Degradation of Aflatoxin B1 by a Sustainable Enzymatic Extract from Spent Mushroom Substrate of Pleurotus eryngii

 , ,
, ,
Abstract
:1. Introduction
2. Results
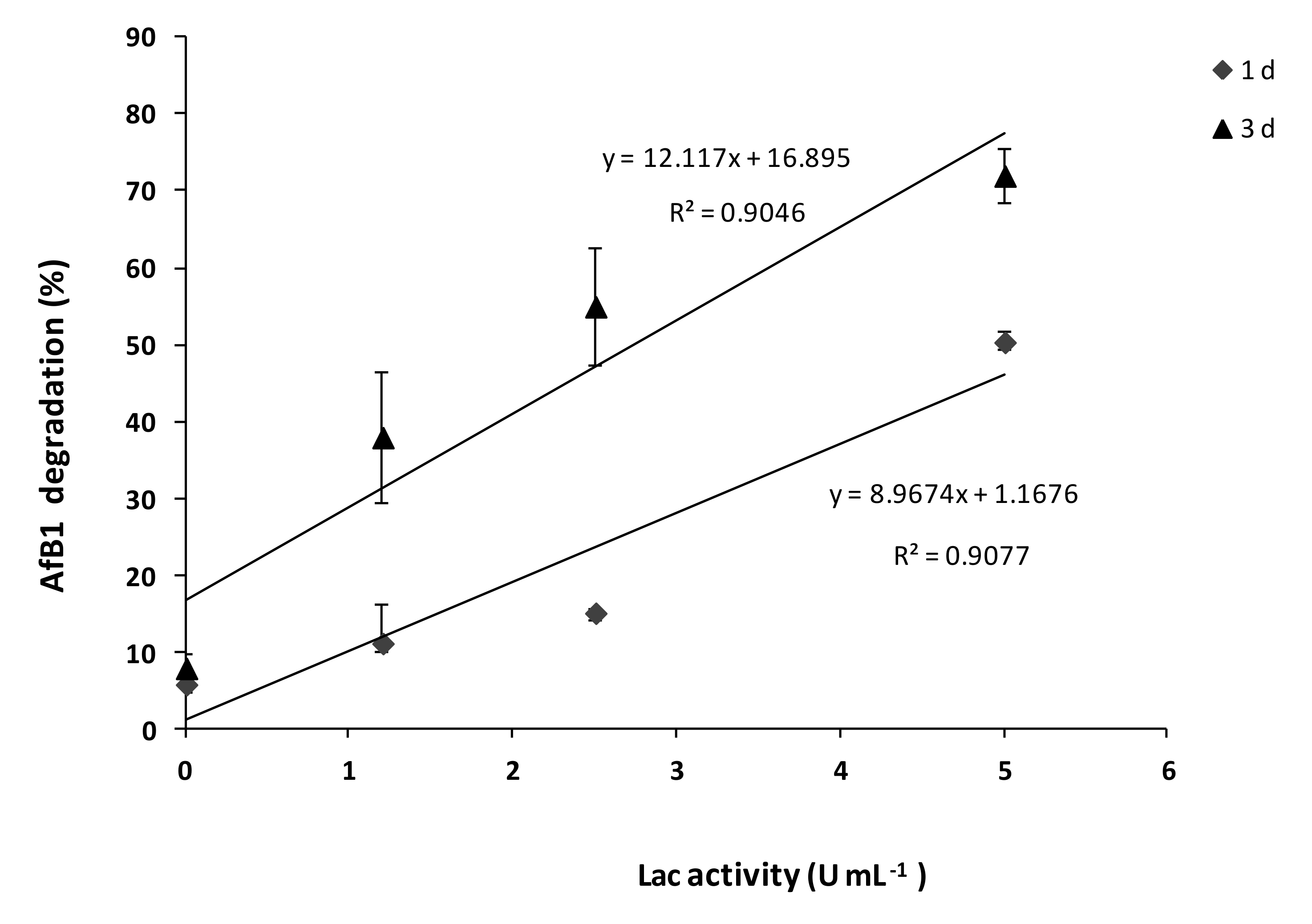
2.1. SMSE Enzymatic Activities
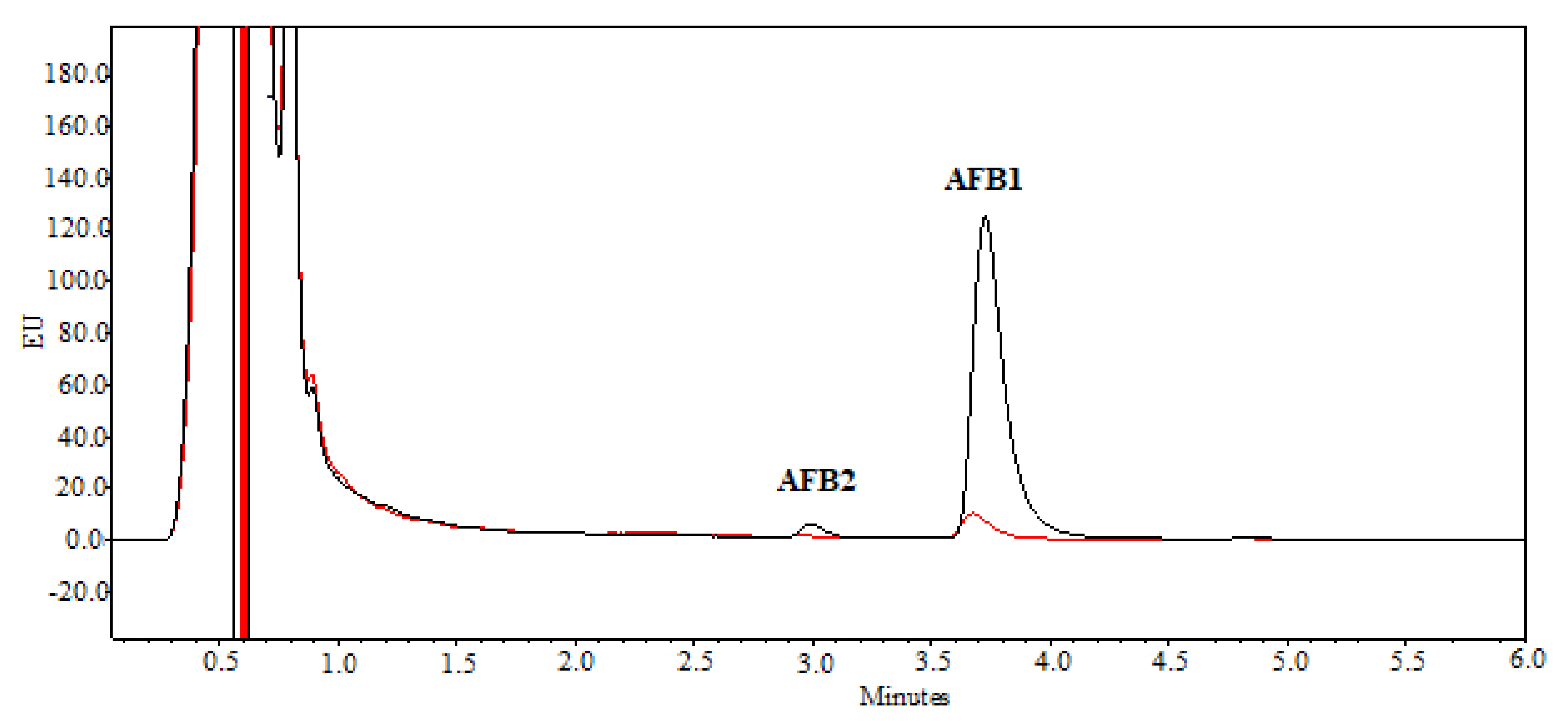
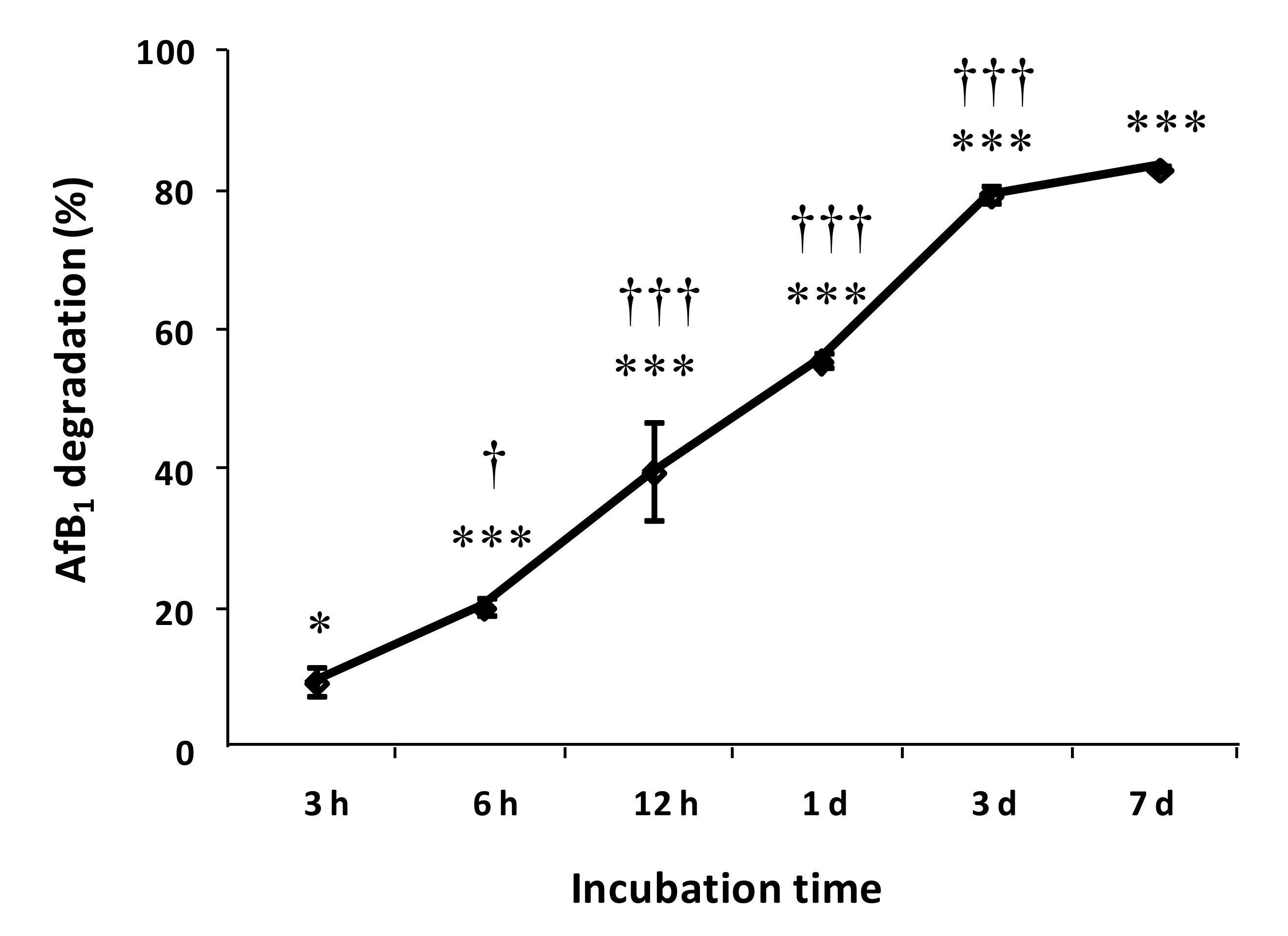
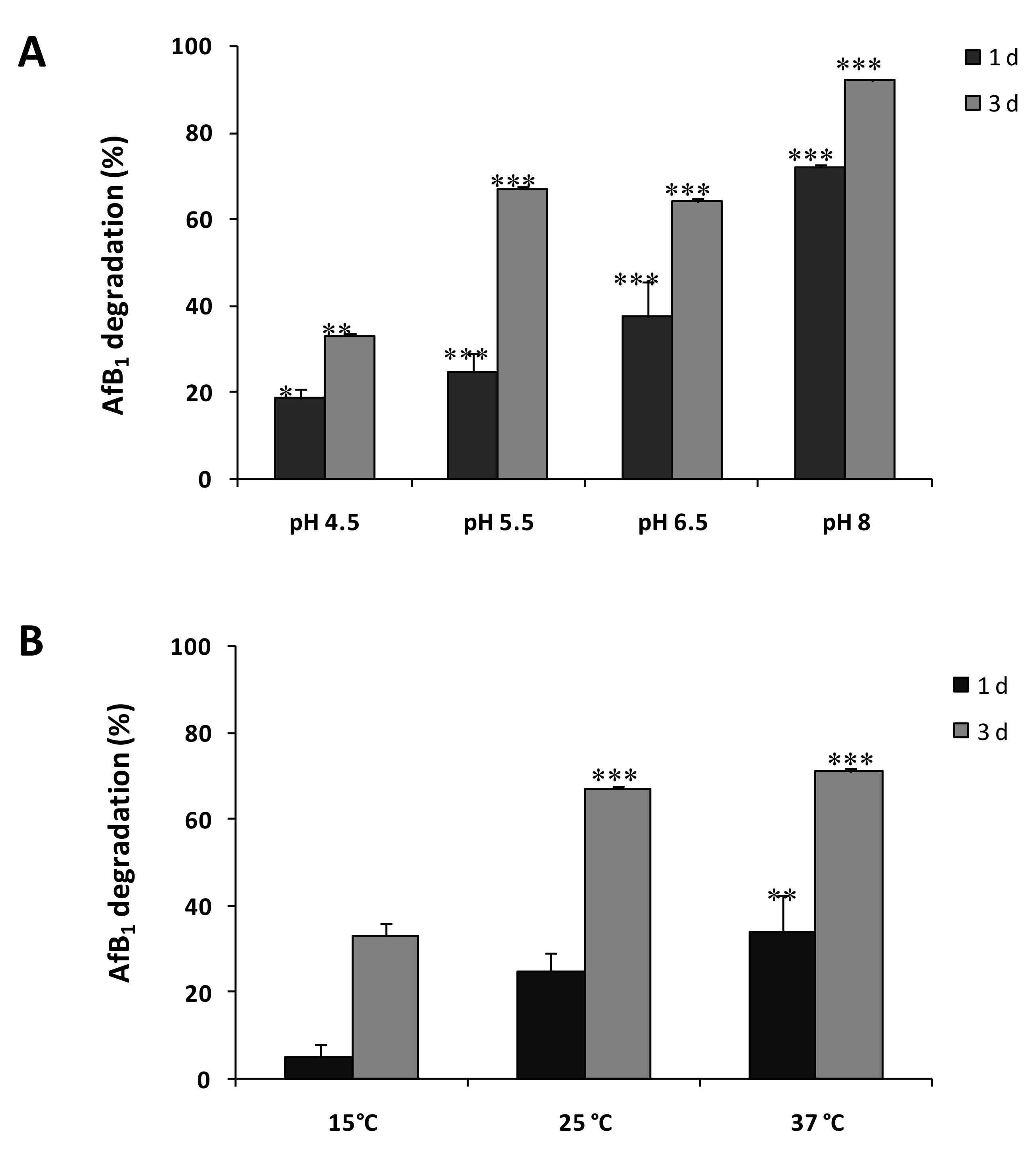
2.2. Degradation of AFB1 by SMSE
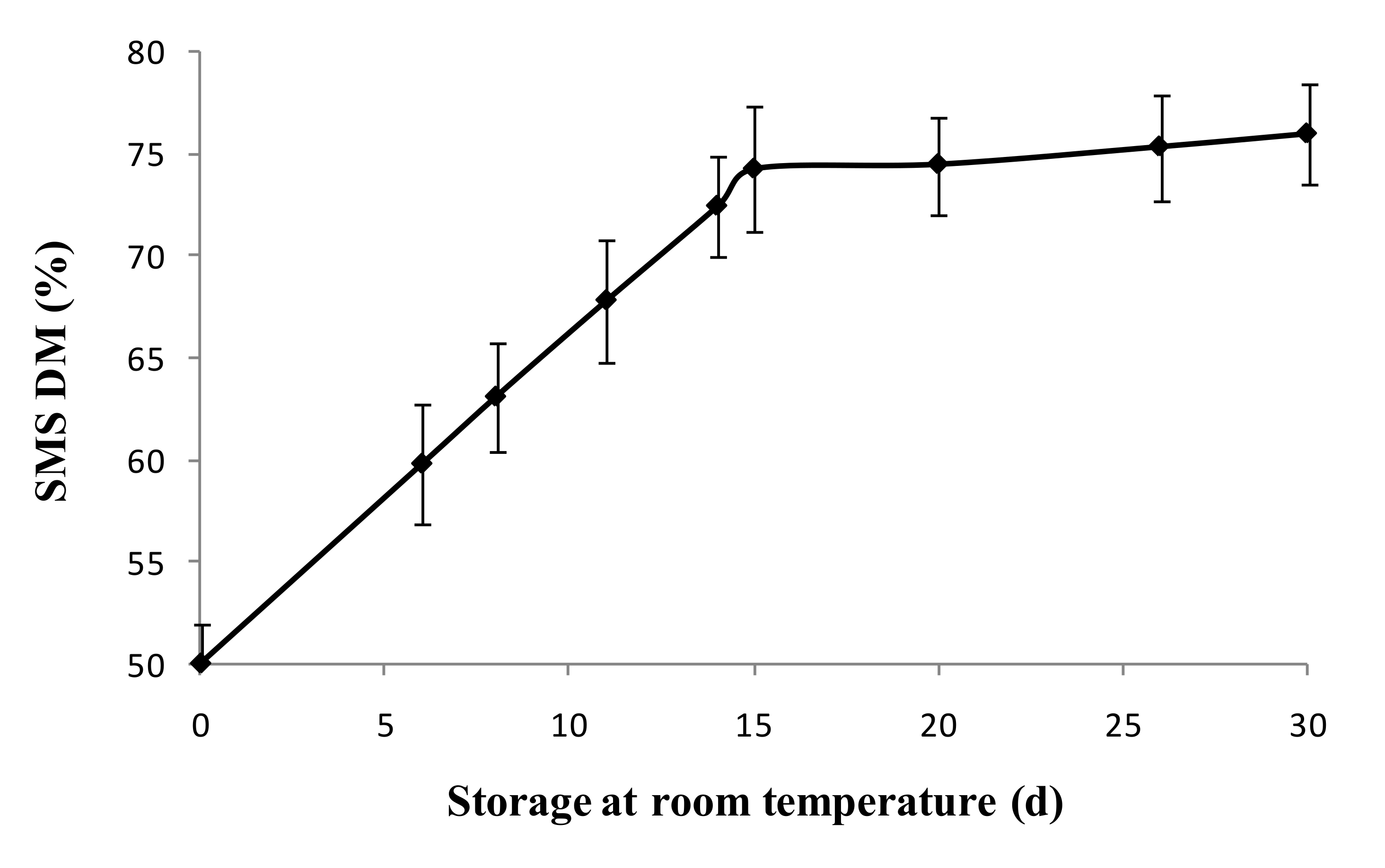
2.3. Storability of SMS
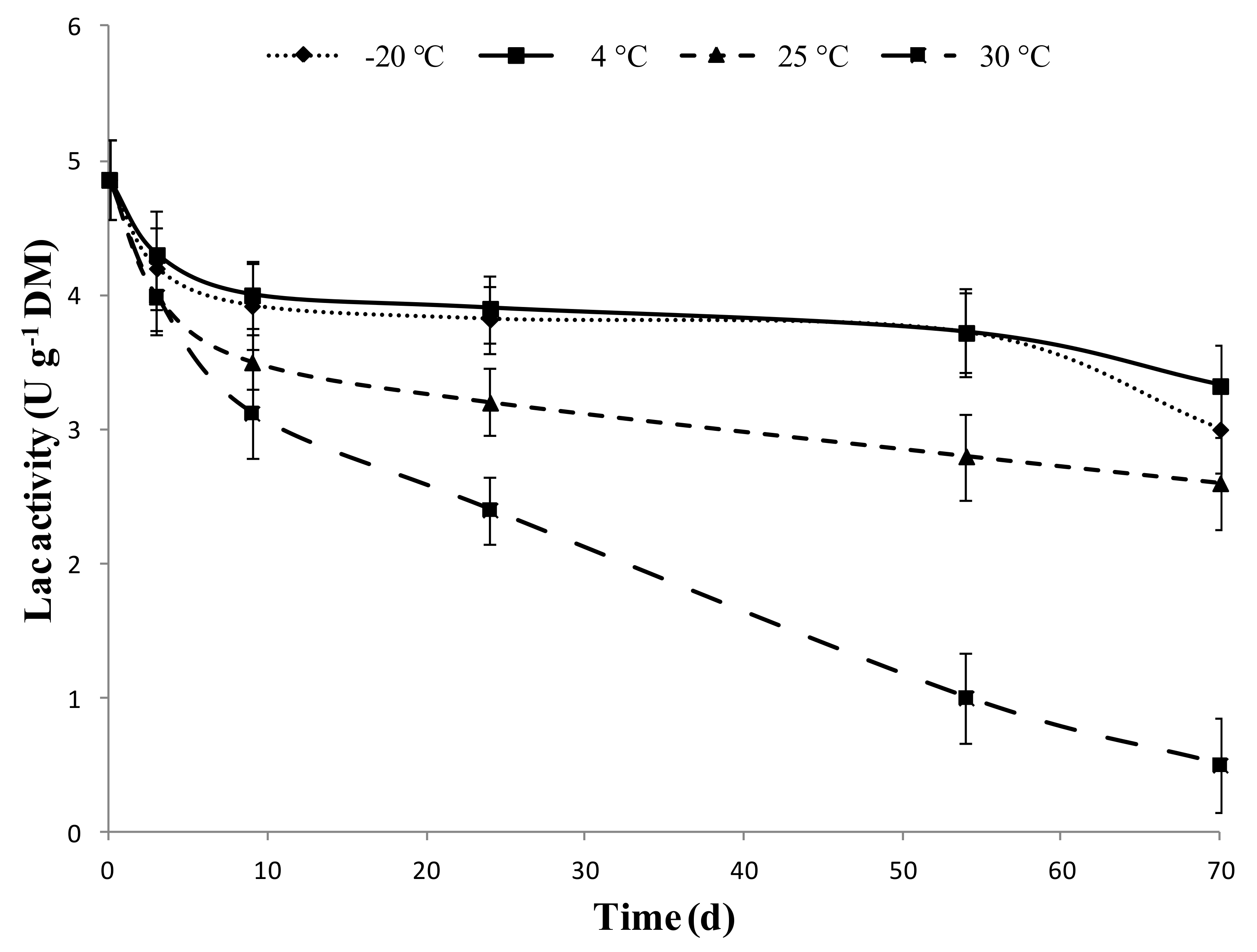
2.4. Storability of SMSE
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Chemicals
5.2. Enzymes Extraction
5.3. SMSE Enzymatic Activities
5.4. AFB1 Determination
5.5. Degradation of AFB1 by P. eryngii SMSE
5.6. Storability of SMS
5.7. Storability of SMSE
5.8. Statistical Analysis
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Williams, J.H.; Phillips, T.D.; Jolly, P.E.; Stiles, J.K.; Jolly, C.M.; Aggarwal, D. Human aflatoxicosis in developing countries: A review of toxicology, exposure, potential health consequences, and interventions. Am. J. Clin. Nutr. 2004, 80, 1106–1122. [Google Scholar] [CrossRef] [PubMed]
- International Agency for Research on Cancer (IARC) Aflatoxins. IARC Monographs on the Evaluation of Carcinogenic Risks to Humans: Some Naturally Occurring Substances: Food Items and Constituents, Heterocyclic Aromatic Amines and Mycotoxins; IARC: Lyon, France, 2012; Volume 56, pp. 245–395. [Google Scholar]
- Rustom, I.Y. Aflatoxin in food and feed: Occurrence, legislation and inactivation by physical methods. Food Chem. 1997, 59, 57–67. [Google Scholar] [CrossRef]
- Filazi, A.; Sireli, U.T. Occurrence of aflatoxins in food. In Aflatoxins Recent Advances and Future Prospects; Razzaghi-Abyaneh, M., Ed.; InTech Open Access Publisher: Rijeka, Croatia, 2013; pp. 143–170. [Google Scholar]
- Bennett, J.W.; Klich, M. Mycotoxins. Clin. Microbiol. Rev. 2003, 16, 497–516. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- RASFF. The Rapid Alert System for Food and Feed Annual Report. Available online: https://ec.europa.eu/food/safety/rasff/reports_publications_en (accessed on 20 November 2019).
- Wu, F. Global impacts of aflatoxin in maize: Trade and human health. World Mycotoxin J. 2014, 8, 137–142. [Google Scholar] [CrossRef]
- Sabzevari, O.; Emrarian, I.; Abdi, K.; Abouhossain, G.; Amirahmadi, M. Detection of aflatoxins in human hair. Iran. J. Pharm. Sci. 2006, 2, 21. [Google Scholar]
- Adebo, O.A.; Njobeh, P.B.; Gbashi, S.; Nwinyi, O.C.; Mavumengwana, V. Review on microbial degradation of aflatoxins. Crit. Rev. Food Sci. Nutr. 2015, 57, 3208–3217. [Google Scholar] [CrossRef]
- Mishra, H.N.; Das, C. A review on biological control and metabolism of aflatoxin. Crit. Rev. Food Sci. Nutr. 2003, 43, 245–264. [Google Scholar] [CrossRef]
- Hormisch, D.; Brost, I.; Kohring, G.W.; Giffhorn, F.; Kroppenstedt, R.M.; Stackebradt, E.; Stackebrandt, E.; Holapfel, H. Mycobacterium fluoranthenivorans sp. nov., a fluoranthene and aflatoxin B1 degrading bacterium from contaminated soil of a former coal gas plant. Syst. Appl. Microbiol. 2004, 27, 653–660. [Google Scholar] [CrossRef]
- Petchkongkaew, A.; Taillandier, P.; Gasaluck, P.; Lebrihi, A. Isolation of Bacillus spp. from Thai fermented soybean (Thua-nao): Screening for aflatoxin B1 and ochratoxin A detoxification. J. Appl. Microbiol. 2008, 104, 1495–1502. [Google Scholar] [CrossRef] [Green Version]
- Tejada-Castañeda, Z.I.; Ávila-Gonzalez, E.; Casaubon-Huguenin, M.T.; Cervantes-Olivares, R.A.; Vásquez-Peláez, C.; Hernández-Baumgarten, E.M.; Moreno-Martínez, E. Biodetoxification of aflatoxin-contaminated chick feed. Poult. Sci. 2008, 87, 1569–1576. [Google Scholar] [CrossRef]
- Topcu, A.; Bulat, T.; Wishah, R.; Boyacõ, I.H. Detoxification of aflatoxin B1 and patulin by Enterococcus faecium strains. Int. J. Food Microbiol. 2010, 139, 202–205. [Google Scholar] [CrossRef] [PubMed]
- Gao, X.; Ma, Q.; Zhao, L.; Lei, Y.; Shan, Y.; Ji, C. Isolation of Bacillus subtilis: Screening for aflatoxins B1, M1, and G1 detoxification. Eur. Food Res. Technol. 2011, 232, 957. [Google Scholar] [CrossRef]
- Samuel, M.S.; Sivaramakrishna, A.; Mehta, A. Degradation and detoxification of aflatoxin B1 by Pseudomonas putida. Int. Biodeterior. Biodegrad. 2014, 86, 202–209. [Google Scholar] [CrossRef]
- Rao, K.R.; Vipin, A.V.; Hariprasad, P.; AnuAppaiah, K.A.; Venkateswaran, G. Biological detoxification of Aflatoxin B1 by Bacillus licheniformis CFR1. Food Control. 2017, 71, 234–241. [Google Scholar]
- Motomura, M.; Toyomasu, T.; Mizuno, K.; Shinozawa, T. Purification and characterization of an aflatoxin degradation enzyme from Pleurotus ostreatus. Microbiol. Res. 2003, 158, 237–242. [Google Scholar] [CrossRef]
- Branà, M.T.; Cimmarusti, M.T.; Haidukowski, M.; Logrieco, A.F.; Altomare, C. Bioremediation of aflatoxin B1-contaminated maize by king oyster mushroom (Pleurotus eryngii). PLoS ONE 2017, 12, e0182574. [Google Scholar]
- Loi, M.; Fanelli, F.; Liuzzi, V.C.; Logrieco, A.F.; Mulé, G. Mycotoxin biotransformation by native and commercial enzymes: Present and future perspectives. Toxins. 2017, 9, 111–142. [Google Scholar] [CrossRef]
- Arora, D.S.; Sharma, R.K. Ligninolytic fungal laccases and their biotechnological Applications. Appl. Biochem. Biotechnol. 2010, 160, 1760–1788. [Google Scholar] [CrossRef]
- Detroy, R.W.; Hesseltine, C.W. Aflatoxicol: Structure of a new transformation product of aflatoxin B1. Can. J. Biochem. 1970, 48, 830–832. [Google Scholar] [CrossRef]
- Yehia, R.S. Aflatoxin detoxification by manganese peroxidase purified from Pleurotus ostreatus. Braz. J. Microbiol. 2014, 45, 127–134. [Google Scholar] [CrossRef] [Green Version]
- Alberts, J.F.; Gelderblom, W.C.A.; Botha, A.; Van Zyl, W.H. Degradation of aflatoxin B1 by fungal laccase enzymes. Int. J. Food Microbiol. 2009, 135, 47–52. [Google Scholar] [CrossRef] [PubMed]
- Loi, M.; Fanelli, F.; Cimmarusti, M.T.; Mirabelli, V.; Haidukowski, M.; Logrieco, A.; Caliandro, R.; Mule, G. In vitro single and combined mycotoxins degradation by Ery4 laccase from Pleurotus eryngii and redox mediators. Food Control. 2018, 8, 245. [Google Scholar]
- Royse, D.J.; Baars, J.; Tan, Q. Current overview of mushroom production in the world. Edible Med. Mushrooms Technol. Appl. 2017, 5–13. [Google Scholar] [CrossRef]
- Stajić, M.; Vukojević, J.; Duletić-Lauševic, S. Biology of Pleurotus eryngii and role in biotechnological processes: A review. Crit. Rev. Biotechnol. 2009, 29, 55–66. [Google Scholar] [CrossRef]
- Gregori, A.; Švagelj, M.; Pohleven, J. Cultivation techniques and medicinal properties of Pleurotus spp. Food Technol. Biotechnol. 2007, 45, 238–249. [Google Scholar]
- Cohen, R.; Persky, L.; Hadar, Y. Biotechnological applications and potential of wood-degrading mushrooms of the genus Pleurotus. Appl. Microbiol. Biotechnol. 2002, 58, 582–594. [Google Scholar] [CrossRef]
- Estrada, A.E.R.; del Mar Jimenez-Gasco, M.; Royse, D.J. Improvement of yield of Pleurotus eryngii var. eryngii by substrate supplementation and use of a casing overlay. Bioresour. Technol. 2009, 100, 5270–5276. [Google Scholar] [CrossRef]
- Rinker, D.L. Handling and using of “Spent” Mushroom Substrate around the world. In Mushroom Biology and Mushroom Products; Sánchez, J.E., Huerta, G., Montiel, E., Eds.; UAEM: Cuernavaca, Mexico, 2002; pp. 43–60. [Google Scholar]
- Ball, A.S.; Jackson, A.M. The recovery of lignocelluloses-degrading enzymes from spent mushroom compost. Bioresour. Technol. 1995, 54, 311–314. [Google Scholar] [CrossRef]
- Ko, H.G.; Park, S.H.; Kim, S.H.; Park, H.G.; Park, W.M. Detection and recovery of hydrolytic enzymes from spent compost of four mushroom species. Folia Microbiol. 2005, 50, 103–106. [Google Scholar] [CrossRef]
- Karla, M.D.; Maria del Refugio, T.H.; Marco, R.P. Recovery of laccase from the residual compost of Agaricus bisporus in aqueous two-phase systems. Proc. Biochem. 2009, 44, 435–439. [Google Scholar]
- Li, X.; Lin, X.; Zhang, J.; Wu, Y.; Yin, R.; Feng, Y.; Wang, Y. Degradation of polycyclic aromatic hydrocarbons by crude extracts from spent mushroom substrate and its possible mechanisms. Curr. Microbiol. 2010, 60, 336–342. [Google Scholar] [CrossRef] [PubMed]
- Phan, C.-W.; Sabaratnam, V. Potential uses of spent mushroom substrate and its associated lignocellulosic enzymes. Appl. Microbiol. Biotechnol. 2012, 96, 863–873. [Google Scholar] [CrossRef] [PubMed]
- Martirani, L.; Giardina, P.; Marzullo, L.; Sannia, G. Reduction of phenol content and toxicity in olive oil mill waste waters with the ligninolytic fungus Pleurotus ostreatus. Water Res. 1996, 30, 1914–1918. [Google Scholar] [CrossRef]
- Lau, K.L.; Tsang, Y.Y.; Chiu, S.W. Use of spent mushroom compost to bioremediate PAH-contaminated samples. Chemosphere. 2003, 52, 1539–1546. [Google Scholar] [CrossRef]
- Mougin, C.; Boukcim, H.; Jolivalt, C. Soil bioremediation strategies based on the use of fungal enzymes. In Advances in Applied Bioremediation; Singh, A., Kuhad, R.C., Ward, O.P., Eds.; Springer: Berlin/Heidelberg, Germany, 2009; Volume 17, pp. 123–149. [Google Scholar]
- Loi, M.; Fanelli, F.; Zucca, P.; Liuzzi, V.C.; Quintieri, L.; Cimmarusti, M.T.; Mulè, G. Aflatoxin B1 and M1 degradation by Lac2 from Pleurotus pulmonarius and redox mediators. Toxins. 2016, 8, 245–261. [Google Scholar] [CrossRef] [Green Version]
- Scarpari, M.; Bello, C.; Pietricola, C.; Zaccaria, M.; Bertocchi, L.; Angelucci, A.; Ricciardi, M.R.; Scala, V.; Parroni, A.; Fabbri, A.A.; et al. Aflatoxin control in maize by Trametes versicolor. Toxins. 2014, 6, 3426–3437. [Google Scholar] [CrossRef] [Green Version]
- Hajare, S.S.; Hajare, S.N.; Sharma, A. Aflatoxin inactivation using aqueous extract of ajowan (Trachyspermum ammi) seeds. J. Food Sci. 2005, 70, C29–C34. [Google Scholar] [CrossRef]
- Wang, J.; Ogata, M.; Hirai, H.; Kawagishi, H. Detoxification of aflatoxin B1 by manganese peroxidase from the white-rot fungus Phanerochaete sordida YK-624. FEMS Microbiol. Lett. 2011, 314, 164–169. [Google Scholar] [CrossRef]
- Adebo, O.A.; Njobeh, P.B.; Mavumengwana, V. Degradation and detoxification of AFB1 by Staphylocococcus warneri, Sporosarcina sp. and Lysinibacillus fusiformis. Food Control. 2016, 68, 92–96. [Google Scholar] [CrossRef]
- Méndez-Albores, A.; Del Río-García, J.C.; Moreno-Martínez, E. Decontamination of aflatoxin duckling feed with aqueous citric acid treatment. Anim. Feed Sci. Technol. 2007, 135, 249–262. [Google Scholar] [CrossRef]
- Méndez-Albores, A.; Veles-Medina, J.; Urbina-Álvarez, E.; Martínez-Bustos, F.; Moreno-Martínez, E. Effect of citric acid on aflatoxin degradation and on functional and textural properties of extruded sorghum. Anim. Feed Sci. Technol. 2009, 150, 316–329. [Google Scholar] [CrossRef]
- Alberts, J.; Schatzmayr, G.; Moll, W.-D.; Davids, I.; Rheeder, J.; Burger, H.-M.; Shephard, G.; Gelderblom, W. Detoxification of the fumonisin mycotoxins in maize: An enzymatic approach. Toxins. 2019, 11, 523. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van der Westhuizen, L.; Shephard, G.S.; Burger, H.M.; Rheeder, J.P.; Gelderblom, W.C.; Wild, C.P.; Gong, Y.Y. Fumonisin B1 as a urinary biomarker of exposure in a maize intervention study among South African subsistence farmers. Cancer Epidemiol. Biomark. Prev. 2011, 20, 483–489. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boer, C.G.; Obici, L.; de Souza, C.G.M.; Peralta, R.M. Decolorization of synthetic dyes by solid state cultures of Lentinula (Lentinus) edodes producing manganese peroxidase as the main ligninolytic enzyme. Bioresour. Technol. 2004, 94, 107–112. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantization of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–253. [Google Scholar] [CrossRef]
- AOAC. Section 49.2.02 (AOAC method 971.22) preparation of standards. In Official Methods of Analysis, 17th ed.; AOAC International: Gaithersburg, MD, USA, 2000. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Control AFB1 (ng mL−1) | SMSE | |
|---|---|---|
| AFB1 (ng mL−1) (a) | D (%) (b) | |
| 154 ± 3 | 19 ± 4 * | 88 a |
| 322 ± 8 | 36 ± 2 * | 89 a |
| 462 ± 11 | 61 ± 2 * | 87 a |
| 885 ± 22 | 146 ± 2 * | 84 a |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Branà, M.T.; Sergio, L.; Haidukowski, M.; Logrieco, A.F.; Altomare, C. Degradation of Aflatoxin B1 by a Sustainable Enzymatic Extract from Spent Mushroom Substrate of Pleurotus eryngii. Toxins 2020, 12, 49. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins12010049
Branà MT, Sergio L, Haidukowski M, Logrieco AF, Altomare C. Degradation of Aflatoxin B1 by a Sustainable Enzymatic Extract from Spent Mushroom Substrate of Pleurotus eryngii. Toxins. 2020; 12(1):49. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins12010049
Chicago/Turabian StyleBranà, Maria Teresa, Lucrezia Sergio, Miriam Haidukowski, Antonio F. Logrieco, and Claudio Altomare. 2020. "Degradation of Aflatoxin B1 by a Sustainable Enzymatic Extract from Spent Mushroom Substrate of Pleurotus eryngii" Toxins 12, no. 1: 49. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins12010049