An Eye on Staphylococcus aureus Toxins: Roles in Ocular Damage and Inflammation
by
 , and
, and
Roger Astley
1 ,
,
Frederick C. Miller
2,
Md Huzzatul Mursalin
3,
Phillip S. Coburn
1 and
and
Michelle C. Callegan
1,3,4,* 1
Department of Ophthalmology, University of Oklahoma Health Sciences Center, Oklahoma City, OK 73104, USA
2
Department of Cell Biology and Department of Family and Preventive Medicine, University of Oklahoma Health Sciences Center, Oklahoma City, OK 73104, USA
3
Department of Microbiology and Immunology, University of Oklahoma Health Sciences Center, Oklahoma City, OK 73104, USA
4
Dean McGee Eye Institute, 608 Stanton L. Young Blvd., DMEI PA-418, Oklahoma City, OK 73104, USA
*
Author to whom correspondence should be addressed.
Toxins 2019, 11(6), 356; https://0-doi-org.brum.beds.ac.uk/10.3390/toxins11060356
Submission received: 13 May 2019
/
Revised: 13 June 2019
/
Accepted: 15 June 2019
/
Published: 19 June 2019
(This article belongs to the Special Issue Staphylococcus aureus Toxins)
Abstract
:Staphylococcus aureus (S. aureus) is a common pathogen of the eye, capable of infecting external tissues such as the tear duct, conjunctiva, and the cornea, as well the inner and more delicate anterior and posterior chambers. S. aureus produces numerous toxins and enzymes capable of causing profound damage to tissues and organs, as well as modulating the immune response to these infections. Unfortunately, in the context of ocular infections, this can mean blindness for the patient. The role of α-toxin in corneal infection (keratitis) and infection of the interior of the eye (endophthalmitis) has been well established by comparing virulence in animal models and α-toxin-deficient isogenic mutants with their wild-type parental strains. The importance of other toxins, such as β-toxin, γ-toxin, and Panton–Valentine leukocidin (PVL), have been analyzed to a lesser degree and their roles in eye infections are less clear. Other toxins such as the phenol-soluble modulins have yet to be examined in any animal models for their contributions to virulence in eye infections. This review discusses the state of current knowledge of the roles of S. aureus toxins in eye infections and the controversies existing as a result of the use of different infection models. The strengths and limitations of these ocular infection models are discussed, as well as the need for physiological relevance in the study of staphylococcal toxins in these models.
Key Contribution: This review highlights the current knowledge base regarding toxins of Staphylococcus aureus and their roles in potentially blinding ocular infections. Although specific toxins have been identified as contributing to ocular tissue damage and inflammation, the exact mechanisms of action of these toxins on ocular tissues and their roles in inciting damaging inflammation need clarification.
1. Introduction
Staphylococcal involvement in wound infections was established in the 1881 report by Ogston [1], who described the formation of new abscesses in guinea pigs and mice injected with pus taken from his patients. He called the clusters of bacteria he observed in the abscesses “staphyle” (“bunch of grapes” in Greek) because the arrangement of the bacterial cells resembled a cluster of grapes [2]. In 1884, Rosenbach [3] published studies on bacteria isolated from human wounds. He isolated and categorized two staphylococci by color: golden colonies were named Staphylococcus aureus (S. aureus), and white colonies named Staphylococcus albus (now called epidermidis).
In 1928, S. aureus became infamous after the organism was identified as the cause of 18 children becoming violently ill after receiving diphtheria vaccinations in Bundaberg, Australia. Hours after inoculation, the children began to vomit, developed high fever, experienced convulsions, and fell into unconsciousness. Twelve of the children died within 36 h of vaccination. All of the surviving children developed severe abscesses at the injection site [4]. S. aureus was isolated from the vaccine vials [5]. The Commonwealth of Australia commissioned Burnet to study why the contamination had been so deadly [5]. Burnet demonstrated that supernatants from the S. aureus culture isolated from the vaccine caused hemolysis in vitro, caused skin-necrosis after intradermal inoculation, and was rapidly lethal to rabbits after intravenous injection. He concluded that a single heat-labile factor was responsible for these diverse pathological features [6]. Burnet further reported that other S. aureus isolates had the same pathological properties as the one isolated from the Bundaberg vaccine [7].
Burnet’s contention that the diverse disease features were due to a single toxin was disputed by various other researchers [8,9,10]. Glenny and Stevens [11] demonstrated that the hemolysis in vitro, skin-necrosis at the site of inoculation, and rapid lethality after intravenous injection were due to one immunologically distinct toxin, which they called “α-toxin”. Whether one toxin or more were responsible for these pathological features was not resolved until researchers were able to purify α-toxin [12,13] and demonstrate that α-toxin alone caused these pathological effects. Once purified toxin was available, researchers were able to study pore formation, identify α-toxin binding receptors, and examine how this single, potent toxin is able to produce such a diverse repertoire of actions.
S. aureus is a Gram-positive, nonmotile coccoid bacteria. S. aureus can be distinguished from other staphylococci by its classic gold pigmentation, β-hemolysis, a positive coagulase reaction, fermentation of mannitol, and a positive deoxyribonuclease test [14] (Figure 1). Although there are over 50 species of staphylococci, S. aureus is by far the most significant pathogen in this group [14]. S. aureus has an extensive array of virulence factors in its arsenal, which are depicted in Figure 2A. These virulence factors include proteins and enzymes associated with the bacterial surface which promote adherence to tissue and assist in biofilm formation, the cell wall itself which interacts with innate immune receptors to initiate acute inflammation, and a myriad of toxins and enzymes which promote bacterial spread by damaging tissue and lysing inflammatory cells.
Although this collection of virulence factors renders S. aureus a formidable pathogen, this organism is part of the normal human flora. S. aureus nasal carriage among the general adult population varies depending on the geographical location [15]. In the Netherlands, the S. aureus carriage rate was reported to be 24–25.2% [16,17]. In the United States from 2001–2004, 30.4% of the population was reported to carry S. aureus [18]. Among healthy Japanese, 35.7% of individuals were carriers [19]. Saxena et al. [20] reported the carriage rate in India to be 29.4%. In Kenya, 18.3% of health care workers were S. aureus carriers [21]. In Mexico, 37.1% of healthy adults carried S. aureus [22]. Taken collectively, a quarter to one-third of individuals are carriers for S. aureus [23]. The most common site of colonization is the nasal mucosa, but S. aureus can also colonize the throat and the perineum [15,16,24,25].
Antibiotic resistance in S. aureus has emerged in waves, as S. aureus has counteracted the introduction of antibiotics as the primary treatment for bacterial diseases [26]. The first wave began after two patients were treated with penicillin in 1941 [27] and 1942 [27,28]. The first report of antibiotic resistance to penicillin was in 1944 [29], and by 1948, 59% of S. aureus cultures were reported to be resistant to penicillin [30], which is now know to be due to a plasmid-encoded penicillinase. S. aureus in this group were most commonly of phage type 80/81 [31]. By the 1950s, this phage group had become pandemic [26]. Phage type 80/81 seemingly disappeared after the introduction of methicillin as the preferred antibiotic for the treatment of staphylococcal infections [26,32].
The second wave of antibiotic resistance emerged with the introduction of methicillin in 1959 [33]. The first isolations of methicillin resistant S. aureus (MRSA) were reported in 1961 [34]. These S. aureus clones carried the mecA gene, which encodes the low-affinity penicillin binding protein PBP2a that imparts broad anti-beta-lactam resistance [35]. These isolates were mainly associated with hospitals and other health care establishments and are referred to as Healthcare-Associated MRSA (HA-MRSA). A third wave of antibiotic resistance occurred with the development of methicillin-resistant strains of S. aureus outside of health care establishments, called Community-Associated MRSA (CA-MRSA). These linages are unrelated to HA-MRSA and arose from numerous genetically distinct lineages across the world. Some have spread internationally while others have remained restricted to certain geographic regions [36,37,38]. In the United States, USA300 is the most prevalent strain of CA-MRSA, but each continent appears to have its own predominant type [26].
2. S. aureus and Eye Infections
S. aureus is a leading cause of eye infections such as dacryocystitis, conjunctivitis, keratitis, cellulitis, corneal ulcers, blebitis, and endophthalmitis [39,40,41]. Contact lens wearers, especially those who wear extended-wear contact lenses or fail at proper lens hygiene, are at greater risk for the development of keratitis. S. aureus is one of the leading causes of bacterial keratitis, along with Pseudomonas aeruginosa. The number of eye infections appears to be rising due to increasing use of ocular surgical procedures, especially in elderly and diabetic populations. In the developed world, the rate of intraocular surgeries, for example for cataract removal and lens replacement, has been steadily increasing [42]. The leading infectious complication from cataract surgery is endophthalmitis, with an incidence of 0.01–0.3% [43,44]. Intravitreal injections are also being performed at greater frequencies for the treatment of neovascular eye disease, neurodegenerative diseases, and intraocular inflammation [45]. It was estimated that in 2014, 18 million intraocular injections of anti-vascular endothelial growth factor (VEGF) agents to treat age-related macular degeneration were performed [46]. The frequencies of injection-related complications has also increased [47], with endophthalmitis incidences after intravitreal injection reported to be 0.006–0.16% per injection and 0.7–1.3% over the course of treatment [48,49,50,51]. While these infection rates are statistically very low compared to diseases such as diabetes or cancer, because of the numbers of procedures, they number in the tens of thousands. In addition, eye infections can cause profound vision loss and disability, costing millions of healthcare dollars and reductions in quality of life for those affected.
S. aureus expresses an amalgam of toxins, enzymes, and secreted proteins whose virulence in humans and animal models has been well documented [13,52,53,54,55]. Studies on the contribution of these virulence factors to eye infections have focused on a few well-characterized toxins, such as α-toxin, β-toxin, γ-toxin, and PVL (Figure 2B). However, S. aureus has many more toxic factors in its armamentarium, some whose roles in eye infections have yet to be defined. Most studies of the activities of S. aureus toxins have focused on cell killing, but many recent studies have indicated that sublytic doses of some S. aureus toxins can have dramatic effects on target cells [56,57,58,59]. These reports have indicated that by altering the permeability of cell membranes, toxins can kill or manipulate the functions of the immune cells, and disrupt epithelial barriers to promote bacterial growth and spreading. By interacting with various cell surface proteins, toxins are able to target diverse cell types—mostly cells of the immune system which only express those receptors [56,57,58,59,60].
S. aureus elaborates many virulence factors which, depending upon the infection site in the eye, can manifest as serious but treatable infections such as conjunctivitis or dacryocystitis, or sight-threating infections such as corneal ulcers, endophthalmitis, or orbital cellulitis [39,40,41]. The eye is an especially vulnerable organ whose integrity, structure, and function are essential for proper sight. For the most part, our eyes resist the negative impact of infecting organisms through the concerted actions of blinking, tear film, and antimicrobial peptides and other enzymes which protect the ocular surface [55,61,62,63]. When S. aureus comes in contact with tissues of the eye and is able to circumvent these protections, infection results (Figure 2B). This review will discuss S. aureus and its toxins in the context of ocular infections and examine studies conducted in animal models to determine the roles and mechanisms of S. aureus toxins in eye infections. The strengths and limitations of these models and future steps for study will be discussed.
3. Membrane-Damaging Toxins
α-toxin is also known as α-hemolysin because it was originally named for its ability to cause β-hemolysis of red blood cells [11]. Generations of microbiology students are therefore condemned to having to remember that α-hemolysin/α-toxin causes β-hemolysis. α-toxin is a beta-barrel pore-forming toxin expressed by 95–100% of S. aureus isolates from various sites [53,64] and, in our small cohort of strains, 94% of ocular S. aureus isolates (Table 1). α-toxin has been widely studied for its role in the pathogenesis of central nervous system infections [65], endocarditis [66], endophthalmitis [52], keratitis [55,67,68], mastitis [69], pneumonia [70,71], sepsis [72,73], and skin and soft tissue infections [74,75].
Monomers of α-toxin combine in the eukaryotic cell membrane to form a seven member (heptameric) pre-pore ring-like structure [76]. The mature pore functions as an ion channel, allowing passage of Ca2+, K+, ATP and other 1–4 kDa-sized molecules into and out of the cell [77,78]. α-toxin targets cells by binding to the receptor a disintegrin and metalloproteinase 10 (ADAM10), which is required for α-toxin-mediated cytotoxicity [73,76]. Binding of α-toxin to ADAM10 results in assembly of an α-toxin-ADAM10 complex in cholesterol/sphingolipid-rich caveolar rafts [76]. At high concentrations of α-toxin, cell death predominates. However, at sublytic concentrations, α-toxin binding of ADAM10 induces the activation of ADAM10 metalloprotease activity which cleaves E-cadherin adherens junctions and results in disruption of focal adhesions and disruption of tissue barriers [76]. Also, the α-toxin/ADAM10 complex stimulates the dephosphorylation of FAD, p130cas, paxillin and src, leading to the tissue disruption that is characteristic of S. aureus infections [5] and enhanced bacterial dissemination [60].
A characteristic feature of S. aureus infections of the cornea are epithelial defects (Figure 3). In an experimental rabbit model of S. aureus keratitis [67], as organisms replicate and form microcolonies, the corneal epithelial cells die in a zone around the colonies, exposing the underlying stroma, and forming a painful ulcer that is similar to that seen in human cases (Figure 3). The role of α-toxin as a major toxin responsible for corneal epithelial ulceration in keratitis was first demonstrated in an experimental rabbit model in which isogenic mutants of the S. aureus strain 8325-4 deficient in α-toxin were injected directly into the corneal stroma. At 15 hours post infection, eyes injected with α-toxin-deficient S. aureus had lower slit lamp scores, no epithelial erosions, and less inflammation as indicated by reduced myeloperoxidase activity, compared to the wild-type S. aureus parent strain [67]. Further work using this experimental model and an α-toxin-deficient isogenic mutant confirmed that the wild-type strain produced a more severe pathology than the α-toxin mutant [68]. Similar results were observed in the same rabbit model when infections were initiated by wild-type strain Newman and its α-toxin-deficient mutant [79], and in a comparison of S. aureus 8325-4 and its α-toxin-deficient mutant in young (6–7 weeks old) and aged mice (36–48 weeks old) using the corneal scarification method [80]. In the latter study, wild-type S. aureus produced a more severe pathology than the α-toxin deficient strain, and the pathology was more severe in the aged mice. Callegan et al. [52] used an intravitreal injection model of endophthalmitis in rabbits to demonstrate that infection with an α-toxin-deficient mutant of S. aureus 8325-4 resulted in significantly reduced retinal damage compared to the wild-type strain. Recently, Putra et al. [81] reported their comparison of S. aureus strain JE2 and its α-toxin-deficient mutant in an experimental mouse model of keratitis initiated after total corneal epithelial debridement. Corneal wound healing was significantly improved following infection with the α-toxin-deficient mutant strain. This mutant was also inefficient in infecting debrided mouse corneas, further confirming the important role of α-toxin in S. aureus keratitis. Although it is clear that an absence of α-toxin results in less pathology during S. aureus keratitis, it is not known whether α-toxin is acting toward cells or triggering the immune response by similar mechanisms in different species. Different mouse strains have different responses following experimental exposure to S. aureus, but whether α-toxin contributes to these differences has not been investigated.
Topical application of purified α-toxin to the eyes of rabbits caused significant and dose-dependent inflammation of the conjunctiva and iris at six hours post application [68]. Injection of purified α-toxin into rabbit corneas also caused significant and dose-dependent inflammation of the cornea and iris [68]. These results were confirmed in the mouse keratitis model after corneal scarification. Again, corneal pathology was more severe in aged mice (36–38 weeks old) than in young mice (6–7 weeks old) [80]. However, when purified α-toxin was injected into the corneas of young (6–8 weeks old) and aged rabbits (about 30 months old), there was significantly greater disease in young rabbits than in aged ones [84]. Histological studies of rabbit eyes four hours after α-toxin injection showed epithelial cell death by necrosis and apoptosis, sloughing of viable cornea epithelial cells, severe corneal edema, and inflammatory cell migration from the tear film and from the limbal vessels into the cornea [85]. Intravitreal injection of purified α-toxin in a mouse model of endophthalmitis produced an inflammatory response, particularly via IL-1β, but only mild retinal damage and edema, and produced no significant decline in retinal function, as measured by a-or b-wave amplitudes via electroretinography [86].
The expression of α-toxin is tightly regulated. Control of toxin expression in vitro is influenced by the interplay of at least three global regulatory loci: the quorum-sensing systems agr (accessory gene regulator), sarA (staphylococcal accessory gene regulator), and sae (staphylococcal accessory protein effector). Loss of any one of these loci negatively impacts toxin production in vitro [87,88]. During in vivo growth, regulation appears to be more complicated. Booth et al. [89] reported that infection with S. aureus strain RN6390 with a defective agr resulted in a significant reduction in retinal pathology in a rabbit model of endophthalmitis. Infection with a double mutant of agr and sar resulted in an even greater reduction of pathology, but a deficiency in sar alone resulted in ocular pathology similar to that of the wild-type S. aureus. Using an implanted device model in guinea pigs, Goerke et al. [87] reported that mutation of agr or sarA in strains RN6390 and Newman did not impact the level of transcription of the gene for α-toxin, hla. However, loss of sae caused an almost total downregulation of α-toxin production in both strains. Xiong et al. [88] examined the α-toxin production of two S. aureus strains: RN6390, which lacks a functional sigB regulon, and SH1000, which has a repaired sigB regulon. SigB negatively influences the production of α-toxin in vitro [90]. Xiong et al. [88] reported that in vitro isogenic mutants in RN6390 of agr, sarA, and agr/sarA had significant reductions in α-toxin production. Mutation of sae completely abolished α-toxin production in both strains during in vitro growth. SH1000 produced less α-toxin overall than RN6390 but the results of the mutations were similar, except that mutating sarA did not reduce α-toxin production. During in vivo growth with RN6390, using an infective endocarditis model in rabbits, only mutation of sae resulted in a significant reduction in α-toxin production, compared to the wild-type strain. With SH1000 there was no significant reduction in toxin production with any of the mutant strains. Even with the sae mutation, both strains produced measurable amounts of αtoxin during in vivo growth. These data indicate that during in vivo growth other factors, in addition to agr, sarA and sae, are involved in α-toxin regulation. O’Callaghan et al. [68] injected agr-deficient mutants of strain 8325-4 into the corneas of rabbits and reported that the agr mutant was even less virulent than the α-toxin mutant. Girgis et al. [80] also reported that the agr mutant was less virulent than the α-toxin mutant in the mouse keratitis scarification model. Both authors surmised that these outcomes were likely due to deficiencies in α-toxin and all the other toxins under its control in the agr-deficient mutants.
The aforementioned studies confirm an important role for α-toxin in the pathogenesis of S. aureus keratitis and endophthalmitis. A rational strategy for improving the therapeutic outcome of keratitis would be to block the effect of α-toxin on the cornea. As nearly all ocular S. aureus isolates have been reported to possess the hla gene (Table 1 and [64]), this virulence factor is a viable target. Passive and active immunization of rabbits against α-toxin has been shown to be effective at reducing corneal disease in the rabbit model of keratitis [79]. Active immunization against α-toxin prevented the formation of corneal epithelial erosions [79]. Whether immunization strategies would be effective in limiting the effects of α-toxin in endophthalmitis or other ocular S. aureus infections remains an open question. Recently, nanoparticles composed of erythrocyte membranes surrounding a biologically inert poly-lactic-co-glycolic core have been developed to function as a type of decoy to neutralize pore-forming toxins [91]. These nanoparticles, termed nanosponges, neutralized α-toxin and reduced hemolytic activity in vitro, protected mice from developing staphylococcal α-toxin induced skin lesions, and decreased mortality after systemic injection of a lethal dose of α-toxin [91]. In a mouse model of S. aureus endophthalmitis, Coburn et al. [92] reported that intraocular injection of nanosponges alone following infection did not result in improved retinal function compared to untreated mice. However, injection of nanosponges in conjunction with the fourth-generation fluoroquinolone gatifloxacin resulted in decreased inflammation, less damage to the retinal architecture, and significantly improved retinal function compared to untreated or gatifloxacin only treated mice following intraocular infection with an MRSA ocular isolate [92]. These studies suggested that nanosponges might be a viable adjunctive therapy for intraocular infections caused by S. aureus. Together, these therapeutics studies imply that blocking α-toxin would improve the therapeutic outcome of ocular S. aureus infections.
4. Bi-Component Toxins
In S. aureus, bi-component leukotoxins are composed of a pair of proteins designated S (slow) and F (fast) based on their elution speeds [93]. Initial binding of the leukotoxin receptor is mediated by the S-component, followed secondarily by binding of the F-component, resulting in formation of a lytic pore-forming octamer (β-barrel) of alternating subunits that insert into the cell membrane, resulting in osmotic imbalance and cell lysis [56,94]. Human S. aureus isolates can encode up to five leukocidin toxins: Panton–Valentine leukocidin (PVL), γ-hemolysin AB (HlgAB), γ-hemolysin CB (HlgCB), LukED, and LukAB (also referred to as LukGH) [94]. Elucidation of the role of these toxins in human disease has been hampered by the species and cell specificities of some of the toxins [95].
Panton–Valentine leukocidin (PVL) is a prophage-encoded toxin present in about 5% of S. aureus isolates and most CA-MRSA strains [53]. PVL binds to complement component C5a anaphylatoxin chemotactic receptors to target neutrophils, monocytes, macrophages, natural killer cells, dendritic cells, and T lymphocytes [60,94]. C5aR1 is highly expressed on phagocytic cells, but C5aR2 has low expression on neutrophils [94,95,96]. The cytotoxicity of PVL can be blocked with C5aR1 antagonists [94,96]. PVL is active against human and rabbit neutrophils, but macaque, cow, and mouse neutrophils are resistant to its effects [93]. PVL stimulates inflammasome activation in monocytes and primary macrophages at sublethal doses [58]. S. aureus β-toxin, δ-toxin, γ-toxin, LukDE, and PSMα3 synergize with PVL to amplify IL-1β release to trigger inflammation [58]. Based on species specificity, the most appropriate animal models for the study of PVL are in rabbits and humanized mice with engrafted primary human haematopoietic cells [95].
Not deterred by an acknowledged lack of lytic activity by PVL against mouse neutrophils, Zaidi et al. [97] tested PVL-deficient isogenic mutants of S. aureus USA400 and USA300 (currently the predominant CA-MRSA strain in the United States) in an experimental scratch model of mouse keratitis. Inactivation of PVL in the LAC strain of USA300 resulted in reduced corneal opacity and reduced corneal bacterial counts compared to the wild-type strain. However, results with USA400 strains depended on the strain tested. Infection with the isogenic PVL-deficient mutant of MW2 and its wild-type parent strain resulted in similar degrees of corneal opacity and corneal bacterial counts. Infection with the isogenic PVL-deficient mutant of NRS 193 had reduced corneal opacity and reduced corneal bacterial counts compared to its wild-type parent. Infection with the isogenic PVL-deficient mutant of NRS 194 had reduced corneal opacity, but did not have reduced corneal bacterial counts, as compared to its wild-type parent strain. In this study, topical applications of neutralizing polyclonal antibodies against PVL resulted in significantly reduced corneal opacity and bacterial numbers in corneas infected with USA300 strains LAC and SF8300, but in corneas infected with USA400 strains MW2 and NRS 193, the bacterial loads were unaffected and there was no reduction in corneal opacity. Corneas infected with strain NRS 194 showed a reduction in corneal opacity, but bacterial counts were unaffected following the application of anti-PVL antibodies [97].
Siqueira et al. [98] demonstrated that intravitreal injections of six different combinations of the PVL and γ-toxin subunits (S and F) from S. aureus ATCC strain 49,775 caused acute inflammatory reactions involving the posterior and anterior chambers as well as the conjunctiva and eyelids. The clinical and histological signs of inflammation began by 4 h after injection and persisted for the 5 days of the experiment. The toxicity was dose-dependent. Certain combinations were more toxic than others. The most virulent combination was the PV-S subunit plus γ-toxin-F subunit, although it is not known if this combination occurs naturally. Liu et al. [99] injected PVL into rabbit eyes and identified retinal ganglion cells as the primary targets of the toxin, and these cells were the only ones identified to express C5a receptors. Binding of PVL triggered increased IL-6 expression in the retina and apoptosis of microglial cells. Liu et al. [100] demonstrated similar results using a rabbit retinal explant model. However, in addition to PVL binding to retinal ganglion cells, the toxin also co-localized to horizontal cells which did not express C5a receptor, and there was no significant increase in IL-6 as compared to the control explants. Peterson et al. [101] recently reported that the PVL gene was detected in only 14.7% of S. aureus keratitis isolates in their study, so it is unclear how clinically relevant this toxin is to ocular disease.
γ-toxin consists of two bicomponent pore-forming toxins: HlgAB and HlgCB, which are encoded in the core genome and are reported to be present in 99.8% of clinical isolates [53,102]. During in vitro culture, γ-toxin is highly upregulated in the presence of blood [103] and upon phagocytosis by neutrophils [104]. HlgAB targets the chemokine receptors CXCR1, CXCR2, and CCR2 [105], while HlgCB targets the compliment component C5a receptors C5aR1 and C5aR2 [97]. These receptors are highly expressed on phagocytic cells [105]. HlgCB is active against neutrophils of human, rabbit, macaque, and cow [105]. Mouse neutrophils are resistant to HlgCB. In mice only CCR2 is able to act as a receptor for HlgAB [105].
Supersac et al. [106] published a study on the role of γ-toxin using a rabbit model of endophthalmitis infected with the S. aureus strain Newman, which the authors believed did not produce α-toxin or β-toxin [107]. After intravitreal injection of either a γ-toxin mutant of Newman, the wild-type, or a γ-toxin complemented strain, all three strains produced a strong inflammatory response, with the only difference being that the γ-toxin mutant did not cause inflammation of the eyelid [106]. More recent studies have shown that while strain Newman has a truncated copy of β-toxin, it does have the α-toxin gene [108], is a weak producer of α-toxin in vitro [79,87], and is a strong producer of α-toxin in vivo [87]. Using the rabbit model of endophthalmitis, Callegan et al. [52] showed with an isogenic mutant of γ-toxin in strain 8325-4 that loss of this gene had no positive effect on retinal function. Dajcs et al. [79], using a rabbit model of keratitis and a γ-toxin mutant of the Newman strain, reported significantly lower slit lamp scores and reduced numbers of PMN in the corneal stroma as compared to the wild-type strain or the genetically rescued strain. Outcomes here appear to be model- and strain-dependent.
Phenol-soluble modulins (PSMs), of which δ-toxin is a member, are a family of small (20–44 amino acids) amphipathic secreted peptides which are only produced by the members of the genus Staphylococcus [109]. All S. aureus have the hld gene and two loci with genes coding for psm α and psmβ [110]. However, the widely used S. aureus strain 8325-4 is unique for not having the gene for psmα [111]. PSMs have multiple roles in S. aureus pathogenesis, such as facilitating biofilm dissemination, cytolytic activity, and proinflammatory activity [110,112]. PSMα induces the release of TLR2-activating lipoproteins from bacterial cells [113]. While micromolar concentrations of PSMs cause cytolysis, nanomolar concentrations cause inflammation [114]. PSMs are detected by the formyl peptide receptor 2 (FPR2) [114,115] on mammalian cells, resulting in the attraction and activation of neutrophils and inflammation [110]. The mechanism of cytolysis is not clear, but PSMs appear to have a detergent-like action on the cell membrane [112]. Cytolysis is likely nonspecific and receptor independent [116]. Many species of staphylococci express PSM, but S. aureus expresses highly cytotoxic forms, such as PSMα1–α4 [116]. Although PSMs are among the most abundant proteins secreted in staphylococcal culture filtrates [109], we are unaware of any studies using animal models to examine their role in eye infections.
5. Enzymes
β-toxin is a neutral sphingomyelinase, not a hemolytic toxin per se. β-toxin is able to hydrolyze the plasma membrane lipid sphingomyelin [117] and is responsible for α-hemolysis on blood agar plates. Reports on the frequency of the hlb gene in S. aureus clinical isolates vary. Although the hlb gene was reported to be present in 39–57% of clinical isolates [64,118] and, in our small cohort, 50% of ocular isolates (Table 1), van Wamel et al. [119] reported that in 90% of the isolates examined, the β-toxin gene had been inactivated by phage φSa3 insertion. However, Salgado-Pabon et al. [120] reported that during in vivo and in vitro growth, phage φSa3 can excise from a subpopulation of S. aureus and translocate to atypical sites in the chromosome, restoring β-toxin production. β-toxin is toxic to human monocytes, but is inactive at equal concentrations against human erythrocytes, fibroblasts, granulocytes, and lymphocytes [121].
β-toxin has not been shown epidemiologically to be associated with a specific disease or infection severity criteria [112]. However, O’Callaghan et al. [68] demonstrated in a rabbit model of keratitis that topical application of β-toxin caused significant conjunctival inflammation, and corneal injection of purified β-toxin resulted in a rapid edematous reaction in the sclera. Intrastromal injection of an isogenic β-toxin-deficient mutant of S. aureus strain 8325-4 caused less scleral edema than its wild-type parental strain, but slit lamp scores, epithelial erosions, and intrastromal ulcers were similar to that of keratitis cause by the wild-type strain [68]. Similar results were reported in an experimental rabbit model of endophthalmitis. S. aureus 8325-4 and the same β-toxin-deficient mutant were compared for their virulence following intravitreal injection, and the β-toxin-deficient mutant resulted in significantly reduced retinal damage compared to the wild-type strain [52]. This reduction in damage, however, was not as significant as the reduction of damage in eyes infected with the α-toxin-deficient mutant of the same wild-type parent strain. Together, these studies suggest that β-toxin may contribute to ocular changes in the cornea and within the eye, but its expression in the eye is not essential for complete virulence.
6. Discussion
The role of α-toxin in eye infections has been well established by studies comparing α-toxin-deficient isogenic mutants with their wild-type parental strains in models of keratitis and endophthalmitis in both rabbits and mice (Table 2). Ocular administration of purified α-toxin has also been tested. α-toxin’s role as a major virulence factor in ocular S. aureus infections is clear cut. However, the studies of the role of the bi-component toxins in ocular infections have been less conclusive. Studies examining the role of PVL were conducted in the mouse [97], the cells of which have been reported to be resistant to PVL [105]. The studies by Supersac et al. [106] and Dajcs et al. [79] both used a γ-toxin-deficient isogenic mutant of the Newman strain in a rabbit model. While Supersac et al. [106] reported a strong inflammatory response following intravitreal injection of wild-type or γ-toxin-deficient Newman, Dajcs et al. [79] reported significantly less inflammation following instrastromal injection of the same γ-toxin-deficient Newman mutant. These two studies suggest a model-dependent role for the virulence of γ-toxin. Studies of the importance of the enzyme β-toxin in corneal and intraocular infection models have reported less virulence in β-toxin-deficient mutants [67]; however, the extent to which ocular virulence is muted in β-toxin-deficient or γ-toxin-deficient ocular infections is not nearly as significant as that of ocular infections caused by α-toxin-deficient S. aureus.
Except for α-toxin, our understanding of the role of S. aureus toxins in eye infections is limited. Limitations that should be accounted for in such studies are the species-specific sensitivities to several of the bi-component toxins, and the variability in toxin types and the amounts produced by the various isolates commonly used to study S. aureus infections. Many of the studies reviewed in this paper used S. aureus isolate 8325-4, which has a published genome, numerous mutants, and has been tested in many non-ocular infection models. S. aureus 8325-4 was isolated in 1960, before the development of CA-MRSA and HA-MRSA, has been phage cured, is known to lack phenol soluble modulin α3, and has shown significant variation in α-toxin production depending on which lab isolate is used [111]. The use of clinically relevant ocular isolates could provide insight into which genes are important in eye infections and give insight into the interplay of the various toxins produced by the pathogens. However, isolates should first be characterized and compared for relevance to the disease model. Most of the data reviewed in this paper was generated using animal models of infection (Table 2). Although obvious, the many physiological similarities and differences must be accounted for when interpreting and translating data to humans [122,123,124]. All animal infection models have limitations, but most can provide useful information. Animal models not only provide us with the control necessary to initiate various aspects of the infection process, but also facilitate study of the host response to infection, an essential player absent in cell or tissue culture models [125]. Understanding the limitations of these models is important in evaluating the usefulness of the data it produces. For example, rabbit keratitis models using intrastromal injections of bacteria are technically simple, highly controlled, and the output is highly reproducible [55]. However, this model does not replicate the normal manner in which humans develop bacterial keratitis and it bypasses important steps in the infection process. Mouse models of bacterial keratitis suffer from the limitation that only certain mouse strains reliably develop infections [80]. Some mouse strains clear the infections without treatment, unlike infections in humans. The debridement of the mouse corneal epithelium to prepare it for application of bacteria [81] is also not the most accurate model of how the human cornea is damaged prior to infection, unless there is a large wound from chemical or mechanical trauma. To date, the scratch model, similar to that which is used in studies of Pseudomonas keratitis [126], is as close an approximation of what may happen to a human cornea prior to S. aureus infection that has been attempted. Mouse and rabbit models of bacterial endophthalmitis initiated by intravitreal injections are an approximation of traumatic endophthalmitis, but they lack the traumatic aspect, which may include significant tissue damage and the presence of blood in the eye [125]. Injection or topical application of purified toxin (Table 3) is a strategy that may approximate the toxin concentrations needed for tissue damage and may be helpful in studying the mechanisms by which toxins affect these tissues, but these studies may lack physiological relevance because the amounts and timing of toxin production have not been quantified, bacteria do not produce a single bolus of toxin in infected tissue, and different strains produce different complements of toxin during infection.
A common and relatively straightforward method for examining the importance of a toxin to the pathogenesis of a disease is to create an isogenic mutant specifically deficient in that toxin and examine how the pathology of the disease is altered in the preferred model. While the absence of a single toxin may significantly alter pathogenicity if an organism produces few toxins, for S. aureus and many other pathogens, this strategy is rather like removing one part of a car to determine how much damage it does when it hits an object. This practice will provide us with valuable information about how important some parts of the car are, but it will vastly underestimate the importance of most of them. However, an informative start has been made into understanding the roles of S. aureus toxins in eye infections. The toxins of S. aureus appear to have robust cytotoxic actions on ocular tissues and perhaps subtoxic activities in targeting the both the innate and adaptive branches of the immune system to both modulate the immune response and to block the development of a protective response. A more complete understanding of these activities will be facilitated by developing and using models that are more physiologically relevant and approach the environments and mechanisms by which S. aureus infects the human eye.
Author Contributions
Conceptualization, R.A., M.H.M., and M.C.C.; Investigation, R.A.; Resources, M.C.C.; Writing—Original Draft Preparation, R.A. and M.C.C.; Writing—Review and Editing, R.A., F.C.M., P.S.C., and M.C.C.; Visualization, R.A., M.H.M., and M.C.C.; Project Administration, M.C.C.; Funding Acquisition, M.C.C.
Funding
Our research and some of the studies mentioned herein were supported by National Institutes of Health grants R01EY028810, R01EY024140, R01EY025947 and R21EY028066 (to MCC), NEI Vision Core Grant P30EY027125 (to MCC, OUHSC), a Presbyterian Health Foundation Equipment Grant (to Robert E. Anderson, OUHSC), and an unrestricted grant to the Dean A. McGee Eye Institute from Research to Prevent Blindness.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Ogston, A. Report upon micro-organisms in surgical diseases. Br. Med. J. 1881, 1, 369–375. [Google Scholar] [CrossRef] [PubMed]
- Fetsch, A. Staphylococcus Aureus; Academic Press: Cambridge, MA, USA, 2018; p. 3. [Google Scholar]
- Rosenbach, F.J. Mikro-organismen bei den Wund-Infection-krankheiten des Menschen; Wiesbaden, J.F. Bergmann: Plano, TX, USA, 1884; pp. 19–21. [Google Scholar]
- Hooker, C. Diphtheria, Immunization and the Bundaberg Tragedy: A Study of Public Health in Australia. Health Hist. 2000, 2, 52–78. [Google Scholar] [CrossRef]
- Berube, B.J.; Wardenburg, J.B. Staphylococcus aureus α-Toxin: Nearly a Century of Intrigue. Toxins 2013, 5, 1140–1166. [Google Scholar] [CrossRef] [PubMed]
- Burnet, F.M. The Exotoxins of Staphylococcus Pyogenes Aureus. J. Pathol. Bacteriol. 1929, 32, 717–734. [Google Scholar] [CrossRef]
- Burnet, F.M. The Production of Staphylococcal Toxin. J. Pathol. Bacteriol. 1930, 33, 1–16. [Google Scholar] [CrossRef]
- Burky, E.L. Studies on Cultures and Broth Filtrates of Staphylococci. J. Immunol. 1933, 25, 419–437. [Google Scholar]
- Woolpert, O.C.; Dack, G.M. Relation of Gastro-Intestinal Poison to Other Toxic Substances Produced by Staphylococci. J. Infect. Dis. 1933, 52, 6–19. [Google Scholar] [CrossRef]
- Bigger, J.W. The Production of Staphylococcal Haemolysin With Observations on Its Mode of Action. J. Pathol. Bacteriol. 1933, 36, 87–114. [Google Scholar] [CrossRef]
- Glenny, A.T.; Stevens, M.F. Staphylococcus Toxins and Antitoxins. J. Pathol. Bacteriol. 1935, 40, 201–210. [Google Scholar] [CrossRef]
- Kumar, S.; Lindorfer, R.K. The Characterization of Staphylococcal Toxins. J. Exp. Med. 1962, 30, 1095–1106. [Google Scholar] [CrossRef]
- Bernheimer, A.W.; Schwartz, L.L. Isolation and Composition of Staphylococcal Alpha Toxin. J. Gen. Microbiol. 1963, 30, 455–468. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lowy, F.D. Staphylococcus Aureus Infections. N. Engl. J. Med. 1998, 339, 520–532. [Google Scholar] [CrossRef] [PubMed]
- Sollid, J.U.E.; Furberg, A.S.; Hanssen, A.M.; Johannessen, M. Staphylococcus aureus: Determinates of human carriage. Infect. Genet. Evolut. 2014, 21, 531–541. [Google Scholar] [CrossRef] [PubMed]
- Wertheim, H.F.L.; Vos, M.C.; Ott, A.; van Belkum, A.; Voss, A.; Kluytmans, J.; van Keulen, P.; Vandenbroucke-Grauls, C.; Meester, M.; Verbrugh, H. Risk and outcome of nosocomial Staphylococcus aureus bacteremia in nasal carriers versus non-carriers. Lancet 2004, 364, 703–705. [Google Scholar] [CrossRef]
- Lebon, A.; Moll, H.A.; Tavakol, M.; van Wamel, W.J.; Jaddoe, V.; Hofman, A.; Verbrugh, H.A.; van Belkum, A. Correlation of Bacterial Colonization Status between Mother and Child: The Generation R. Study. J. Clin. Microbiol. 2010, 48, 960–962. [Google Scholar] [CrossRef] [PubMed]
- Gorwitz, R.J.; Kruszon-Moran, D.; McAllister, S.K.; McQuillan, G.; McDougal, L.K.; Fosheim, G.E.; Jensen, B.J.; Killgore, G.; Tenover, F.C.; Kuehnert, M.J. Changes in the Prevalence of Nasal Colonization with Staphylococcus aureus in the United States 2001–2004. J. Infect. Dis. 2008, 197, 1226–1234. [Google Scholar] [CrossRef] [PubMed]
- Uemura, E.; Kakinohana, S.; Higa, N.; Toma, C.; Nakasone, N. Comparative characterization of Staphylococcus aureus isolates from throats and noses of healthy volunteers. Jpn. J. Infect. Dis. 2004, 57, 21–24. [Google Scholar]
- Saxena, S.; Singh, K.; Talwar, V. Methicillin-resistant Staphylococcus aureus prevalence in community in the East Delhi area. Jpn. J. Infect. Dis. 2003, 56, 54–56. [Google Scholar]
- Omuse, G.; Kariuki, S.; Revathi, G. Unexpected absence of methicillin-resistant Staphylococcus aureus nasal carriage by healthcare workers in a tertiary hospital in Kenya. J. Hosp. Infect. 2012, 80, 71–73. [Google Scholar] [CrossRef]
- Hamdan-Partida, A.; Sainz-Espunes, T.; Bustos-Martinez, J. Characterization and Persistence of Staphylococcus aureus Strains Isolated from the Anterior Nares and Throats of Healthy Carriers in a Mexican Community. J. Clin. Microbiol. 2010, 48, 1701–1705. [Google Scholar] [CrossRef]
- Eriksen, N.; Espersen, F.; Rosdahl, V.T.; Jensen, K. Carriage of Staphylococcus aureus among 104 healthy persons during a 19-month period. Epidemiol. Infect. 1995, 115, 51–60. [Google Scholar] [CrossRef] [PubMed]
- Kluytmans, J.; van Belkum, A.; Verbruch, H. Nasal Carriage of Staphylococcus aureus: Epidemiology, Underlying Mechanisms, and Associated Risks. Clin. Microbiol. Rev. 1997, 10, 505–520. [Google Scholar] [CrossRef] [PubMed]
- Acton, D.S.; Plat-Sinnige, M.J.; van Wamel, W.; de Groot, N.; van Belkum, A. Intestinal carriage of Staphylococcus aureus: how does its frequency compare with that of nasal carriage and what is its impact? Eur. Clin. Microbiol. Infect. Dis. 2009, 28, 115–127. [Google Scholar] [CrossRef] [PubMed]
- Chambers, H.F.; DeLeo, F.R. Waves of Resistance: Staphylococcus aureus in the Antibiotic Era. Nat. Rev. Microbiol. 2009, 7, 629–641. [Google Scholar] [CrossRef] [PubMed]
- Gaynes, R. The Discovery of Penicillin–New Insights After More Than 75 years of Clinical Use. Emerging Infect. Dis. J. 2017, 23, 849–853. [Google Scholar] [CrossRef]
- Fleming, A. Penicillin its Practical Applications, 2nd ed.; Butterworth & Co.: London, UK, 1950; p. 15. [Google Scholar]
- Kirby, W.M.M. Extraction of a highly potent penicillin inactivator from penicillin resistant staphylococci. Science 1944, 99, 452–453. [Google Scholar] [CrossRef]
- Barber, M.; Rozwadowska-Dowzenko, M. Infection by Penicillin-Resistant Staphylococci. Lancet 1948, 23, 641–644. [Google Scholar] [CrossRef]
- Blair, J.E.; Carr, M. Distribution of Phage Groups of Staphylococcus aureus in the Years 1927 through 1947. Science 1960, 132, 1247–1248. [Google Scholar] [CrossRef] [PubMed]
- Jevons, M.P.; Parker, M.T. The evolution of new hospital strains of Staphylococcus aureus. J. Clin. Pathol. 1964, 17, 243–250. [Google Scholar] [CrossRef] [Green Version]
- Chambers, H.F. Methicillin-Resistant Staphylococci. Clin. Microbiol. Rev. 1988, 1, 173–186. [Google Scholar] [CrossRef]
- Barber, M. Methicillin-resistant staphylococci. J. Clin. Pathol. 1961, 14, 385–393. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, A.S.; de Lencastre, H.; Garau, J.; Kluytmans, J.; Malhotra-Kumar, S.; Peschel, A.; Harbarth, S. Methicillin-resistant Staphylococcus aureus. Nat. Rev. 2018, 4, 1188–1196. [Google Scholar]
- Vandensch, F.; Naimi, T.; Enright, M.C.; Lina, G.; Nimmo, G.R.; Heffernan, H.; Liassine, N.; Bes, M.; Greenland, T.; Reverdy, M.; et al. Community-Acquired Methicillin-Resistant Staphylococcus aureus Carrying Panton-Valentine Leukocidin Genes: Worldwide Emergence. Emerg. Infect. Dis. 2003, 9, 978–984. [Google Scholar] [CrossRef] [PubMed]
- Mediavilla, J.R.; Chen, L.; Mathema, B.; Kreiswirth, B.N. Global epidemiology of community-associated methicillin resistant Staphylococcus aureus (CA-MRSA). Curr. Opin. Microbiol. 2012, 15, 588–595. [Google Scholar] [CrossRef] [PubMed]
- Boswihi, S.S.; Udo, E.E. Methicillin-resistant Staphylococcus aureus: An update on the epidemiology, treatment options and infection control. Curr. Med. Res. Pract. 2018, 8, 18–24. [Google Scholar] [CrossRef]
- Blomquist, P.H. Methicillin-Resistant Staphylococcus aureus Infections of the Eye and Orbit. Trans. Am. Ophthalmol. Soc. 2006, 104, 322–345. [Google Scholar]
- Hsiao, C.; Chuang, C.; Tan, H.; Ma, D.; Lin, K.; Chang, C.; Huang, Y. Methicillin-Resistant Staphylococcus aureus Ocular Infection: A 10-Year Hospital-Based Study. Ophthalmology 2012, 119, 522–527. [Google Scholar] [CrossRef] [PubMed]
- Chuang, C.; Hsiao, C.; Tan, H.; Ma, D.; Lin, K.; Chang, C.; Huang, Y. Staphylococcus aureus Ocular Infection: Methicillin-Resistance, Clinical Features, and Antibiotic Susceptibilities. PLoS ONE 2012, 8, e42437. [Google Scholar] [CrossRef]
- Brian, G.; Taylor, H. Cataract blindness-challenges for the 21st century. Bull. World Health Organ. 2001, 79, 249–256. [Google Scholar]
- Endophthalmitis Study Group. Prophylaxis of postoperative endophthalmitis following cataract surgery: Results of the ESCRS multicenter study and identification of risk factors. J. Cataract Refract. Surg. 2007, 33, 978–988. [Google Scholar] [CrossRef]
- Taban, M.; Behrens, A.; Newcomb, R.L.; Nobe, M.Y.; Saedi, G.; Sweet, P.M.; McDonnell, P.J. Acute endophthalmitis following cataract surgery: A systemic review of the literature. Arch. Ophthalmol. 2005, 123, 613–620. [Google Scholar] [CrossRef] [PubMed]
- Peyman, G.A.; Lad, E.M.; Moshfeghi, D.M. Intravitreal injection of therapeutic agents. Retina 2009, 29, 875–912. [Google Scholar] [CrossRef] [PubMed]
- Rupenthal, I.D. Sector Overview Ocular Drug Delivery: Exciting Times Ahead. Available online: http://ondrugdelivery.com/publications/54/Sector_Overview.pdf (accessed on 1 June 2019).
- Sadaka, A.; Durand, M.L.; Gilmore, M.S. Bacterial endophthalmitis in the age of outpatient intravitreal therapies and cataract surgeries: Host-microbe interactions in intraocular infection. Prog. Retin. Eye Res. 2012, 31, 316–331. [Google Scholar] [CrossRef] [PubMed]
- VEGF Inhibition Study in Ocular Neovascularization (V.I.S.I.O.N.) Clinical Trial Group; D’Amico, D.J.; Masonson, H.N.; Patel, M.; Adamis, A.P.; Cunningham, E.T., Jr.; Guyer, D.R.; Katz, B. Pegaptanib sodium for neovascular age-related macular degeneration: Two-year safety results of the two prospective, multicenter, controlled clinical trials. Ophthalmology 2006, 113, 992–1001.e6. [Google Scholar]
- Diago, T.; McCannel, C.A.; Bakri, S.J.; Pulido, J.S.; Edwards, A.O.; Pach, J.M. Infectious endophthalmitis after intravitreal injection of antiangiogenic agents. Retina 2009, 29, 601–605. [Google Scholar] [CrossRef] [PubMed]
- Klein, K.S.; Walsh, M.K.; Hassan, T.S.; Halperin, L.S.; Castellarian, A.A.; Roth, D.; Driscoll, S.; Prenner, J.L. Endophthalmitis after anti-VEGF injections. Ophthalmology 2009, 116, 1225. [Google Scholar] [CrossRef]
- Sampat, K.M.; Garg, S.J. Complications of intravitreal injections. Curr. Opin. Ophthalmol. 2010, 21, 178–183. [Google Scholar] [CrossRef] [PubMed]
- Callegan, M.C.; Engelbert, M.; Parke, D.W.; Jett, B.D.; Gilmore, M.S. Bacterial Endophthalmitis: Epidemiology, Therapeutics, and Bacterium-Host Interactions. Clin. Microbiol. Rev. 2002, 15, 111–124. [Google Scholar] [CrossRef] [Green Version]
- Grumann, D.; Nubel, U.; Borker, B.M. Staphylococcus aureus toxins–Their functions and genetics. Infect. Genet. Evolut. 2014, 21, 583–592. [Google Scholar] [CrossRef]
- Marquart, M.E.; O’Callaghan, R.J. Infectious Keratitis: Secreted Bacterial Proteins That Mediate Corneal Damage. J. Ophthalmol. 2013, 2013, 369094. [Google Scholar] [CrossRef]
- O’Callaghan, R.J. The Pathogenesis of Staphylococcus aureus Eye Infections. Pathogens 2018, 7, 9. [Google Scholar] [CrossRef] [PubMed]
- Alonzo, F.; Torres, V.J. Bacterial Survival Amidst and Immune Onslaught: The Contribution of the Staphylococcus aureus Leukotoxins. PLoS Pathog. 2013, 9, e1003143. [Google Scholar] [CrossRef] [PubMed]
- Inoshima, I.; Inoshima, N.; Wilke, G.A.; Powers, M.E.; Frank, K.M.; Wang, Y.; Wardenburg, J.B. Staphylococcus aureus pore-forming toxin subverts the activity of ADAM10 to cause lethal infection in mice. Nat. Med. 2011, 17, 1310–1314. [Google Scholar] [CrossRef] [PubMed]
- Perret, M.; Badiou, C.; Lina, G.; Burbaud, S.; Benito, Y.; Bes, M.; Cottin, V.; Couzon, F.; Juruj, C.; Dauwalder, O.; et al. Cross-talk between Staphylococcus aureus leukocidins-intoxicated macrophages and lung epithelial cells trigger chemokine secretion in an inflammasome-dependent manner. Cell. Microbiol. 2012, 14, 1019–1036. [Google Scholar] [CrossRef] [PubMed]
- Seilie, E.S.; Wardenburg, J.B. Staphylococcus aureus pore-forming toxins: The interface of pathogen and host complexity. Semin. Cell Dev. Biol. 2017, 72, 101–116. [Google Scholar] [CrossRef] [PubMed]
- Peraro, M.D.; van der Goot, F. Pore-forming toxins: Ancient, but never really out of fashion. Nat. Rev. 2016, 14, 77–92. [Google Scholar] [CrossRef] [PubMed]
- Ueta, M.; Kinoshita, S. Innate immunity of the ocular surface. Brain Res. Bull. 2010, 81, 219–228. [Google Scholar] [CrossRef] [PubMed]
- Ueta, M.; Kinoshita, S. Ocular surface inflammation is regulated by innate immunity. Prog. Retin. Eye Res. 2012, 31, 551–575. [Google Scholar] [CrossRef] [PubMed]
- Foulsham, W.; Coco, G.; Amouzegar, A.; Chauhan, S.K.; Dana, R. When Clarity Is Crucial: Regulating Ocular Surface Immunity. Trends Immunol. 2018, 39, 288–301. [Google Scholar] [CrossRef]
- Booth, M.C.; Pence, L.M.; Mahasreshti, P.; Callegan, M.C.; Gilmore, M.S. Clonal Associations among Staphylococcus aureus Isolates from Various Sites of Infection. Infect. Immun. 2001, 69, 345–352. [Google Scholar] [CrossRef]
- Kielian, T.; Cheung, A.; Hickey, W.F. Diminished Virulence of an Alpha-Toxin Mutant of Staphylococcus aureus in Experimental Brain Abscesses. Infect. Immun. 2001, 69, 6902–6911. [Google Scholar] [CrossRef] [PubMed]
- Bayer, A.; Ramos, M.D.; Menzies, B.E.; Yeaman, M.R.; Shen, A.J.; Cheung, A.L. Hyperproduction of Alpha-Toxin by Staphylococcus aureus Results in Paradoxically Reduced Virulence in Experimental Endocarditis: A Host Defense Role for Platelet Microbicidal Proteins. Infect. Immun. 1997, 65, 4652–4660. [Google Scholar] [PubMed]
- Callegan, M.C.; Engel, L.S.; Hill, J.M.; O’Callaghan, R.J. Corneal Virulence of Staphylococcus aureus: Roles of Alpha-Toxin and Protein A in Pathogenesis. Infect. Immun. 1994, 62, 2478–2482. [Google Scholar] [PubMed]
- O’Callaghan, R.J.; Callegan, M.C.; Moreau, J.M.; Green, L.C.; Foster, T.J.; Hartford, O.M.; Engel, L.S.; Hill, J.M. Specific Roles of Alpha-toxin and Beta-Toxin during Staphylococcus aureus Corneal Infection. Infect. Immun. 1997, 65, 1571–1578. [Google Scholar] [PubMed]
- Jonsson, P.; Lindberg, M.; Haraldsson, I.; Wadstrom, T. Virulence of Staphylococcus aureus in a Mouse Mastitis Model: Studies of Alpha Hemolysin, Coagulase, and Protein A as Possible Virulence Determinates with Protoplast Fusion and Gene Cloning. Infect. Immun. 1985, 49, 765–769. [Google Scholar] [PubMed]
- Wardenburg, J.B.; Patel, R.J.; Schneewind, O. Surface Proteins and Exotoxins Are Required for the Pathogenesis of Staphylococcus aureus Pneumonia. Infect. Immun. 2007, 75, 1040–1044. [Google Scholar] [CrossRef] [PubMed]
- Wardenburg, J.B.; Bae, T.; Otto, M.; Deleo, F.R.; Schneewind, O. Poring over pores: Alpha-hemolysin and Panton-Valentine leukocidin in Staphylococcus aureus pneumonia. Nat. Med. 2007, 13, 1405–1406. [Google Scholar] [CrossRef] [PubMed]
- Menzies, B.E.; Kernodle, D.S. Passive Immunization with Antiserum to a Nontoxic Alpha-Toxin Mutant from Staphylococcus aureus Is Protective in a Murine Model. Infect. Immun. 1996, 64, 1839–1841. [Google Scholar] [PubMed]
- Powers, M.E.; Kim, H.K.; Wang, Y.; Wardenburg, J.B. ADAM10 Mediates Vascular Injury Induced by Staphylococcus aureus α-Hemolysin. J. Infect. Dis. 2012, 206, 352–356. [Google Scholar] [CrossRef]
- Kennedy, A.D.; Wardenburg, J.B.; Gardner, D.J.; Long, D.; Whitney, A.R.; Braughton, K.R.; Schneewind, O.; DeLeo, F.R. Targeting of Alpha-Hemolysin by Active or Passive Immunization Decreases Severity of USA300 Skin Infection in a Mouse Model. J. Infect. Dis. 2010, 202, 1050–1058. [Google Scholar] [CrossRef]
- Patel, A.H.; Nowland, P.; Weavers, E.D.; Foster, T. Virulence of Protein A-Deficient and Alpha-Toxin-Deficient Mutants of Staphylococcus aureus Isolated by Allele Replacement. Infect. Immun. 1987, 55, 3103–3110. [Google Scholar] [PubMed]
- Wilke, G.A.; Wardenburg, J.B. Role of a disintegrin and metalloprotease 10 in Staphylococcus aureus α-hemolysin-mediated cellular injury. Proc. Natl. Acad. Sci. USA 2010, 107, 13473–13478. [Google Scholar] [CrossRef] [PubMed]
- Bhakdi, S.; Tranum-Jensen, J. Alpha-toxin of Staphylococcus aureus. Microbiol. Rev. 1991, 55, 733–751. [Google Scholar] [PubMed]
- Parker, M.W.; Feil, S.C. Pore-forming protein toxins: From structure to function. Prog. Biophys. Mol. Biol. 2005, 88, 91–142. [Google Scholar] [CrossRef] [PubMed]
- Dajcs, J.J.; Austin, M.S.; Sloop, G.D.; Moreau, J.M.; Hume, E.B.H.; Thompson, H.W.; McAleese, F.M.; Foster, T.J.; O’Callaghan, R.J. Corneal Pathogenesis of Staphylococcus aureus Strain Newman. Investig. Ophthalmol. Vis. Sci. 2002, 43, 1109–1115. [Google Scholar]
- Girgis, D.O.; Sloop, G.D.; Reed, J.M.; O’Callaghan, R.J. Effects of Toxin Production in a Murine Model of Staphylococcus aureus Keratitis. Investig. Ophthalmol. Vis. Sci. 2005, 46, 2064–2070. [Google Scholar] [CrossRef] [PubMed]
- Putra, I.; Rabiee, B.; Anwar, K.N.; Gidfar, S.; Shen, X.; Babalooee, M.; Ghassemi, M.; Afsharkhamseh, N.; Bakhsh, S.; Missiakas, D.; et al. Staphylococcus aureus Alpha-Hemolysin Impairs Corneal Epithelial Wound Healing and Promotes Intracellular Bacterial Invasion. Exp. Eye Res. 2019, 181, 263–270. [Google Scholar] [CrossRef]
- Lee, K.; Lee, H.; Kim, M. Two Cases of Corneal Ulcer due to Methicillin-Resistant Staphylococcus aureus in High Risk Groups. Korean J. Ophthalmol. 2010, 24, 240–244. [Google Scholar] [CrossRef] [PubMed]
- Marquart, M.E. Animal Models of Bacterial Keratitis. J. Biomed. Biotechnol. 2011, 2011, 1–12. [Google Scholar] [CrossRef] [Green Version]
- O’Callaghan, R.J.; McCormick, C.C.; Caballero, A.R.; Marquart, M.E.; Gatlin, H.P.; Fratkin, J.D. Age-Related Differences in Rabbits during Experimental Staphylococcus aureus Keratitis. Investig. Ophthalmol. Vis. Sci. 2007, 48, 5125–5131. [Google Scholar] [CrossRef]
- Moreau, J.M.; Sloop, G.D.; Engel, L.S.; Hill, J.M.; O’Callaghan, J. Histopathological studies of staphylococcal alpha-toxin: Effects on rabbit corneas. Curr. Eye Res. 1997, 16, 1221–1228. [Google Scholar] [CrossRef] [PubMed]
- Kumar, A.; Kumar, A. Role of Staphylococcus aureus Virulence Factors in Inducing Inflammation and Vascular Permeability in a Mouse Model of Bacterial Endophthalmitis. PLoS ONE 2015, 10, e0128423. [Google Scholar] [CrossRef]
- Goerke, C.; Fluckiger, U.; Steinhuber, A.; Zimmerli, W.; Wolz, C. Impact of the regulatory loci agr, sarA, and sae of Staphylococcus aureus on the induction of α-toxin during device-related infection resolved by direct quantitative transcript analysis. Mol. Microb. 2001, 40, 1439–1447. [Google Scholar] [CrossRef]
- Xiong, Y.Q.; Willard, J.; Yeaman, M.R.; Cheung, A.L.; Bayer, A.S. Regulation of Staphylococus aureus α-Toxin Gene (hla) Expression by agr, sarA, and sae In Vitro and in Experimental Infective Endocarditis. J. Infect. Dis. 2006, 194, 1267–1275. [Google Scholar] [CrossRef] [PubMed]
- Booth, M.C.; Cheung, A.L.; Hatter, K.L.; Jett, B.D.; Callegan, M.C.; Gilmore, M.S. Staphylococcal Accessory Regulator (sar) in Conjunction with agr Contributes to Staphylococcus aureus Virulence in Endophthalmitis. Infect. Immun. 1997, 65, 1550–1556. [Google Scholar] [PubMed]
- Horsburgh, M.J.; Aish, J.L.; White, I.J.; Shaw, L.; Lithgow, J.K.; Foster, S.J. σB Modulates Virulence Determinant Expression and Stress Resistance: Characterization of a Functional rsbU Strain Derived from Staphylococcus aureus 8325-4. J. Bacteriol. 2002, 184, 5457–5467. [Google Scholar] [CrossRef] [PubMed]
- Hu, C.M.; Fang, R.H.; Copp, J.; Luk, B.T.; Zhang, L. A biomimetic nanosponge that absorbs pore-forming toxins. Nat. Nanotechnol. 2013, 8, 336–340. [Google Scholar] [CrossRef] [Green Version]
- Coburn, P.S.; Miller, F.C.; LaGrow, A.L.; Land, C.; Mursalin, H.; Livingston, E.; Amayem, O.; Chen, Y.; Gao, L.; Callegan, M.C. Disarming Pore-Forming Toxins with Biomimetic Nanosponges in Intraocular Infections. mSphere 2019, 4, e00262-19. [Google Scholar] [CrossRef] [Green Version]
- Gravet, A.; Colin, D.A.; Keller, R.; Giradot, R.; Monteil, H.; Prevost, G. Characterization of a novel structural member, LukE-LukD, of the bi-component staphylococcal leucotoxins family. FEBS Lett. 1998, 136, 202–208. [Google Scholar] [CrossRef]
- Spaan, A.N.; Schiepers, A.; de Haas, C.J.C.; van Hooijdonk, D.; Badiou, C.; Contamin, H.; Vandenesch, F.; Lina, G.; Gerard, N.P.; Gerard, C.; et al. Differential Interactions of the Staphylococcal Toxins Panton-Valentine Leukocidin and γ-Hemolysin CB with Human C5a Receptors. J. Immunol. 2015, 195, 1034–1043. [Google Scholar] [CrossRef]
- Spaan, A.N.; van Strijp, J.A.G.; Torres, V.J. Leukocidins: Staphylococcal bi-component pore-forming toxins find their receptors. Nat. Rev. 2017, 15, 435–447. [Google Scholar] [CrossRef] [PubMed]
- Spaan, A.N.; Henry, T.; van Rooijen, W.; Perret, M.; Badiou, C.; Aerts, P.C.; Kemmink, J.; de Haas, C.; van Kessel, K.; Vandenesch, F.; et al. The Staphylocccal Toxin Panton-Valentine Leukocidin Targets Human C5a Receptors. Cell Host Microbe 2013, 13, 584–594. [Google Scholar] [CrossRef] [PubMed]
- Zaidi, T.; Zaidi, T.; Yoong, P.; Pier, G.B. Staphylococcus aureus Corneal Infections: Effect of the Panton-Valentine Leukocidin (PVL) and Antibody to PVL on Virulence and Pathology. Investig. Ophthalmol. Vis. Sci. 2013, 54, 4430–4438. [Google Scholar] [CrossRef] [PubMed]
- Siqueira, J.A.; Speeg-Schatz, C.; Freitas, F.I.S.; Sahel, J.; Monteil, H.; Prevost, G. Channel-forming leucotoxins from Staphylococcus aureus cause severe inflammatory reactions in a rabbit eye model. J. Med. Microbiol. 1997, 46, 486–494. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.L.; Heitz, P.; Roux, M.; Keller, D.; Bourcier, T.; Sauer, A.; Pervost, G.; Gaucher, D. Panton-Valentine Leukocidin Colocalizes with Retinal Ganglion and Amacrine Cells and Acivates Glial Reactions and Microglial Apoptosis. Sci. Rep. 2018, 8, 2953. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.L.; Roux, M.J.; Picaud, S.; Keller, D.; Sauer, A.; Heitz, P.; Pervost, G.; Gaucher, D. Panton-Valentine Leucocidin Proves Direct Neuronal Targeting and Its Early Neuronal and Glial Impacts a Rabbit Retinal Explant Model. Toxins 2018, 10, 455. [Google Scholar] [CrossRef] [PubMed]
- Peterson, J.C.; Durkee, H.; Miller, D.; Maestre-Mesa, J.; Arboleda, A.; Aguilar, M.C.; Relhan, N.; Flynn, H.W.; Amescua, G.; Parel, J.; et al. Molecular epidemiology and resistance profiles among healthcare- and community-associated Staphylococcus aureus keratitis isolates. Infect. Drug Resist. 2019, 12, 831–843. [Google Scholar] [CrossRef] [PubMed]
- von Eiff, C.; Friedrich, A.W.; Peters, G.; Becker, K. Prevalence of genes encoding for members of the staphylococcal leukotoxin family among clinical isolates of Staphylococcus aureus. Diagn. Microbiol. Infect. Dis. 2004, 49, 157–162. [Google Scholar] [CrossRef]
- Malachowa, N.; Whitney, A.R.; Kobayashi, S.D.; Sturdevant, D.E.; Kennedy, A.D.; Braughton, K.R.; Shabb, D.W.; Diep, B.A.; Chambers, H.F.; Otto, M.; et al. Global Changes in Staphylococcus aureus Gene Expression in Human Blood. PLoS ONE 2011, 6, e18617. [Google Scholar] [CrossRef]
- Voyich, J.M.; Braughton, K.R.; Sturdevant, D.E.; Whitney, A.R.; Said-Salim, B.; Porcella, S.F.; Long, R.D.; Dorward, D.W.; Gardener, D.J.; Kreiswirth, B.N.; et al. Insights into Mechanisms Used by Staphylococcus aureus to Avoid Destruction by Human Neutrophils. J. Immunol. 2005, 175, 3907–3919. [Google Scholar] [CrossRef]
- Spaan, A.N.; Vrieling, M.; Wallet, P.; Badiou, C.; Reyes-Robles, T.; Ohneck, E.A.; Benito, Y.; de Haas, C.; Day, C.J.; Jennings, M.P.; et al. The staphylococcal toxins γ-haemolysin AB and CB differentially target phagocytes by employing specific chemokine receptors. Nat. Commun. 2014, 5, 5438. [Google Scholar] [CrossRef] [PubMed]
- Supersac, G.; Piemont, Y.; Kubina, M.; Prevost, G.; Foster, T.J. Assessment of the role of γ-toxin in experimental endophthalmitis using a hlg-deficient mutant of Staphylococcus aureus. Microb. Pathog. 1998, 24, 241–251. [Google Scholar] [CrossRef] [PubMed]
- Dassy, B.; Hogan, T.; Foster, T.J.; Fournier, J. Involvement of the accessory gene regulator (agr) in expression of the type 5 capsular polysaccharide by Staphylococcus aureus. Microbiology 1993, 139, 1301–1306. [Google Scholar] [CrossRef] [PubMed]
- Baba, T.; Bae, T.; Schneewind, O.; Takeuchi, F.; Hiramatsu, K. Genome Sequence of Staphylococcus aureus Strain Newman and Comparative Analysis of Staphylococcal Genomes: Polymorphism and Evolution of Two Major Pathogenicity Islands. J. Bacteriol. 2008, 190, 300–310. [Google Scholar] [CrossRef] [PubMed]
- Periasamy, S.; Chatterjee, S.S.; Cheung, G.Y.C.; Otto, M. Phenol-soluble modulins in staphylococci What are they originally for? Commun. Integr. Biol. 2012, 5, 275–276. [Google Scholar] [CrossRef] [PubMed]
- Wang, R.; Braughton, K.R.; Kretschmer, D.; Bach, T.L.; Queck, S.Y.; Li, M.; Kennedy, A.D.; Dorward, D.W.; Kelbanoff, S.J.; Peschel, A.; et al. Identification of novel cytolytic peptides as key virulence determinates for community-associated MRSA. Nat. Med. 2007, 13, 1510–1514. [Google Scholar] [CrossRef]
- Baek, K.T.; Frees, D.; Renzoni, A.; Barras, C.; Rodriguez, N.; Manzano, N.; Kelley, W.L. Genetic Variation in the Staphylococcus aureus 8325 Strain Lineage Revealed by Whole-Genome Sequencing. PLoS ONE 2013, 8, e77122. [Google Scholar] [CrossRef]
- Vandenesch, F.; Lina, G.; Henry, T. Staphylococcus aureus hemolysins, bi-component leukocidins, and cytolytic peptides: A redundant arsenal of membrane-damaging virulence factors? Front. Cell. Infect. Microbiol. 2012, 2, 12. [Google Scholar] [CrossRef]
- Hanzelmann, D.; Joo, H.; Franz-Wachel, M.; Hertlein, T.; Stevanovic, S.; Macek, B.; Wolz, C.; Gotz, F.; Otto, M.; Krestschmer, D.; et al. Toll-like receptor 2 activation depends on lipopeptide shedding by bacterial surfactants. Nat. Commun. 2016, 7, 12304. [Google Scholar] [CrossRef]
- Kretschmer, D.; Gleske, A.; Rautenberg, M.; Wang, R.; Koberle, M.; Bohn, E.; Schoneberg, T.; Rabiet, M.; Boulay, F.; Klebanoff, S.J.; et al. Human Formyl Peptide Receptor 2 Senses Highly Pathogenic Staphylococcus aureus. Cell Host Microbe 2010, 7, 463–473. [Google Scholar] [CrossRef] [Green Version]
- Weiss, E.; Hanzelmann, D.; Fehlhaber, B.; Klos, A.; von Leowenich, F.; Liese, J.; Peschel, A.; Kretschmer, D. Formyl-peptide receptor 2 governs leukocyte influx in local Staphylococcus aureus infections. FASEB J. 2018, 32, 26–36. [Google Scholar] [CrossRef] [PubMed]
- Otto, M. Staphylococcus aureus toxins. Curr. Opin. Microbiol. 2014, 17, 32–37. [Google Scholar] [CrossRef] [PubMed]
- Freer, J.H.; Arbuthnott, J.P. Toxins of Staphylococcus aureus. Pharmacol. Ther. 1983, 19, 55–106. [Google Scholar] [CrossRef]
- Aarestrup, F.M.; Larsen, H.D.; Eriksen, N.H.R.; Elsberg, C.S.; Jensen, N.E. Frequency of α-and β-haemolysin in Staphylococcus aureus of bovine and human origin. Apmis 1999, 107, 425–430. [Google Scholar] [CrossRef] [PubMed]
- van Wamel, W.; Rooijakkers, S.; Ruyken, M.; van Kessel, K.; van Strijp, J. The Innate Immune Modulators Staphylococcal Complement Inhibitor and Chemotaxis Inhibitory Protein of Staphylococcus aureus Are Located on β-Hemolysin-Converting Bacteriophages. J. Bacteriol. 2006, 188, 1310–1315. [Google Scholar] [CrossRef] [PubMed]
- Salgado-Pabon, W.; Herrera, A.; Vu, B.G.; Stach, C.S.; Merriman, J.A.; Spaulding, A.R.; Schlievert, P.M. Staphylococcus aureus β-toxin Production is Common in Strains with the β-toxin Gene Inactivation by Bacteriophage. J. Infect. Dis. 2014, 210, 784–792. [Google Scholar] [CrossRef] [PubMed]
- Walev, I.; Weller, U.; Strauch, S.; Foster, T.; Bhakdi, S. Selective Killing of Human Monocytes and Cytokine Release Provoked by Sphingomyelinase (Beta-Toxin) of Staphylococcus aureus. Infect. Immun. 1996, 64, 2974–2979. [Google Scholar]
- Mestas, J.; Hughes, C.C.W. Of Mice and Not Men: Differences between Mouse and Human Immunology. J. Immunol. 2004, 172, 2731–2738. [Google Scholar] [CrossRef] [Green Version]
- Seok, J.; Warren, H.S.; Cuenca, A.G.; Mindrinos, M.N.; Baker, H.V.; Xu, W.; Richards, D.R.; McDonald-Smith, G.P.; Gao, H.; Hennessy, L.; et al. Genomic responses in mouse models poorly mimic human inflammatory diseases. Proc. Natl. Acad. Sci. USA 2013, 110, 3507–3512. [Google Scholar] [CrossRef] [Green Version]
- Warren, H.S.; Tompkins, R.G.; Modawer, L.L.; Seok, J.; Xu, W.; Mindrinos, M.N.; Maier, R.V.; Xiao, W.; Davis, R.W. Mice are not men. Proc. Natl. Acad. Sci. USA 2015, 112, E345. [Google Scholar] [CrossRef]
- Astley, R.A.; Coburn, P.S.; Parkunan, S.M.; Callegan, M.C. Modeling intraocular bacterial infections. Prog. Retina Eye Res. 2016, 54, 30–48. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kwon, B.; Hazlett, L.D. Association of CD4+ T Cell-Dependent Keratitis with Genetic Susceptibility to Pseudomonas aeruginosa Ocular Infection. J. Immunol. 1997, 159, 6283–6290. [Google Scholar] [PubMed]
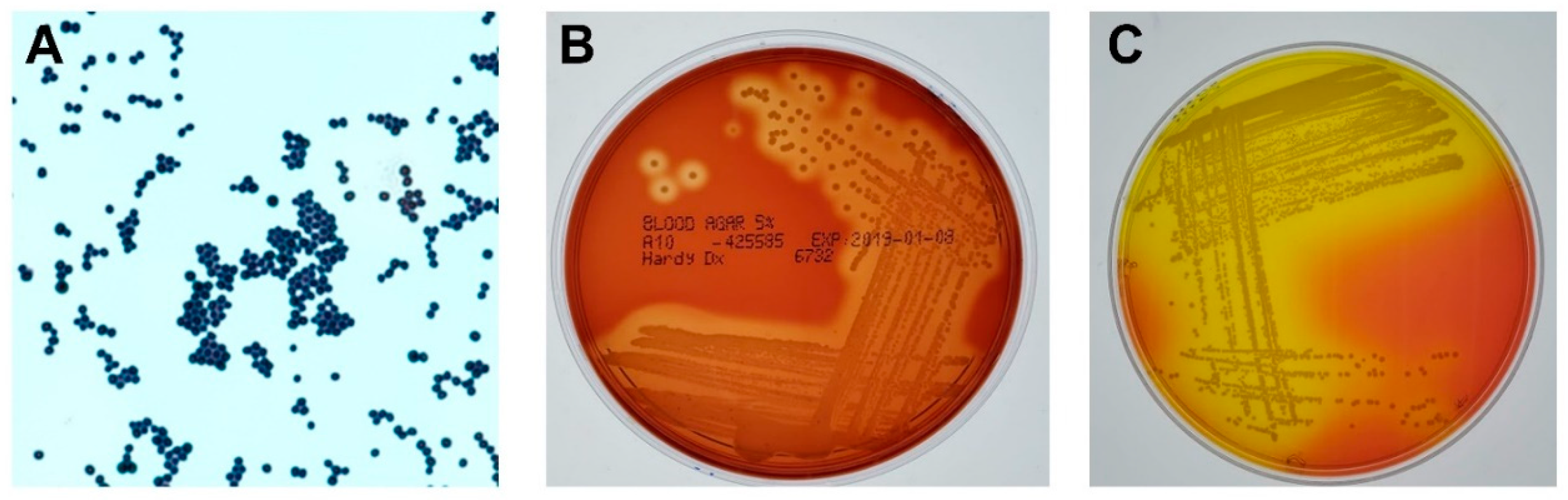
Figure 1.
Phenotypes of Staphylococcus aureus (S. aureus). (A) Gram stain of S. aureus. Magnification, 100×. (B) S. aureus grown on 5% sheep blood agar overnight at 37 °C. Note the characteristic zones of β-hemolysis surrounding each colony. (C) S. aureus grown on mannitol salt agar overnight at 37 °C. S. aureus, unlike other staphylococci, ferments mannitol, resulting in acid production and the classic yellow halo within the deep pink agar.
Figure 1.
Phenotypes of Staphylococcus aureus (S. aureus). (A) Gram stain of S. aureus. Magnification, 100×. (B) S. aureus grown on 5% sheep blood agar overnight at 37 °C. Note the characteristic zones of β-hemolysis surrounding each colony. (C) S. aureus grown on mannitol salt agar overnight at 37 °C. S. aureus, unlike other staphylococci, ferments mannitol, resulting in acid production and the classic yellow halo within the deep pink agar.

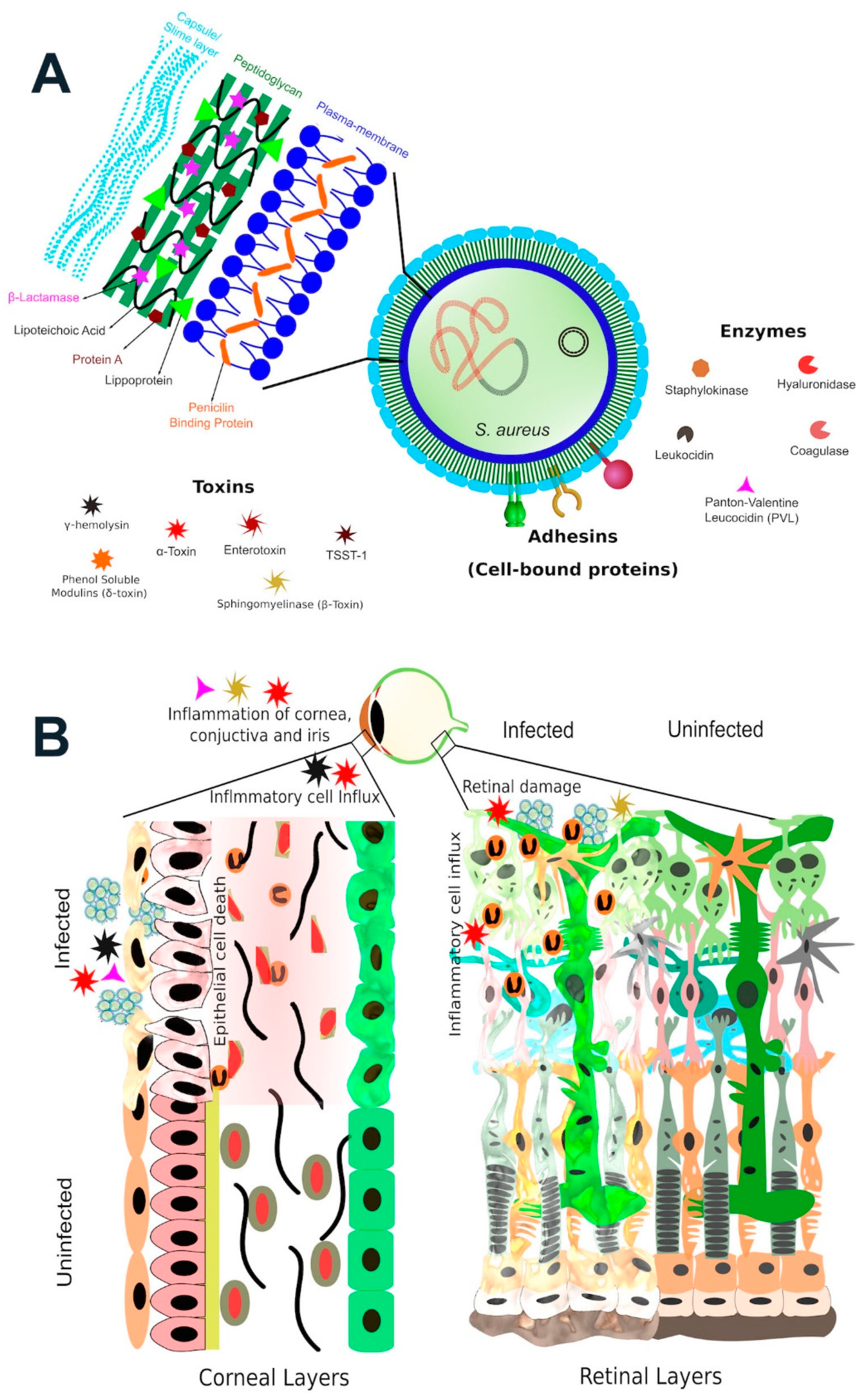
Figure 2.
S. aureus virulence factors contribute to the pathogenesis of ocular infections. (A) S. aureus contains numerous virulence factors in its armamentarium which assists in antibiotic resistance, adherence to tissue, interactions with innate immunity, and spread of infection. (B) S. aureus toxins which have been analyzed in experimental infection models include α-toxin, β-toxin, γ-toxin, and Panton–Valentine leukocidin (PVL). The contribution of these toxins to ocular infections is strain-and model-dependent, as discussed below.
Figure 2.
S. aureus virulence factors contribute to the pathogenesis of ocular infections. (A) S. aureus contains numerous virulence factors in its armamentarium which assists in antibiotic resistance, adherence to tissue, interactions with innate immunity, and spread of infection. (B) S. aureus toxins which have been analyzed in experimental infection models include α-toxin, β-toxin, γ-toxin, and Panton–Valentine leukocidin (PVL). The contribution of these toxins to ocular infections is strain-and model-dependent, as discussed below.

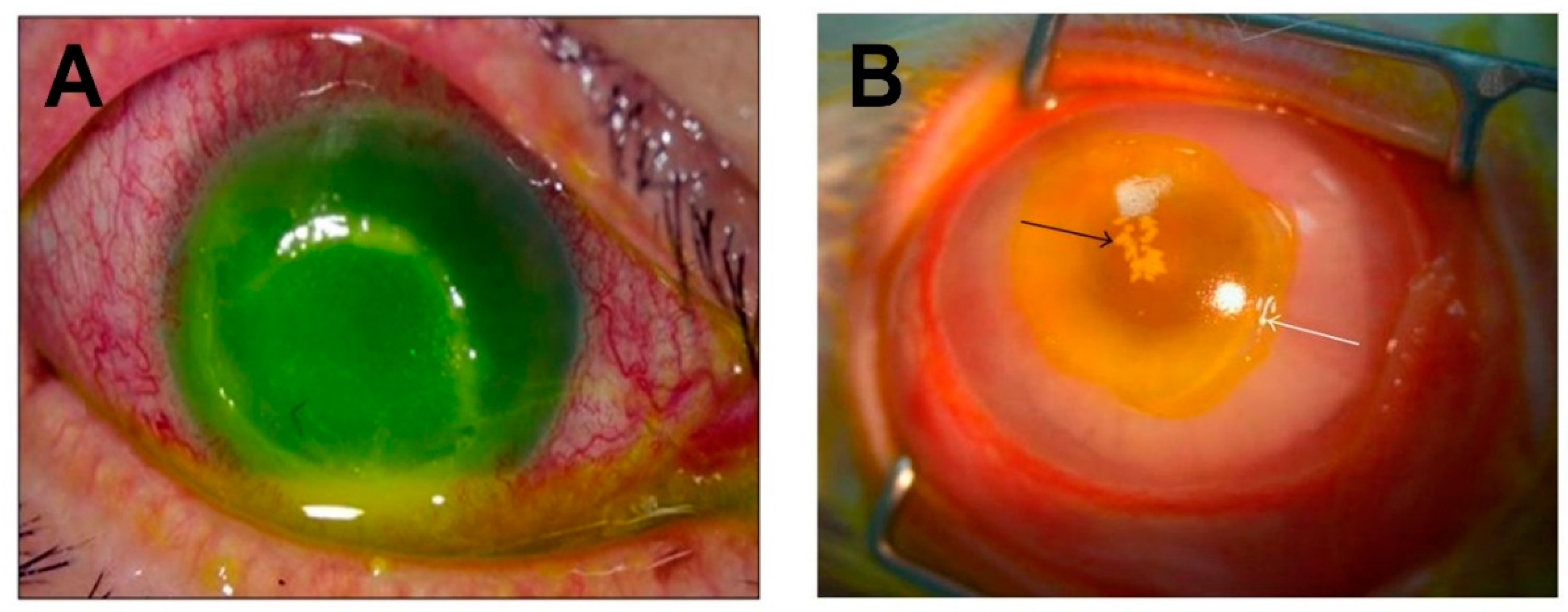
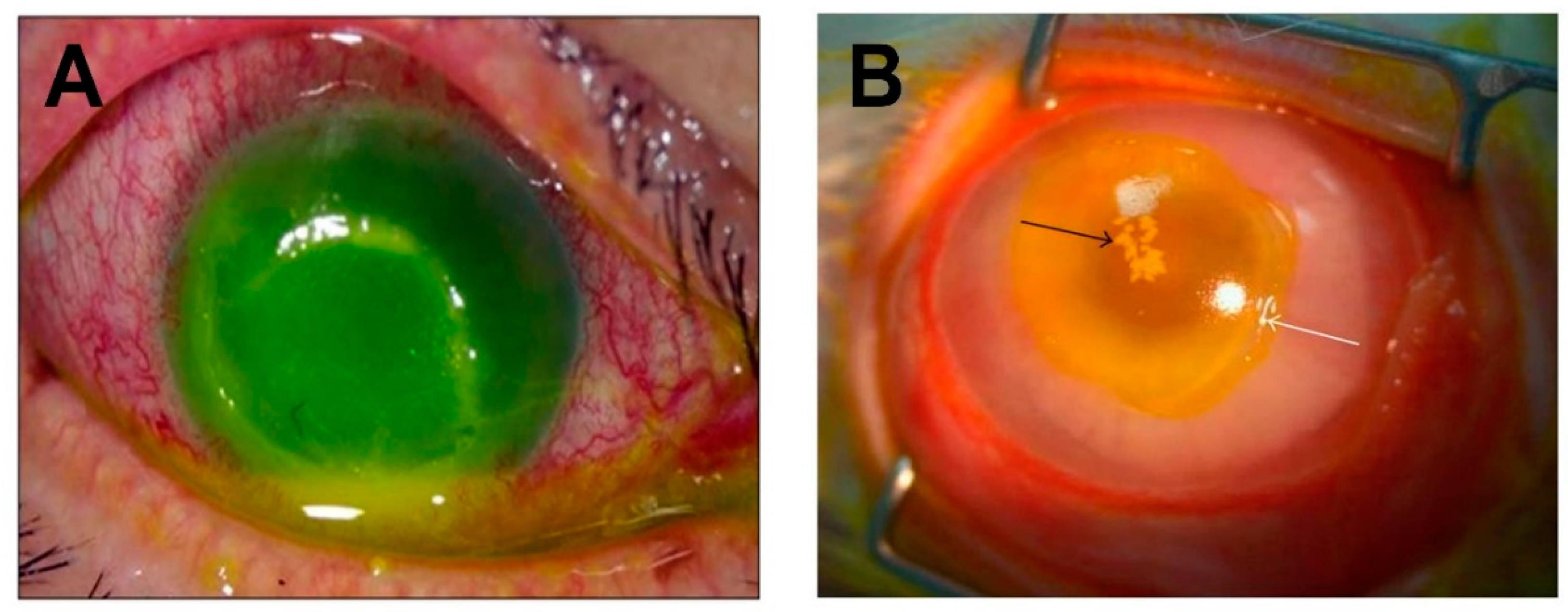
Figure 3.
S. aureus-induced corneal ulcers in humans and rabbits. (A) A corneal ulcer with superficial stromal infiltration and anterior chamber hypopyon in a human eye infected with MRSA. Copyright © Lee et al., 2010 [82]. (B) S. aureus keratitis in a rabbit eye. Black arrow indicates inflammatory cell infiltration and staphylococcal microcolonies formed in the corneal stroma. White arrow indicates the edge of the epithelial erosion stained with fluorescein. Copyright © Marquart et al., 2011 [83]. These are open access articles distributed under the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited.
Figure 3.
S. aureus-induced corneal ulcers in humans and rabbits. (A) A corneal ulcer with superficial stromal infiltration and anterior chamber hypopyon in a human eye infected with MRSA. Copyright © Lee et al., 2010 [82]. (B) S. aureus keratitis in a rabbit eye. Black arrow indicates inflammatory cell infiltration and staphylococcal microcolonies formed in the corneal stroma. White arrow indicates the edge of the epithelial erosion stained with fluorescein. Copyright © Marquart et al., 2011 [83]. These are open access articles distributed under the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited.

{kind=link}
{kind=link}
{kind=link}
Table 1.
Evaluation of toxin genes in a random sampling of Staphylococcus aureus isolated from ocular infections at the Dean A. McGee Eye Institute (USA) from April 2011 to February 2018.
Table 1.
Evaluation of toxin genes in a random sampling of Staphylococcus aureus isolated from ocular infections at the Dean A. McGee Eye Institute (USA) from April 2011 to February 2018.
| Gene | MSSA (7) | MRSA (9) | All (16) |
|---|---|---|---|
| hla | 6 (86%) | 9 (100%) | 15 (94%) |
| hlb | 4 (57%) | 5 (56%) | 8 (50%) |
| mecA | 0 | 9 | 9 |
Numbers in parentheses in the top row indicate total number of isolates. Percentages in the table indicate percent positive of the column total by PCR, using primers described previously [64]. PCR for mecA was conducted to confirm clinical laboratory results. MSSA, methicillin-sensitive Staphylococcus aureus; MRSA, methicillin-resistant Staphylococcus aureus.
Table 2.
Studies analyzing the role of S. aureus toxins in ocular infections using staphylococcal strains specifically deficient in the toxin of interest.
Table 2.
Studies analyzing the role of S. aureus toxins in ocular infections using staphylococcal strains specifically deficient in the toxin of interest.
| Toxin Absent | S. Aureus Strain | Ocular Infection Model | Result | References |
|---|---|---|---|---|
| α-toxin | 8325-4 | Keratitis rabbit | 🡫 Slit lamp scores 🡫 Myeloperoxidase activity 🡫 Corneal erosions | [67] |
| α-toxin | 8325-4 | Keratitis rabbit | 🡫 Slit lamp scores 🡫 Inflammation | [68] |
| β-toxin | 8325-4 | Keratitis rabbit | 🡫 Scleral edema | [68] |
| γ-toxin | Newman | Endophthalmitis rabbit | 🡫 Lid inflammation | [106] |
| α-toxin | Newman | Keratitis rabbit | 🡫 Slit lamp scores 🡫 Inflammation | [81] |
| γ-toxin | Newman | Keratitis rabbit | 🡫 Slit lamp scores 🡫 Corneal PMN | [81] |
| α-toxin | 8325-4 | Endophthalmitis rabbit | 🡫 Retinal damage | [52] |
| β-toxin | 8325-4 | Endophthalmitis rabbit | 🡫 Retinal damage | [52] |
| γ-toxin | 8325-4 | Endophthalmitis rabbit | No change | [52] |
| α-toxin | 8325-4 | Keratitis mouse | More severe in aged mice | [80] |
| PVL | Various USA 300 and 400 strains | Keratitis mouse | Enhanced virulence in a subset of MRSA strains | [97] |
| α-toxin | JE2 | Keratitis mouse | 🡡 Corneal healing | [81] |
Table 3.
Summary of studies analyzing the role of S. aureus toxins on ocular tissue using purified toxins applied directly to the eye.
Table 3.
Summary of studies analyzing the role of S. aureus toxins on ocular tissue using purified toxins applied directly to the eye.
| Toxin | Inoculation Method | Ocular Model | Result | References |
|---|---|---|---|---|
| α-toxin | Topical | Rabbit cornea | Inflammation of conjunctiva and iris | [68] |
| α-toxin | Injection | Rabbit cornea | Inflammation of cornea and iris, corneal epithelial defect | [68] |
| β-toxin | Topical | Rabbit cornea | Inflammation of conjunctiva | [68] |
| β-toxin | Injection | Rabbit cornea | Scleral edema | [68] |
| γ-toxin | Injection | Rabbit cornea | Acute inflammatory reactions | [98] |
| PVL | Injection | Rabbit cornea | Acute inflammatory reactions | [98] |
| α-toxin | Injection | Rabbit cornea | ↑SLE score, edema, epithelia cell death | [85] |
| α-toxin | Topical | Mouse cornea | Corneal pathology more severe in aged mice | [80] |
| α-toxin | Injection | Rabbit cornea | Corneal pathology more severe in young rabbits | [81] |
| α-toxin | Injection | Mouse Vitreous | Mild retinal damage, no reduction in retinal function | [89] |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Astley, R.; Miller, F.C.; Mursalin, M.H.; Coburn, P.S.; Callegan, M.C. An Eye on Staphylococcus aureus Toxins: Roles in Ocular Damage and Inflammation. Toxins 2019, 11, 356. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins11060356
AMA Style
Astley R, Miller FC, Mursalin MH, Coburn PS, Callegan MC. An Eye on Staphylococcus aureus Toxins: Roles in Ocular Damage and Inflammation. Toxins. 2019; 11(6):356. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins11060356
Chicago/Turabian StyleAstley, Roger, Frederick C. Miller, Md Huzzatul Mursalin, Phillip S. Coburn, and Michelle C. Callegan. 2019. "An Eye on Staphylococcus aureus Toxins: Roles in Ocular Damage and Inflammation" Toxins 11, no. 6: 356. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins11060356
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.