Identification of the Fungal Pathogens of Postharvest Disease on Peach Fruits and the Control Mechanisms of Bacillus subtilis JK-14

Abstract
:1. Introduction
2. Results
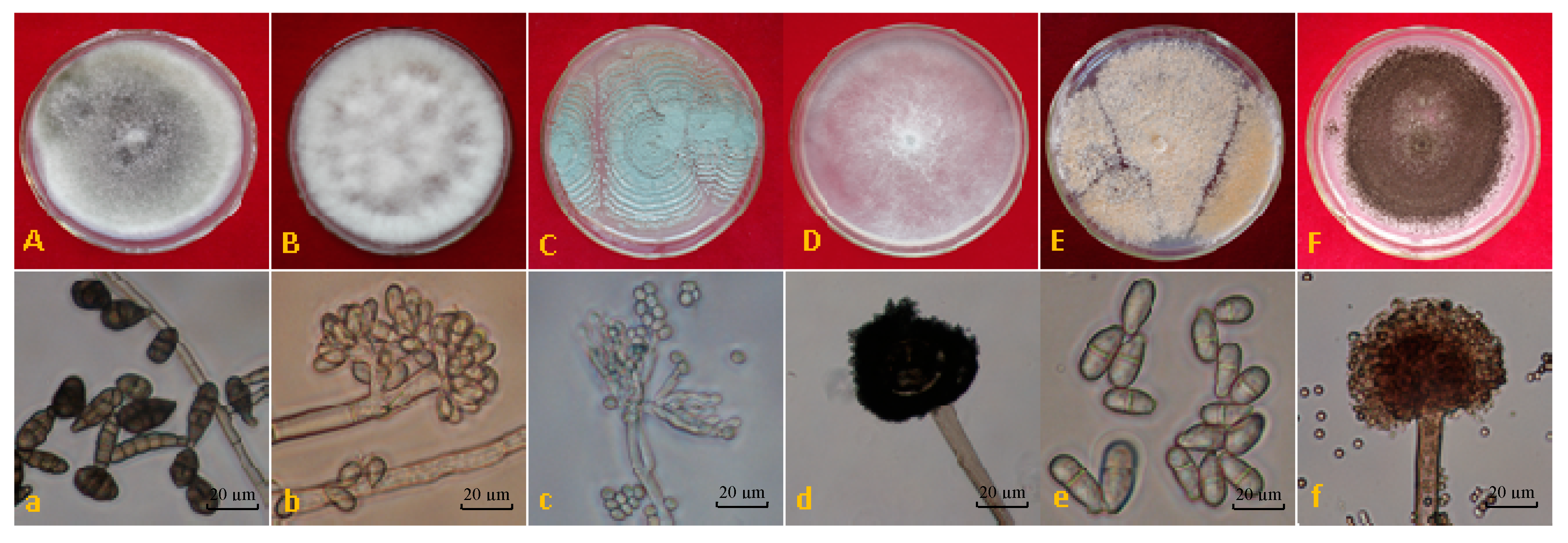
2.1. Isolation and Identification of Postharvest Fungal Pathogens
2.2. Determination of the Pathogenicity of the Isolates
2.3. Inhibitory Effect of Bacillus subtilis JK-14 against Alternaria tenuis and Botrytis cinerea
2.4. Effect of Bacillus subtilis JK-14 in Controlling Alternaria tenuis and Botrytis cinerea Decay on Peaches
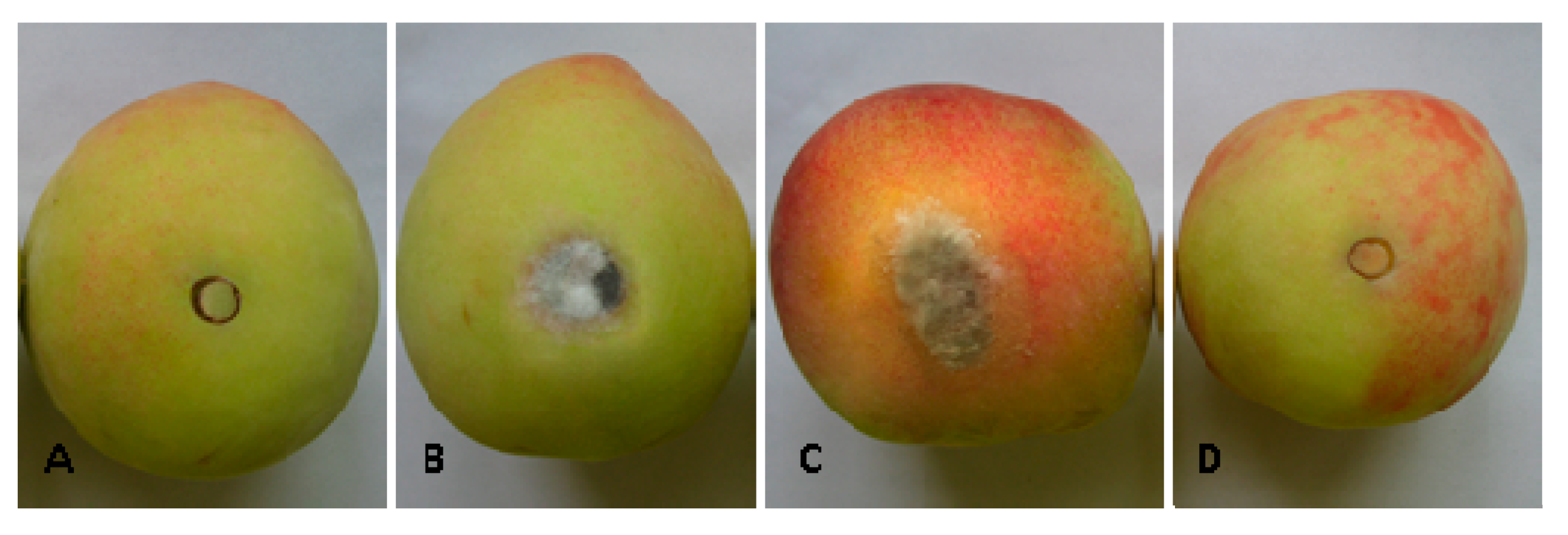
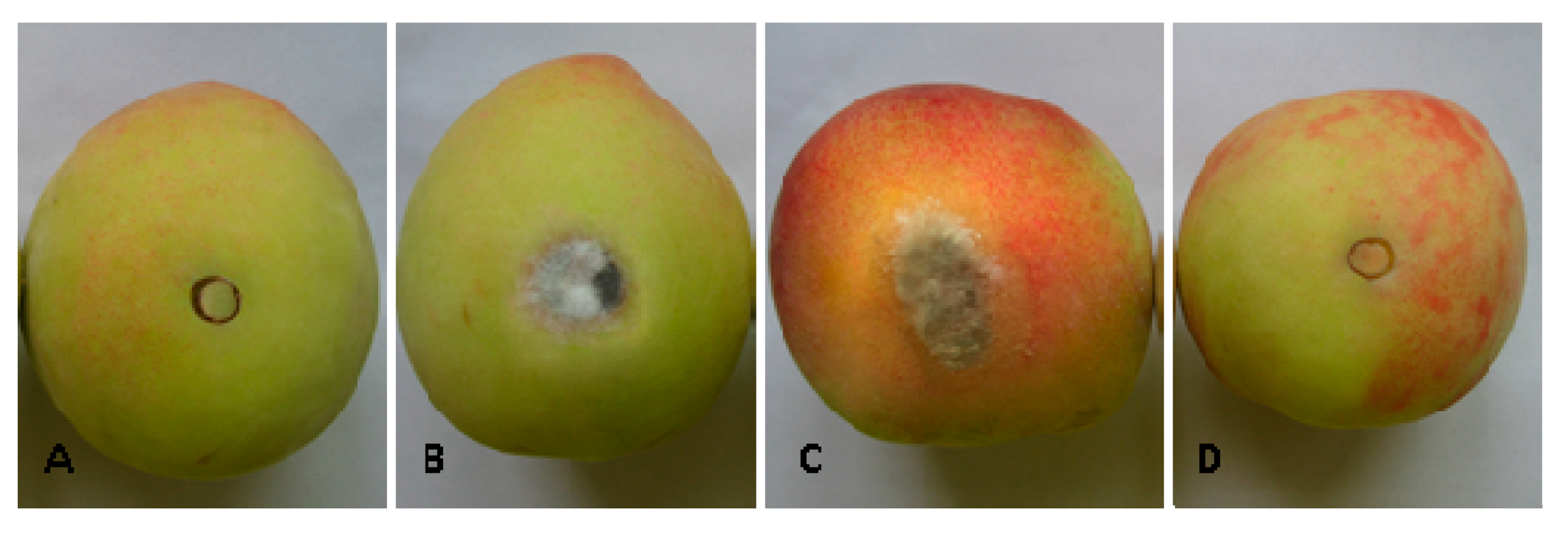
2.5. Effect of Bacillus subtilis JK-14 on the Symptoms of Fruits Decay after Inoculation with the Pathogens on Peaches
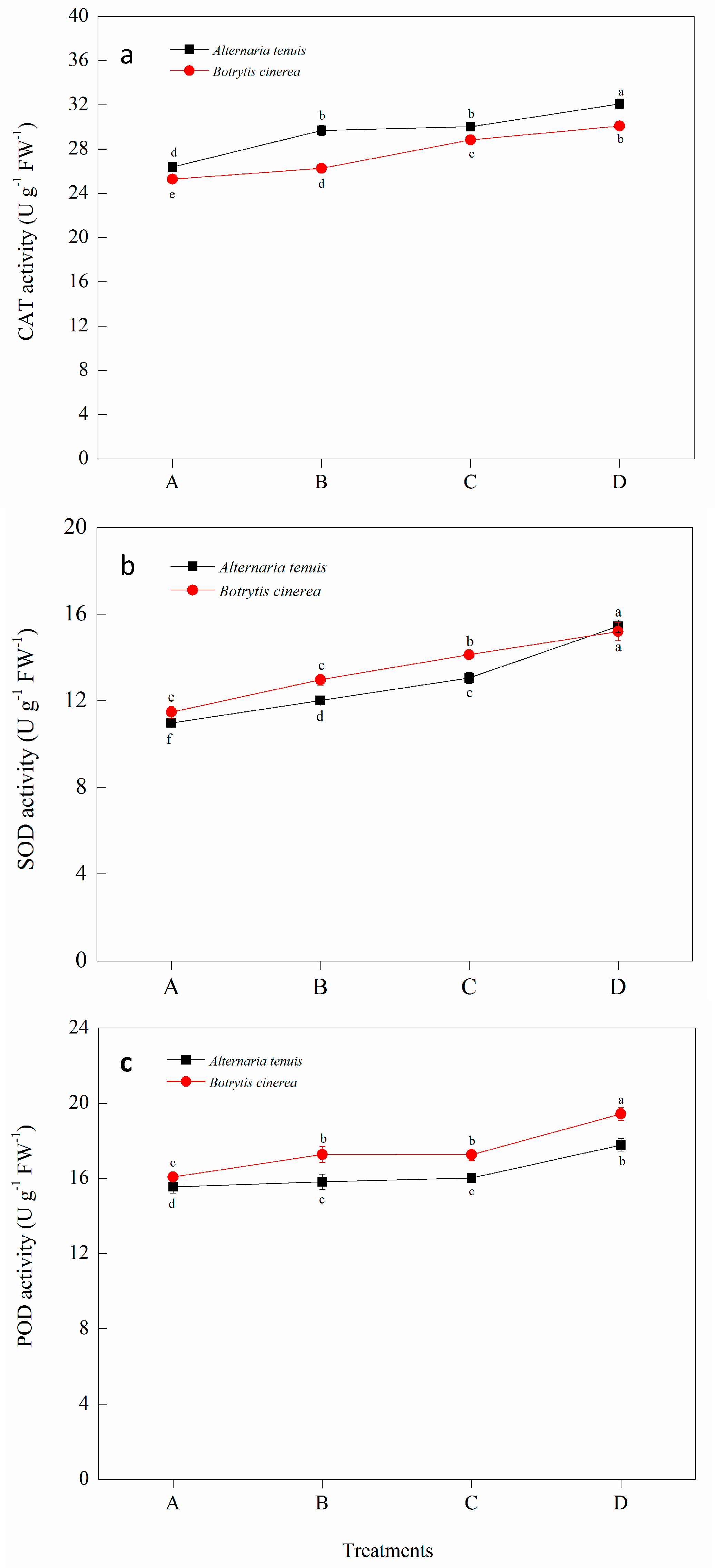
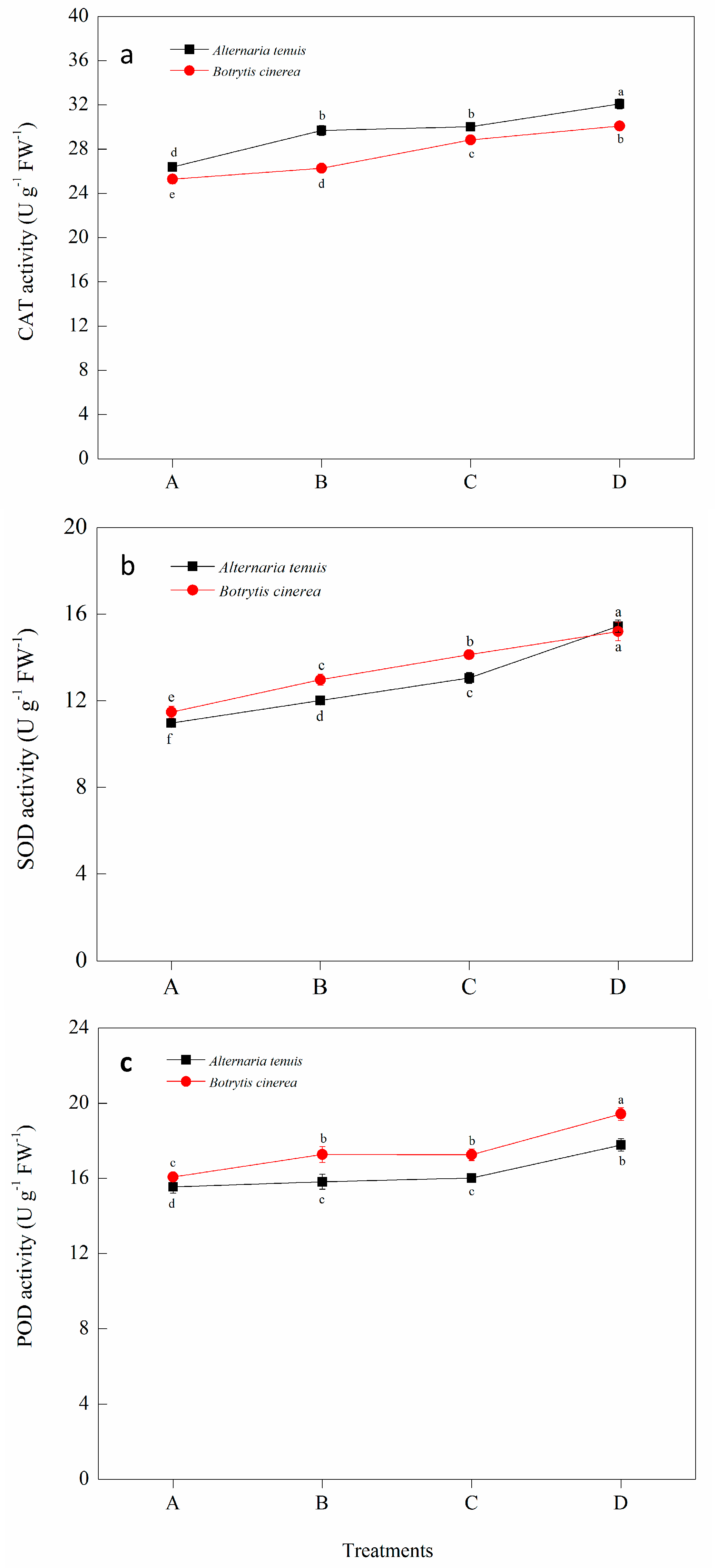
2.6. Effect of Bacillus subtilis JK-14 on the Activities of Defense-Related Enzymes of Peaches
3. Discussion and Conclusions
4. Materials and Methods
4.1. Fungal Pathogens Isolation and Identification
4.2. Spore Suspensions of Fungal Pathogen Preparation
4.3. Fruit Preparation
4.4. Pathogenicity of the Isolates on Peach Fruits
4.5. Formulations of Bacillus Subtilis JK-14 Preparation
4.6. In Vitro and in Vivo Antagonistic Activity Determination
4.7. Efficacy of Bacillus subtilis JK-14 in Controlling of Peach Postharvest Disease
4.8. Effects of Bacillus subtilis JK-14 on the Activities of Defense-Related Enzymes of Peaches
4.9. Determination and Analysis of Defense-Related Enzyme Activities of Peaches
4.10. Statistical Analysis
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Kader, A.A. Increasing food availability by reducing postharvest losses of fresh produce. Int. Postharvest Symp. Acta Hort. 2005, 682, 2169–2178. [Google Scholar] [CrossRef]
- Parfitt, J.; Barthel, M.; Macnaughton, S. Food waste within food supply chains: Quantification and potential for change to 2050. Philos. Trans. R. Soc. 2010, 365, 3065–3081. [Google Scholar] [CrossRef] [PubMed]
- Food and Agriculture Organization of the United Nations. Global Food Losses and Food Waste: Extent, Causes and Prevention; FAO: Rome, Italy, 2011. [Google Scholar]
- Liu, H.X.; Jiang, W.B.; Bi, Y.; Luo, Y.B. Postharvest BTH treatment induces resistance of peach (Prunus persica L. cv. Jiubao) fruit to infection by Penicillium expansum and enhances activity of fruit defense mechanisms. Postharvest Biol. Technol. 2005, 35, 263–269. [Google Scholar] [CrossRef]
- Karabulut, O.A.; Baykal, N. Integrated control of postharvest diseases of peaches with a yeast antagonist, hot water and modified atmosphere packaging. Crop Prot. 2004, 23, 431–435. [Google Scholar] [CrossRef]
- Gatto, M.A.; Ippolito, A.; Linsalata, V.; Cascarano, N.A.; Nigro, F.; Vanadia, S.; Di Venere, D. Activity of extracts from wild edible herbs against postharvest fungal diseases of fruit and vegetables. Postharvest Biol. Technol. 2011, 61, 72–82. [Google Scholar] [CrossRef]
- Xu, B.T.; Zhang, H.Y.; Chen, K.P.; Xu, Q.; Yao, Y.; Gao, H. Biocontrol of postharvest Rhizopus decay of peaches with Pichia caribbica. Curr. Microbiol. 2013, 67, 255–261. [Google Scholar] [CrossRef] [PubMed]
- Karabulut, O.A.; Cohen, L.; Wiess, B.; Daus, A.; Lurie, S.; Droby, S. Control of brown rot and blue mold of peach and nectarine by short hot water brushing and yeast antagonists. Postharvest Biol. Technol. 2002, 24, 103–111. [Google Scholar] [CrossRef]
- Vitoratos, A.; Dimitrios, B.; Karkanis, A.; Efthimiadou, A. Antifungal activity of plant essential oils against Botrytis cinerea, Penicillium italicum and Penicillium digitatum. Not. Bot. Hort. Agrobot. 2013, 41, 86–92. [Google Scholar] [CrossRef]
- Spadaro, D.; Droby, S. Development of biocontrol products for postharvest diseases of fruit: The importance of elucidating the mechanisms of action of yeast antagonists. Trends Food Sci. Tech. 2016, 47, 39–49. [Google Scholar] [CrossRef]
- Arrebola, E.; Sivakumar, D.; Bacigalupo, R.; Korsten, L. Combined application of antagonist Bacillus amyloliquefaciens and essential oils for the control of peach postharvest diseases. Crop Prot. 2010, 29, 369–377. [Google Scholar] [CrossRef]
- Mari, M.; Bautista-Banos, S.; Sivakumar, D. Decay control in the postharvest system: Role of microbial and plant volatile organic compounds. Postharvest Biol. Technol. 2016, 122, 70–81. [Google Scholar] [CrossRef]
- Usall, J.; Casals, C.; Sisquella, M.; Palou, L.; de Cal, A. Alternative technologies to control postharvest diseases of stone fruits. Stewart Postharvest Rev. 2015, 11, 1–6. [Google Scholar]
- Usall, J.; Torres, R.; Vias, I.; Abadias, M.; Teixidó, N. Principales enfermedades de postcosecha y su control. In Poscosecha de pera, manzana y melocotón; Mundiprensa: Madrid, Spain, 2013; pp. 247–280. [Google Scholar]
- Sharma, R.; Singh, D.; Singh, R. Biological control of postharvest diseases of fruits and vegetables by microbial antagonists: A review. Biol. Control 2009, 50, 205–221. [Google Scholar] [CrossRef]
- Wisniewski, M.; Droby, S.; Norelli, J.; Liu, J.; Schena, L. Alternative management technologies for postharvest disease control: The journey from simplicity to complexity. Postharvest Biol. Technol. 2016, 122, 3–10. [Google Scholar] [CrossRef]
- Holmes, G.J.; Eckert, J.W. Sensitivity of Penicillium digitatum and P. italicum to postharvest citrus fungicides in California. Phytopathology 1999, 89, 716–721. [Google Scholar] [CrossRef] [PubMed]
- Sansone, G.; Rezza, I.; Calvente, V.; Benuzzi, D.; Tosetti, M.I.S. Control of Botrytis cinerea strains resistant to iprodione in apple with rhodotorulic acid and yeasts. Postharvest Biol. Technol. 2005, 35, 245–251. [Google Scholar] [CrossRef]
- Adaskaveg, J.E.; Michailides, T.J. Cancellation of postharvest use of Rovral 50WP on stone fruit crops and other label changes of the fungicide. Cent. Vall. Postharvest Newslett. 1996, 5, 9–10. [Google Scholar]
- He, D.; Zheng, X.D.; Yin, Y.M.; Sun, P.; Zhang, H.Y. Yeast application for controlling apple postharvest diseases associated with Penicillium expansum. Bot. Bull Acad. Sinica 2003, 44, 211–216. [Google Scholar]
- Hong, C.X.; Michailides, T.J.; Holtz, B.A. Effects of wounding, inoculum density and biological control agents on postharvest brown rot of stone fruits. Plant Dis. 1998, 82, 1210–1216. [Google Scholar] [CrossRef]
- Lopez-Reyes, J.G.; Spadaro, D.; Prelle, A.; Garibaldi, A.; Gullino, M.L. Efficacy of plant essential oils on postharvest control of rots caused by fungi on different stone fruits in vivo. J. Food Prot. 2013, 76, 631–639. [Google Scholar] [CrossRef]
- Droby, S.; Wisniewski, M.; Macarisin, D.; Wilson, C. Twenty years of postharvest biocontrol research: Is it time for a new paradigm. Postharvest Biol. Technol. 2009, 52, 137–145. [Google Scholar] [CrossRef]
- Romanazzi, G.; Lichter, A.; Mlikota Gabler, F.; Smilanick, J.L. Recent advances on the use of natural and safe alternatives to conventional methods to control postharvest gray mold of table grapes. Postharvest Biol. Technol. 2012, 63, 141–147. [Google Scholar] [CrossRef]
- Janisiewicz, W.J.; Korsten, L. Biological control of postharvest diseases of fruits. Annu. Rev. Phytopathol. 2002, 40, 411–441. [Google Scholar] [CrossRef] [PubMed]
- Cavaglieri, L.; Orlando, J.; Rodriguez, M.I.; Chulze, S.; Etcheverry, M. Biocontrol of Bacillus subtilis against Fusarium verticillioides in vitro and at the maize root level. Res. Microbiol. 2005, 156, 748–754. [Google Scholar] [CrossRef]
- De Jensen, C.D.; Percich, J.A.; Graham, P.H. Integrated management strategies of bean root rot with Bacillus subtilis and Rhizobium in Minnesota. Field Crop Res. 2002, 74, 107–115. [Google Scholar] [CrossRef]
- Sharma, N.; Sharma, S. Control of foliar diseases of mustard by Bacillus from reclaimed soil. Microbiol. Res. 2008, 163, 408–413. [Google Scholar] [CrossRef] [PubMed]
- Scherm, H.; Ngugi, H.K.; Savelle, A.T.; Edwards, J.R. Biological control of infection of blueberry flowers caused by Monilinia vaccinii-corymbosi. Biol. Control 2004, 29, 199–206. [Google Scholar] [CrossRef]
- Acea, M.J.; Moore, C.R.; Alexander, M. Survival and growth of bacteria introduced into soil. Soil Biol. Biochem. 1988, 20, 509–515. [Google Scholar] [CrossRef]
- Lurie, S.; Crisosto, C.H. Chilling injury in peach and nectarine. Postharvest Biol. Technol. 2005, 37, 195–208. [Google Scholar] [CrossRef]
- Vreeland, R.H.; Rosenzweig, W.D.; Powers, D.W. Isolation of a 250 million-year-old halo tolerant bacterium from a primary salt crystal. Nature 2000, 407, 897–900. [Google Scholar] [CrossRef]
- Lee, S.C.; Yoo, J.S.; Kim, S.H.; Chung, S.Y.; Hwang, C.W.; Joo, C.H.; Choi, Y.L. Production and characterization of lipopeptide biosurfactant from Bacillus subtilis A8-8. J. Microbiol. Biotechnol. 2006, 16, 716–723. [Google Scholar]
- Snowdon, A.L. Colour Atlas of Postharvest-Harvest Diseases and Disorders of Fruit and Vegetables, Volume 1: General Introduction and Fruits; Manson Publishing Ltd.: London, UK, 2010. [Google Scholar]
- Hong, C.X.; Michailides, T.J. Prune, plum, and nectarine as hosts of Trichothecium roseum in California orchards. Plant Dis. 1997, 81, 112. [Google Scholar] [CrossRef] [PubMed]
- Mclaughlin, R.J.; Wilson, C.L.; Droby, S.; Benarie, R.; Chalutz, E. Biological control of postharvest diseases of grape, peach and apple with the yeasts kloeckera-apiculata and candida-guilliermondii. Plant Dis. 1992, 76, 470–473. [Google Scholar] [CrossRef]
- De Cal, A.; Sandín-Espana, P.; Martinez, F.; Egüen, B.; Chien-Ming, C.; Lee, M.H.; Melgarejo, P.; Prusky, D. Role of gluconic acid and pH modulation in virulence of Monilinia fructicola on peach fruit. Postharvest Biol. Technol. 2013, 86, 418–423. [Google Scholar] [CrossRef]
- Gell, I.; De Cal, A.; Torres, R.; Usall, J.; Melgarejo, P. Relationship between the incidence of latent infections caused by Monilinia spp. and the incidence of brown rot of peach fruit: Factors affecting latent infection. Eur. J. Plant Pathol. 2008, 121, 487–498. [Google Scholar] [CrossRef]
- Fan, Q.; Tian, S.P. Postharvest biological control of Rhizopus rot of nectarine fruits by Pichia membranefaciens. Plant Dis. 2000, 84, 1212–1216. [Google Scholar]
- Zhang, H.Y.; Wang, L.; Dong, Y.; Jiang, S. Postharvest biological control of gray mold decay of strawberry with Rhodotorula glutinis. Biol. Control 2007, 40, 287–292. [Google Scholar] [CrossRef]
- Zhang, H.Y.; Yang, Q.Y.; Ge, L.L.; Zhang, G.C.; Zhang, X.L.; Zhang, X.Y. Chitin enhances biocontrol of Rhodotorula mucilaginosa to postharvest decay of peaches. Int. J. Biol. Macromol. 2016, 88, 465–475. [Google Scholar] [CrossRef]
- Noble, R.; Coventry, E. Suppression of soil-borne plant diseases with composts: A review. Biocontrol Sci. Technol. 2005, 15, 3–20. [Google Scholar] [CrossRef]
- Jacobsen, A.M.; Hallingsørensen, B.; Ingerslev, F.; Hansen, S.H. Simultaneous extraction of tetracycline, macrolide and sulfonamide antibiotics from agricultural soils using pressurised liquid extraction, followed by solid-phase extraction and liquid chromatography-tandem mass spectrometry. J. Chromatogr. A 2004, 1038, 157–170. [Google Scholar] [CrossRef]
- Zhou, T.; Schneider, K.E.; Li, X.Z. Development of biocontrol agents from food microbial isolates for controlling post-harvest peach brown rot caused by Monilinia fructicola. Int. J. Food Microbiol. 2008, 126, 180–185. [Google Scholar] [CrossRef] [PubMed]
- Abdallah, B.; Stedel, R.A.; Garagounis, C.; Nefzi, C.; Jabnoun-Khiareddine, A.; Papadopoulou, H.; Daami-Remadi, K.K. Involvement of lipopeptide antibiotics and chitinase genes and induction of host defense in suppression of Fusarium wilt by endophytic Bacillus spp. in tomato. Crop Prot. 2017, 99, 45–58. [Google Scholar] [CrossRef]
- Fu, L.; Penton, C.R.; Ruan, Y. Inducing the rhizosphere microbiome by biofertilizer application to suppress banana Fusarium wilt disease. Soil Biol. Biochem. 2017, 104, 39–48. [Google Scholar] [CrossRef]
- Wu, B.; Wang, X.; Yang, L. Effects of Bacillus amyloliquefaciens ZM9 on bacterial wilt and rhizosphere microbial communities of tobacco. Appl. Soil Ecol. 2016, 103, 1–12. [Google Scholar] [CrossRef]
- Zheng, X.L.; Tian, S.P.; Meng, X.H.; Li, B.Q. Physiological and biochemical responses in peach fruit to oxalic acid treatment during storage at room temperature. Food Chem. 2007, 45, 281–284. [Google Scholar] [CrossRef]
- Zhang, S.W.; Gan, Y.T.; Xu, B.L.; Xue, Y.Y. The parasitic and lethal effects of Trichoderma longibrachiatum against Heterodera avenae. Biol. Control 2014, 72, 1–8. [Google Scholar] [CrossRef]
- Liu, J.; Tian, S.P.; Meng, X.H.; Xu, Y. Control effects of chitosan on postharvest diseases and physiological response of tomato fruit. Postharvest Biol. Tech. 2007, 44, 300–306. [Google Scholar] [CrossRef]
- Bell, D.K.; Wells, H.D.; Markham, C.R. In vitro antagonism of Trichoderma species against six fungal plant pathogens. Phytopathology 1982, 72, 379–382. [Google Scholar] [CrossRef]
- Liu, B.; Zhou, J. Effect study on isolation and screening of antagonistic actinomycetes in soil. Mod. Agric. Technol. 2013, 17, 225–228. [Google Scholar]
- Dasari, V.; Nikku, M.Y.; Donthireddy, S.R.R. Screening of antagonistic marine actinomycetes: Optimization of process parameters for the production of novel antibiotic by Amycolatopsis alba var. nov. DVR D4. J. Antimicrob. Chemother. 2011, 3, 92–98. [Google Scholar]
- Meng, X.H.; Li, B.Q.; Liu, J.; Tian, S.P. Physiological responses and quality attributes of table grape fruit to chitosan postharvest spray and postharvest coating during storage. Food Chem. 2008, 106, 501–508. [Google Scholar] [CrossRef]
- Giannopolitis, C.N.; Ries, S.K. Superoxide dismutases: I. occurrence in higher plants. Plant Physiol. 1977, 59, 309–314. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.S.; Tian, S.P.; Xu, Y.; Qin, G.Z.; Yao, H.J. Changes in the activities of protein and antioxidant enzymes in peach fruit inoculated with Cryptococcus laurentii or Penicillium expansum at 0 or 20 °C. Postharvest Biol. Tech. 2004, 34, 21–28. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Isolates | Disease Incidences (%) | |
|---|---|---|
| Wound Inoculation | Intact Inoculation | |
| Alternaria tenuis | 100.00 ± 0.00 a | 100.00 ± 0.00 a |
| Botrytis cinerea | 100.00 ± 0.00 a | 92.33 ± 3.16 c |
| Penicillium digitatum | 95.46 ± 4.21 b | 56.67 ± 3.02 f |
| Trichothecium roseum | 34.54 ± 2.56 g | 0.00 ± 0.00 i |
| Rhizopus nigricans | 100.00 ± 0.00 a | 83.33 ± 2.89 d |
| Aspergillus niger | 67.63 ± 2.33 e | 17.34 ± 1.54 h |
| Control | 0.00 ± 0.00 i | 0.00 ± 0.00 i |
| Treatments | Inhibitory Rates (%) | |
|---|---|---|
| Alternaria tenuis | Botrytis cinerea | |
| Bacillus subtilis JK-14 | 81.32 ± 2.11 b | 83.45 ± 1.54 a |
| Control | - | - |
| Treatments | Alternaria tenuis | Botrytis cinerea | ||
|---|---|---|---|---|
| Disease Incidences (%) | Lesion Diameters (mm) | Disease Incidences (%) | Lesion Diameters (mm) | |
| Bacillus subtilis JK-14 | 14.8 ± 3.40 b | 3.0 ± 0.06 d | 14.1 ± 3.40 b | 3.2 ± 0.16 d |
| Control | 94.8 ± 1.29 a | 12.0 ± 0.12 c | 92.6 ± 2.57 a | 13.1 ± 0.17 bc |
| Concentrations (CFU mL−1) | Disease Incidences (%) | Lesion Diameters (mm) | ||
|---|---|---|---|---|
| FLBC | BCS | FLBC | BCS | |
| 1 × 109 | 47.41 ± 1.28 c | 45.18 ± 3.40 cd | 6.13 ± 0.13 c | 5.69 ± 0.08 d |
| 1 × 108 | 38.52 ± 3.39 d | 36.30 ± 3.40 de | 5.61 ± 0.06 d | 5.16 ± 0.16 e |
| 1 × 107 | 18.52 ± 2.56 f | 14.82 ± 3.40 f | 3.79 ± 0.08 fg | 3.06 ± 0.06 g |
| 1 × 106 | 34.07 ± 3.40 e | 31.11 ± 2.22 e | 4.95 ± 0.06 e | 4.11 ± 0.09 f |
| 1 × 105 | 65.92 ± 2.60 b | 61.48 ± 3.39 b | 7.36 ± 0.14 b | 6.72 ± 0.19 bc |
| Control | 92.59 ± 1.28 a | 11.95 ± 0.09 a | ||
| Concentrations (CFU mL−1) | Disease Incidences (%) | Lesion Diameters (mm) | ||
|---|---|---|---|---|
| FLBC | BCS | FLBC | BCS | |
| 1 × 109 | 44.44 ± 2.23 c | 43.70 ± 3.40 c | 6.03 ± 0.19 c | 5.74 ± 0.22 c |
| 1 × 108 | 39.26 ± 1.28 d | 36.30 ± 1.28 e | 5.37 ± 0.25 d | 5.13 ± 0.15 d |
| 1 × 107 | 17.78 ± 2.22 h | 14.07 ± 3.40 i | 3.74 ± 0.16 f | 3.19 ± 0.16 g |
| 1 × 106 | 28.15 ± 1.28 f | 24.44 ± 2.23 g | 4.80 ± 0.17 e | 4.22 ± 0.16 e |
| 1 × 105 | 62.22 ± 2.22 b | 60.00 ± 2.22 b | 7.15 ± 0.08 b | 6.66 ± 0.34 b |
| Control | 92.59 ± 2.57 a | 13.11 ± 0.17 a | ||
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, S.; Zheng, Q.; Xu, B.; Liu, J. Identification of the Fungal Pathogens of Postharvest Disease on Peach Fruits and the Control Mechanisms of Bacillus subtilis JK-14. Toxins 2019, 11, 322. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins11060322
Zhang S, Zheng Q, Xu B, Liu J. Identification of the Fungal Pathogens of Postharvest Disease on Peach Fruits and the Control Mechanisms of Bacillus subtilis JK-14. Toxins. 2019; 11(6):322. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins11060322
Chicago/Turabian StyleZhang, Shuwu, Qi Zheng, Bingliang Xu, and Jia Liu. 2019. "Identification of the Fungal Pathogens of Postharvest Disease on Peach Fruits and the Control Mechanisms of Bacillus subtilis JK-14" Toxins 11, no. 6: 322. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins11060322