From Discovery of Snake Venom Disintegrins to A Safer Therapeutic Antithrombotic Agent


1
Department of Medicine, Mackay Medical College, New Taipei City 25245, Taiwan
2
Graduate Institute of Pharmacology, College of Medicine, National Taiwan University, Taipei 10051, Taiwan
*
Author to whom correspondence should be addressed.
Toxins 2019, 11(7), 372; https://doi.org/10.3390/toxins11070372
Submission received: 2 June 2019
/
Revised: 22 June 2019
/
Accepted: 24 June 2019
/
Published: 26 June 2019
(This article belongs to the Special Issue From Toxins to Drugs)
{kind=link}
Abstract
:Snake venoms affect blood coagulation and platelet function in diverse ways. Some venom components inhibit platelet function, while other components induce platelet aggregation. Among the platelet aggregation inhibitors, disintegrins have been recognized as unique and potentially valuable tools for examining cell–matrix and cell–cell interactions and for the development of antithrombotic and antiangiogenic agents according to their anti-adhesive and anti-migration effect on tumor cells and antiangiogenesis activities. Disintegrins represent a family of low molecular weight, cysteine-rich, Arg-Gly-Asp(RGD)/Lys-Gly-Asp(KGD)-containing polypeptides, which inhibit fibrinogen binding to integrin αIIbβ3 (i.e., platelet glycoprotein IIb/IIIa), as well as ligand binding to integrins αvβ3, and α5β1 expressed on cells (i.e., fibroblasts, tumor cells, and endothelial cells). This review focuses on the current efforts attained from studies using disintegrins as a tool in the field of arterial thrombosis, angiogenesis, inflammation, and tumor metastasis, and briefly describes their potential therapeutic applications and side effects in integrin-related diseases. Additionally, novel R(K)GD-containing disintegrin TMV-7 mutants are being designed as safer antithrombotics without causing thrombocytopenia and bleeding.
Keywords:
snake venom proteins; disintegrins; antiplatelet agent; arterial thrombosis; angiogenesis; septic inflammationKey Contribution: Bleeding risk limits the dose and use of current integrin αIIbβ3 antagonists, thus limiting their therapeutic utilization. This review provides a picture of the pathological mechanism of drug-induced thrombocytopenia and bleeding, and may provide a strategy for the development of safer antithrombotic drugs.
1. Introduction
1.1. Role of Platelet Integrin in Thrombosis and Hemostasis
Integrins, one of the adhesion receptor super-families, play vital roles in regulating cell–matrix and cell–cell interactions. Integrins exist as an α:β heterodimeric complex of transmembrane proteins, and play important roles in platelet aggregation, adhesion, spreading, retraction, migration, angiogenesis, inflammatory reactions, and other biological processes [1,2].
Circulating platelets respond very rapidly to vascular injury. The coverage of the exposed endothelium by activated platelets depends on the recognition of adhesive proteins (i.e., fibrinogen, collagen, von Willebrand factor, and fibronectin) by their specific platelet membrane glycoproteins (i.e., αIIbβ3, α2β1, αvβ3, α5β1, and α6β1). At present, α2β1/GPVI is identified to mediate collagen or fibrin adhesion/activation of platelets, whereas αIIbβ3 mediates the adhesion of fibrinogen and the subsequent platelet–platelet interaction, a common final step of platelet aggregation shared by stimulating agonists [3,4].
The processes of platelet activation consist of the stimulation of several signal transductions and result in a shape change, thromboxane A2 formation, release reaction of adenosine diphosphate (ADP), the augmentation of coagulation activity, and finally, the exposure of latent αIIbβ3. Consequently, the plasma fibrinogen binds to the activated integrin αIIbβ3 of the platelets and leads to platelet aggregation, not only maintaining hemostasis and preventing excessive bleeding under normal conditions, but also forming a thrombus under pathological conditions, such as an endothelial injury due to an atherosclerotic lesion [5,6].
1.2. Discovery of Disintegrins from Viper Venom as Antithrombotic Agents
Disintegrin was first discovered in 1987, since trigramin, a non-enzymatic small molecular polypeptide, was isolated from the snake venom of Trimeresurus gramineus [7]. Trigramin inhibited platelet aggregation by blocking fibrinogen binding to aggregation agonist-stimulated platelets. At that time, it was proposed that disintegrins derived from Agkistrodon rhodostoma, Agkistrodon halys, and T. gramineus [8,9,10] inhibited platelet aggregation induced by various agonists, including collagen, ADP, sodium arachidonate, and epinephrine that neither affected the shape change nor the cyclic adenosine monophosphate (cAMP) level. Further studies reported that the disintegrin trigramin inhibited fibrinogen binding to ADP-stimulated platelets, and the binding ability of 125I-trigramin toward ADP-stimulated platelets was almost completely abolished in patients with a genetic αIIbβ3-defect disease (i.e., Glanzmann’s thrombasthenia) when compared to normal platelets, demonstrating that the fibrinogen receptor αIIbβ3 is the target of trigramin [7,11]. In addition, mAb 7E3 raised against αIIbβ3 and RGDS showed an inhibitory effect on 125I-tragramin binding to platelets, indicating that its binding target is αIIbβ3 and the Arg-Gly-Asp (RGD) tripeptide sequence is important for its binding activity. Importantly, the trigramin sequence clearly identified that it is an RGD-containing single polypeptide of 72 amino acid residues with six disulfide bonds [7]. Alkylated and reduced trigramin lost activity in inhibiting platelet aggregation and binding capacity toward platelets, demonstrating that the binding capacity of αIIbβ3 depends on RGD tripeptides, and its secondary structure and the disulfide bridges are essential in the expression of biological activities [7,12]. Upon intravenous administration, the bleeding time of severed mesentery arteries was significantly prolonged by trigramin, indicating that it decreases the ability of platelets to form thromboemboli in vivo [13]. Subsequently, emerging reports have shown that many disintegrins exhibit an inhibitory effect on platelet adhesion to extracorporeal circuit surfaces [14,15]. Since then, disintegrins have been thought to be potential candidates as antithrombotic agents, and this finding has motivated many pharmaceutical companies to develop a series of RGD-mimetic drugs based on the specific steric structure of the RGD loop of disintegrins.
Among these disintegrins, a Lys-Gly-Asp (KGD) containing peptide barbourin exhibited specificity toward integrin αIIbβ3 than to αvβ3 [16]. Therefore, a cyclic KGD peptide, Integrilin, was successfully developed as an antithrombotic agent, and used clinically for the prevention of restenosis after percutaneous transluminal coronary angioplasty [17,18]. To date, three current Food and Drug Administration-approved platelet integrin antagonists, the anti-adhesive agent Eptifibatide (Integrilin, COR Therapeutics), Tirofiban (Aggrastat, Merck), and the chimeric 7E3 Fab (Abciximab, Repro) mAb raised against αIIbβ3, have been successfully developed in this field. Tirofiban is a 495-Da synthetic compound engineered to mimic the RGD sequence and acts as an anti-aggregation agent [19] and Abciximab, a mouse/human chimeric monoclonal c7E3Fab raised against integrin αIIbβ3 [20,21]. These three FDA-approved αIIbβ3 antagonists are administered intravenously. Orally active integrin antagonists have also been developed, but the clinical trials of these oral agents have resulted in increased mortality instead of beneficial effects [22]. Nevertheless, in high-risk patients undergoing percutaneous coronary intervention and transluminal coronary angioplasty, the current integrin antagonists have each demonstrated clear therapeutic benefits, as indicated by a significant decline in death rates and the reoccurrence of myocardial infarction [23].
1.3. Bleeding Side Effects of Current Antithrombotic Agents for Acute Coronary Syndromes
Despite the successful clinical use of integrin antagonists as therapeutic antithrombotic drugs, the risk of life-threatening bleeding directly limited their utilization in patients undergoing percutaneous coronary intervention [24]. The intrinsic mechanism of abnormal bleeding is due to the binding of αIIbβ3 antagonists to the integrin αIIbβ3, inducing conformational changes. After integrin antagonists disassociated from αIIbβ3, the conformationally changed receptor αIIbβ3 exposed the epitopes LIBSs (ligand-induced binding sites) [25]. Recent studies reported that LIBSs represent the binding sites of the intrinsic antibodies in patients administered with these drugs [26]. The complex of LIBS/intrinsic antibody recruited FcγRIIa and induced its downstream ITAM–Syk–phospholipase (PLC)γ2 signaling pathway, leading to FcγRIIa-mediated immune clearance of platelets and thrombocytopenia [27].
After bleeding, drug-induced immune thrombocytopenia and severe reactions to re-administration are the most severe side effects of αIIbβ3 antagonists [28]. These include commonly used drugs such as RGD-mimetic agents, antibiotics, and anticonvulsants, which are used to prevent in-stent thrombosis in patients undergoing percutaneous coronary intervention. Thrombocytopenia and gastrointestinal bleeding induced by RGD-mimetic drugs occur in patients administrated with αIIbβ3 antagonists [29,30]. Although the thrombocytopenia is usually resolved within two weeks after drug withdrawal, at present, there is no treatment to halt severe bleeding or rapidly upsurge the patient’s platelet count if it occurs during the thrombocytopenic phase. Given the widespread use of these efficacious drugs and the relatively high incidence, it is crucial to develop a new class of αIIbβ3 antagonists with a minimal risk of bleeding.
1.4. New Insights into Antiplatelet Strategies
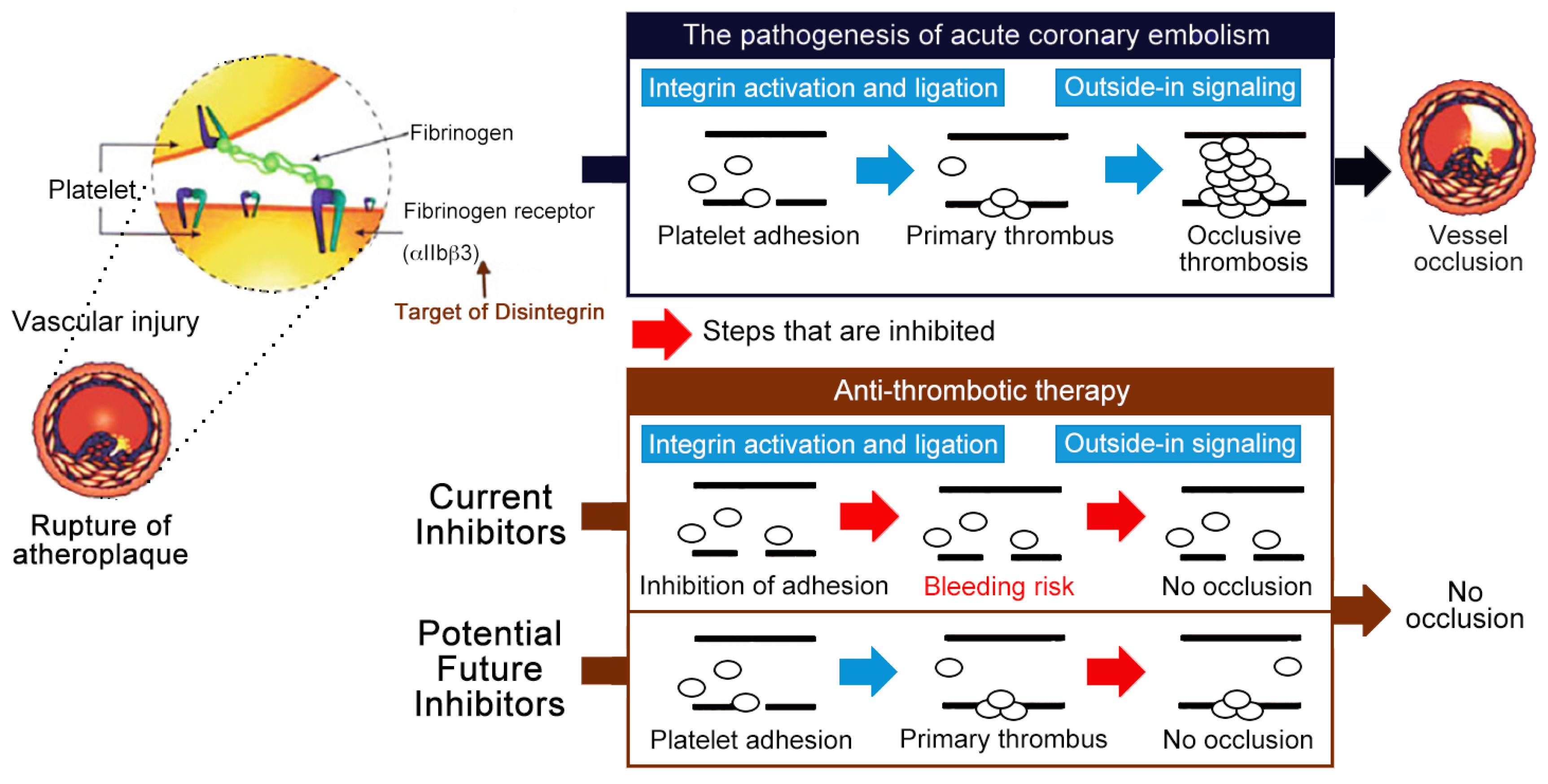
As established by expanded clinical trials and usage, the clinical scenarios in which the current αIIbβ3 antagonists present efficacy are more limited than originally expected. However, the important role of integrin αIIbβ3 in platelet aggregation and thrombosis remains irrefutable. Since the premise that targeting αIIbβ3 remains a fundamentally solid strategy, some investigators have looked to identify a new class of αIIbβ3 antagonists, ones that will either not cause conformational changes on dissociation or association from αIIbβ3, or reduce the occurrences of severe bleeding and thrombocytopenia in some patients. Two potential strategies have been suggested to achieve this end: targeting factors that mediate the propagation of the thrombus without affecting the initial formation of the thrombus core [31], or discovering inhibitors that bind to the extracellular domain of the integrin without causing receptor activation and conformational changes [32]. These may hold the key to safer antithrombotic therapy (Figure 1).
1.4.1. Integrin Antagonists that Minimally Affect Conformational Changes
Tirofiban, Eptifibatide, and Abciximab all cause thrombocytopenia and bleeding, which is associated with integrin conformational changes after drug dissociation. Immune thrombocytopenia occurs on the first exposure to RGD-mimetic drugs and subsequently the platelet counts decrease abruptly within hours of the commencement of drug administration [29], indicating the presence of an intrinsic antiplatelet antibody that occurs naturally. In the majority of patients with RGD-mimetic drug-induced thrombocytopenia, the binding of the antibody was drug specific and only occurred as the integrin combined with the drugs that caused the thrombocytopenia. RGD-mimetic drugs binding to the RGD recognition site of αIIbβ3 cause conformational changes and the exposure of cryptic epitopes, termed MIBSs (integrin mimetic-induced binding sites) [33], or LIBSs (ligand-induced binding sites) [34]. Recently, novel small-molecule integrin antagonists and pure disintegrins from snake venom have been developed that exhibit improved specificity and potency without causing the emergence of the LIBS epitopes of integrin β3. The small-molecule RUC-1 and the second congener RUC-2 block fibrinogen binding to activated αIIbβ3, but do not induce major conformational changes in the protein β3 subunit nor the emergence of LIBSs and presumably MIBSs [32,35], and may not be antigenic and cause immune thrombocytopenia. RUC-1 interacts with αIIb, while RUC-2 interacts with the Mg2+ coordinating sites of β3, termed MIDAS (metal ion-dependent adhesion site) [36]. The RGD-containing disintegrin TMV-7 from snake venom and its more potent derivatives inhibit the ligand-binding function of integrins, platelet aggregation, and thrombus formation, and importantly do not cause major conformational changes in the integrin β3 subunit or induce integrin activation [27].
1.4.2. Integrin Antagonists that Target Integrin Outside-in Signaling
Emerging evidence has shown that mutational disruption or deletion of the talin-binding site prevented mice from thrombosis, but defected physiological hemostasis has been revealed to prolong tail bleeding times in vivo [37,38]. Since then, the inhibition of inside-out signaling has led to impairing the talin-driven integrin activation and its ligand-binding function, therefore, it is expected that these inside-out signaling inhibitors should show characteristics similar to current αIIbβ3 antagonists, which inhibit both thrombosis and hemostasis. Current conceptual advances in integrin outside-in signaling have shown the potential in developing selective inhibitors of integrin outside-in signaling as new antithrombotic drugs [31]. A permeable myristoylated ExE motif peptide was designed to selectively inhibit Gα13-mediated outside-in signaling, platelet spreading, and thrombus formation without causing excessive hemorrhage in vivo. Thus, selective inhibitors of outside-in signaling as a potential antithrombotic strategy may find the right balance between potent antithrombotic efficacy and bleeding adverse reactions to allow antithrombotic therapy with an appropriate control of bleeding risk.
1.5. Disintegrins as Anti-Inflammatory Agents
The immune system recognizes pathogen-related molecular patterns via a repertoire of pattern recognition receptors, among which the family of Toll-like receptors (TLRs) features prominently [39]. TLR4 recognizes lipopolysaccharide (LPS)-containing Gram-negative bacteria, while TLR2 recognizes the peptidoglycan of Gram-positive bacteria. The β2 and β3 integrins form heterodimers with an α subunit to regulate leukocyte trafficking and function [40]. Vitronectin and integrin αvβ3 contribute to the initiation of TLR2 responses to bacterial lipopeptide [41]. Our recent studies have indicated that the disintegrin rhodostomin (Rn) interacts with the αvβ3 integrin of monocytes/macrophages, leading to interference with the LPS-induced NFκB and MAPK pathways, and MyD88-dependent TLRs in the production of cytokines in phagocytes. Thus, Rn significantly represses pro-inflammatory cytokine and chemokine release, inhibits cell adhesion and migration in vitro, and elevates the survival rate of septic mice in the LPS-administration and cecal ligation puncture (CLP) model by attenuating the acute inflammatory activity caused by bacterial infections [42,43]. The capacity of Rn to reduce thrombus formation may be responsible for its antiplatelet activity through the αIIbβ3 blockade. The potential application of disintegrins, and their safer derivatives, as an inhibitor of TLR2 and TLR4 activation raises the possibility of drug development in inflammatory diseases caused by complicated microbial patterns.
1.6. Disintegrins as an Anti-Tumor and Anti-Angiogenesis Agents
Angiogenesis plays a vital role in normal physiological processes such as tissue repair, embryonic development, and luteal formation [44]. Integrin αvβ3 expressed on smooth muscle cells, endothelial cells, transformed cells, and fibroblasts modulates cell migration, proliferation, and has a great impact on restenosis, tumor cell migration, angiogenesis, and atherosclerosis [45]. Several classes of integrins recognize the RGD sequence present in extracellular matrix (ECM) proteins [46], resulting in linking cytoskeletal proteins with the ECM, and are involved in bidirectional signaling that changes cellular functions. Our previous studies have reported that some disintegrins inhibit adhesion between ECMs and tumor cells via blocking αvβ3 and α5β1 integrins [47,48,49,50]. These disintegrins, as well as the synthetic RGD-containing peptides, have been shown to inhibit the experimental metastasis of melanoma cells [51,52,53]. We also found that RGD containing the antiplatelet disintegrin trifavin inhibited B16F10 melanoma cell-induced lung colonization in an experimental model [54], and trigramin inhibited the cell-substratum adhesion of human melanoma cells, and spreading on fibronectin and fibrinogen [55], as well as that the rhodostomin inhibited the ancrod-generated fibrin-triggered prostaglandin I2 formation of human umbilical vein endothelial cells (HUVECs) through blocking αvβ3 integrin [56,57,58]. Rhodostomin was also found to cause the cleavage of β-catenin and poly (ADP-ribose) polymerase during the apoptosis of endothelial cells [59]. However, efforts to apply αvβ3-specific RGD-mimetics in tumor therapy are still under active investigation. Further development of the recombinant venom-derived disintegrin, along with new technologies looking at additional disintegrin-like proteins with pure antagonist specificity, may offer novel therapeutic approaches in targeting tumor-induced angiogenesis
1.7. Novel αIIbβ3 Antagonists Derived from Disintegrins
Life-threatening thrombocytopenia and bleeding risks limit clinical antithrombotic use in patients undergoing percutaneous coronary intervention (PCI), however, given the widespread use of these efficacious drugs and the relatively high incidence, ideal antithrombotic agents with minimal bleeding side effects are currently under active investigation. Recently, we found a unique RGD-containing disintegrin, TMV-7, purified from Trimeresurus mucrosquamatus venom, which, like RUC-2, binds to the domain lying between the αIIb and β3 subunits, mostly stabilized through strong interactions with the αIIb subunit, thus neither primes the platelets to bind the ligands nor causes a conformational change of β3 as identified with the ligand-induced binding site (LIBS) mAb AP5 [27]. TMV-7 has also been shown to be an efficacious antithrombotic in illumination-induced mesenteric venous thrombosis and FeCl3-induced carotid artery thrombosis models. At an efficacious antithrombotic dosage, TMV-7 did not increase bleeding risk in vivo [27]. Its unique mechanism of action may be related to targeting outside-in signaling without affecting the talin-driven inside-out signaling and platelet function (i.e., platelet fibrin adhesion and clot retraction) responsible for primary hemostasis. A recent report indicated that Gα13 and talin play vital roles in thrombin-induced αIIbβ3 bidirectional signaling and bind mutually exclusive sites of β3 cytoplasmic domain in opposing waves, demonstrating that selectively targeting integrin outside-in signaling allows for the inhibitory activity of thrombus formation, while maintaining physiological hemostasis in animal models [31,60,61]. Therefore, the elucidation of the docking structure of the disintegrin TMV-7-αIIbβ3 complex and the structure–activity relationship between TMV-7 and αIIbβ3 on a molecular level may provide clues for the drug development of an ideal antithrombotic R(K)GD-mimetic with a better safety profile. Furthermore, TMV-7 inhibits LPS-TLR4 ligation-mediated release of pro-inflammatory mediators by downregulating reactive oxygen species (ROS) production and FAK/NFκB/MAPK axis signaling through the blockade of integrin αVβ3 in RAW 264.7 macrophages and human THP-1 cells [62]. Furthermore, TMV-7 significantly reduced myocardial ischemia-reperfusion (I-R)-induced arrhythmias, infarct volume, as well as mortality, via downregulating apoptotic proteins Bax and Caspase-3 in rats with I-R injury, demonstrating that TMV-7 is also possibly a potent antiarrhythmic agent with cardio-protective properties [63].
1.8. Other Possible Applications of Disintegrins
It has been suggested that the intracellular changes during ischemia trigger the accumulation of reactive oxygen species (ROS) and metabolic intermediates. The abnormal increase of ROS is associated with oxidative stress, and implicated in ischemic stroke and acute myocardial infarction [64,65,66], resulting in potential damage to proteins, lipids, and nucleic acids. Both of these diseases are also caused by arterial thrombosis. Current clinical trials have examined a combination of fibrinolytic therapy by using αIIbβ3 antagonists and a recombinant tissue-type plasminogen activator [67,68,69], and indicate that αIIbβ3 antagonists may have a beneficial effect by reducing adverse outcomes caused by strokes, although there is a risk of increased bleeding, especially with Abciximab [69]. Therefore, whether a pure αIIbβ3 antagonist, instead of the partial antagonist, can be revaluated in this add-on therapeutic strategy on stroke treatment remains to be examined.
2. Conclusions
The journey of snake venom disintegrin research has been initiated since the first naturally occurring disintegrin was discovered [7] and has inspired many studies in the molecular interaction of R(K)GD-containing disintegrins with integrin αIIbβ3, αvβ3, and other integrins, leading to the fruitful discovery of potential therapeutic agents in field of inflammation, angiogenesis, tumor metastasis, and atherothrombotic diseases (i.e., acute coronary syndrome, myocardial infarction, or ischemia stroke). With the aid of advanced molecular biology techniques, and the elucidation of the physiological and pathological roles of integrins, we can utilize these disintegrins and their derivatives to target specific integrins for the further evaluation of their potential efficacy and the associated adverse reaction mechanisms. Several potential antiplatelet strategies have shown considerable promise toward preventing thrombus formation while sparing hemorrhage. These include inhibitors of the αIIbβ3 outside-in signaling [31], conformation-specific targeting of αIIbβ3 [27,32,70], and a competitive inhibitor of the fibrin D-dimer region of monomeric Glycoprotein VI [71,72]. In keeping with the concept that the differential targeting of the thrombus formation may preserve physiological hemostasis, all of these inhibitors do not seem to affect the initial adhesion of platelets in vivo and thus allow for the formation of a platelet hemostatic core in response to vascular injury. The most challenging task is how to design a safer therapeutic agent, namely a pure integrin antagonist, which specifically blocks the αIIbβ3/αvβ3 target without intrinsically activating or changing the conformation into an activated state, leading to bleeding, thrombocytopenia/activation of inflammation/angiogenesis.
The discovery of novel therapeutic antithrombotics, especially the small-mass RGD-mimetic drugs derived from naturally-occurring disintegrins, may be accelerated through clarifying the detailed ligand-receptor docking, structural biology of integrins, and ligand-integrin complex aided by X-ray crystallography. Despite the challenges of translating promising animal data into the clinic, the emergence of novel antithrombotic targets holds the promise of improving cardiovascular outcomes, minimizing iatrogenic bleeding complications, and significantly increasing the number of patients who are afforded protection by antithrombotic drugs. Likewise, these safety-improved integrin antagonists derived from disintegrins can be widely used in patients afflicted with integrin-related diseases, such as ischemia stroke, septic inflammation, angiogenesis, and tumors.
Author Contributions
Review concept and design: Y.-J.K., T.-F.H.; Acquisition of data: Y.-J.K., C.-H.C.; Analysis and interpretation of data: Y.-J.K., C.-H.C., T.-F.H.; Drafting of the manuscript: Y.-J.K.; Study supervision: T.-F.H. had full access to all the data in the review and take responsibility for the integrity of the manuscript. All authors read and approved the final manuscript.
Funding
These studies were supported by the Ministry of Science and Technology of Taiwan [grant number: 107-2320-B-715-004-MY3]; Mackay Medical College [grant number: MMC-1071B2 and 1072B3].
Acknowledgments
We acknowledge grants from the Ministry of Science and Technology of Taiwan MOST 107-2320-B-715-004.
Conflicts of Interest
The authors declare that they have no conflict of interest.
References
- Hynes, R.O. Integrins: A family of cell surface receptors. Cell 1987, 48, 549–554. [Google Scholar] [CrossRef]
- Hynes, R.O. Integrins: Versatility, modulation, and signaling in cell adhesion. Cell 1992, 69, 11–25. [Google Scholar] [CrossRef]
- Ginsberg, M.H.; Loftus, J.C.; Plow, E.F. Cytoadhesins, integrins, and platelets. Thromb. Haemost. 1988, 59, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Watson, S.P.; Auger, J.M.; McCarty, O.J.; Pearce, A.C. Gpvi and integrin alphaiib beta3 signaling in platelets. J. Thromb. Haemost. 2005, 3, 1752–1762. [Google Scholar] [CrossRef] [PubMed]
- Coller, B.S. Platelets and thrombolytic therapy. N. Engl. J. Med. 1990, 322, 33–42. [Google Scholar] [CrossRef] [PubMed]
- Stein, B.; Fuster, V.; Halperin, J.L.; Chesebro, J.H. Antithrombotic therapy in cardiac disease. An emerging approach based on pathogenesis and risk. Circulation 1989, 80, 1501–1513. [Google Scholar] [CrossRef] [PubMed]
- Huang, T.F.; Holt, J.C.; Lukasiewicz, H.; Niewiarowski, S. Trigramin. A low molecular weight peptide inhibiting fibrinogen interaction with platelet receptors expressed on glycoprotein iib-iiia complex. J. Biol. Chem. 1987, 262, 16157–16163. [Google Scholar]
- Huang, T.F.; Ouyang, C. Action mechanism of the potent platelet aggregation inhibitor from trimeresurus gramineus snake venom. Thromb. Res. 1984, 33, 125–138. [Google Scholar] [CrossRef]
- Huang, T.F.; Liu, C.Z.; Ouyang, C.H.; Teng, C.M. Halysin, an antiplatelet arg-gly-asp-containing snake venom peptide, as fibrinogen receptor antagonist. Biochem. Pharm. 1991, 42, 1209–1219. [Google Scholar]
- Huang, T.F.; Wu, Y.J.; Ouyang, C. Characterization of a potent platelet aggregation inhibitor from agkistrodon rhodostoma snake venom. Biochim. Biophys. Acta 1987, 925, 248–257. [Google Scholar] [CrossRef]
- Gan, Z.R.; Gould, R.J.; Jacobs, J.W.; Friedman, P.A.; Polokoff, M.A. Echistatin. A potent platelet aggregation inhibitor from the venom of the viper, echis carinatus. J. Biol. Chem. 1988, 263, 19827–19832. [Google Scholar] [PubMed]
- Huang, T.F.; Holt, J.C.; Kirby, E.P.; Niewiarowski, S. Trigramin: Primary structure and its inhibition of von willebrand factor binding to glycoprotein iib/iiia complex on human platelets. Biochemistry 1989, 28, 661–666. [Google Scholar] [CrossRef]
- Cook, J.J.; Huang, T.F.; Rucinski, B.; Strzyzewski, M.; Tuma, R.F.; Williams, J.A.; Niewiarowski, S. Inhibition of platelet hemostatic plug formation by trigramin, a novel rgd-peptide. Am. J. Physiol. 1989, 256, H1038–H1043. [Google Scholar] [CrossRef] [PubMed]
- Musial, J.; Niewiarowski, S.; Rucinski, B.; Stewart, G.J.; Cook, J.J.; Williams, J.A.; Edmunds, L.H., Jr. Inhibition of platelet adhesion to surfaces of extracorporeal circuits by disintegrins. Rgd-containing peptides from viper venoms. Circulation 1990, 82, 261–273. [Google Scholar] [CrossRef] [PubMed]
- Rote, W.E.; Mu, D.X.; Roncinske, R.A.; Frelinger, A.L., 3rd; Lucchesi, B.R. Prevention of experimental carotid artery thrombosis by applaggin. J. Pharm. Exp. 1993, 267, 809–814. [Google Scholar]
- Scarborough, R.M.; Rose, J.W.; Hsu, M.A.; Phillips, D.R.; Fried, V.A.; Campbell, A.M.; Nannizzi, L.; Charo, I.F. Barbourin. A gpiib-iiia-specific integrin antagonist from the venom of sistrurus m. Barbouri. J. Biol. Chem. 1991, 266, 9359–9362. [Google Scholar] [PubMed]
- Scarborough, R.M.; Rose, J.W.; Naughton, M.A.; Phillips, D.R.; Nannizzi, L.; Arfsten, A.; Campbell, A.M.; Charo, I.F. Characterization of the integrin specificities of disintegrins isolated from american pit viper venoms. J. Biol. Chem. 1993, 268, 1058–1065. [Google Scholar] [PubMed]
- Scarborough, R.M.; Naughton, M.A.; Teng, W.; Rose, J.W.; Phillips, D.R.; Nannizzi, L.; Arfsten, A.; Campbell, A.M.; Charo, I.F. Design of potent and specific integrin antagonists. Peptide antagonists with high specificity for glycoprotein iib-iiia. J. Biol. Chem. 1993, 268, 1066–1073. [Google Scholar]
- Gould, R.J.; Polokoff, M.A.; Friedman, P.A.; Huang, T.F.; Holt, J.C.; Cook, J.J.; Niewiarowski, S. Disintegrins: A family of integrin inhibitory proteins from viper venoms. Proc. Soc. Exp. Biol. Med. 1990, 195, 168–171. [Google Scholar] [CrossRef]
- Coller, B.S.; Peerschke, E.I.; Scudder, L.E.; Sullivan, C.A. A murine monoclonal antibody that completely blocks the binding of fibrinogen to platelets produces a thrombasthenic-like state in normal platelets and binds to glycoproteins iib and/or iiia. J. Clin. Investig. 1983, 72, 325–338. [Google Scholar] [CrossRef]
- Coller, B.S.; Scudder, L.E. Inhibition of dog platelet function by in vivo infusion of f (ab’)2 fragments of a monoclonal antibody to the platelet glycoprotein iib/iiia receptor. Blood 1985, 66, 1456–1459. [Google Scholar] [PubMed]
- Chew, D.P.; Bhatt, D.L.; Sapp, S.; Topol, E.J. Increased mortality with oral platelet glycoprotein iib/iiia antagonists: A meta-analysis of phase iii multicenter randomized trials. Circulation 2001, 103, 201–206. [Google Scholar] [CrossRef] [PubMed]
- Bledzka, K.; Smyth, S.S.; Plow, E.F. Integrin alphaiibbeta3: From discovery to efficacious therapeutic target. Circ. Res. 2013, 112, 1189–1200. [Google Scholar] [CrossRef]
- Bassand, J.P. Current antithrombotic agents for acute coronary syndromes: Focus on bleeding risk. Int. J. Cardiol. 2013, 163, 5–18. [Google Scholar] [CrossRef] [PubMed]
- Chong, B.H. Drug-induced thrombocytopenia: Mibs trumps libs. Blood 2012, 119, 6177–6178. [Google Scholar] [CrossRef] [PubMed]
- Gao, C.; Boylan, B.; Bougie, D.; Gill, J.C.; Birenbaum, J.; Newman, D.K.; Aster, R.H.; Newman, P.J. Eptifibatide-induced thrombocytopenia and thrombosis in humans require fcgammariia and the integrin beta3 cytoplasmic domain. J. Clin. Invest. 2009, 119, 504–511. [Google Scholar] [CrossRef] [PubMed]
- Kuo, Y.J.; Chen, Y.R.; Hsu, C.C.; Peng, H.C.; Huang, T.F. An alphaiib beta3 antagonist prevents thrombosis without causing fc receptor gamma-chain iia-mediated thrombocytopenia. J. Thromb. Haemost. 2017, 15, 2230–2244. [Google Scholar] [CrossRef]
- Reese, J.A.; Li, X.; Hauben, M.; Aster, R.H.; Bougie, D.W.; Curtis, B.R.; George, J.N.; Vesely, S.K. Identifying drugs that cause acute thrombocytopenia: An analysis using 3 distinct methods. Blood 2010, 116, 2127–2133. [Google Scholar] [CrossRef]
- Aster, R.H.; Curtis, B.R.; McFarland, J.G.; Bougie, D.W. Drug-induced immune thrombocytopenia: Pathogenesis, diagnosis, and management. J. Thromb. Haemost. 2009, 7, 911–918. [Google Scholar] [CrossRef]
- Hanna, E.B.; Rao, S.V.; Manoukian, S.V.; Saucedo, J.F. The evolving role of glycoprotein iib/iiia inhibitors in the setting of percutaneous coronary intervention strategies to minimize bleeding risk and optimize outcomes. JACC Cardiovasc. Interv. 2010, 3, 1209–1219. [Google Scholar] [CrossRef]
- Shen, B.; Zhao, X.; O’Brien, K.A.; Stojanovic-Terpo, A.; Delaney, M.K.; Kim, K.; Cho, J.; Lam, S.C.; Du, X. A directional switch of integrin signalling and a new anti-thrombotic strategy. Nature 2013, 503, 131–135. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Negri, A.; Li, J.; Naini, S.; Coller, B.S.; Filizola, M. Structure-based virtual screening of small-molecule antagonists of platelet integrin alphaiibbeta3 that do not prime the receptor to bind ligand. J. Comput. Aided Mol. Des. 2012, 26, 1005–1015. [Google Scholar] [CrossRef] [PubMed]
- Bougie, D.W.; Rasmussen, M.; Zhu, J.; Aster, R.H. Antibodies causing thrombocytopenia in patients treated with rgd-mimetic platelet inhibitors recognize ligand-specific conformers of alphaiib/beta3 integrin. Blood 2012, 119, 6317–6325. [Google Scholar] [CrossRef]
- Frelinger, A.L., 3rd; Du, X.P.; Plow, E.F.; Ginsberg, M.H. Monoclonal antibodies to ligand-occupied conformers of integrin alpha iib beta 3 (glycoprotein iib-iiia) alter receptor affinity, specificity, and function. J. Biol. Chem. 1991, 266, 17106–17111. [Google Scholar]
- Zhu, J.; Zhu, J.; Negri, A.; Provasi, D.; Filizola, M.; Coller, B.S.; Springer, T.A. Closed headpiece of integrin alphaiibbeta3 and its complex with an alphaiibbeta3-specific antagonist that does not induce opening. Blood 2010, 116, 5050–5059. [Google Scholar] [CrossRef] [PubMed]
- Zhu, J.; Choi, W.S.; McCoy, J.G.; Negri, A.; Zhu, J.; Naini, S.; Li, J.; Shen, M.; Huang, W.; Bougie, D.; et al. Structure-guided design of a high-affinity platelet integrin alphaiibbeta3 receptor antagonist that disrupts mg(2)(+) binding to the midas. Sci. Transl. Med. 2012, 4, 125ra132. [Google Scholar] [CrossRef] [PubMed]
- Nieswandt, B.; Moser, M.; Pleines, I.; Varga-Szabo, D.; Monkley, S.; Critchley, D.; Fassler, R. Loss of talin1 in platelets abrogates integrin activation, platelet aggregation, and thrombus formation in vitro and in vivo. J. Exp. Med. 2007, 204, 3113–3118. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Petrich, B.G.; Fogelstrand, P.; Partridge, A.W.; Yousefi, N.; Ablooglu, A.J.; Shattil, S.J.; Ginsberg, M.H. The antithrombotic potential of selective blockade of talin-dependent integrin alpha iib beta 3 (platelet gpiib-iiia) activation. J. Clin. Investig. 2007, 117, 2250–2259. [Google Scholar] [CrossRef]
- West, A.P.; Koblansky, A.A.; Ghosh, S. Recognition and signaling by toll-like receptors. Annu. Rev. Cell Dev. Biol. 2006, 22, 409–437. [Google Scholar] [CrossRef]
- Ley, K.; Laudanna, C.; Cybulsky, M.I.; Nourshargh, S. Getting to the site of inflammation: The leukocyte adhesion cascade updated. Nat. Rev. Immunol. 2007, 7, 678–689. [Google Scholar] [CrossRef] [PubMed]
- Gerold, G.; Abu Ajaj, K.; Bienert, M.; Laws, H.J.; Zychlinsky, A.; de Diego, J.L. A toll-like receptor 2-integrin beta3 complex senses bacterial lipopeptides via vitronectin. Nat. Immunol. 2008, 9, 761–768. [Google Scholar] [CrossRef] [PubMed]
- Hsu, C.C.; Chuang, W.J.; Chang, C.H.; Tseng, Y.L.; Peng, H.C.; Huang, T.F. Improvements in endotoxemic syndromes using a disintegrin, rhodostomin, through integrin alphavbeta3-dependent pathway. J. Thromb. Haemost. 2011, 9, 593–602. [Google Scholar] [CrossRef] [PubMed]
- Hsu, C.C.; Chuang, W.J.; Chung, C.H.; Chang, C.H.; Peng, H.C.; Huang, T.F. Snake venom disintegrin inhibits the activation of toll-like receptors and alleviates sepsis through integrin alphavbeta3 blockade. Sci. Rep. 2016, 6, 23387. [Google Scholar] [CrossRef] [PubMed]
- Folkman, J.; Shing, Y. Angiogenesis. J. Biol. Chem. 1992, 267, 10931–10934. [Google Scholar] [PubMed]
- Jin, H.; Varner, J. Integrins: Roles in cancer development and as treatment targets. Br. J. Cancer 2004, 90, 561–565. [Google Scholar] [CrossRef] [PubMed]
- Ruoslahti, E. Integrins. J. Clin. Investig. 1991, 87, 1–5. [Google Scholar] [CrossRef]
- Sheu, J.R.; Lin, C.H.; Huang, T.F. Triflavin, an antiplatelet peptide, inhibits tumor cell-extracellular matrix adhesion through an arginine-glycine-aspartic acid-dependent mechanism. J. Lab. Clin. Med. 1994, 123, 256–263. [Google Scholar]
- Sheu, J.R.; Huang, T.F. Triflavin, an arg-gly-asp-containing peptide, inhibits b16-f10 mouse melanoma cell adhesion to matrix proteins via direct binding to tumor cells. J. Biomed. Sci. 1996, 3, 359–364. [Google Scholar] [CrossRef]
- Chiang, H.S.; Yang, R.S.; Huang, T.F. The arg-gly-asp-containing peptide, rhodostomin, inhibits in vitro cell adhesion to extracellular matrices and platelet aggregation caused by saos-2 human osteosarcoma cells. Br. J. Cancer 1995, 71, 265–270. [Google Scholar] [CrossRef]
- Sheu, J.R.; Lin, C.H.; Peng, H.C.; Huang, T.F. Triflavin, an arg-gly-asp-containing peptide, inhibits the adhesion of tumor cells to matrix proteins via binding to multiple integrin receptors expressed on human hepatoma cells. Proc. Soc. Exp. Biol. Med. 1996, 213, 71–79. [Google Scholar] [CrossRef] [PubMed]
- Rucinski, B.; Niewiarowski, S.; Holt, J.C.; Soszka, T.; Knudsen, K.A. Batroxostatin, an arg-gly-asp-containing peptide from bothrops atrox, is a potent inhibitor of platelet aggregation and cell interaction with fibronectin. Biochim. Biophys. Acta 1990, 1054, 257–262. [Google Scholar] [CrossRef]
- Soszka, T.; Knudsen, K.A.; Beviglia, L.; Rossi, C.; Poggi, A.; Niewiarowski, S. Inhibition of murine melanoma cell-matrix adhesion and experimental metastasis by albolabrin, an rgd-containing peptide isolated from the venom of trimeresurus albolabris. Exp. Cell Res. 1991, 196, 6–12. [Google Scholar] [CrossRef]
- Morris, V.L.; Schmidt, E.E.; Koop, S.; MacDonald, I.C.; Grattan, M.; Khokha, R.; McLane, M.A.; Niewiarowski, S.; Chambers, A.F.; Groom, A.C. Effects of the disintegrin eristostatin on individual steps of hematogenous metastasis. Exp. Cell Res. 1995, 219, 571–578. [Google Scholar] [CrossRef] [PubMed]
- Sheu, J.R.; Lin, C.H.; Chung, J.L.; Teng, C.M.; Huang, T.F. Triflavin, an arg-gly-asp-containing antiplatelet peptide inhibits cell-substratum adhesion and melanoma cell-induced lung colonization. Jpn. J. Cancer Res. 1992, 83, 885–893. [Google Scholar] [CrossRef] [PubMed]
- Knudsen, K.A.; Tuszynski, G.P.; Huang, T.F.; Niewiarowski, S. Trigramin, an rgd-containing peptide from snake venom, inhibits cell-substratum adhesion of human melanoma cells. Exp. Cell Res. 1988, 179, 42–49. [Google Scholar] [CrossRef]
- Chang, M.C.; Wang, B.R.; Huang, T.F. Characterization of endothelial cell differential attachment to fibrin and fibrinogen and its inhibition by arg-gly-asp-containing peptides. Thromb. Haemost. 1995, 74, 764–769. [Google Scholar] [CrossRef]
- Chang, M.C.; Jeng, J.H.; Cheong, T.C.; Huang, T.F. The morphologic change of endothelial cells by ancrod-generated fibrin is triggered by alpha v beta 3 integrin binding and the subsequent activation of a g-protein coupled phospholipase c. Biochim. Biophys. Acta 1995, 1269, 115–121. [Google Scholar] [CrossRef]
- Chang, M.C.; Yang, R.S.; Lin, C.H.; Huang, T.F. Integrin alpha v beta 3 and phospholipase c regulate prostacyclin formation of endothelial cells caused by ancrod-generated fibrin. Eur. J. Pharm. 1996, 297, 129–136. [Google Scholar] [CrossRef]
- Wu, W.B.; Peng, H.C.; Huang, T.F. Disintegrin causes proteolysis of beta-catenin and apoptosis of endothelial cells. Involvement of cell-cell and cell-ecm interactions in regulating cell viability. Exp. Cell Res. 2003, 286, 115–127. [Google Scholar] [CrossRef]
- Tadokoro, S.; Shattil, S.J.; Eto, K.; Tai, V.; Liddington, R.C.; de Pereda, J.M.; Ginsberg, M.H.; Calderwood, D.A. Talin binding to integrin beta tails: A final common step in integrin activation. Science 2003, 302, 103–106. [Google Scholar] [CrossRef] [PubMed]
- Gong, H.; Shen, B.; Flevaris, P.; Chow, C.; Lam, S.C.; Voyno-Yasenetskaya, T.A.; Kozasa, T.; Du, X. G protein subunit galpha13 binds to integrin alphaiibbeta3 and mediates integrin “outside-in” signaling. Science 2010, 327, 340–343. [Google Scholar] [CrossRef] [PubMed]
- Hung, Y.C.; Hsu, C.C.; Chung, C.H.; Huang, T.F. The disintegrin, trimucrin, suppresses lps-induced activation of phagocytes primarily through blockade of nf-kappab and mapk activation. Naunyn. Schmiedeberg’s Arch. Pharm. 2016, 389, 723–737. [Google Scholar] [CrossRef] [PubMed]
- Hung, Y.C.; Kuo, Y.J.; Huang, S.S.; Huang, T.F. Trimucrin, an arg-gly-asp containing disintegrin, attenuates myocardial ischemia-reperfusion injury in murine by inhibiting platelet function. Eur. J. Pharm. 2017, 813, 24–32. [Google Scholar] [CrossRef] [PubMed]
- Becatti, M.; Fiorillo, C.; Gori, A.M.; Marcucci, R.; Paniccia, R.; Giusti, B.; Violi, F.; Pignatelli, P.; Gensini, G.F.; Abbate, R. Platelet and leukocyte ros production and lipoperoxidation are associated with high platelet reactivity in non-st elevation myocardial infarction (nstemi) patients on dual antiplatelet treatment. Atherosclerosis 2013, 231, 392–400. [Google Scholar] [CrossRef] [PubMed]
- Davis, S.M.; Pennypacker, K.R. Targeting antioxidant enzyme expression as a therapeutic strategy for ischemic stroke. Neurochem. Int. 2017, 107, 23–32. [Google Scholar] [CrossRef]
- Jamasbi, J.; Ayabe, K.; Goto, S.; Nieswandt, B.; Peter, K.; Siess, W. Platelet receptors as therapeutic targets: Past, present and future. Thromb. Haemost. 2017, 117, 1249–1257. [Google Scholar] [CrossRef]
- Siebler, M.; Hennerici, M.G.; Schneider, D.; von Reutern, G.M.; Seitz, R.J.; Rother, J.; Witte, O.W.; Hamann, G.; Junghans, U.; Villringer, A.; et al. Safety of tirofiban in acute ischemic stroke: The satis trial. Stroke 2011, 42, 2388–2392. [Google Scholar] [CrossRef]
- Pancioli, A.M.; Adeoye, O.; Schmit, P.A.; Khoury, J.; Levine, S.R.; Tomsick, T.A.; Sucharew, H.; Brooks, C.E.; Crocco, T.J.; Gutmann, L.; et al. Combined approach to lysis utilizing eptifibatide and recombinant tissue plasminogen activator in acute ischemic stroke-enhanced regimen stroke trial. Stroke 2013, 44, 2381–2387. [Google Scholar] [CrossRef]
- Eckert, B.; Koch, C.; Thomalla, G.; Kucinski, T.; Grzyska, U.; Roether, J.; Alfke, K.; Jansen, O.; Zeumer, H. Aggressive therapy with intravenous abciximab and intra-arterial rtpa and additional pta/stenting improves clinical outcome in acute vertebrobasilar occlusion: Combined local fibrinolysis and intravenous abciximab in acute vertebrobasilar stroke treatment (fast): Results of a multicenter study. Stroke 2005, 36, 1160–1165. [Google Scholar]
- Armstrong, P.C.; Peter, K. Gpiib/iiia inhibitors: From bench to bedside and back to bench again. Thromb. Haemost. 2012, 107, 808–814. [Google Scholar] [CrossRef] [PubMed]
- Onselaer, M.B.; Hardy, A.T.; Wilson, C.; Sanchez, X.; Babar, A.K.; Miller, J.L.C.; Watson, C.N.; Watson, S.K.; Bonna, A.; Philippou, H.; et al. Fibrin and d-dimer bind to monomeric gpvi. Blood Adv. 2017, 1, 1495–1504. [Google Scholar] [CrossRef] [PubMed]
- Alshehri, O.M.; Hughes, C.E.; Montague, S.; Watson, S.K.; Frampton, J.; Bender, M.; Watson, S.P. Fibrin activates gpvi in human and mouse platelets. Blood 2015, 126, 1601–1608. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Current αIIbβ3 antagonists are effective antithrombotics, but have a significant bleeding risk. Schematic representation of the antithrombotic strategy selectively inhibiting outside-in signaling without causing integrin activation nor affecting the processes of primary hemostasis (i.e., platelet adhesion and fibrin clot retraction), thus these potential antithrombotics do not increase bleeding risk and have greater safety profiles than the current αIIbβ3 antagonists (i.e., Eptifibatide) [31].
Figure 1.
Current αIIbβ3 antagonists are effective antithrombotics, but have a significant bleeding risk. Schematic representation of the antithrombotic strategy selectively inhibiting outside-in signaling without causing integrin activation nor affecting the processes of primary hemostasis (i.e., platelet adhesion and fibrin clot retraction), thus these potential antithrombotics do not increase bleeding risk and have greater safety profiles than the current αIIbβ3 antagonists (i.e., Eptifibatide) [31].

© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Kuo, Y.-J.; Chung, C.-H.; Huang, T.-F. From Discovery of Snake Venom Disintegrins to A Safer Therapeutic Antithrombotic Agent. Toxins 2019, 11, 372. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins11070372
AMA Style
Kuo Y-J, Chung C-H, Huang T-F. From Discovery of Snake Venom Disintegrins to A Safer Therapeutic Antithrombotic Agent. Toxins. 2019; 11(7):372. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins11070372
Chicago/Turabian StyleKuo, Yu-Ju, Ching-Hu Chung, and Tur-Fu Huang. 2019. "From Discovery of Snake Venom Disintegrins to A Safer Therapeutic Antithrombotic Agent" Toxins 11, no. 7: 372. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins11070372
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.