Cytotoxic and Proapoptotic Activity of Sanguinarine, Berberine, and Extracts of Chelidonium majus L. and Berberis thunbergii DC. toward Hematopoietic Cancer Cell Lines

 , , and
, , and
Abstract
:1. Introduction
2. Results
2.1. Sanguinarine, Berberine, and Chelidonium majus L. Extract Exhibit Cytotoxic Activity against Hematopoietic Cell Lines
2.2. Sanguinarine, Berberine, and Chelidionium majus L. Extract Affect the Expression of Genes Related to Apoptosis
2.2.1. U266B1 Cell Line
2.2.2. CEM/C1 Cell Line
2.2.3. CCRF/CEM Cell Line
2.2.4. HL60 Cell Line
2.2.5. HL60/MX1 Cell Line
2.2.6. HL60/MX2 Cell Line
2.2.7. J45.01 Cell Line
2.3. Clustering and PARAFAC Analysis
3. Discussion
4. Materials and Methods
4.1. Sanguinarine and Berberine Stock Solutions
4.2. Plant Extracts Preparation
4.3. Cell Lines
4.4. Trypan Blue Staining
4.5. RNA Isolation
4.6. cDNA Synthesis
4.7. Real-Time PCR
4.8. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A

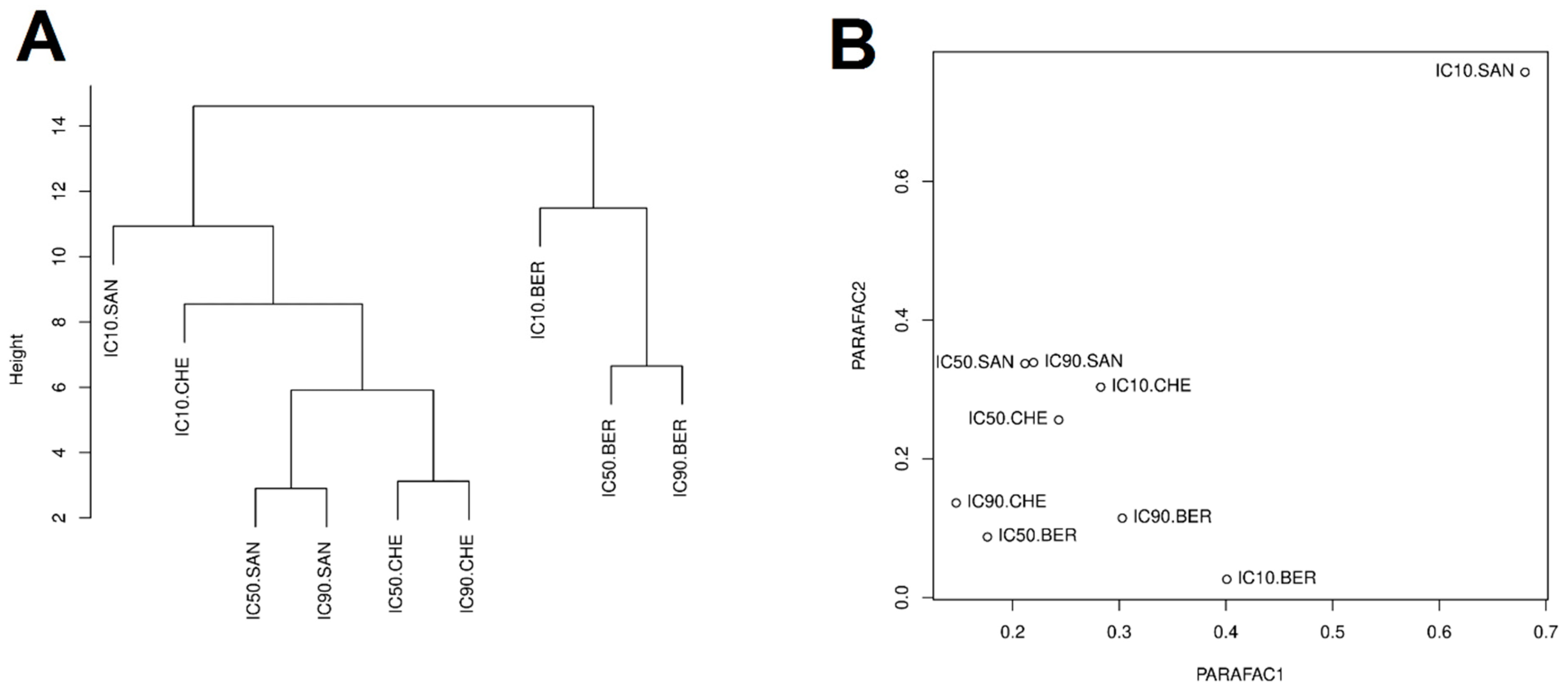{kind=link}
{kind=link}
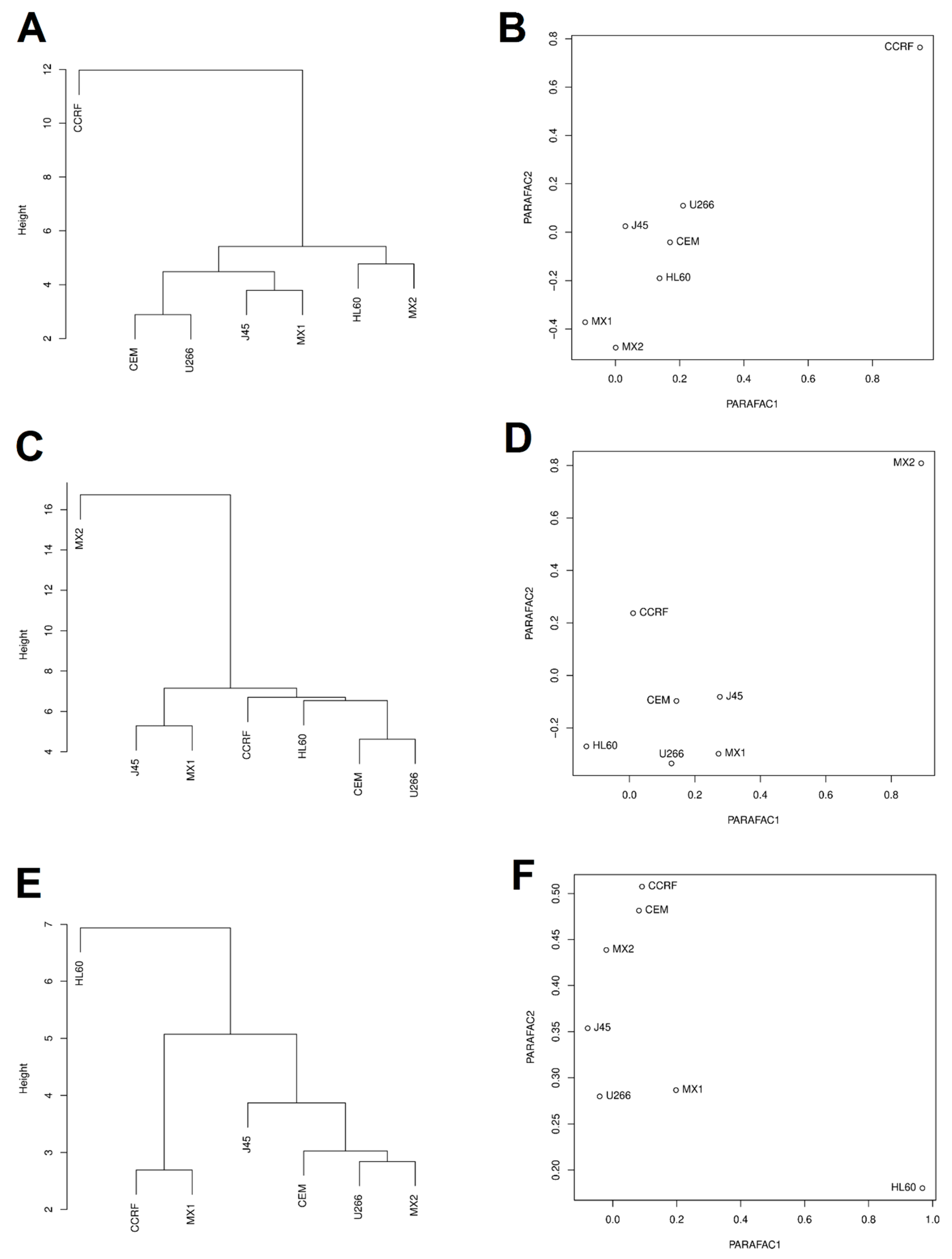{kind=link}
| Gene Symbol | Probe | Gene Name |
|---|---|---|
| BAK1 | Hs00940249_m1 | BCL2 antagonist/killer 1 |
| BAX | Hs00180363_m1 | BCL2 associated X, apoptosis regulator |
| BCL2 | Hs00608023_m1 | BCL2 apoptosis regulator |
| MCL1 | Hs01050896_m1 | MCL1 apoptosis regulator, BCL2 family member |
| BCL2L1 | Hs00236329_m1 | BCL2 like 1 |
| BCL2L2 | Hs00187848_m1 | BCL2 like 2 |
| BID | Hs01026792_m1 | BH3 interacting domain death agonist |
| BIK | Hs00154189_m1 | BCL2 interacting killer |
| BNIP1 | Hs00241824_m1 | BCL2 interacting protein 1 |
| BNIP2 | Hs00188939_m1 | BCL2 interacting protein 2 |
| BNIP3 | Hs00969291_m1 | BCL2 interacting protein 3 |
| PMAIP1 | Hs00560402_m1 | Phorbol-12-myristate-13-acetate-induced protein 1 |
| BAD | Hs00188930_m1 | BCL2 associated agonist of cell death |
| CASP3 | Hs00234387_m1 | Caspase 3 |
| CASP8 | Hs01018151_m1 | Caspase 8 |
| CASP9 | Hs00154261_m1 | Caspase 9 |
| TP53 | Hs01034249_m1 | Tumor protein p53 |
| GAPDH (Endogenous control) | Hs99999905_m1 | Glyceraldehyde-3-phosphate dehydrogenase |
References
- Trujillo-Villanueva, K.; Rubio-Piña, J.; Monforte-González, M.; Ramírez-Benítez, E.; Vázquez-Flota, F. The sequential exposure to jasmonate, salicylic acid and yeast extract promotes sanguinarine accumulation in Argemone mexicana cell cultures. Biotechnol. Lett. 2012, 34, 379–385. [Google Scholar] [CrossRef] [PubMed]
- Cho, H.Y.; Rhee, H.S.; Yoon, S.Y.; Park, J.M. Differential induction of protein expression and benzophenanthridine alkaloid accumulation in Eschscholtzia californica suspension cultures by methyl jasmonate and yeast extract. J. Microbiol. Biotechnol. 2008, 18, 255–262. [Google Scholar] [PubMed]
- Obiang-Obounou, B.W.; Kang, O.H.; Choi, J.G.; Keum, J.H.; Kim, S.B.; Mun, S.H.; Shin, D.W.; Kim, K.W.; Park, C.B.; Kim, Y.G.; et al. The mechanism of action of sanguinarine against methicillin-resistant Staphylococcus aureus. J. Toxicol. Sci. 2011, 36, 277–283. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Q.; Dai, P.; Bao, H.; Liang, P.; Wang, W.; Xing, A.; Sun, J. Anti-inflammatory and neuroprotective effects of sanguinarine following cerebral ischemia in rats. Exp. Ther. Med. 2017, 13, 263–268. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.; Sun, X.; Huang, K.; Shen, S.; Lin, X.; Xie, Z.; Wang, J.; Fan, S.; Ma, J.; Zhao, X. Sanguinarine protects against osteoarthritis by suppressing the expression of catabolic proteases. Oncotarget 2017, 8, 62900–62913. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, S.S.; Kai, M.; Lee, M.K. Inhibitory effects of sanguinarine on monoamine oxidase activity in mouse brain. Phytother. Res. 2001, 15, 167–169. [Google Scholar] [CrossRef] [PubMed]
- Jursky, F.; Baliova, M. Differential effect of the benzophenanthridine alkaloids sanguinarine and chelerythrine on glycine transporters. Neurochem. Int. 2011, 58, 641–647. [Google Scholar] [CrossRef] [PubMed]
- Singh, R.; Mackraj, I.; Naidoo, R.; Gathiram, P. Sanguinarine downregulates AT1a gene expression in a hypertensive rat model. J. Cardiovasc. Pharmacol. 2006, 48, 14–21. [Google Scholar] [CrossRef] [PubMed]
- Jeng, J.H.; Wu, H.L.; Lin, B.R.; Lan, W.H.; Chang, H.H.; Ho, Y.S.; Lee, P.H.; Wang, Y.J.; Wang, J.S.; Chen, Y.J.; et al. Antiplatelet effect of sanguinarine is correlated to calcium mobilization, thromboxane and cAMP production. Atherosclerosis 2007, 191, 250–258. [Google Scholar] [CrossRef]
- Kuznetsova, L.P.; Nikol’skaya, E.B.; Sochilina, E.E.; Faddeeva, M.D. Inhibition of Human Blood Acetylcholinesterase and Butyrylcholinesterase by Some Alkaloids. J. Evol. Biochem. Physiol. 2002, 38, 35–39. [Google Scholar] [CrossRef]
- Zajoncova, L.; Kosina, P.; Vicar, J.; Ulrichová, J.; Pec, P. Study of the inhibition of alpha-amylase by the benzo[c]phenanthridine alkaloids sanguinarine and chelerythrine. J. Enzym. Inhib. Med. Chem. 2005, 20, 261–267. [Google Scholar] [CrossRef] [PubMed]
- Imenshahidi, M.; Hosseinzadeh, H. Berberine and barberry (Berberis vulgaris): A clinical review. Phytother. Res. 2019, 33, 504–523. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Wang, Y.; Yan, L.; Liang, R.M.; Dai, B.D.; Tang, R.J.; Gao, P.H.; Jiang, Y.Y. Proteomic analysis reveals a synergistic mechanism of fluconazole and berberine against fluconazole-resistant Candida albicans: Endogenous ROS augmentation. J. Proteome Res. 2009, 8, 5296–5304. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Geng, Y.N.; Jiang, J.D.; Kong, W.J. Antioxidant and anti-inflammatory activities of berberine in the treatment of diabetes mellitus. Evid. Based Complement. Altern. Med. 2014, 2014, 289264. [Google Scholar] [CrossRef] [PubMed]
- Chang, W.; Chen, L.; Hatch, G.M. Berberine as a therapy for type 2 diabetes and its complications: From mechanism of action to clinical studies. Biochem. Cell Biol. 2015, 93, 479–486. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Liu, J.; Ma, A.; Chen, Y. Cardioprotective effect of berberine against myocardial ischemia/reperfusion injury via attenuating mitochondrial dysfunction and apoptosis. Int. J. Clin. Exp. Med. 2015, 8, 14513–14519. [Google Scholar] [PubMed]
- Fan, J.; Zhang, K.; Jin, Y.; Li, B.; Gao, S.; Zhu, J.; Cui, R. Pharmacological effects of berberine on mood disorders. J. Cell Mol. Med. 2019, 23, 21–28. [Google Scholar] [CrossRef] [PubMed]
- Kongkiatpaiboon, S.; Duangdee, N.; Prateeptongkum, S.; Chaijaroenkul, W. Acetylcholinesterase Inhibitory Activity of Alkaloids Isolated from Stephania venosa. Nat. Prod. Commun. 2016, 11, 1805–1806. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Galadari, S.; Rahman, A.; Pallichankandy, S.; Thayyullathil, F. Molecular targets and anticancer potential of sanguinarine—A benzophenanthridine alkaloid. Phytomedicine 2017, 34, 143–153. [Google Scholar] [CrossRef] [PubMed]
- Gaziano, R.; Moroni, G.; Buè, C.; Miele, M.T.; Sinibaldi-Vallebona, P.; Pica, F. Antitumor effects of the benzophenanthridine alkaloid sanguinarine: Evidence and perspectives. World J. Gastrointest. Oncol. 2016, 8, 30–39. [Google Scholar] [CrossRef] [PubMed]
- Hussain, A.R.; Al-Jomah, N.A.; Siraj, A.K.; Manogaran, P.; Al-Hussein, K.; Abubaker, J.; Platanias, L.C.; Al-Kuraya, K.S.; Uddin, S. Sanguinarine-dependent induction of apoptosis in primary effusion lymphoma cells. Cancer Res. 2007, 67, 3888–3897. [Google Scholar] [CrossRef] [PubMed]
- Weerasinghe, P.; Hallock, S.; Tang, S.C.; Liepins, A. Role of Bcl-2 family proteins and caspase-3 in sanguinarine-induced bimodal cell death. Cell Biol. Toxicol. 2001, 17, 371–381. [Google Scholar] [CrossRef] [PubMed]
- Weerasinghe, P.; Hallock, S.; Tang, S.C.; Liepins, A. Sanguinarine induces bimodal cell death in K562 but not in high Bcl-2-expressing JM1 cells. Pathol. Res. Pract. 2001, 197, 717–726. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.; Lee, T.J.; Leem, J.; Choi, K.S.; Park, J.W.; Kwon, T.K. Sanguinarine-induced apoptosis: Generation of ROS, down-regulation of Bcl-2, c-FLIP, and synergy with TRAIL. J. Cell Biochem. 2008, 104, 895–907. [Google Scholar] [CrossRef] [PubMed]
- Park, S.Y.; Jin, M.L.; Kim, Y.H.; Lee, S.J.; Park, G. Sanguinarine inhibits invasiveness and the MMP-9 and COX-2 expression in TPA-induced breast cancer cells by inducing HO-1 expression. Oncol. Rep. 2014, 31, 497–504. [Google Scholar] [CrossRef] [PubMed]
- Basini, G.; Bussolati, S.; Santini, S.E.; Grasselli, F. Sanguinarine inhibits VEGF-induced angiogenesis in a fibrin gel matrix. Biofactors 2007, 29, 11–18. [Google Scholar] [CrossRef]
- Basini, G.; Santini, S.E.; Bussolati, S.; Grasselli, F. Sanguinarine inhibits VEGF-induced Akt phosphorylation. Ann. N. Y. Acad. Sci. 2007, 1095, 371–376. [Google Scholar] [CrossRef]
- Eun, J.P.; Koh, G.Y. Suppression of angiogenesis by the plant alkaloid, sanguinarine. Biochem. Biophys. Res. Commun. 2004, 317, 618–624. [Google Scholar] [CrossRef]
- Zou, K.; Li, Z.; Zhang, Y.; Zhang, H.Y.; Li, B.; Zhu, W.L.; Shi, J.Y.; Jia, Q.; Li, Y.M. Advances in the study of berberine and its derivatives: A focus on anti-inflammatory and anti-tumor effects in the digestive system. Acta Pharmacol. Sin. 2017, 38, 157–167. [Google Scholar] [CrossRef]
- Yan, K.; Zhang, C.; Feng, J.; Hou, L.; Yan, L.; Zhou, Z.; Liu, Z.; Liu, C.; Fan, Y.; Zheng, B.; et al. Induction of G1 cell cycle arrest and apoptosis by berberine in bladder cancer cells. Eur. J. Pharmacol. 2011, 661, 1–7. [Google Scholar] [CrossRef]
- Goto, H.; Kariya, R.; Shimamoto, M.; Kudo, E.; Taura, M.; Katano, H.; Okada, S. Antitumor effect of berberine against primary effusion lymphoma via inhibition of NF-κB pathway. Cancer Sci. 2012, 103, 775–781. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Liu, Q.; Xu, B.; Wu, J.; Guo, C.; Zhu, F.; Yang, Q.; Gao, G.; Gong, Y.; Shao, C. Berberine induces p53-dependent cell cycle arrest and apoptosis of human osteosarcoma cells by inflicting DNA damage. Mutat. Res. 2009, 662, 75–83. [Google Scholar] [CrossRef] [PubMed]
- Jin, F.; Xie, T.; Huang, X.; Zhao, X. Berberine inhibits angiogenesis in glioblastoma xenografts by targeting the VEGFR2/ERK pathway. Pharm. Biol. 2018, 56, 665–671. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, J.; Kang, M.; Wen, Q.; Qin, Y.T.; Wei, Z.X.; Xiao, J.J.; Wang, R.S. Berberine sensitizes nasopharyngeal carcinoma cells to radiation through inhibition of Sp1 and EMT. Oncol. Rep. 2017, 37, 2425–2432. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Yu, H.; Zhang, C.; Cheng, Y.; Hu, L.; Meng, X.; Zhao, Y. Protective effects of berberine on radiation-induced lung injury via intercellular adhesion molecular-1 and transforming growth factor-beta-1 in patients with lung cancer. Eur. J. Cancer 2008, 44, 2425–2432. [Google Scholar] [CrossRef] [PubMed]
- Singh, N.; Sharma, B. Toxicological Effects of Berberine and Sanguinarine. Front. Mol. Biosci. 2018, 5, 21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Das, M.; Ansari, K.M.; Dhawan, A.; Shukla, Y.; Khanna, S.K. Correlation of DNA damage in epidemic dropsy patients to carcinogenic potential of argemone oil and isolated sanguinarine alkaloid in mice. Int. J. Cancer 2005, 117, 709–717. [Google Scholar] [CrossRef] [PubMed]
- Dixit, R.; Srivastava, P.; Basu, S.; Srivastava, P.; Mishra, P.K.; Shukla, V.K. Association of mustard oil as cooking media with carcinoma of the gallbladder. J. Gastrointest. Cancer 2013, 44, 177–181. [Google Scholar] [CrossRef]
- Hossain, M.; Kabir, A.; Suresh Kumar, G. Binding of the anticancer alkaloid sanguinarine with tRNA (phe): Spectroscopic and calorimetric studies. J. Biomol. Struct. Dyn. 2012, 30, 223–234. [Google Scholar] [CrossRef]
- Adhami, V.M.; Aziz, M.H.; Mukhtar, H.; Ahmad, N. Activation of prodeath Bcl-2 family proteins and mitochondrial apoptosis pathway by sanguinarine in immortalized human HaCaT keratinocytes. Clin. Cancer Res. 2003, 9, 3176–3182. [Google Scholar]
- Selvi, B.R.; Pradhan, S.K.; Shandilya, J.; Das, C.; Sailaja, B.S.; Shankar, G.N.; Gadad, S.S.; Reddy, A.; Dasgupta, D.; Kundu, T.K. Sanguinarine interacts with chromatin, modulates epigenetic modifications, and transcription in the context of chromatin. Chem. Biol. 2009, 16, 203–216. [Google Scholar] [CrossRef] [PubMed]
- Maiti, M.; Kumar, G.S. Polymorphic nucleic Acid binding of bioactive isoquinoline alkaloids and their role in cancer. J. Nucleic Acids 2010, 2010, 593408. [Google Scholar] [CrossRef] [PubMed]
- Ji, X.; Sun, H.; Zhou, H.; Xiang, J.; Tang, Y.; Zhao, C. The interaction of telomeric DNA and C-myc22 G-quadruplex with 11 natural alkaloids. Nucleic Acid Ther. 2012, 22, 127–136. [Google Scholar] [CrossRef] [PubMed]
- Choy, C.S.; Cheah, K.P.; Chiou, H.Y.; Li, J.S.; Liu, Y.H.; Yong, S.F.; Chiu, W.T.; Liao, J.W.; Hu, C.M. Induction of hepatotoxicity by sanguinarine is associated with oxidation of protein thiols and disturbance of mitochondrial respiration. J. Appl. Toxicol. 2008, 28, 945–956. [Google Scholar] [CrossRef] [PubMed]
- Ferraroni, M.; Bazzicalupi, C.; Bilia, A.R.; Gratteri, P. X-Ray diffraction analyses of the natural isoquinoline alkaloids Berberine and Sanguinarine complexed with double helix DNA d(CGTACG). Chem. Commun. (Camb.) 2011, 47, 4917–4919. [Google Scholar] [CrossRef] [PubMed]
- Maiti, M.; Kumar, G.S. Molecular aspects on the interaction of protoberberine, benzophenanthridine, and aristolochia group of alkaloids with nucleic acid structures and biological perspectives. Med. Res. Rev. 2007, 27, 649–695. [Google Scholar] [CrossRef] [PubMed]
- Li, X.L.; Hu, Y.J.; Wang, H.; Yu, B.Q.; Yue, H.L. Molecular spectroscopy evidence of berberine binding to DNA: Comparative binding and thermodynamic profile of intercalation. Biomacromolecules 2012, 13, 873–880. [Google Scholar] [CrossRef] [PubMed]
- Hu, X.; Wu, X.; Huang, Y.; Tong, Q.; Takeda, S.; Qing, Y. Berberine induces double-strand DNA breaks in Rev3 deficient cells. Mol. Med. Rep. 2014, 9, 1883–1888. [Google Scholar] [CrossRef] [Green Version]
- Yuan, Z.Y.; Lu, X.; Lei, F.; Chai, Y.S.; Wang, Y.G.; Jiang, J.F.; Feng, T.S.; Wang, X.P.; Yu, X.; Yan, X.J.; et al. TATA boxes in gene transcription and poly (A) tails in mRNA stability: New perspective on the effects of berberine. Sci. Rep. 2015, 5, 18326. [Google Scholar] [CrossRef] [Green Version]
- Chai, Y.S.; Yuan, Z.Y.; Lei, F.; Wang, Y.G.; Hu, J.; Du, F.; Lu, X.; Jiang, J.F.; Xing, D.M.; Du, L.J. Inhibition of retinoblastoma mRNA degradation through Poly (A) involved in the neuroprotective effect of berberine against cerebral ischemia. PLoS ONE 2014, 9, e90850. [Google Scholar] [CrossRef]
- Och, A.; Szewczyk, K.; Pecio, Ł.; Stochmal, A.; Załuski, D.; Bogucka-Kocka, A. UPLC-MS/MS Profile of Alkaloids with Cytotoxic Properties of Selected Medicinal Plants of the Berberidaceae and Papaveraceae Families. Oxid. Med. Cell. Longev. 2017, 2017, 9369872. [Google Scholar] [CrossRef] [PubMed]
- Aburai, N.; Yoshida, M.; Ohnishi, M.; Kimura, K. Sanguinarine as a potent and specific inhibitor of protein phosphatase 2C in vitro and induces apoptosis via phosphorylation of p38 in HL60 cells. Biosci. Biotechnol. Biochem. 2010, 74, 548–552. [Google Scholar] [CrossRef] [PubMed]
- Slaninová, I.; Slanina, J.; Táborská, E. Quaternary benzo[c]phenanthridine alkaloids—Novel cell permeant and red fluorescing DNA probes. Cytometry A 2007, 71, 700–708. [Google Scholar] [CrossRef] [PubMed]
- Bogucka-Kocka, A.; Zalewski, D. Qualitative and quantitative determination of main alkaloids of Chelidonium majus L. using thin-layer chromatographic-densitometric method. Acta Chromatogr. 2017, 29, 385–397. [Google Scholar] [CrossRef]
- Nadova, S.; Miadokova, E.; Alfoldiova, L.; Kopaskova, M.; Hasplova, K.; Hudecova, A.; Vaculcikova, D.; Gregan, F.; Cipak, L. Potential antioxidant activity, cytotoxic and apoptosis-inducing effects of Chelidonium majus L. extract on leukemia cells. Neuro Endocrinol. Lett. 2008, 29, 649–652. [Google Scholar] [PubMed]
- Zielińska, S.; Jezierska-Domaradzka, A.; Wójciak-Kosior, M.; Sowa, I.; Junka, A.; Matkowski, A.M. Greater Celandine’s Ups and Downs-21 Centuries of Medicinal Uses of Chelidonium majus From the Viewpoint of Today’s Pharmacology. Front. Pharmacol. 2018, 9, 299. [Google Scholar] [CrossRef] [PubMed]
- Hu, H.Y.; Li, K.P.; Wang, X.J.; Liu, Y.; Lu, Z.G.; Dong, R.H.; Guo, H.B.; Zhang, M.X. Set9, NF-κB, and µRNA-21 mediate berberine-induced apoptosis of human multiple myeloma cells. Acta Pharmacol. Sin. 2013, 34, 157–166. [Google Scholar] [CrossRef] [PubMed]
- Efferth, T.; Davey, M.; Olbrich, A.; Rücker, G.; Gebhart, E.; Davey, R. Activity of drugs from traditional Chinese medicine toward sensitive and MDR1-or MRP1-overexpressing multidrug-resistant human CCRF-CEM leukemia cells. Blood Cells Mol. Dis. 2002, 28, 160–168. [Google Scholar] [CrossRef]
- Bui, N.L.; Pandey, V.; Zhu, T.; Ma, L.; Basappa; Lobie, P.E. BAD phosphorylation as a target of inhibition in oncology. Cancer Lett. 2018, 415, 177–186. [Google Scholar] [CrossRef]
- Macher-Goeppinger, S.; Keith, M.; Hatiboglu, G.; Hohenfellner, M.; Schirmacher, P.; Roth, W.; Tagscherer, K.E. Expression and Functional Characterization of the BNIP3 Protein in Renal Cell Carcinomas. Transl. Oncol. 2017, 10, 869–875. [Google Scholar] [CrossRef]
- Singh, A.; Azad, M.; Shymko, M.D.; Henson, E.S.; Katyal, S.; Eisenstat, D.D.; Gibson, S.B. The BH3 only Bcl-2 family member BNIP3 regulates cellular proliferation. PLoS ONE 2018, 13, e0204792. [Google Scholar] [CrossRef] [PubMed]
- Bishayee, A.; Rao, D.V.; Bouchet, L.G.; Bolch, W.E.; Howell, R.W. Protection by DMSO against cell death caused by intracellularly localized iodine-125, iodine-131 and polonium-210. Radiat. Res. 2000, 153, 416–427. [Google Scholar] [CrossRef]
- Chomczynski, P.; Sacchi, N. The single-step method of RNA isolation by acid guanidinium thiocyanate-phenol-chloroform extraction: Twenty-something years on. Nat. Protoc. 2006, 1, 581–585. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Schmittgen, T.D.; Livak, K.J. Analyzing real-time PCR data by the comparative C (T) method. Nat. Protoc. 2008, 3, 1101–1108. [Google Scholar] [CrossRef] [PubMed]



| Cell Line | IC10 | IC50 | IC90 |
|---|---|---|---|
| Sanguinarine (µM ± SD) | |||
| J45.01 | 0.10 ± 0.04 | 0.50 ± 0.04 | 1.00 ± 0.05 |
| U266B1 | 0.80 ± 0.04 | 1.05 ± 0.05 | 1.80 ± 0.04 |
| HL-60 | 0.20 ± 0.03 | 0.60 ± 0.06 | 1.80 ± 0.03 |
| HL-60/MX1 | 0.15 ± 0.03 | 0.50 ± 0.04 | 1.80 ± 0.05 |
| HL-60/MX2 | 0.06 ± 0.05 | 0.10 ± 0.05 | 1.20 ± 0.05 |
| CCRF/CEM | 0.50 ± 0.04 | 0.70 ± 0.03 | 1.20 ± 0.03 |
| CEM/C1 | 0.30 ± 0.04 | 0.50 ± 0.04 | 1.00 ± 0.03 |
| Berberine (µM ± SD) | |||
| J45.01 | 25.15 ± 3.15 | 80.15 ± 4.65 | 200.80 ± 4.65 |
| U266B1 | 125.15 ± 2.68 | 240.45 ± 4.15 | 250.00 ± 1.10* |
| HL-60 | 50.32 ± 4.56 | 90.45 ± 5.83 | 250.00 ± 4.35* |
| HL-60/MX1 | 25.05 ± 2.13 | 110.05 ± 6.72 | 250.00 ± 3.15* |
| HL-60/MX2 | 75.25 ± 6.52 | 250.00 ± 2.15* | - |
| CCRF/CEM | 50.40 ± 1.18 | 80.00 ± 2.13 | 130.25 ± 1.18 |
| CEM/C1 | 50.25 ± 4.25 | 225.15 ± 5.25 | 250.00 ± 2.85* |
| Chelidonium majus L. extract (µg/mL ± SD) | |||
| J45.01 | 5.05 ± 2.15 | 12.25 ± 2.85 | 38.65 ± 5.23 |
| U266B1 | 8.05 ± 3.45 | 21.50 ± 5.65 | 264.50 ± 4.12 |
| HL-60 | 9.01 ± 2.35 | 13.82 ± 3.15 | 280.02 ± 6.15 |
| HL-60/MX1 | 7.81 ± 6.18 | 20.15 ± 4.16 | 202.11 ± 4.32 |
| HL-60/MX2 | 19.85 ± 5.68 | 64.50 ± 5.48 | 236.0 ± 4.82 |
| CCRF/CEM | 7.58 ± 2.89 | 10.75 ± 2.15 | 27.75 ± 1.63 |
| CEM/C1 | 7.33 ± 5.48 | 13.25 ± 3.23 | 27.30 ± 1.89 |
| Cell Line | IC10 | IC50 | IC90 |
|---|---|---|---|
| Sanguinarine | |||
| J45.01 | ↑(BNIP3), ↓(BCL2L2, TP53) | ↑(BCL2, BNIP3), ↓(BAD, TP53) | ↑(BCL2, BNIP3), ↓(TP53) |
| U266B1 | ↑(BAK1, BNIP3, CASP8, MCL1, PMAIP1) | ↑(BAK1, BCL2, BNIP3) | ↑(BNIP3) |
| HL-60 | ↑(PMAIP1, TP53) | ↑(BAD) | NA |
| HL-60/MX1 | ↑(BAK1, CASP8, PMAIP1), ↓(BCL2L2, BID) | ↑(BCL2), ↓(TP53) | ↓(BCL2L2, BNIP2, CASP3, TP53) |
| HL-60/MX2 | ↑(BAK1, BAX, BIK, CASP3, CASP8, MCL1, PMAIP1, TP53) | ↓(BAD, BNIP2, PMAIP1, TP53) | ↓(BAD, BNIP2, TP53) |
| CCRF/CEM | (B2M, BAD, BAK1, BAX, BCL2, BCL2L1, BCL2L2, BID, BIK, BNIP1, BNIP2, BNIP3, CASP3, CASP8, CASP9, MCL1, PMAIP1, TP53) | ↑(BAK1, BNIP3, CASP8, PMAIP1, TP53) | ↑(BAD, BAK1, BCL2, BNIP2, BNIP3, CASP8, CASP9, MCL1, PMAIP1, TP53) |
| CEM/C1 | ↑(BAD, BAK1, BAX, BCL2, BCL2L2, BNIP1, BNIP2, CASP3, CASP8, MCL1, PMAIP1, TP53) | ↑(BAK1, MCL1) | ↓(BNIP3) |
| Berberine | |||
| J45.01 | ↑(BAK1, BCL2, BCL2L2, BID, BNIP3, CASP9) | ↓(BCL2L2) | ↑(BNIP3, CASP9) |
| U266B1 | NA | ↑(BAK1, BNIP3, CASP9), ↓(BCL2L2, BNIP2, MCL1, TP53) | ↑(BAK1, BAX, BNIP3, CASP9), ↓(BCL2, BIK, BNIP2) |
| HL-60 | ↓(BAK1, BNIP2, TP53) | ↓(BAK1, BAX, BNIP2, BNIP3, TP53) | ↓(TP53) |
| HL-60/MX1 | ↑(BAD, BAK1, BCL2, BNIP3, CASP9, TP53), ↓(BNIP2, MCL1) | ↑(BCL2), ↓(BAK1, BNIP2, BNIP3, TP53) | ↑(BAK1, BCL2, BNIP3, CASP9, PMAIP1), ↓(BNIP2, CASP8, MCL1) |
| HL-60/MX2 | ↑(BAD, BAK1, BAX, BCL2, BCL2L1, BCL2L2, BID, BIK, BNIP1, BNIP2, BNIP3, CASP3, CASP8, CASP9, MCL1, PMAIP1, TP53) | ↑(BAX, BIK, CASP3, CASP9, MCL1, TP53) | NA |
| CCRF/CEM | ↑(TP53), ↓(BAD, BAK1, BNIP2) | ↑(TP53), ↓(BAD, BAK1) | ↑(TP53), ↓(BAD, BAK1) |
| CEM/C1 | NA | NA | ↑(B2M, BAK1, BAX, BCL2, BNIP3, CASP9, PMAIP1), ↓(BNIP2) |
| Chelidonium majus L. extract | |||
| J45.01 | ↑(BAK1, BCL2, BNIP3, CASP8, CASP9, MCL1, PMAIP1) | ↑(BNIP3), ↓(BAD, BCL2L2, TP53) | ↑(BNIP3), ↓(TP53) |
| U266B1 | ↑(BAK1) | ↑(BAK1, BCL2, BCL2L2, BNIP2) | ↑(BAK1) |
| HL-60 | ↑(BAK1, BCL2L2, MCL1) | ↑(BAK1) | ↓(TP53) |
| HL-60/MX1 | ↑(BAK1, BCL2L2, MCL1, TP53) | ↑(BAK1, TP53) | ↑(TP53) |
| HL-60/MX2 | ↑(BAK1, BAX, MCL1), ↓(TP53) | ↑(BAK1, BAX, CASP3, MCL1), ↓(TP53) | ↑(BAK1, BAX, MCL1) |
| CCRF/CEM | ↑(BCL2, BCL2L2, BNIP2, BNIP3, CASP8, MCL1, PMAIP1, TP53) | ↑(BAK1, BCL2L2, BIK, BNIP2, TP53) | ↑(TP53), ↓(BAD) |
| CEM/C1 | ↑(BAK1, BCL2L2, CASP8, MCL1, PMAIP3) | ↑(BAK1) | ↑(BAD, BAK1, BCL2L1, BCL2L2, BNIP2) |
| Berberis thunbergii DC. extract (145.5 µg/mL) | |||
| J45.01 | ↑(BAK1, BAX, BCL2, BID, BNIP1, BNIP3, CASP9), ↓(BNIP2, TP53) | ||
| U266B1 | ↓(BNIP3) | ||
| HL-60 | ↑(BAK1, BNIP3, CASP9, TP53), ↓(BCL2L1, BIK, BNIP2, CASP3, CASP8, MCLI) | ||
| HL-60/MX1 | ↑(BCL2), ↓(BNIP2, TP53) | ||
| HL-60/MX2 | ↑(BAX, BIK, CASP3, MCL1), ↓(TP53) | ||
| CCRF/CEM | NA | ||
| CEM/C1 | ↑(BCL2), ↓(BAD, BAK1, TP53) | ||
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Och, A.; Zalewski, D.; Komsta, Ł.; Kołodziej, P.; Kocki, J.; Bogucka-Kocka, A. Cytotoxic and Proapoptotic Activity of Sanguinarine, Berberine, and Extracts of Chelidonium majus L. and Berberis thunbergii DC. toward Hematopoietic Cancer Cell Lines. Toxins 2019, 11, 485. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins11090485
Och A, Zalewski D, Komsta Ł, Kołodziej P, Kocki J, Bogucka-Kocka A. Cytotoxic and Proapoptotic Activity of Sanguinarine, Berberine, and Extracts of Chelidonium majus L. and Berberis thunbergii DC. toward Hematopoietic Cancer Cell Lines. Toxins. 2019; 11(9):485. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins11090485
Chicago/Turabian StyleOch, Anna, Daniel Zalewski, Łukasz Komsta, Przemysław Kołodziej, Janusz Kocki, and Anna Bogucka-Kocka. 2019. "Cytotoxic and Proapoptotic Activity of Sanguinarine, Berberine, and Extracts of Chelidonium majus L. and Berberis thunbergii DC. toward Hematopoietic Cancer Cell Lines" Toxins 11, no. 9: 485. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins11090485