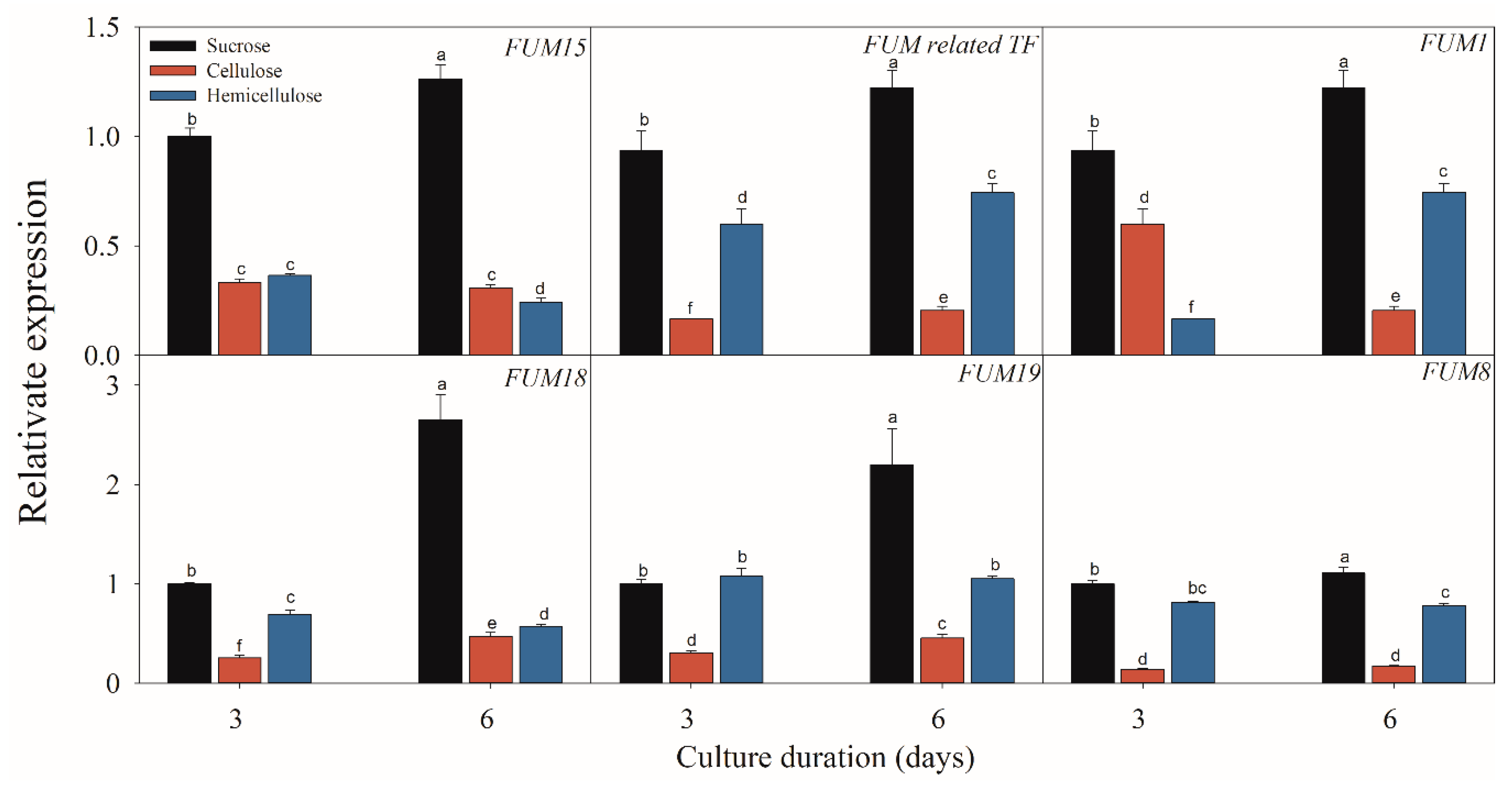
Figure 7.
Effect of sucrose and polysaccharide extracted from unripe or ripe banana peel on the expressions of the fumonisin-related genes of F. proliferatum. The detailed information of these genes is shown in Table 1. The data are presented as means of three independent replicates. The vertical bars indicate standard errors of three replicates. Different letters represent significant differences (p < 0.05).
Figure 7.
Effect of sucrose and polysaccharide extracted from unripe or ripe banana peel on the expressions of the fumonisin-related genes of F. proliferatum. The detailed information of these genes is shown in Table 1. The data are presented as means of three independent replicates. The vertical bars indicate standard errors of three replicates. Different letters represent significant differences (p < 0.05).

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Prime pairs used for RT-qPCR.
| Gene | Description | Sequence of Primer (5’ to 3’) |
|---|---|---|
| FUM1 | Polyketide synthase | For: ACTTTGCCATTTCCAACCGTAT |
| Rev: GGGAGTTTTTCCATCCGAATTT | ||
| FUM6 | Cytochrome P450 Monooxygenase | For: CGCTGGTACAGAAACGACGGCTAC |
| Rev: TCGCGTAGGCACGCACTGAGATA | ||
| FUM8 | Aminotransferase | For: ATTCCATGAGGAGGCAATGCAG |
| Rev: GGTGCTATTCCTTCGAGGTCAC | ||
| FUM9 | Dioxygenase | For: GAGCGTGGATGCTTGGCTGTTACT |
| Rev: GGACTGGGAGCTTCTTTGCGGTATC | ||
| FUM15 | Cytochrome P450 monooxygenase | For: CCATTCCACTCACGATGCGAGAAGC |
| Rev: GCCAGGATTATTCTAGTGCCAGCAGGTA | ||
| FUM18 | Longevity assurance factor | For: TGGTAGATGATGTGAGGAGCGACGA |
| Rev: TCAAGTAGCCGTTGCCGTCATTCC | ||
| FUM19 | ABC transporter | For: GGCTATGGATTCGGACGCTCTCAG |
| Rev: ACCGTGCTGTGCTTGACCTAACATC | ||
| FUM related TF | Transcription factor | For: GCGGTGGAGGTGTCGGATTGAGTAA |
| Rev: TGTCGGTGGAGGTAATGTAGTGGCTATTC | ||
| ZFR1 | ZFR1 regulator of fumonisin biosynthesis | For: GCTCGTCTTCTCCTACATCGGCATCA |
| Rev: CGGAATATGTGCGTTGTCAACAAGGTAGT | ||
| FUM related Zn(II)2Cys6 | Fumonisin biosynthetic Related Zn(II)2Cys6 protein | For: CAACTGCCAATAGCGAGGATGTGATGTC |
| Rev: GACCTTCTCAACAATCCCGATTCCATTAC | ||
| Histone H3 | Histone H3 | For: ACTAAGCAGACCGCCCGCAGG |
| Rev: GCGGGCGAGCTGGATGTCCTT |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Wu, Y.; Li, T.; Gong, L.; Wang, Y.; Jiang, Y. Effects of Different Carbon Sources on Fumonisin Production and FUM Gene Expression by Fusarium proliferatum. Toxins 2019, 11, 289. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins11050289
AMA Style
Wu Y, Li T, Gong L, Wang Y, Jiang Y. Effects of Different Carbon Sources on Fumonisin Production and FUM Gene Expression by Fusarium proliferatum. Toxins. 2019; 11(5):289. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins11050289
Chicago/Turabian StyleWu, Yu, Taotao Li, Liang Gong, Yong Wang, and Yueming Jiang. 2019. "Effects of Different Carbon Sources on Fumonisin Production and FUM Gene Expression by Fusarium proliferatum" Toxins 11, no. 5: 289. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins11050289
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.