The Urgent Need to Develop Novel Strategies for the Diagnosis and Treatment of Snakebites

 , and
, and 
Abstract
:1. Introduction
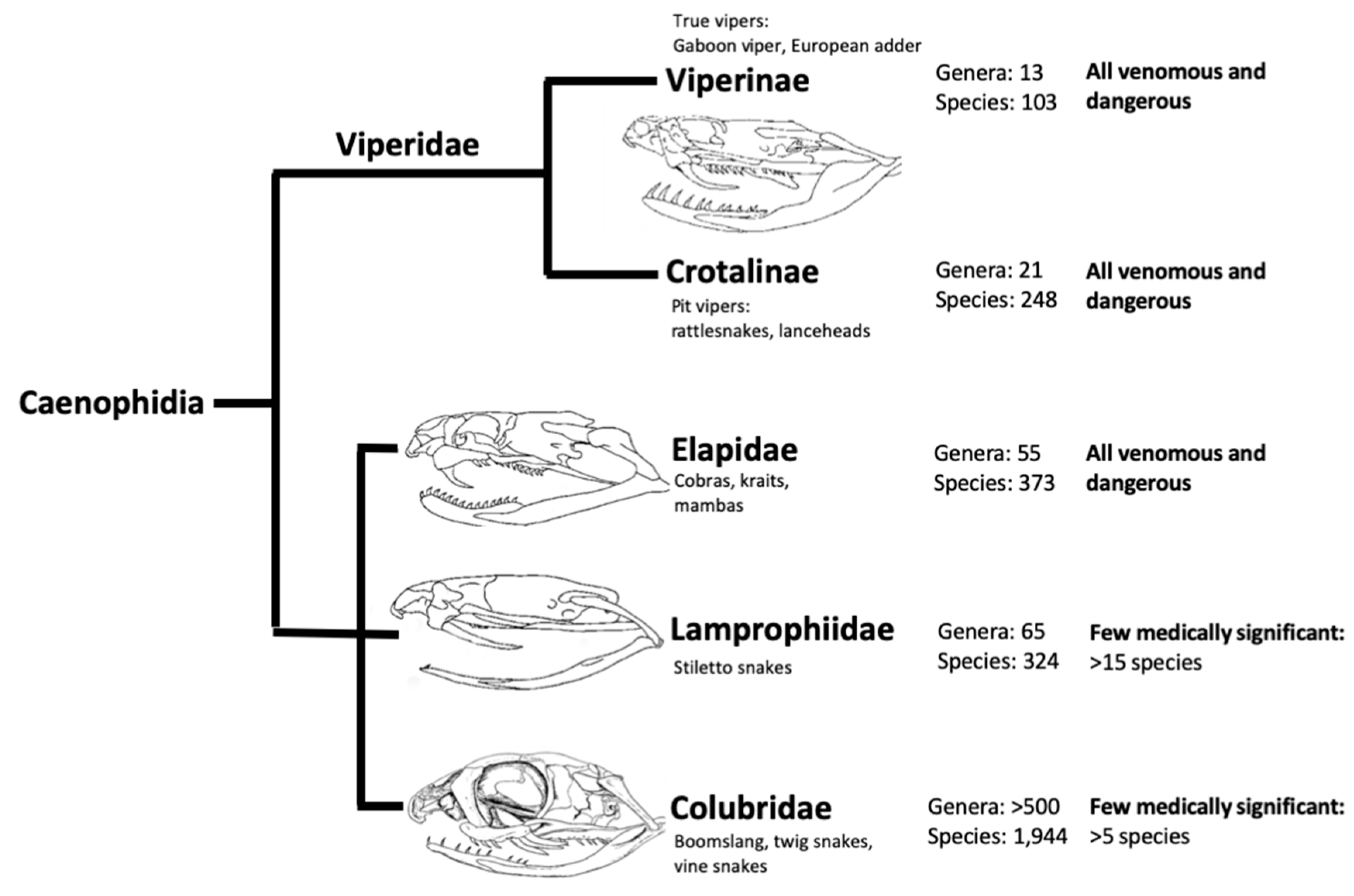
2. The Complexity of Snake Venoms
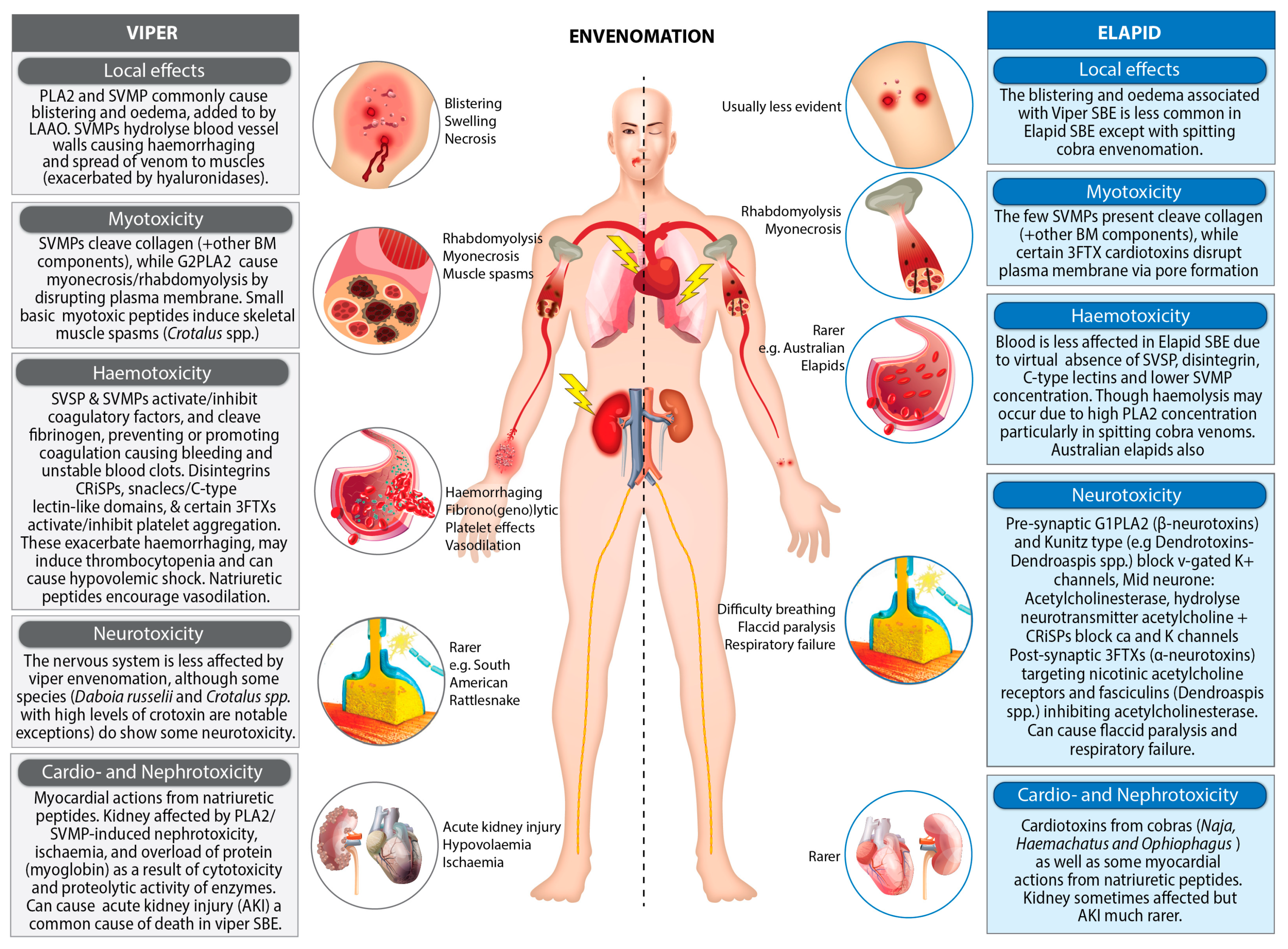
2.1. Enzymatic Components
2.2. Non-Enzymatic Components
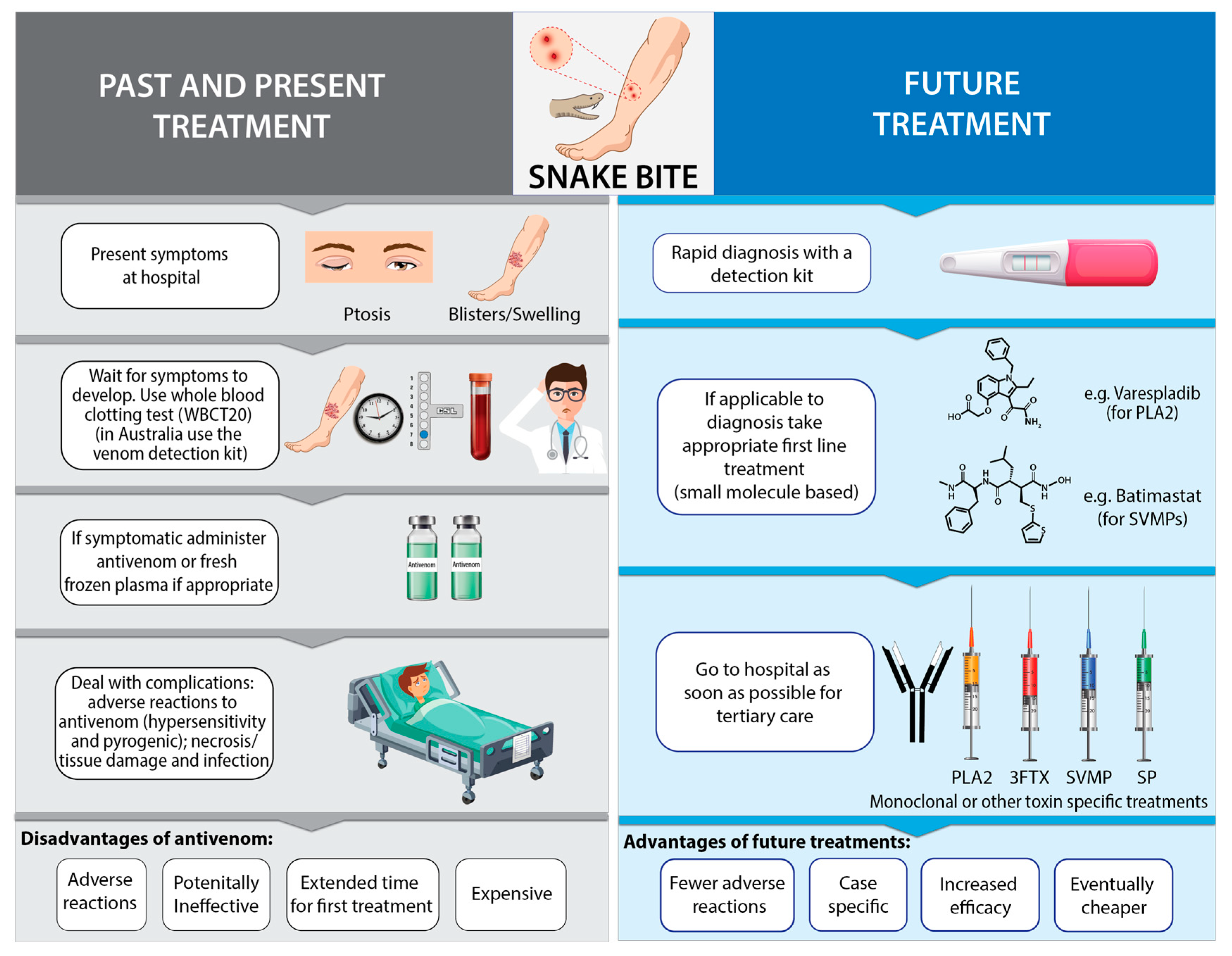
3. Antivenom (Anti-Snake Venom/Venin/ASV) and Its Associated Problems
3.1. Reproducibility Issues Associated with Antivenom Production
3.2. Relative Instability of Antivenom
3.3. Adverse Reactions to Antivenom
3.4. Expense of Antivenom
4. Diagnosis of Snakebites
- Where were you bitten? Leading to examination of bite site.
- When were you bitten? In recent bites symptoms may be absent.
- What were you doing when you were bitten? Activity may be diagnostical.
- Where is the snake that bit you or what did it look like? Actual snake or photo can aid in diagnosis.
- How are you feeling now? Check for further symptoms of envenoming.
4.1. Clinical Diagnosis
4.2. Venom Detection Kits
4.3. Improving the Diagnosis of SBE
5. Future Treatment Approaches for SBE
5.1. Small Molecular Therapeutics (SMTs)
5.2. Protein, Peptide and Oligomer Based Technologies
6. Diagnostics Feeding into Treatment
7. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Harrison, R.A.; Hargreaves, A.; Wagstaff, S.C.; Faragher, B.; Lalloo, D.G. Snake envenoming: A disease of poverty. PLoS Negl. Trop. Dis. 2009, 3, e569. [Google Scholar] [CrossRef] [PubMed]
- WHO. Neglected Tropical Diseases. Available online: http://www.who.int/neglected_diseases/diseases/en/ (accessed on 20 June 2019).
- Williams, D.; Gutiérrez, J.M.; Harrison, R.; Warrell, D.A.; White, J.; Winkel, K.D.; Gopalakrishnakone, P. The global snake bite initiative: An antidote for snake bite. Lancet 2010, 375, 89–91. [Google Scholar] [CrossRef]
- Hernandez, H.W.; Soeung, M.; Zorn, K.M.; Ashoura, N.; Mottin, M.; Andrade, C.H.; Caffrey, C.R.; de Siqueira-Neto, J.L.; Ekins, S. High throughput and computational repurposing for neglected diseases. Pharm. Res. 2019, 36, 27. [Google Scholar] [CrossRef] [PubMed]
- Welton, R.E.; Liew, D.; Braitberg, G. Incidence of fatal snake bite in Australia: A coronial based retrospective study (2000–2016). Toxicon 2017, 131, 11–15. [Google Scholar] [CrossRef] [PubMed]
- Chippaux, J.P. Snake-bites: Appraisal of the global situation. Bull. World Health Org. 1998, 76, 515. [Google Scholar] [PubMed]
- Kasturiratne, A.; Wickremasinghe, A.R.; de Silva, N.; Gunawardena, N.K.; Pathmeswaran, A.; Premaratna, R.; Savioli, L.; Lalloo, D.G.; de Silva, H.J. The global burden of snakebite: A literature analysis and modelling based on regional estimates of envenoming and deaths. PLoS Med. 2008, 5, e218. [Google Scholar] [CrossRef]
- Mohapatra, B.; Warrell, D.A.; Suraweera, W.; Bhatia, P.; Dhingra, N.; Jotkar, R.M.; Rodriguez, P.S.; Mishra, K.; Whitaker, R.; Jha, P. Snakebite mortality in India: A nationally representative mortality survey. PLoS Negl. Trop. Dis. 2011, 5, e1018. [Google Scholar] [CrossRef]
- Ralph, R.; Sharma, S.K.; Faiz, M.A.; Ribeiro, I.; Rijal, S.; Chappuis, F.; Kuch, U. The timing is right to end snakebite deaths in South Asia. Bmj 2019, 364, k5317. [Google Scholar] [CrossRef] [Green Version]
- Chippaux, J.-P. Incidence and mortality due to snakebite in the Americas. PLoS Negl. Trop. Dis. 2017, 11, e0005662. [Google Scholar] [CrossRef]
- Abubakar, S.B.; Habib, A.G.; Mathew, J. Amputation and disability following snakebite in Nigeria. Trop. Dr. 2010, 40, 114–116. [Google Scholar] [CrossRef]
- Williams, S.S.; Wijesinghe, C.A.; Jayamanne, S.F.; Buckley, N.A.; Dawson, A.H.; Lalloo, D.G.; de Silva, H.J. Delayed psychological morbidity associated with snakebite envenoming. PLoS Negl. Trop. Dis. 2011, 5, e1255. [Google Scholar] [CrossRef] [PubMed]
- Habib, A.G.; Kuznik, A.; Hamza, M.; Abdullahi, M.I.; Chedi, B.A.; Chippaux, J.-P.; Warrell, D.A. Snakebite is under appreciated: Appraisal of burden from West Africa. PLoS Negl. Trop. Dis. 2015, 9, e0004088. [Google Scholar] [CrossRef] [PubMed]
- O’neal, C.J.; Jobling, M.G.; Holmes, R.K.; Hol, W.G. Structural basis for the activation of cholera toxin by human ARF6-GTP. Science 2005, 309, 1093–1096. [Google Scholar] [CrossRef]
- Sorobetea, D.; Svensson-Frej, M.; Grencis, R. Immunity to gastrointestinal nematode infections. Mucosal Immunol. 2018, 11, 304. [Google Scholar] [CrossRef] [PubMed]
- Gutiérrez, J.M.; Calvete, J.J.; Habib, A.G.; Harrison, R.A.; Williams, D.J.; Warrell, D.A. Snakebite envenoming. Nat. Rev. Dis. Prim. 2017, 3, 17063. [Google Scholar] [CrossRef] [PubMed]
- Sack, D.A.; R Bradley Sack, S. Getting serious about cholera. N. Engl. J. Med. 2006, 355, 649. [Google Scholar] [CrossRef] [PubMed]
- Aagaard-Hansen, J.; Chaignat, C.L. Neglected tropical diseases: Equity and social determinants. In Equity, Social Determinants and Public Health Programmes; Blas, E., Kurup, A.S., Eds.; WHO Press: Geneva, Switzerland, 2010; p. 135. [Google Scholar]
- Hotez, P.J.; Molyneux, D.H.; Fenwick, A.; Kumaresan, J.; Sachs, S.E.; Sachs, J.D.; Savioli, L. Control of neglected tropical diseases. N. Engl. J. Med. 2007, 357, 1018–1027. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Naghavi, M.; Allen, C.; Barber, R.M.; Bhutta, Z.A.; Carter, A.; Casey, D.C.; Charlson, F.J.; Chen, A.Z.; Coates, M.M. Global, regional, and national life expectancy, all-cause mortality, and cause-specific mortality for 249 causes of death, 1980–2015: A systematic analysis for the global burden of disease study 2015. Lancet 2016, 388, 1459–1544. [Google Scholar] [CrossRef]
- Ali, M.; Lopez, A.L.; You, Y.; Kim, Y.E.; Sah, B.; Maskery, B.; Clemens, J. The global burden of cholera. Bull. World Health Org. 2012, 90, 209–218. [Google Scholar] [CrossRef]
- De Silva, N.R.; Brooker, S.; Hotez, P.J.; Montresor, A.; Engels, D.; Savioli, L. Soil-transmitted helminth infections: Updating the global picture. Trends Parasitol. 2003, 19, 547–551. [Google Scholar] [CrossRef]
- Bethony, J.; Brooker, S.; Albonico, M.; Geiger, S.M.; Loukas, A.; Diemert, D.; Hotez, P.J. Soil-transmitted helminth infections: Ascariasis, trichuriasis, and hookworm. Lancet 2006, 367, 1521–1532. [Google Scholar] [CrossRef]
- Ottesen, E.A.; Hooper, P.J.; Bradley, M.; Biswas, G. The global programme to eliminate lymphatic filariasis: Health impact after 8 years. PLoS Negl. Trop. Dis. 2008, 2, e317. [Google Scholar] [CrossRef] [PubMed]
- Ottesen, E.A. Lymphatic filariasis: Treatment, control and elimination. Adv. Parasitol. 2006, 61, 395–441. [Google Scholar] [PubMed]
- Basáñez, M.-G.; Sébastien, D.; Churcher, T.S.; Breitling, L.P.; Little, M.P.; Boussinesq, M. River blindness: A success story under threat? PLoS Med. 2006, 3, e371. [Google Scholar] [CrossRef] [PubMed]
- Smith, W.C.; van Brakel, W.; Gillis, T.; Saunderson, P.; Richardus, J.H. The Missing Millions: A Threat to the Elimination of Leprosy. PLoS Negl. Trop. Dis. 2015, 9, e0003658. [Google Scholar] [CrossRef]
- Hopkins, D.R.; Ruiz-Tiben, E.; Eberhard, M.L.; Weiss, A.; Withers, P.C., Jr.; Roy, S.L.; Sienko, D.G. Dracunculiasis eradication: Are we there yet? Am. J. Trop.Med. Hyg. 2018, 99, 388–395. [Google Scholar] [CrossRef] [PubMed]
- Modahl, C.M.; Saviola, A.J.; Mackessy, S.P. Venoms of colubrids. Venom Genom. Proteom. 2016, 2016, 51–79. [Google Scholar]
- Warrell, D.A. Snake bite. Lancet 2010, 375, 77–88. [Google Scholar] [CrossRef]
- Warrell, D.A. Venomous bites, stings, and poisoning: An update. Infect. Dis. Clin. 2019, 33, 17–38. [Google Scholar] [CrossRef]
- Tilbury, C.R.; Verster, J. A fatal bite from the burrowing asp atractaspis corpulenta (Hallowell 1854). Toxicon 2016, 118, 21–26. [Google Scholar] [CrossRef]
- Kardong, K.; Weinstein, S.; Smith, T.; Mackessy, S. Reptile venom glands: Form, function, and future. In Handbook of Venoms and Toxins of Reptiles; Mackessy, S.P., Ed.; CRC Press Inc.: London, UK, 2009; pp. 65–91. [Google Scholar]
- Chara, K.; Baccouche, N.; Turki, O.; Regaig, K.; Chaari, A.; Bahloul, M.; Bouaziz, M. A rare complication of viper envenomation: Cardiac failure. A case report. Med. Sante Trop. 2017, 27, 52–55. [Google Scholar] [CrossRef] [PubMed]
- El Zahran, T.; Kazzi, Z.; Chehadeh, A.A.; Sadek, R.; El Sayed, M.J. Snakebites in Lebanon: A descriptive study of snakebite victims treated at a tertiary care center in Beirut, Lebanon. J. Emerg. Trauma Shock 2018, 11, 119–124. [Google Scholar] [CrossRef] [PubMed]
- Vikrant, S.; Jaryal, A.; Parashar, A. Clinicopathological spectrum of snake bite-induced acute kidney injury from India. World J. Nephrol. 2017, 6, 150–161. [Google Scholar] [CrossRef] [PubMed]
- Silva, A.; Kuruppu, S.; Othman, I.; Goode, R.J.; Hodgson, W.C.; Isbister, G.K. Neurotoxicity in Sri Lankan Russell’s viper (Daboia russelii) envenoming is primarily due to U1-viperitoxin-Dr1a, a pre-synaptic neurotoxin. Neurotox. Res. 2017, 31, 11–19. [Google Scholar] [CrossRef] [PubMed]
- Segura, A.; Herrera, M.; Reta Mares, F.; Jaime, C.; Sanchez, A.; Vargas, M.; Villalta, M.; Gomez, A.; Gutierrez, J.M.; Leon, G. Proteomic, toxicological and immunogenic characterization of Mexican west-coast rattlesnake (Crotalus basiliscus) venom and its immunological relatedness with the venom of central American rattlesnake (Crotalus simus). J. Proteom. 2017, 158, 62–72. [Google Scholar] [CrossRef]
- Gillissen, A.; Theakston, R.D.G.; Barth, J.; May, B.; Krieg, M.; Warrell, D.A. Neurotoxicity, haemostatic disturbances and haemolytic anaemia after a bite by a Tunisian saw-scaled or carpet viper (Echis ‘pyramidum’-complex): Failure of antivenom treatment. Toxicon 1994, 32, 937–944. [Google Scholar] [CrossRef]
- Tan, C.H.; Tan, K.Y.; Yap, M.K.K.; Tan, N.H. Venomics of tropidolaemus wagleri, the sexually dimorphic temple pit viper: Unveiling a deeply conserved atypical toxin arsenal. Sci. Rep. 2017, 7, 43237. [Google Scholar] [CrossRef]
- Kalita, B.; Singh, S.; Patra, A.; Mukherjee, A.K. Quantitative proteomic analysis and antivenom study revealing that neurotoxic phospholipase A2 enzymes, the major toxin class of Russell’s viper venom from southern India, shows the least immuno-recognition and neutralization by commercial polyvalent antivenom. Int. J. Boil. Macromol. 2018, 118, 375–385. [Google Scholar]
- Maduwage, K.; Isbister, G.K. Current treatment for venom-induced consumption coagulopathy resulting from snakebite. PLoS Negl. Trop. Dis. 2014, 8, e3220. [Google Scholar] [CrossRef]
- Fry, B.G. Structure–function properties of venom components from Australian elapids. Toxicon 1999, 37, 11–32. [Google Scholar] [CrossRef]
- Reyes-Velasco, J.; Card, D.C.; Andrew, A.L.; Shaney, K.J.; Adams, R.H.; Schield, D.R.; Casewell, N.R.; Mackessy, S.P.; Castoe, T.A. Expression of venom gene homologs in diverse python tissues suggests a new model for the evolution of snake venom. Mol. Biol. Evolut. 2014, 32, 173–183. [Google Scholar] [CrossRef] [PubMed]
- Kardong, K.V. ‘Protovipers’ and the evolution of snake fangs. Evolution 1979, 33, 433–443. [Google Scholar]
- Deufel, A.; Cundall, D. Functional plasticity of the venom delivery system in snakes with a focus on the poststrike prey release behavior. Zool. Anz. A J. Comp. Zool. 2006, 245, 249–267. [Google Scholar] [CrossRef]
- Uetz, P.; Hallermann, J. The Reptile Database. Available online: http://reptile-database.reptarium.cz (accessed on 20 June 2019).
- Chippaux, J.-P.; Williams, V.; White, J. Snake venom variability: Methods of study, results and interpretation. Toxicon 1991, 29, 1279–1303. [Google Scholar] [CrossRef]
- Shashidharamurthy, R.; Kemparaju, K. Region-specific neutralization of Indian cobra (Naja naja) venom by polyclonal antibody raised against the eastern regional venom: A comparative study of the venoms from three different geographical distributions. Int. Immunopharmacol. 2007, 7, 61–69. [Google Scholar] [CrossRef] [PubMed]
- Tsai, I.-H.; Tsai, H.-Y.; Wang, Y.-M.; Warrell, D.A. Venom phospholipases of Russell’s vipers from myanmar and eastern India—Cloning, characterization and phylogeographic analysis. Biochim. Biophys. Acta (BBA)-Proteins Proteom. 2007, 1774, 1020–1028. [Google Scholar] [CrossRef] [PubMed]
- Fry, B.G.; Winkel, K.D.; Wickramaratna, J.C.; Hodgson, W.C.; Wüster, W. Effectiveness of snake antivenom: Species and regional venom variation and its clinical impact. J. Toxicol. Toxin Rev. 2003, 22, 23–34. [Google Scholar] [CrossRef]
- Chatrath, S.T.; Chapeaurouge, A.; Lin, Q.; Lim, T.K.; Dunstan, N.; Mirtschin, P.; Kumar, P.P.; Kini, R.M. Identification of novel proteins from the venom of a cryptic snake drysdalia coronoides by a combined transcriptomics and proteomics approach. J. Proteome Res. 2011, 10, 739–750. [Google Scholar] [CrossRef] [PubMed]
- Watt, G.; Padre, L.; Tuazon, L.; Theakston, R.; Laughlin, L. Bites by the philippine cobra (Naja naja philippinensis): Prominent neurotoxicity with minimal local signs. Am. J. Trop.Med. Hyg. 1988, 39, 306–311. [Google Scholar] [CrossRef]
- Warrell, D.A. Tropical snake bite: Clinical studies in south-east Asia. Toxicon 1986, 23, 25–45. [Google Scholar]
- Wüster, W.; McCarthy, C.J. Venomous Snake Systematics: Implications for Snake Bite Treatment and Toxinology. In Envenomings and Their Treatments; Bon, C., Goyffon, M., Eds.; Fondation MÉrieux: Lyon, France, 1996; pp. 13–23. [Google Scholar]
- Tasoulis, T.; Isbister, G.K. A Review and database of snake venom proteomes. Toxins 2017, 9, 290. [Google Scholar] [CrossRef] [PubMed]
- Williams, H.F.; Hayter, P.; Ravishankar, D.; Baines, A.; Layfield, H.J.; Croucher, L.; Wark, C.; Bicknell, A.B.; Trim, S.; Vaiyapuri, S. Impact of Naja nigricollis venom on the production of methaemoglobin. Toxins 2018, 10, 539. [Google Scholar] [CrossRef] [PubMed]
- Pungerčar, J.; Križaj, I. Understanding the molecular mechanism underlying the presynaptic toxicity of secreted phospholipases A2. Toxicon 2007, 50, 871–892. [Google Scholar] [CrossRef] [PubMed]
- Prasarnpun, S.; Walsh, J.; Awad, S.; Harris, J. Envenoming bites by kraits: The biological basis of treatment-resistant neuromuscular paralysis. Brain 2005, 128, 2987–2996. [Google Scholar] [CrossRef] [PubMed]
- Vulfius, C.A.; Kasheverov, I.E.; Kryukova, E.V.; Spirova, E.N.; Shelukhina, I.V.; Starkov, V.G.; Andreeva, T.V.; Faure, G.; Zouridakis, M.; Tsetlin, V.I.; et al. Pancreatic and snake venom presynaptically active phospholipases A2 inhibit nicotinic acetylcholine receptors. PLoS ONE 2017, 12, e0186206. [Google Scholar] [CrossRef] [PubMed]
- Paoli, M.; Rigoni, M.; Koster, G.; Rossetto, O.; Montecucco, C.; Postle, A.D. Mass spectrometry analysis of the phospholipase A2 activity of snake pre-synaptic neurotoxins in cultured neurons. J. Neurochem. 2009, 111, 737–744. [Google Scholar] [CrossRef] [PubMed]
- Dixon, R.W.; Harris, J.B. Myotoxic activity of the toxic phospholipase, notexin, from the venom of the Australian tiger snake. J. Neuropathol. Exp. Neurol. 1996, 55, 1230–1237. [Google Scholar] [CrossRef] [PubMed]
- Gutiérrez, J.M.; Ownby, C.L. Skeletal muscle degeneration induced by venom phospholipases A2: Insights into the mechanisms of local and systemic myotoxicity. Toxicon 2003, 42, 915–931. [Google Scholar] [CrossRef]
- Fuly, A.; Machado, O.; Alves, E.; Carlini, C. Mechanism of inhibitory action on platelet activation of a phospholipase A2 isolated from Lachesis muta (Bushmaster) snake venom. Thromb. Haemost. 1997, 78, 1372–1380. [Google Scholar] [CrossRef]
- Tzeng, M.-C.; Yen, C.-H.; Hseu, M.-J.; Dupureur, C.M.; Tsai, M.-D. Conversion of bovine pancreatic phospholipase A at a single site into a competitor of neurotoxic phospholipases A by site-directed mutagenesis. J. Biol. Chem. 1995, 270, 2120–2123. [Google Scholar] [CrossRef]
- Kini, R.M. Excitement ahead: Structure, function and mechanism of snake venom phospholipase A2 enzymes. Toxicon 2003, 42, 827–840. [Google Scholar] [CrossRef] [PubMed]
- Ramos, O.; Selistre-de-Araujo, H. Snake venom metalloproteases—Structure and function of catalytic and disintegrin domains. Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2006, 142, 328–346. [Google Scholar] [CrossRef] [PubMed]
- Mackessy, S.P. Handbook of Venoms and Toxins of Reptiles; CRC Press: Boca Raton, FL, USA, 2010. [Google Scholar]
- Williams, H.F.; Mellows, B.A.; Mitchell, R.; Sfyri, P.; Layfield, H.J.; Salamah, M.; Vaiyapuri, R.; Collins-Hooper, H.; Bicknell, A.B.; Matsakas, A.; et al. Mechanisms underpinning the permanent muscle damage induced by snake venom metalloprotease. PLoS Negl. Trop. Dis. 2019, 13, e0007041. [Google Scholar] [CrossRef] [PubMed]
- Siigur, E.; Siigur, J. Purification and characterization of lebetase, a fibrinolytic enzyme from vipera lebetina (snake) venom. Biochim. Biophys. Acta (BBA) Gene. Subj. 1991, 1074, 223–229. [Google Scholar] [CrossRef]
- Fox, J.W.; Serrano, S.M.T. Structural considerations of the snake venom metalloproteinases, key members of the M12 reprolysin family of metalloproteinases. Toxicon 2005, 45, 969–985. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, B.A.; Deconte, S.R.; de Moura, F.B.R.; Tomiosso, T.C.; Clissa, P.B.; Andrade, S.P.; Araújo, F.d.A. Inflammation, angiogenesis and fibrogenesis are differentially modulated by distinct domains of the snake venom metalloproteinase jararhagin. Int. J. Biol. Macromol. 2018, 119, 1179–1187. [Google Scholar] [CrossRef] [PubMed]
- Clissa, P.B.; Lopes-Ferreira, M.; Della-Casa, M.S.; Farsky, S.H.P.; Moura-da-Silva, A.M. Importance of jararhagin disintegrin-like and cysteine-rich domains in the early events of local inflammatory response. Toxicon 2006, 47, 591–596. [Google Scholar] [CrossRef] [PubMed]
- Clemetson, K.J. Snaclecs (snake C-type lectins) that inhibit or activate platelets by binding to receptors. Toxicon 2010, 56, 1236–1246. [Google Scholar] [CrossRef] [PubMed]
- Serrano, S.M.; Maroun, R.C. Snake venom serine proteinases: Sequence homology vs. substrate specificity, a paradox to be solved. Toxicon 2005, 45, 1115–1132. [Google Scholar] [CrossRef] [PubMed]
- Stocker, K.; Fischer, H.; Meier, J. Thrombin-like snake venom proteinases. Toxicon 1982, 20, 265–273. [Google Scholar] [CrossRef]
- Xiong, S.; Huang, C. Synergistic strategies of predominant toxins in snake venoms. Toxicol. Lett. 2018, 287, 142–154. [Google Scholar] [CrossRef] [PubMed]
- Vaiyapuri, S.; Sunagar, K.; Gibbins, J.; Jackson, T.; Reeks, T.; Fry, B. Kallikrein enzymes. In Venomous Reptiles and Their Toxins: Evolution, Pathophysiology and Biodiscovery; Fry, B.G., Ed.; Oxford University Press: Oxford, UK, 2015; pp. 267–280. [Google Scholar]
- Kisiel, W. Effect of snake venoms on factor V. Handb. Nat. Toxins Reptil. Venom. Toxins 2018, 2018, 253–264. [Google Scholar]
- Santos, B.F.; Serrano, S.M.; Kuliopulos, A.; Niewiarowski, S. Interaction of viper venom serine peptidases with thrombin receptors on human platelets. FEBS Lett. 2000, 477, 199–202. [Google Scholar] [CrossRef]
- Vaiyapuri, S.; Thiyagarajan, N.; Hutchinson, E.G.; Gibbins, J.M. Sequence and phylogenetic analysis of viper venom serine proteases. Bioinformation 2012, 8, 763–772. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sanchez, E.F.; Santos, C.I.; Magalhaes, A.; Diniz, C.R.; Figueiredo, S.; Gilroy, J.; Richardson, M. Isolation of a proteinase with plasminogen-activating activity from Lachesis muta muta (bushmaster) snake venom. Arch. Biochem. Biophys. 2000, 378, 131–141. [Google Scholar] [CrossRef] [PubMed]
- Kisiel, W.; Kondo, S.; Smith, K.; McMullen, B.; Smith, L. Characterization of a protein C activator from agkistrodon contortrix contortrix venom. J. Biol. Chem. 1987, 262, 12607–12613. [Google Scholar] [PubMed]
- Meléndez-Martínez, D.; Muñoz, J.M.; Barraza-Garza, G.; Cruz-Peréz, M.S.; Gatica-Colima, A.; Alvarez-Parrilla, E.; Plenge-Tellechea, L.F. Rattlesnake crotalus molossus nigrescens venom induces oxidative stress on human erythrocytes. J. Venom. Anim. Toxins Incl. Trop. Dis. 2017, 23, 24. [Google Scholar] [CrossRef]
- Sharma, R.D.; Katkar, G.D.; Sundaram, M.S.; Paul, M.; NaveenKumar, S.K.; Swethakumar, B.; Hemshekhar, M.; Girish, K.S.; Kemparaju, K. Oxidative stress-induced methemoglobinemia is the silent killer during snakebite: A novel and strategic neutralization by melatonin. J. Pineal Res. 2015, 59, 240–254. [Google Scholar] [CrossRef]
- Izidoro, L.F.M.; Ribeiro, M.C.; Souza, G.R.; Sant’Ana, C.D.; Hamaguchi, A.; Homsi-Brandeburgo, M.I.; Goulart, L.R.; Beleboni, R.O.; Nomizo, A.; Sampaio, S.V. Biochemical and functional characterization of an L-amino acid oxidase isolated from Bothrops pirajai snake venom. Bioorg. Med. Chem. 2006, 14, 7034–7043. [Google Scholar] [CrossRef]
- Ande, S.R.; Kommoju, P.R.; Draxl, S.; Murkovic, M.; Macheroux, P.; Ghisla, S.; Ferrando-May, E. Mechanisms of cell death induction by L-amino acid oxidase, a major component of ophidian venom. Apoptosis 2006, 11, 1439–1451. [Google Scholar] [CrossRef]
- Sakurai, Y.; Shima, M.; Matsumoto, T.; Takatsuka, H.; Nishiya, K.; Kasuda, S.; Fujimura, Y.; Yoshioka, A. Anticoagulant activity of M-LAO, l-amino acid oxidase purified from agkistrodon halys blomhoffii, through selective inhibition of factor IX. Biochim. Biophys. Acta (BBA)-Proteins Proteom. 2003, 1649, 51–57. [Google Scholar] [CrossRef]
- de Queiroz, M.R.; de Sousa, B.B.; da Cunha Pereira, D.F.; Mamede, C.C.N.; Matias, M.S.; de Morais, N.C.G.; de Oliveira Costa, J.; de Oliveira, F. The role of platelets in hemostasis and the effects of snake venom toxins on platelet function. Toxicon 2017, 133, 33–47. [Google Scholar] [CrossRef] [PubMed]
- Schetinger, M.R.; Rocha, J.B.T.; Ahmed, M.; Morsch, V.M. Snake venom acetylcholinesterase. In Handbook of Venoms and Toxins of Reptiles; Mackessy, S.P., Ed.; CRC Press Inc.: London, UK, 2009; pp. 207–219. [Google Scholar]
- Isoyama, T.; Thwaites, D.; Selzer, M.G.; Carey, R.I.; Barbucci, R.; Lokeshwar, V.B. Differential selectivity of hyaluronidase inhibitors toward acidic and basic hyaluronidases. Glycobiology 2006, 16, 11–21. [Google Scholar] [CrossRef] [PubMed]
- Suwansrinon, K.; Khow, O.; Mitmoonpitak, C.; Daviratanasilpa, S.; Chaiyabutr, N.; Sitprija, V. Effects of Russell’s viper venom fractions on systemic and renal hemodynamics. Toxicon 2007, 49, 82–88. [Google Scholar] [CrossRef] [PubMed]
- Dhananjaya, B.L.; Vishwanath, B.S.; D’Souza, C.J. Snake venom nucleases, nucleotidases, and phosphomonoesterases. Handb. Venom. Toxins Reptil. 2010, 155, 171. [Google Scholar]
- Aird, S.D. Ophidian envenomation strategies and the role of purines. Toxicon 2002, 40, 335–393. [Google Scholar] [CrossRef]
- Fry, B.G. Venomous Reptiles and Their Toxins: Evolution, Pathophysiol. and Biodiscovery; Oxford University Press: Oxford, UK, 2015. [Google Scholar]
- Aird, S.D.; Watanabe, Y.; Villar-Briones, A.; Roy, M.C.; Terada, K.; Mikheyev, A.S. Quantitative high-throughput profiling of snake venom gland transcriptomes and proteomes (Ovophis okinavensis and Protobothrops flavoviridis). BMC Genom. 2013, 14, 790. [Google Scholar] [CrossRef] [PubMed]
- Pawlak, J.; Mackessy, S.P.; Fry, B.G.; Bhatia, M.; Mourier, G.; Fruchart-Gaillard, C.; Servent, D.; Ménez, R.; Stura, E.; Ménez, A. Denmotoxin, a three-finger toxin from the colubrid snake Boiga dendrophila (Mangrove Catsnake) with bird-specific activity. J. Biol. Chem. 2006, 281, 29030–29041. [Google Scholar] [CrossRef] [PubMed]
- Heyborne, W.H.; Mackessy, S.P. Identification and characterization of a taxon-specific three-finger toxin from the venom of the green vinesnake (Oxybelis fulgidus; family Colubridae). Biochimie 2013, 95, 1923–1932. [Google Scholar] [CrossRef]
- Kini, R.M.; Doley, R. Structure, function and evolution of three-finger toxins: Mini proteins with multiple targets. Toxicon 2010, 56, 855–867. [Google Scholar] [CrossRef] [PubMed]
- Nirthanan, S.; Gwee, M.C.E. Three-Finger & alpha;-Neurotoxins and the Nicotinic Acetylcholine Receptor, Forty Years On. J. Pharmacol. Sci. 2004, 94, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Palud, D.; Soioulata, A.; Haw, C.; Mugnier, L.; Jarrosson, F.; Tarbe, M.; Mollet, C.; de Pomyers, H.; Gigmes, D.; Mabrouk, K. Isolation and characterization of Fasciculin II: From Dendroaspis angusticeps snake venom and from chemical synthesis. Toxicon 2018, 149, 94–95. [Google Scholar] [CrossRef]
- Harvey, A.L. Cardiotoxins from cobra venoms. In Handbook of Natural Toxins; Routledge: London, UK, 2018; pp. 85–106. [Google Scholar]
- Fry, B.G.; Vidal, N.; Norman, J.A.; Vonk, F.J.; Scheib, H.; Ramjan, S.R.; Kuruppu, S.; Fung, K.; Hedges, S.B.; Richardson, M.K. Early evolution of the venom system in lizards and snakes. Nature 2006, 439, 584. [Google Scholar] [CrossRef] [PubMed]
- Nobile, M.; Noceti, F.; Prestipino, G.; Possani, L.D. Helothermine, a lizard venom toxin, inhibits calcium current in cerebellar granules. Exp. Brain Res. 1996, 110, 15–20. [Google Scholar] [CrossRef] [PubMed]
- Nobile, M.; Magnelli, V.; Lagostena, L.; Mochca-Morales, J.; Possani, L.; Prestipino, G. The toxin helothermine affects potassium currents in newborn rat cerebellar granule cells. J. Membr Biol. 1994, 139, 49–55. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, N.; Yamazaki, Y.; Brown, R.L.; Fujimoto, Z.; Morita, T.; Mizuno, H. Structures of pseudechetoxin and pseudecin, two snake-venom cysteine-rich secretory proteins that target cyclic nucleotide-gated ion channels: Implications for movement of the C-terminal cysteine-rich domain. Acta Crystallogr. Sect. D: Biol. Crystallogr. 2008, 64, 1034–1042. [Google Scholar] [CrossRef] [PubMed]
- Yamazaki, Y.; Morita, T. Structure and function of snake venom cysteine-rich secretory proteins. Toxicon 2004, 44, 227–231. [Google Scholar] [CrossRef]
- Yamazaki, Y.; Koike, H.; Sugiyama, Y.; Motoyoshi, K.; Wada, T.; Hishinuma, S.; Mita, M.; Morita, T. Cloning and characterization of novel snake venom proteins that block smooth muscle contraction. Eur. J. Biochem. 2002, 269, 2708–2715. [Google Scholar] [CrossRef]
- Yuan, C.-H.; He, Q.-Y.; Peng, K.; Diao, J.-B.; Jiang, L.-P.; Tang, X.; Liang, S.-P. Discovery of a distinct superfamily of Kunitz-type toxin (KTT) from tarantulas. PLoS ONE 2008, 3, e3414. [Google Scholar] [CrossRef]
- Laustsen, A.H.; Lomonte, B.; Lohse, B.; Fernández, J.; Gutiérrez, J.M. Unveiling the nature of black mamba (Dendroaspis polylepis) venom through venomics and antivenom immunoprofiling: Identification of key toxin targets for antivenom development. J. Proteom. 2015, 119, 126–142. [Google Scholar] [CrossRef]
- Harvey, A.L. Twenty years of dendrotoxins. Toxicon 2001, 39, 15–26. [Google Scholar] [CrossRef]
- Laustsen, A.H.; Karatt-Vellatt, A.; Masters, E.W.; Arias, A.S.; Pus, U.; Knudsen, C.; Oscoz, S.; Slavny, P.; Griffiths, D.T.; Luther, A.M.; et al. In vivo neutralization of dendrotoxin-mediated neurotoxicity of black mamba venom by oligoclonal human IgG antibodies. Nat. Commun. 2018, 9, 3928. [Google Scholar] [CrossRef] [PubMed]
- Laustsen, A.H. Toxin synergism in snake venoms. Toxin Rev. 2016, 35, 165–170. [Google Scholar] [CrossRef] [Green Version]
- Kwong, P.D.; McDonald, N.Q.; Sigler, P.B.; Hendrickson, W.A. Structure of β2-bungarotoxin: Potassium channel binding by Kunitz modules and targeted phospholipase action. Structure 1995, 3, 1109–1119. [Google Scholar] [CrossRef]
- Baconguis, I.; Bohlen, C.J.; Goehring, A.; Julius, D.; Gouaux, E. X-Ray structure of acid-sensing Ion channel 1–Snake toxin complex reveals open state of a Na+-selective channel. Cell 2014, 156, 717–729. [Google Scholar] [CrossRef]
- Ebner, S.; Sharon, N.; Ben-Tal, N. Evolutionary analysis reveals collective properties and specificity in the C-type lectin and lectin-like domain superfamily. Proteins Struct. Funct. Bioinform. 2003, 53, 44–55. [Google Scholar] [CrossRef]
- Vaiyapuri, S.; Hutchinson, E.G.; Ali, M.S.; Dannoura, A.; Stanley, R.G.; Harrison, R.A.; Bicknell, A.B.; Gibbins, J.M. Rhinocetin, a venom-derived integrin-specific antagonist inhibits collagen-induced platelet and endothelial cell functions. J. Biol. Chem. 2012, 287, 26235–26244. [Google Scholar] [CrossRef]
- Clemetson, K.J.; Lu, Q.; Clemetson, J.M. Snake C-type lectin-like proteins and platelet receptors. Pathophysiol. Haemost. Thromb. 2005, 34, 150–155. [Google Scholar] [CrossRef]
- Du, X.-Y.; Navdaev, A.; Clemetson, J.M.; Magnenat, E.; Wells, T.N.; Clemetson, K.J. Bilinexin, a snake C-type lectin from Agkistrodon bilineatus venom agglutinates platelets via GPIb and α2β1. Thromb. Haemost. 2001, 86, 1277–1283. [Google Scholar] [CrossRef]
- Suzuki-Inoue, K.; Fuller, G.L.; García, Á.; Eble, J.A.; Pöhlmann, S.; Inoue, O.; Gartner, T.K.; Hughan, S.C.; Pearce, A.C.; Laing, G.D. A novel Syk-dependent mechanism of platelet activation by the C-type lectin receptor CLEC-2. Blood 2006, 107, 542–549. [Google Scholar] [CrossRef] [Green Version]
- Markland, F.S. Snake venoms and the hemostatic system. Toxicon 1998, 36, 1749–1800. [Google Scholar] [CrossRef]
- Nymalm, Y.; Puranen, J.S.; Nyholm, T.K.; Käpylä, J.; Kidron, H.; Pentikäinen, O.T.; Airenne, T.T.; Heino, J.; Slotte, J.P.; Johnson, M.S. Jararhagin-derived RKKH peptides induce structural changes in α1I domain of human integrin α1β1. J. Biol. Chem. 2004, 279, 7962–7970. [Google Scholar] [CrossRef] [PubMed]
- Calvete, J.J.; Marcinkiewicz, C.; Monleón, D.; Esteve, V.; Celda, B.; Juárez, P.; Sanz, L. Snake venom disintegrins: Evolution of structure and function. Toxicon 2005, 45, 1063–1074. [Google Scholar] [CrossRef] [PubMed]
- Marcinkiewicz, C.; Calvete, J.J.; Marcinkiewicz, M.M.; Raida, M.; Vijay-Kumar, S.; Huang, Z.; Lobb, R.R.; Niewiarowski, S. EC3, a novel heterodimeric disintegrin from Echis carinatus venom, inhibits α4 and α5 integrins in an RGD-independent manner. J. Biol. Chem. 1999, 274, 12468–12473. [Google Scholar] [CrossRef] [PubMed]
- Madrigal, M.; Sanz, L.; Flores-Díaz, M.; Sasa, M.; Núñez, V.; Alape-Girón, A.; Calvete, J.J. Snake venomics across genus Lachesis. Ontogenetic changes in the venom composition of Lachesis stenophrys and comparative proteomics of the venoms of adult Lachesis melanocephala and Lachesis acrochorda. J. Proteom. 2012, 77, 280–297. [Google Scholar] [CrossRef] [PubMed]
- Lainchbury, J.G.; Burnett Jr, J.C.; Meyer, D.; Redfield, M.M. Effects of natriuretic peptides on load and myocardial function in normal and heart failure dogs. Am. J. Physiol. Heart Circ. Physiol. 2000, 278, H33–H40. [Google Scholar] [CrossRef] [PubMed]
- Collins, E.; Bracamonte, M.P.; Burnett Jr, J.C.; Miller, V.M. Mechanism of relaxations to dendroaspis natriuretic peptide in canine coronary arteries. J. Cardiovasc. Pharmacol. 2000, 35, 614–618. [Google Scholar] [CrossRef] [PubMed]
- Sciani, J.M.; Pimenta, D.C. The modular nature of bradykinin-potentiating peptides isolated from snake venoms. J. Venom. Anim. Toxins Incl. Trop. Dis. 2017, 23, 45. [Google Scholar] [CrossRef] [PubMed]
- Sunagar, K.; Fry, B.G.; Jackson, T.N.; Casewell, N.R.; Undheim, E.A.; Vidal, N.; Ali, S.A.; King, G.F.; Vasudevan, K.; Vasconcelos, V.; et al. Molecular evolution of vertebrate neurotrophins: Co-option of the highly conserved nerve growth factor gene into the advanced snake venom arsenalf. PLoS ONE 2013, 8, e81827. [Google Scholar] [CrossRef]
- Wei, C.; Chen, J. A novel lipocalin homologue from the venom gland of Deinagkistrodon acutus similar to mammalian lipocalins. J. Venom. Anim. Toxins Incl. Trop. Dis. 2012, 18, 16–23. [Google Scholar] [Green Version]
- Bdolah, A.; Ducancel, F.; Sunagar, K.; Jackson, T.; Fry, B. Sarafotoxin peptides. Venomous Reptiles and Their Toxins: Evolution, Pathophysiology and Biodiscovery; Oxford University Press: New York, NY, USA, 2015; p. 341. [Google Scholar]
- The World Health Organization. WHO Guidelines for the Production, Control and Regulation of Snake Antivenom Immunoglobulins; WHO: Geneva, Switzerland, 2010; pp. 1–134. [Google Scholar]
- Gutiérrez, J.M. Global availability of antivenoms: The relevance of public manufacturing laboratories. Toxins 2018, 11, 5. [Google Scholar] [CrossRef] [PubMed]
- Zolfagharian, H.; Mohammadpour Dounighi, N. Progress and improvement of the manufacturing process of snake antivenom. Arch. Razi Inst. 2016, 68, 1–10. [Google Scholar]
- Abbas, A.K.; Lichtman, A.H.; Pillai, S. Cellular and Molecular Immunology; Elsevier Health Sciences: Philadelphia, PA, USA, 2011. [Google Scholar]
- Brown, N.; Landon, J. Antivenom: The most cost-effective treatment in the world? Toxicon 2010, 55, 1405–1407. [Google Scholar] [CrossRef] [PubMed]
- Williams, D.J.; Gutiérrez, J.-M.; Calvete, J.J.; Wüster, W.; Ratanabanangkoon, K.; Paiva, O.; Brown, N.I.; Casewell, N.R.; Harrison, R.A.; Rowley, P.D. Ending the drought: New strategies for improving the flow of affordable, effective antivenoms in Asia and Africa. J. Proteom. 2011, 74, 1735–1767. [Google Scholar] [CrossRef]
- Herrera, M.; Paiva, O.K.; Pagotto, A.H.; Segura, A.; Serrano, S.M.; Vargas, M.; Villalta, M.; Jensen, S.D.; León, G.; Williams, D.J. Antivenomic characterization of two antivenoms against the venom of the taipan, oxyuranus scutellatus, from Papua New Guinea and Australia. Am. J. Trop.Med. Hyg. 2014, 91, 887–894. [Google Scholar] [CrossRef] [PubMed]
- Segura, Á.; Herrera, M.; Villalta, M.; Vargas, M.; Gutiérrez, J.M.; León, G. Assessment of snake antivenom purity by comparing physicochemical and immunochemical methods. Biologicals 2013, 41, 93–97. [Google Scholar] [CrossRef]
- Al-Abdulla, I.; Garnvwa, J.M.; Rawat, S.; Smith, D.S.; Landon, J.; Nasidi, A. Formulation of a liquid ovine Fab-based antivenom for the treatment of envenomation by the nigerian carpet viper (Echis ocellatus). Toxicon 2003, 42, 399–404. [Google Scholar] [CrossRef]
- Rojas, G.; Espinoza, M.; Lomonte, B.; Gutiérrez, J. Effect of storage temperature on the stability of the liquid polyvalent antivenom produced in Costa Rica. Toxicon 1990, 28, 101–105. [Google Scholar] [CrossRef]
- O’Leary, M.A.; Kornhauser, R.S.; Hodgson, W.C.; Isbister, G.K. An examination of the activity of expired and mistreated commercial Australian antivenoms. Trans. R. Soc. Trop. Med. Hyg. 2009, 103, 937–942. [Google Scholar] [CrossRef]
- American College of Medical, T.; American Academy of Clinical, T. Antidote shortages in the USA: Impact and response. J. Med. Toxicol 2015, 11, 144–146. [Google Scholar] [CrossRef]
- Snow, R.; Bronzan, R.; Roques, T.; Nyamawi, C.; Murphy, S.; Marsh, K. The prevalence and morbidity of snake bite and treatment-seeking behaviour among a rural Kenyan population. Ann. Trop. Med. Parasitol. 1994, 88, 665–671. [Google Scholar] [CrossRef] [PubMed]
- Omidfar, K.; Rasaee, M.J.; Kashanian, S.; Paknejad, M.; Bathaie, Z. Studies of thermostability in Camelus bactrianus (Bactrian camel) single-domain antibody specific for the mutant epidermal-growth-factor receptor expressed by Pichia. Biotechnol. Appl. Biochem. 2007, 46, 41–49. [Google Scholar] [PubMed]
- Harrison, R.A.; Cook, D.A.; Renjifo, C.; Casewell, N.R.; Currier, R.B.; Wagstaff, S.C. Research strategies to improve snakebite treatment: Challenges and progress. J. Proteom. 2011, 74, 1768–1780. [Google Scholar] [CrossRef] [PubMed]
- Gutiérrez, J.M.; Theakston, R.D.G.; Warrell, D.A. Confronting the neglected problem of snake bite envenoming: The need for a global partnership. PLoS Med. 2006, 3, e150. [Google Scholar] [CrossRef] [PubMed]
- The World Health Organisation. Guidelines for the Management of Snakebite; WHO: Geneva, Switzerland, 2016. [Google Scholar]
- Morais, V.; Massaldi, H. Snake antivenoms: Adverse reactions and production technology. J. Venom. Anim. Toxins Incl. Trop. Dis. 2009, 15, 2–18. [Google Scholar] [CrossRef]
- de Silva, H.A.; Pathmeswaran, A.; Ranasinha, C.D.; Jayamanne, S.; Samarakoon, S.B.; Hittharage, A.; Kalupahana, R.; Ratnatilaka, G.A.; Uluwatthage, W.; Aronson, J.K.; et al. Low-Dose adrenaline, promethazine, and hydrocortisone in the prevention of acute adverse reactions to antivenom following snakebite: A randomised, double-blind, placebo-controlled trial. PLoS Med. 2011, 8, e1000435. [Google Scholar] [CrossRef]
- Malasit, P.; Warrell, D.A.; Chanthavanich, P.; Viravan, C.; Mongkolsapaya, J.; Singhthong, B.; Supich, C. Prediction, prevention, and mechanism of early (anaphylactic) antivenom reactions in victims of snake bites. Br. Med. J. 1986, 292, 17–20. [Google Scholar] [CrossRef]
- Otero, R.; Gutiérrez, J.; Rojas, G.; Núñez, V.; Dıaz, A.; Miranda, E.; Uribe, A.; Silva, J.; Ospina, J.; Medina, Y. A randomized blinded clinical trial of two antivenoms, prepared by caprylic acid or ammonium sulphate fractionation of IgG, in bothrops and porthidium snake bites in Colombia: Correlation between safety and biochemical characteristics of antivenoms. Toxicon 1999, 37, 895–908. [Google Scholar] [CrossRef]
- Lomonte, B.; León, G.; Hanson, L.Å. Similar effectiveness of Fab and F (ab′) 2 antivenoms in the neutralization of hemorrhagic activity of vipera berus snake venom in mice. Toxicon 1996, 34, 1197–1202. [Google Scholar] [CrossRef]
- Theakston, R.D.G.; Warrell, D.; Griffiths, E. Report of a WHO workshop on the standardization and control of antivenoms. Toxicon 2003, 41, 541–557. [Google Scholar] [CrossRef]
- Warrell, D. The global problem of snake bite: Its prevention and treatment. Recent Adv. Toxinol. Res. 1992, 1, 121–153. [Google Scholar]
- Cook, D.A.; Owen, T.; Wagstaff, S.C.; Kinne, J.; Wernery, U.; Harrison, R.A. Analysis of camelid IgG for antivenom development: Serological responses of venom-immunised camels to prepare either monospecific or polyspecific antivenoms for West Africa. Toxicon 2010, 56, 363–372. [Google Scholar] [CrossRef]
- Darvish, M.; Ebrahimi, S.A.; Shahbazzadeh, D.; Bagheri, K.-P.; Behdani, M.; Shokrgozar, M.A. Camelid antivenom development and potential in vivo neutralization of hottentotta saulcyi scorpion venom. Toxicon 2016, 113, 70–75. [Google Scholar] [CrossRef] [PubMed]
- Hamers-Casterman, C.; Atarhouch, T.; Muyldermans, S.; Robinson, G.; Hammers, C.; Songa, E.B.; Bendahman, N.; Hammers, R. Naturally occurring antibodies devoid of light chains. Nature 1993, 363, 446–448. [Google Scholar] [CrossRef] [PubMed]
- De Genst, E.; Silence, K.; Decanniere, K.; Conrath, K.; Loris, R.; Kinne, J.; Muyldermans, S.; Wyns, L. Molecular basis for the preferential cleft recognition by dromedary heavy-chain antibodies. Proc. Natl. Acad. Sci. USA 2006, 103, 4586–4591. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vaiyapuri, S.; Vaiyapuri, R.; Ashokan, R.; Ramasamy, K.; Nattamaisundar, K.; Jeyaraj, A.; Chandran, V.; Gajjeraman, P.; Baksh, M.F.; Gibbins, J.M. Snakebite and its socio-economic impact on the rural population of Tamil Nadu, India. PLoS ONE 2013, 8, e80090. [Google Scholar] [CrossRef]
- Boyer, L.V. On 1000-fold pharmaceutical price markups, and why drugs cost more in the US than in Mexico. Am. J. Med. 2015, 128, 1265–1267. [Google Scholar] [CrossRef]
- Seth, A.; Varma, P.; Pakhetra, R. Randomised control trial on the effective dose of anti-snake venom in cases of snake bite with systemic envenomation. J. Assoc. Physicians India 2000, 48, 756. [Google Scholar]
- Vijeth, S.; Dutta, T.; Shahapurkar, J.; Sahai, A. Dose and frequency of anti-snake venom injection in treatment of Echis carinatus (saw-scaled viper) bite. J. Assoc. Physicians India 2000, 48, 187–191. [Google Scholar]
- Paul, V.; Pratibha, S.; Prahlad, K.; Earali, J.; Francis, S.; Lewis, F. High-dose anti-snake venom versus low-dose anti-snake venom in the treatment of poisonous snake bites-a critical study. J. Assoc. Physicians India 2004, 52, 14–17. [Google Scholar]
- Panda, S.K.; Padhi, L.; Sahoo, G. Oral bacterial flora of Indian cobra (Naja naja) and their antibiotic susceptibilities. Heliyon 2018, 4, e01008. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blaylock, R. Normal oral bacterial flora from some southern African snakes. Onderstepoort J. Vet. Res. 2001, 68, 175–182. [Google Scholar] [PubMed]
- Esmaeilishirazifard, E.; Usher, L.; Trim, C.; Denise, H.; Sangal, V.; Tyson, G.H.; Barlow, A.; Redway, K.; Taylor, J.D.; Kremmyda-Vlachou, M. Microbial adaptation to venom is common in snakes and spiders. bioRxiv 2018, 348433. [Google Scholar] [CrossRef] [Green Version]
- Habib, A.G.; Brown, N.I. The snakebite problem and antivenom crisis from a health-economic perspective. Toxicon 2018, 150, 115–123. [Google Scholar] [CrossRef] [PubMed]
- Mahmood, M.A.; Halliday, D.; Cumming, R.; Thwin, K.T.; Myitzu, M.; White, J.; Alfred, S.; Warrell, D.A.; Bacon, D.; Naing, W.; et al. Inadequate knowledge about snakebite envenoming symptoms and application of harmful first aid methods in the community in high snakebite incidence areas of myanmar. PLoS Negl. Trop. Dis. 2019, 13, e0007171. [Google Scholar] [CrossRef] [PubMed]
- Chappuis, F.; Sharma, S.K.; Jha, N.; Loutan, L.; Bovier, P.A. Protection against snake bites by sleeping under a bed net in southeastern Nepal. Am. J. Trop.Med. Hyg. 2007, 77, 197–199. [Google Scholar] [CrossRef]
- Isbister, G.K. Antivenom efficacy or effectiveness: The Australian experience. Toxicology 2010, 268, 148–154. [Google Scholar] [CrossRef]
- Williams, H.F.; Vaiyapuri, R.; Gajjeraman, P.; Hutchinson, G.; Gibbins, J.M.; Bicknell, A.B.; Vaiyapuri, S. Challenges in diagnosing and treating snakebites in a rural population of Tamil Nadu, India: The views of clinicians. Toxicon 2017, 130, 44–46. [Google Scholar] [CrossRef] [Green Version]
- Warrell, D.A.; Theakston, R.D.G.; Sheriff, M.; Ariaratnam, C.A. Distinctive epidemiologic and clinical features of common krait (Bungarus caeruleus) bites in sri lanka. Am. J. Trop. Med. Hyg. 2008, 79, 458–462. [Google Scholar]
- Stone, R.; Seymour, J.; Marshall, O. Plastic containers and the whole-blood clotting test: Glass remains the best option. Trans. R. Soc. Trop. Med. Hyg. 2006, 100, 1168–1172. [Google Scholar] [CrossRef]
- Gaus, D.P.; Herrera, D.F.; Troya, C.J.; Guevara, A.H. Management of snakebite and systemic envenomation in rural ecuador using the 20-minute whole blood clotting test. Wilderness Environ. Med. 2013, 24, 345–350. [Google Scholar] [CrossRef] [PubMed]
- Theakston, R.D.G.; Laing, G.D. Diagnosis of snakebite and the importance of immunological tests in Venom research. Toxins 2014, 6, 1667–1695. [Google Scholar] [CrossRef]
- CSL, C.S.L. Snake venom detection Kit (SVDK)-detection and identification of snake venom. Clin. Toxinol. Clin. Toxinol. 2007, 1–19. [Google Scholar]
- Steuten, J.; Winkel, K.; Carroll, T.; Williamson, N.A.; Ignjatovic, V.; Fung, K.; Purcell, A.W.; Fry, B.G. The molecular basis of cross-reactivity in the Australian snake venom detection kit (SVDK). Toxicon 2007, 50, 1041–1052. [Google Scholar] [CrossRef]
- Jelinek, G.A.; Tweed, C.; Lynch, D.; Celenza, T.; Bush, B.; Michalopoulos, N. Cross reactivity between venomous, mildly venomous, and non-venomous snake venoms with the commonwealth serum laboratories venom detection kit. Emerg. Med. 2004, 16, 459–464. [Google Scholar] [CrossRef] [PubMed]
- Sharma, S.K.; Kuch, U.; Höde, P.; Bruhse, L.; Pandey, D.P.; Ghimire, A.; Chappuis, F.; Alirol, E. Use of molecular diagnostic tools for the identification of species responsible for snakebite in Nepal: A pilot study. PLoS Negl. Trop. Dis. 2016, 10, e0004620. [Google Scholar] [CrossRef] [PubMed]
- Pawade, B.S.; Salvi, N.C.; Shaikh, I.K.; Waghmare, A.B.; Jadhav, N.D.; Wagh, V.B.; Pawade, A.S.; Waykar, I.G.; Potnis-Lele, M. Rapid and selective detection of experimental snake envenomation—Use of gold nanoparticle based lateral flow assay. Toxicon 2016, 119, 299–306. [Google Scholar] [CrossRef]
- Liu, C.-C.; Yu, J.-S.; Wang, P.-J.; Hsiao, Y.-C.; Liu, C.-H.; Chen, Y.-C.; Lai, P.-F.; Hsu, C.-P.; Fann, W.-C.; Lin, C.-C. Development of sandwich ELISA and lateral flow strip assays for diagnosing clinically significant snakebite in Taiwan. PLoS Negl. Trop. Dis. 2018, 12, e0007014. [Google Scholar] [CrossRef]
- Hung, D.Z.; Lin, J.H.; Mo, J.F.; Huang, C.F.; Liau, M.Y. Rapid diagnosis of Naja atra snakebites. Clin. Toxicol. 2014, 52, 187–191. [Google Scholar] [CrossRef]
- Naik, B.S. “Dry bite” in venomous snakes: A review. Toxicon 2017, 133, 63–67. [Google Scholar] [CrossRef]
- Pore, S.M.; Ramanand, S.J.; Patil, P.T.; Gore, A.D.; Pawar, M.P.; Gaidhankar, S.L.; Ghanghas, R.R. A retrospective study of use of polyvalent anti-snake venom and risk factors for mortality from snake bite in a tertiary care setting. Indian J. Pharmacol. 2015, 47, 270–274. [Google Scholar] [CrossRef]
- Litschka-Koen, T.; Williams, D. Snake antivenoms in southern Africa. Contin. Med. Educ. 2011, 29, 75. [Google Scholar]
- Brown, N.I. Consequences of Neglect: Analysis of the sub-saharan African snake antivenom market and the global context. PLoS Negl. Trop. Dis. 2012, 6, e1670. [Google Scholar] [CrossRef]
- Gupta, Y.; Peshin, S. Do herbal medicines have potential for managing snake bite envenomation? Toxicol. Int. 2012, 19, 89. [Google Scholar] [CrossRef] [PubMed]
- Bulfone, T.C.; Samuel, S.P.; Bickler, P.E.; Lewin, M.R. Developing small molecule therapeutics for the Initial and adjunctive treatment of snakebite. J. Trop. Med. 2018, 2018. [Google Scholar] [CrossRef] [PubMed]
- Knudsen, C.; Laustsen, A. Recent advances in next generation snakebite antivenoms. Trop. Med. Infect. Dis. 2018, 3, 42. [Google Scholar] [CrossRef] [PubMed]
- Laustsen, A.; Solà, M.; Jappe, E.; Oscoz, S.; Lauridsen, L.; Engmark, M. Biotechnological trends in spider and scorpion antivenom development. Toxins 2016, 8, 226. [Google Scholar] [CrossRef] [PubMed]
- H Laustsen, A.; Engmark, M.; Milbo, C.; Johannesen, J.; Lomonte, B.; Maria Gutierrez, J.; Lohse, B. From fangs to pharmacology: The future of snakebite envenoming therapy. Current Pharm. Des. 2016, 22, 5270–5293. [Google Scholar] [CrossRef]
- Knudsen, C.; Ledsgaard, L.; Dehli, R.I.; Ahmadi, S.; Sørensen, C.V.; Laustsen, A.H. Engineering and design considerations for next-generation snakebite antivenoms. Toxicon 2019. [Google Scholar] [CrossRef]
- Arias, A.S.; Rucavado, A.; Gutiérrez, J.M. Peptidomimetic hydroxamate metalloproteinase inhibitors abrogate local and systemic toxicity induced by Echis ocellatus (saw-scaled) snake venom. Toxicon 2017, 132, 40–49. [Google Scholar] [CrossRef]
- Lewin, M.; Samuel, S.; Merkel, J.; Bickler, P. Varespladib (LY315920) Appears to Be a potent, broad-spectrum, inhibitor of snake venom Phospholipase A2 and a possible pre-referral treatment for envenomation. Toxins 2016, 8, 248. [Google Scholar] [CrossRef] [PubMed]
- Bittenbinder, M.A.; Zdenek, C.N.; Op den Brouw, B.; Youngman, N.J.; Dobson, J.S.; Naude, A.; Vonk, F.J.; Fry, B.G. Coagulotoxic cobras: Clinical implications of strong anticoagulant actions of African spitting Naja venoms that are not neutralised by antivenom but are by LY315920 (Varespladib). Toxins 2018, 10, 516. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Zhang, J.; Zhang, D.; Xiao, H.; Xiong, S.; Huang, C. Exploration of the inhibitory potential of varespladib for snakebite envenomation. Molecules 2018, 23, 391. [Google Scholar] [CrossRef] [PubMed]
- Bryan-Quiros, W.; Fernandez, J.; Gutierrez, J.M.; Lewin, M.R.; Lomonte, B. Neutralizing properties of LY315920 toward snake venom group I and II myotoxic phospholipases A2. Toxicon 2019, 157, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Escalante, T.; Franceschi, A.; Rucavado, A.; Gutiérrez, J.M. Effectiveness of batimastat, a synthetic inhibitor of matrix metalloproteinases, in neutralizing local tissue damage induced by BaP1, a hemorrhagic metalloproteinase from the venom of the snake Bothrops asper. Biochem. Pharmacol. 2000, 60, 269–274. [Google Scholar] [CrossRef]
- Nicholls, S.J.; Kastelein, J.J.; Schwartz, G.G.; Bash, D.; Rosenson, R.S.; Cavender, M.A.; Brennan, D.M.; Koenig, W.; Jukema, J.W.; Nambi, V. Varespladib and cardiovascular events in patients with an acute coronary syndrome: The VISTA-16 randomized clinical trial. Jama 2014, 311, 252–262. [Google Scholar] [CrossRef] [PubMed]
- Zhu, W.-H.; Guo, X.; Villaschi, S.; Nicosia, R.F. Regulation of vascular growth and regression by matrix metalloproteinases in the rat aorta model of angiogenesis. Lab. Investig. 2000, 80, 545. [Google Scholar] [CrossRef] [PubMed]
- Shah, S.; Naqvi, S.S.; Abbas, M.A. Use of neostigmine in black mamba snake bite: A case report. Anaesth. Pain Intensive Care 2016, 20, 77–79. [Google Scholar]
- Faiz, M.A.; Ahsan, M.F.; Ghose, A.; Rahman, M.R.; Amin, R.; Hossain, M.; Tareq, M.N.; Jalil, M.A.; Kuch, U.; Theakston, R.D.G. Bites by the monocled cobra, Naja kaouthia, in chittagong division, bangladesh: Epidemiology, clinical features of envenoming and management of 70 identified cases. Am. J. Trop. Med. Hyg. 2017, 96, 876–884. [Google Scholar] [CrossRef] [PubMed]
- Lewin, M.R.; Samuel, S.P.; Wexler, D.S.; Bickler, P.; Vaiyapuri, S.; Mensh, B.D. Early treatment with intranasal neostigmine reduces mortality in a mouse model of Naja naja (Indian Cobra) envenomation. J. Trop. Med. 2014, 2014. [Google Scholar] [CrossRef] [PubMed]
- O’Brien, J.; Lee, S.-H.; Onogi, S.; Shea, K.J. Engineering the protein corona of a synthetic polymer nanoparticle for broad-spectrum sequestration and neutralization of venomous biomacromolecules. J. Am. Chem. Soc. 2016, 138, 16604–16607. [Google Scholar] [CrossRef] [PubMed]
- Karain, B.D.; Lee, M.K.H.; Hayes, W.K. C60 Fullerenes as a novel treatment for poisoning and envenomation: A proof-of-concept study for snakebite. J. Nanosci. Nanotechnol. 2016, 16, 7764–7771. [Google Scholar] [CrossRef]
- Silva, L.C.; Pucca, M.B.; Pessenda, G.; Campos, L.B.; Martinez, E.Z.; Cerni, F.A.; Barbosa, J.E. Discovery of human scFvs that cross-neutralize the toxic effects of B. jararacussu and C. d. terrificus venoms. Acta Trop. 2018, 177, 66–73. [Google Scholar] [CrossRef] [PubMed]
- Laustsen, A.H.; Johansen, K.H.; Engmark, M.; Andersen, M.R. Recombinant snakebite antivenoms: A cost-competitive solution to a neglected tropical disease? PLoS Negl. Trop. Dis. 2017, 11, e0005361. [Google Scholar] [CrossRef] [PubMed]
- Gutierrez, J.M.; Rucavado, A. Snake venom metalloproteinases: Their role in the pathogenesis of local tissue damage. Biochimie 2000, 82, 841–850. [Google Scholar] [CrossRef]
- Cook, D.A.; Samarasekara, C.L.; Wagstaff, S.C.; Kinne, J.; Wernery, U.; Harrison, R.A. Analysis of camelid IgG for antivenom development: Immunoreactivity and preclinical neutralisation of venom-induced pathology by IgG subclasses, and the effect of heat treatment. Toxicon 2010, 56, 596–603. [Google Scholar] [CrossRef] [PubMed]
- Anderson, G.P.; Liu, J.H.; Zabetakis, D.; Liu, J.L.; Goldman, E.R. Thermal stabilization of anti-α-cobratoxin single domain antibodies. Toxicon 2017, 129, 68–73. [Google Scholar] [CrossRef] [PubMed]
- Laustsen, A.H.; Gutiérrez, J.M.; Knudsen, C.; Johansen, K.H.; Bermúdez-Méndez, E.; Cerni, F.A.; Jürgensen, J.A.; Ledsgaard, L.; Martos-Esteban, A.; Øhlenschlæger, M. Pros and cons of different therapeutic antibody formats for recombinant antivenom development. Toxicon 2018, 146, 151–175. [Google Scholar] [CrossRef]
- Aubrey, N.; Devaux, C.; Sizaret, P.-Y.; Rochat, H.; Goyffon, M.; Billiald, P. Design and evaluation of a diabody to improve protection against a potent scorpion neurotoxin. Cell. Mol. Life Sci. CMLS 2003, 60, 617–628. [Google Scholar] [CrossRef]
- El-Aziz, T.M.A.; Ravelet, C.; Molgo, J.; Fiore, E.; Pale, S.; Amar, M.; Al-Khoury, S.; Dejeu, J.; Fadl, M.; Ronjat, M. Efficient functional neutralization of lethal peptide toxins in vivo by oligonucleotides. Sci. Rep. 2017, 7, 7202. [Google Scholar] [CrossRef]
- Chen, Y.-J.; Tsai, C.-Y.; Hu, W.-P.; Chang, L.-S. DNA Aptamers against Taiwan banded krait α-bungarotoxin recognize Taiwan cobra cardiotoxins. Toxins 2016, 8, 66. [Google Scholar] [CrossRef] [PubMed]
- Jenkins, T.P.; Fryer, T.; Dehli, R.I.; Jürgensen, J.A.; Fuglsang-Madsen, A.; Føns, S.; Laustsen, A.H. Toxin neutralization using alternative binding proteins. Toxins 2019, 11, 53. [Google Scholar] [CrossRef] [PubMed]
- Griffin, L.C.; Tidmarsh, G.F.; Bock, L.C.; Toole, J.J.; Leung, L. In vivo anticoagulant properties of a novel nucleotide-based thrombin inhibitor and demonstration of regional anticoagulation in extracorporeal circuits. Blood 1993, 81, 3271–3276. [Google Scholar] [PubMed]
- Wuellner, U.; Klupsch, K.; Buller, F.; Attinger-Toller, I.; Santimaria, R.; Zbinden, I.; Henne, P.; Grabulovski, D.; Bertschinger, J.; Brack, S. Bispecific CD3/HER2 targeting FynomAb induces redirected T cell-mediated cytolysis with high potency and enhanced tumor selectivity. Antibodies 2015, 4, 426–440. [Google Scholar] [CrossRef]
- Roopenian, D.C.; Akilesh, S. FcRn: The neonatal Fc receptor comes of age. Nat. Rev. Immunol. 2007, 7, 715. [Google Scholar] [CrossRef] [PubMed]
- Akilesh, S.; Christianson, G.J.; Roopenian, D.C.; Shaw, A.S. Neonatal FcR expression in bone marrow-derived cells functions to protect serum IgG from catabolism. J. Immunol. 2007, 179, 4580–4588. [Google Scholar] [CrossRef] [PubMed]
- Halilu, S.; Iliyasu, G.; Hamza, M.; Chippaux, J.-P.; Kuznik, A.; Habib, A.G. Snakebite burden in Sub-Saharan Africa: Estimates from 41 countries. Toxicon 2019, 159, 1–4. [Google Scholar] [CrossRef] [PubMed]
- Williams, D.J.; Faiz, M.A.; Abela-Ridder, B.; Ainsworth, S.; Bulfone, T.C.; Nickerson, A.D.; Habib, A.G.; Junghanss, T.; Fan, H.W.; Turner, M.; et al. Strategy for a globally coordinated response to a priority neglected tropical disease: Snakebite envenoming. PLoS Negl. Trop. Dis. 2019, 13, e0007059. [Google Scholar] [CrossRef] [PubMed]
- Bouazza, A.; El Hidan, M.A.; Aimrane, A.; Kahime, K.; Lansari, A.; Laaradia, M.A.; Lahouaoui, H.; Moukrim, A. Climate change effects on venomous snakes: Distribution and snakebite epidemiology. In Handbook of Research on Global Environmental Changes and Human Health; IGI Global: Hershey, PA, USA, 2019; pp. 475–490. [Google Scholar]
- Kannt, A.; Wieland, T. Managing risks in drug discovery: Reproducibility of published findings. Naunyn-Schmiedeberg’s Arch. Pharmacol. 2016, 389, 353–360. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Disease (Source of Data) | Causal Species | Estimated Deaths/An | Global Prevalence | Population at Risk | Clinical Manifestations | Treatment | Diagnostics |
|---|---|---|---|---|---|---|---|
| Snakebite Envenomation [7] | Snakes: >90 Genera, >700 Species | 81,000–137,000 | Up to 2,700,000 | 6–7 Billion | Neurotoxicity and paralysis or cardiovascular toxicity and hypovolemic shock. Cytotoxicity leading to tissue damage and amputation. | Anti-venom | Fang marks, local tissue damage, immunoassay (Aus) Clinical/laboratory markers give other indications |
| Cholera [20,21] | Bacterium: Vibrio cholerae | 68,400 | 2,800,000 | 1.4 Billion | Watery diarrhoea | Oral or intravenous rehydration | Stool examination |
| Leishmaniasis [20] | Protist: Leishmania spp. Transmitted by female sandflies; Phlebotomus/Lutzomyia spp. | 24,200 | 12,000,000 | 350 Million | Cutaneous and mucocutaneous disease, kala-azar | Anti-monials, amphotericin B, pentamidine, miltefosine | Biopsy |
| Chagas’ Disease [20] | Protist: Trypanosoma cruzi | 8000 | 5,700,000 | 70 Million | Cardiomyopathy, megacolon, Mega esophagus | Benznidazole, nifurtimox | Blood smear |
| Schistosomiasis (Bilharzia) [20] | Trematodes: Schistosoma spp. | 4400 | 207,000,000 | 779 Million | Hematuria and urogenital disease, intestinal and liver fibrosis, growth and cognitive delays | Praziquantel | Stool examination |
| Human African Trypanosomiasis [20] | Protist: Trypanosoma brucei amongst other species. Transmitted by tsetse flies; Glossina spp. | 3500 | 300,000 | 60 Million | Sleeping sickness | Pentamidine, suramin, melarsoprol, eflornithine | Biopsy or blood smear |
| Ascariasis [20] | Nematode: Ascaris lumbricoides | 2700 | 807,000,000 | 4.2 Billion | Malnutrition, growth and cognitive delays | Albendazole/mabendazole | Stool examination |
| Trichuriasis [22,23] | Nematode: Trichuris trichiura | Deaths rarely direct | 604,000,000 | 3.2 Billion | Inflammatory bowel disease, growth and cognitive delays | Albendazole/mabendazole | Stool examination |
| Hookworm Infection [22,23] | Nematodes: Ancylostoma duodenale/Necatora americanus | Deaths rarely direct | 576,000,000 | 3.2 Billion | Anemia, malnutrition, growth and cognitive delays, poor pregnancy outcome | Albendazole/mabendazole | Stool examination |
| Lymphatic Filariasis [24,25] | Nematodes: Wuchereria bancrofti, Brugia spp. | Deaths rarely direct | 120,000,000 | 1.3 Billion | Adenolymphangitis, lymphedema, hydrocele | Ivermectin/diethylcarbamazine (plus albendazole) | LFA test strip (Alere) |
| Trachoma [19] | Bacterium: Chlamydia trachomatis | Deaths rarely direct | 84,000,000 | 590 Million | Trachomatous folliculitis and inflammation, trichiasis, blindness | Surgery, aziromycin | Clinal diagnosis using loupes (magnifiers) |
| Onchocerciasis [26] | Nematode: Onchocerca volvulus. Transmitted by blackflies; Simulium spp. | Deaths rarely direct | 37,000,000 | 90 Million | Onchocerca, skin disease, blindness | Ivermectin | Biopsy/slit lamp examination/antibody tests |
| Leprosy [27] | Bacterium: Mycobacterium leprae | Deaths rarely direct | 200,000 | ND | Lepromatous leprosy, tuberculoid leprosy | Multidrug therapy, rifampicin, clofazimine, dapsone | Biopsy |
| Dracunculiasis [28] | Nematode: Dracunculus medinensis | Deaths rarely direct | 30 | ND | Disfiguring ulcer, secondary bacterial infection | Metronidazole/thiobendazole adjunctive to self-care and stick therapy | Clinical presentation |
| Venom Component | Approximate Abundance (% (±SD)) | Major Described Functions | ||
|---|---|---|---|---|
| Elapidae | Viperinae | Crotalinae | ||
| Phospholipase A2 (PLA2) | 31 (±24) | 22 (±17) | 22 (±20) | Presynaptic neurotoxicity (β-neurotoxins), membrane phospholipolysis, haemolysis, myotoxicity, necrosis and inhibition/activation of platelets |
| Snake venom metalloprotease (SVMP) | 3 (±3) | 35 (±20) | 36 (±20) | Haemorrhaging, fibrin(ogen)olytic activity, endothelial damage and myotoxicity |
| Snake venom serine protease (SVSP) | 1 (±1) | 12 (±9) | 16 (±14) | Hypotension, fibrin(ogen)olytic activity and bleeding |
| L-amino acid oxidase (LAAO) | 1 (±2) | 2 (±2) | 5 (±4) | Apoptosis, oedema, cytotoxicity via products and anticoagulant effects via inhibition factor IX |
| Three-finger toxin (3FTX) | 55 (±27) | NA | NA | Postsynaptic neurotoxicity via binding of cholinergic receptors (α-neurotoxins), cardiotoxicity, myotoxicity and cytotoxicity |
| Kunitz type serine protease inhibitors (KSPi) | 4 (±10) | 3 (±6) | NA | Neurotoxicity via binding of voltage gated potassium channels or anticoagulopathic effects due to serine protease inhibition |
| Cysteine rich secretory protein (CRiSP) | 2 (±3) | 4 (±4) | 2 (±2) | Smooth muscle inhibition via blocking of calcium channels |
| Natriuretic peptides | 1 (±1) | 1 (±3) | 7 (±9) | Promote excretion of sodium by kidneys causing hypotension and cardiotoxicity |
| Snake C-type lectins (Snaclec) | NA | 9 (±6) | 6 (±8) | Platelet inhibition and activation via an array of receptors |
| Disintegrin | NA | 6 (±5) | 2 (±4) | Binding of integrins causing inhibition of platelet aggregation |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Williams, H.F.; Layfield, H.J.; Vallance, T.; Patel, K.; Bicknell, A.B.; Trim, S.A.; Vaiyapuri, S. The Urgent Need to Develop Novel Strategies for the Diagnosis and Treatment of Snakebites. Toxins 2019, 11, 363. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins11060363
Williams HF, Layfield HJ, Vallance T, Patel K, Bicknell AB, Trim SA, Vaiyapuri S. The Urgent Need to Develop Novel Strategies for the Diagnosis and Treatment of Snakebites. Toxins. 2019; 11(6):363. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins11060363
Chicago/Turabian StyleWilliams, Harry F., Harry J. Layfield, Thomas Vallance, Ketan Patel, Andrew B. Bicknell, Steven A. Trim, and Sakthivel Vaiyapuri. 2019. "The Urgent Need to Develop Novel Strategies for the Diagnosis and Treatment of Snakebites" Toxins 11, no. 6: 363. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins11060363