1. Introduction
Deoxynivalenol (DON) is a mycotoxin produced by the Fusarium species, commonly contaminating cereal-based foods, such as wheat, rye, barley, oats, and corn [
1,
2]. DON is resistant to milling, processing, and heating [
3] and, therefore, readily enters the food chain, causing food poisoning. Several outbreaks of acute human gastrointestinal disorders, including nausea, vomiting, diarrhea, and gastrointestinal discomfort, have been reported in Asia [
4,
5].
The gastrointestinal tract is the first physiologic barrier against food contaminants, as well as the first target for toxicants. DON reportedly affects the small intestine, and, most notably, its negative effects on intestinal barrier functions were demonstrated in a previous work [
6]. Intestinal barrier functions are primarily formed by intestinal epithelial cells connected by tight junction proteins, forming an anastomosing network that seals adjacent epithelial cells near the luminal surface [
7,
8]. Tight junctions are composed of (1) transmembrane proteins, including occludins and claudins, which form a linear barrier at the luminal lateral cell membranes, and (2) peripheral membrane proteins, such as zonula occludens (ZO) proteins, which link transmembrane tight junction proteins together [
9]. DON also reportedly affects tight junction proteins by decreasing claudin proteins after DON exposure in vitro [
10].
Interestingly, DON is rapidly absorbed while passing through the stomach and the proximal small intestine [
11]. This report suggests that DON affects the intestinal cells not only from the luminal side but also from the basolateral side through the blood stream. A few scientists previously explored the different effects of DON from both the luminal and basolateral routes using porcine intestinal cell lines [
12,
13,
14] or human epithelial cell lines [
15]. They reported that basolateral DON exposure has greater impacts on intestinal barrier functions, cell proliferation, and interleukin-8 secretion than its luminal exposure. However, the intestinal cell lines used in previous studies were composed of enterocyte monolayers without intestinal crypt-villus structures and native intestinal proliferation and differentiation functions. Additionally, these cell lines lacked differentiated cells, such as goblet cells, enteroendocrine cells, Paneth cells, and tuft cells, because intestinal stem cells were absent.
Recently, intestinal organoids (enteroids) are widely used as in vitro models that mimic most features of intestinal epithelium in vivo. They consist of intestinal stem cells, as well as most types of differentiated cells derived from stem cells, and are easily generated from isolated crypts as previously shown [
16,
17]. Establishing enteroid cultures was a breakthrough method in evaluating the effects of substances on intestinal stem cells in vitro. Until recently, little information was available on the DON toxicity on intestinal stem cells. Only a few enteroid-based studies showed that adding DON to the enteroid culture medium suppressed intestinal cell proliferation and decreased the protein level of leucine-rich repeat-containing G-protein-coupled receptor 5 (Lgr5), a stem cell marker in the small intestine [
18,
19]. However, their studies only reflected the effects of basolateral DON exposure on enteroids due to their reversed basolateral-out structure, in contrast to the native intestine. Therefore, the effects of luminal DON exposure on intestinal stem cells remain unclear.
Our goal was to investigate the impacts of luminal or basolateral DON exposure on the small intestine using enteroids. In the present study, we focused on the harmful effects of DON on intestinal barrier functions, as well as intestinal stem cells, after luminally or basolaterally exposed to DON.
3. Discussion
Several studies have reported that DON disrupts normal intestinal barrier functions [
6,
10,
24,
25,
26,
27]. However, only a few studies reported the effects of luminal or basolateral DON exposure on the intestine using intestinal cell lines [
12,
14]. Thus, we first addressed if luminal or basolateral DON exposure affects intestinal barrier functions using our enteroid model, which mimics most features of the native intestine. Recently, Yokoi et al. [
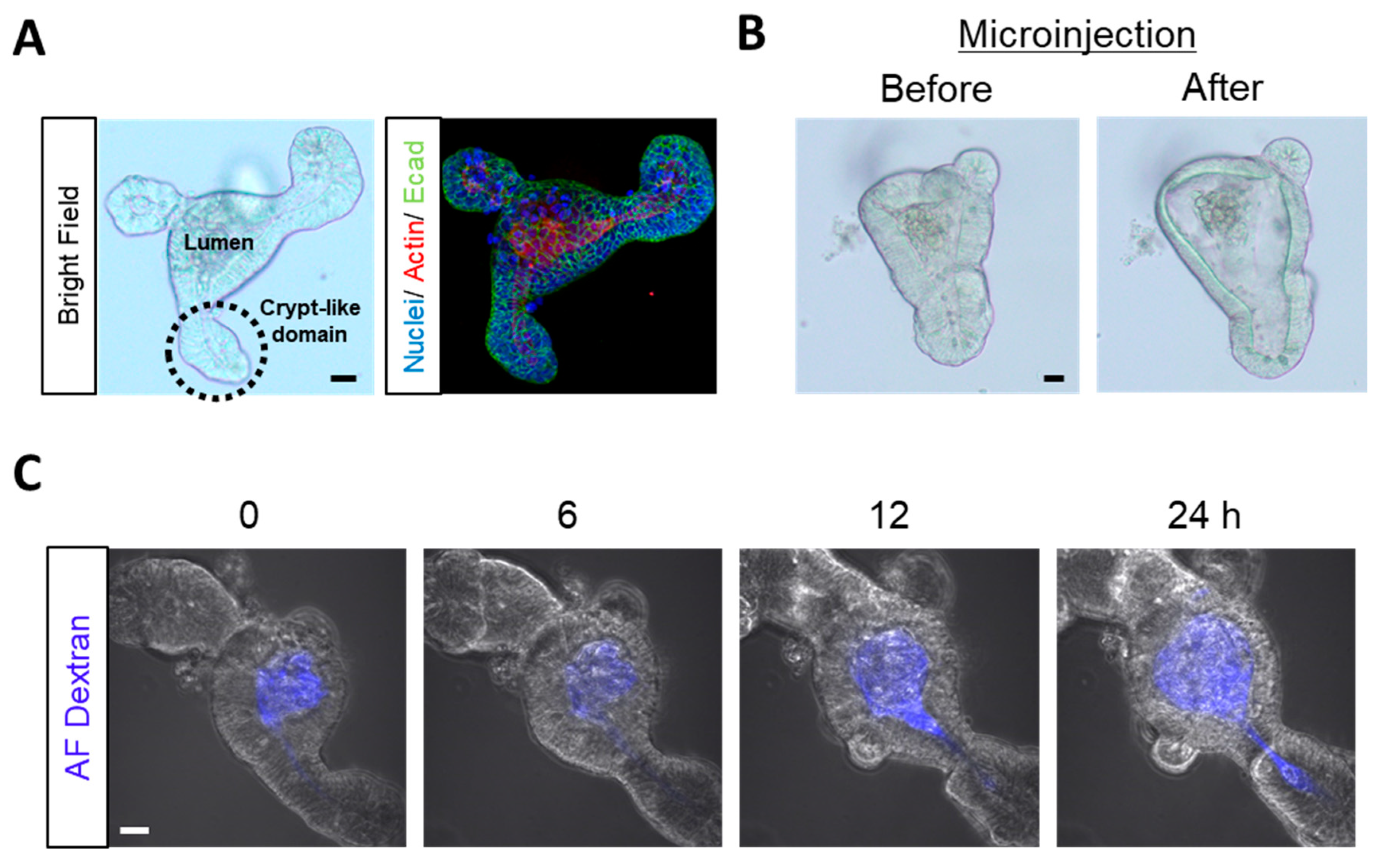
20] devised a novel microinjection method that quantitatively luminally delivers substances to enteroids. This method allowed us to compare between the effects of luminal and basolateral DON treatment at equal concentrations (
Figure 1). Our experiment using fluorescent dextran (
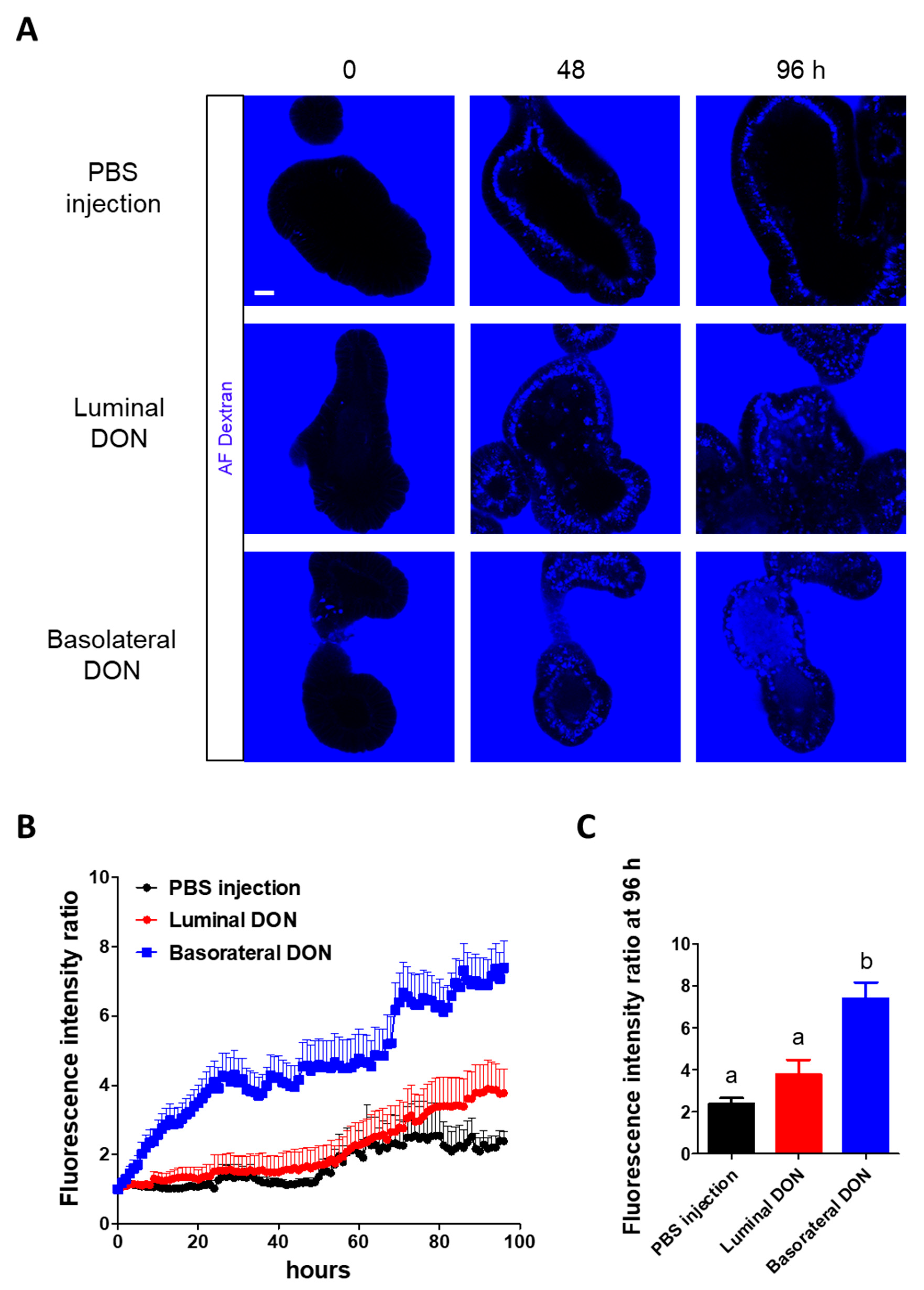
Figure 2) demonstrated that basolateral DON exposure had more impact on intestinal barrier functions than luminal DON exposure, agreeing with previous reports using intestinal cell lines [
12,
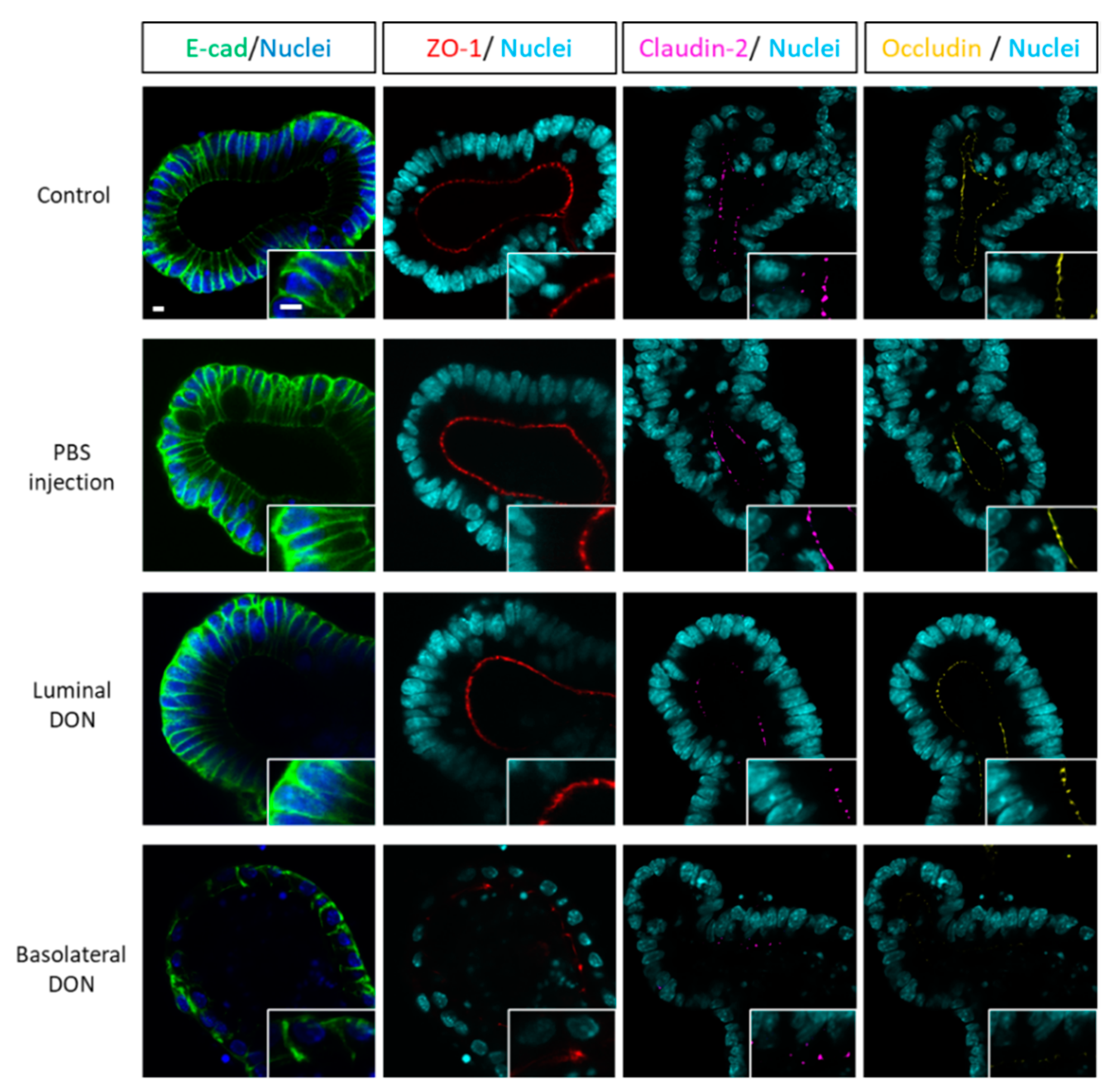
14]. We hypothesized that basolateral DON exposure may disrupt the intestinal barrier by affecting the intestinal epithelial structure, since the intestinal barrier is assisted by the intestinal epithelial proteins involved in the adherens junction or tight junction. Immunofluorescence of intestinal epithelial proteins (E-cadherin, claudin, and occludin) in enteroids (
Figure 3) revealed that only basolateral DON exposure broke down intestinal epithelial integrity. Taken together, basolateral DON exposure had more negative effects on intestinal epithelial proteins that have important roles in intestinal barrier functions, compared with luminal DON exposure. Moreover, our enteroid model enabled us to visually assess intestinal barrier functions, which has more similarity to native intestinal epithelium than 2-D cell line models used in previous studies.
The molecular mechanism by which basolateral DON exposure affected intestinal barrier functions is reportedly due to the occurrence where DON decreases tight junction proteins by activating the MAPK pathway [
24,
26]. In the present study, the basolateral DON exposure did not significantly change extracellular signal-regulated kinase (ERK), p38 mitogen activated protein kinase (p38), or c-jun N-terminal kinase (JNK) gene expression in enteroids in comparison with the control (
Figure S4). Based on our RNA-seq results, MAPK signaling activation possibly caused the disruption of intestinal barrier functions in enteroids basolaterally exposed to DON in the present study, which is similar to previous report [
24,
26].
Although we demonstrated that the basolateral DON has lager impacts on intestinal barrier function in our enteroid model, the same as previous reports, our interest was how luminal or basolateral DON affects intestinal stem cells. The effects of DON exposure on intestinal stem cells have not been tested in vitro due to the lack of a model that contains intestinal stem cells, although the shortened villi observed in DON-administrated mice implied that it affects intestinal cell proliferation and/or intestinal stem cells [
6,
28,
29,
30]. Subsequently, establishing a protocol to generate enteroids was the breakthrough in studying intestinal stem cells in vitro [
17]. Using this model, Li et al. [
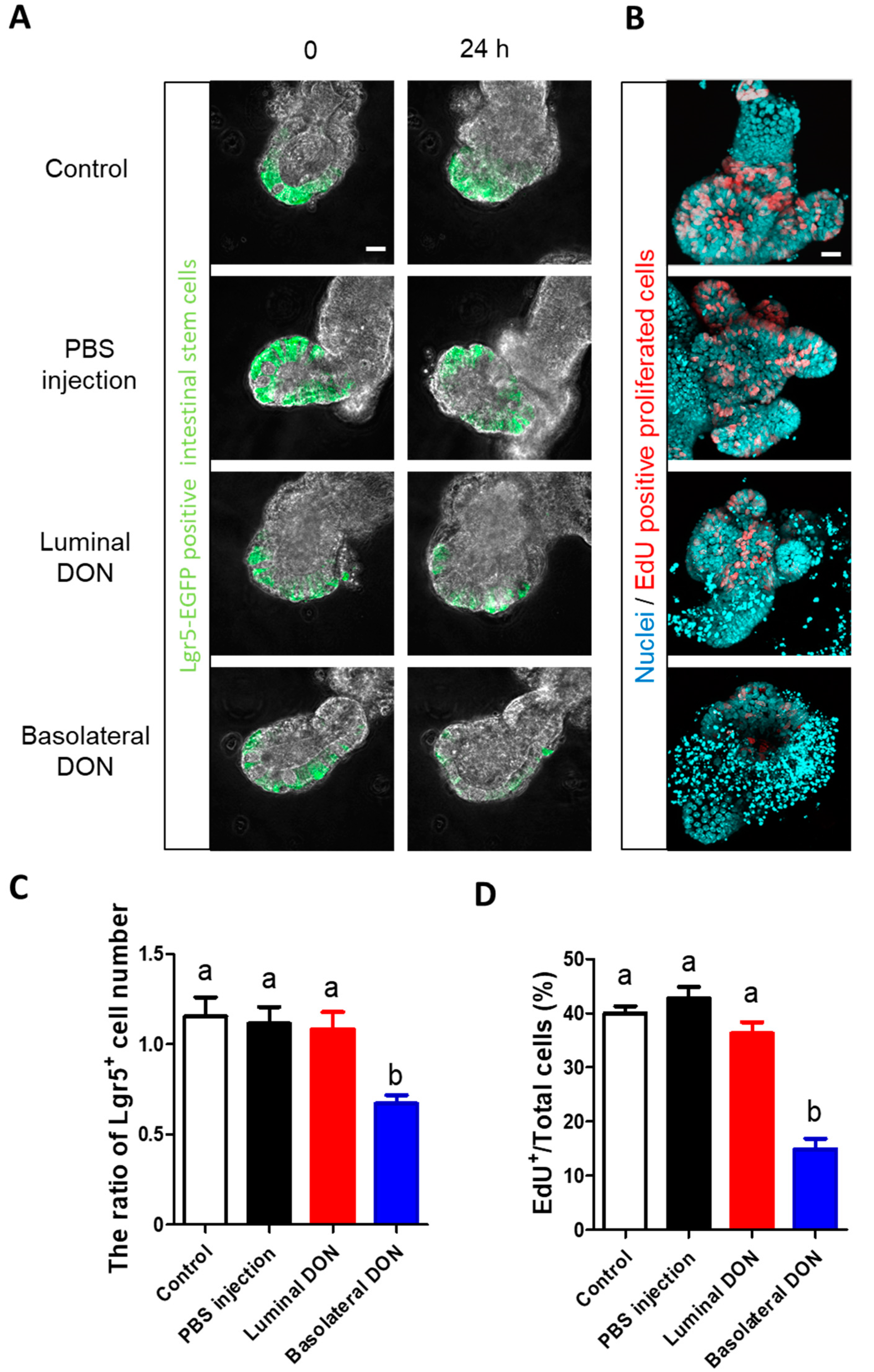
22] demonstrated that basolateral DON exposure (adding DON to a porcine enteroid culture medium) inhibited intestinal stem cell activity through the Wnt/β-catenin pathway. Thus, we further explored the effects of luminal or basolateral DON exposure on intestinal stem cells. Our outcomes (
Figure 4) demonstrated that basolateral DON exposure greatly affected intestinal stem cells than did luminal DON exposure at equal concentrations. Next, we examined the effects of DON on stem cells in native small intestine of mice with oral administration (50 mg/kg BW DON) (
Figure 5). Our purpose of conducting the in vivo experiment was to know whether DON has similar toxic effects in vivo to those observed on stem cells of enteroids. Thus, we used 50 mg/kg BW of DON as the dose, based on our preliminary experiments. Although we did not measure DON concentration in the blood, Pestka et al. [
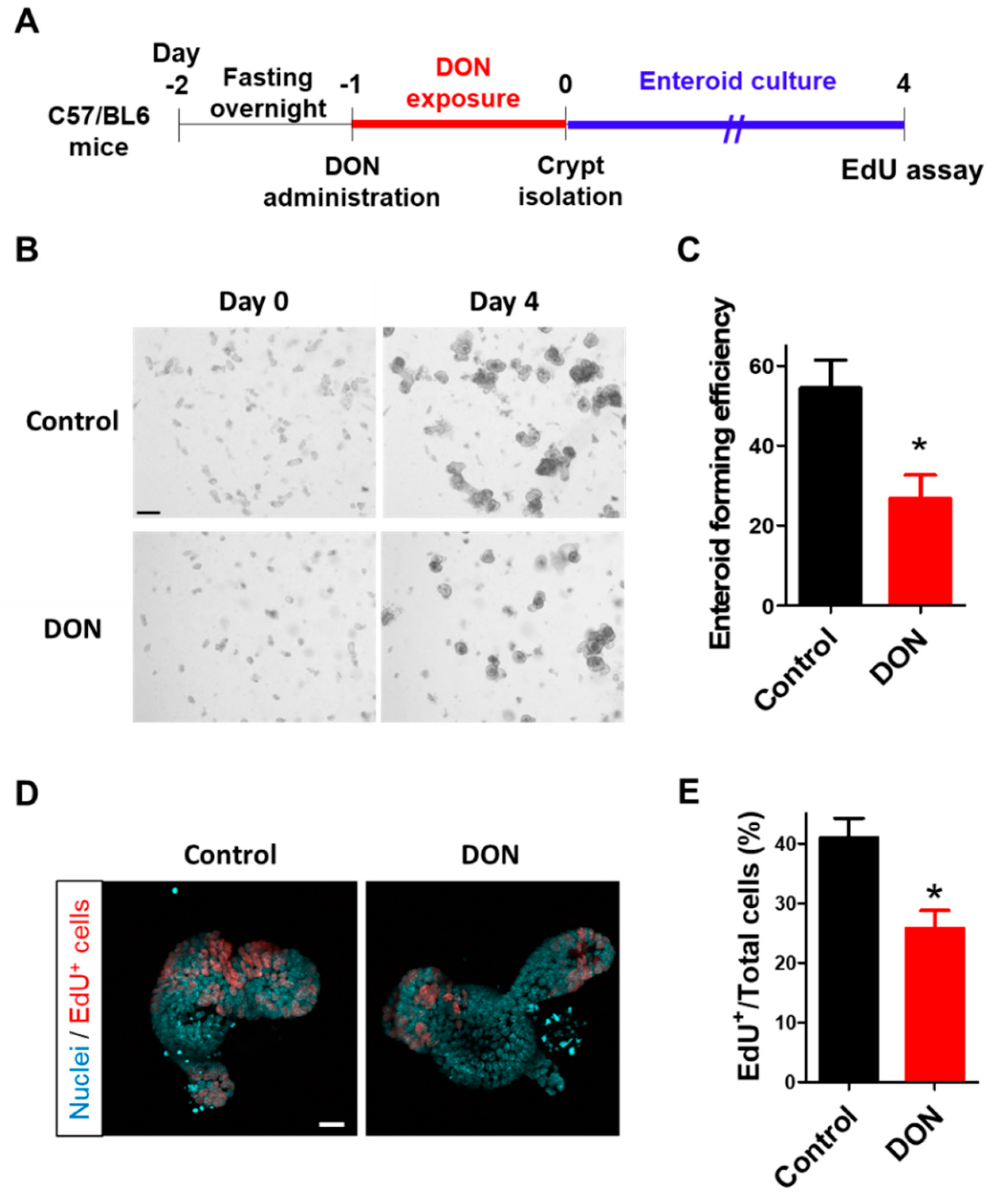
31] showed that DON level in the blood was about 0.4 μg/mL at 24 h after a single oral exposure of 25 mg/kg BW of DON. In the case of 50 mg/kg BW of DON, DON concentration in the blood is expected to be about 0.8 μg/mL (2.7 μM), which is almost comparable to the concentration that was used in our in vitro experiment (1 μM DON). Enteroids derived from mice orally administrated with DON showed reduced forming efficiency and proliferative activity in comparison with the control enteroids (
Figure 5), which is consistent with our in vitro results, showing that the in vitro enteroid model can be a powerful tool to test the effects of toxins on intestinal stem cells.
We assumed that DON has greater impacts on intestinal stem cells than on differentiated cells, although the present study did not investigate DON’s effects on differentiated cells. Proliferating cells, such as intestinal stem cells, are frequently expected to be highly susceptible to DON, since DON has an ability to inhibit protein synthesis [
32,
33]. Indeed, the DON concentration (1 µM) used in our enteroid experiment, which negatively affected intestinal stem cells, was lower than that in previous works using cell lines [
32,
33]. The RNA-seq analysis in our study revealed that only the basolateral DON exposure downregulated the gene expression of cyclin D, a target gene of the Wnt signaling pathway (
Figure S4B). This is supported by a previous study that reported DON to have an effect on stem cells through the Wnt/β-catenin pathway [
22]. However, based on our RNA-seq results, the gene expressions of Wnt or frizzled-class receptors were upregulated only after basolateral DON exposure (
Figure S4B). This may be the negative feedback regulation to compensate for the suppression of Wnt/β-catenin pathway caused by DON exposure. Taken together, the high susceptibility to DON and the downregulation of the Wnt/β-catenin pathway may be attributed to the decreased stem cell number.
The question of why basolateral DON exposure has more impacts on the intestine than luminal DON exposure at the same concentration still remains. There are three possible answers: Firstly, basolateral DON could enter the cells potentially faster or more easily, compared with apical entry, because P-glycoprotein (P-gp) in the apical membrane exports drugs out of the cells, as previously discussed [
14]. Certainly, Ivanova et al. have reported that DON is exported by P-gp, causing it to be less cytotoxic in P-gp-overexpressing cells [
34]. Secondly, our RNA-seq analysis showed that several gene expressions, including extracellular matrix (ECM) in the focal adhesion pathway (mmu04510), were upregulated not by luminal DON exposure, but by basolateral exposure (
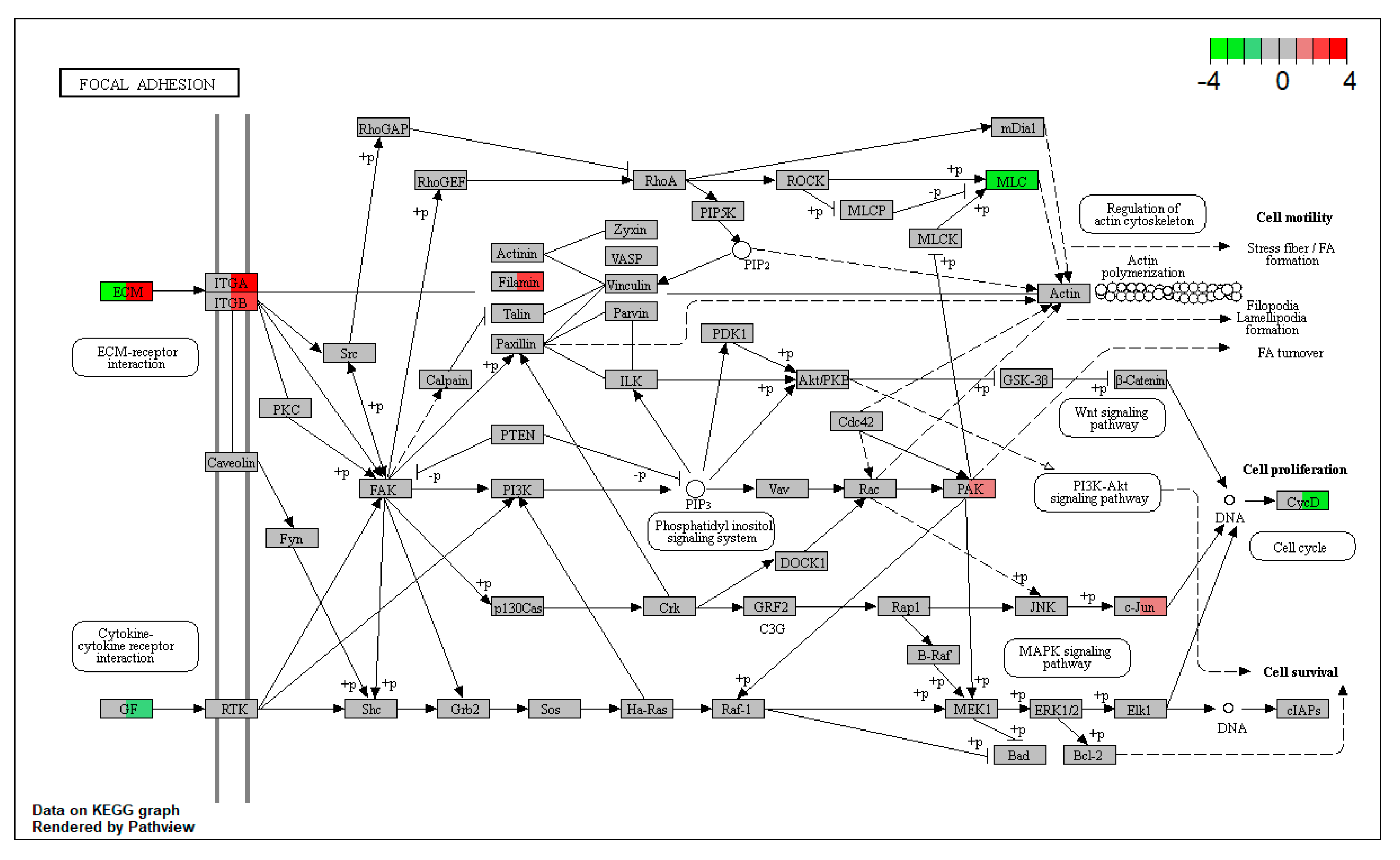
Figure 7). This result implies that basolateral DON exposure caused extracellular damage to the focal adhesion pathway, subsequently inducing feedback regulation. Gene expression changes in the focal adhesion pathway suggest that the basolateral DON is likely to enter the intestinal epithelium, leading to greater impacts than luminal DON. Finally, the luminal mucus layer of enteroids may inhibit DON entry from the luminal side, thereby lowering the toxic effects of luminal DON exposure. Enteroids stained with periodic acid-Schiff showed that there were goblet cells and secreted mucus substances that covered the enteroid lumen surface (
Figure S5) (see
Supplementary Materials and Methods: 4). However, in the present study, the detailed, involved mechanisms still remain unclear. Further studies on these mechanisms are required.
In conclusion, our study revealed that basolateral DON exposure was more toxic than luminal DON exposure in terms of intestinal barrier functions and stem cells. In particular, the present study is the first to show that the basolateral DON exposure had greater negative effects on intestinal stem cells than luminal DON exposure. Individually clarifying the effects of DON on the intestine both from the luminal and basolateral sides is important, since DON is rapidly and nearly completely absorbed in the stomach and the proximal small intestine [
11]. Our findings provide useful information in accurately understanding DON toxicity on intestinal barrier functions and stem cells. Additionally, our enteroid model is animal-friendly and may be technically useful for evaluating the effects of toxins on intestinal stem cells and intestinal epithelium functions, because this enteroid-based in vitro model reduces the experimental period, cost, and number of animals used, compared with in vivo models.
4. Materials and Methods
4.1. Experimental Animals
C57BL/6J (wild-type; WT) mice (CLEA Japan, Inc., Tokyo, Japan) and Lgr5-EGFP-internal ribosome entry site (IRES)-CreERT2 (Lgr5-EGFP) mice (C57BL/6 background; The Jackson Laboratory, Bar Harbor, ME, USA) were fed with standard laboratory rodent food (MF; Oriental Yeast Co., Ltd., Tokyo, Japan), with free access to water. Male and female mice were used for experimentation at 2–4 months of age. The Lgr5-EGFP mice were used to visualize intestinal stem cells in enteroids. For in vitro experiments, we prepared enteroids from three animals (approximately half male and female mice) in each experiment. For oral DON administration, the WT mice were fasted overnight with free access to water and then orally treated with DON at a dose of 50 mg/kg body weight (two male mice for the control and three male mice for the DON-treated group). Mice were sacrificed for crypt isolation at 24 h after oral DON administration. All animal experiments adhered to the guidelines for maintaining and handling of experimental animals established by the Tokyo University of Agriculture Ethics Committee. Approval code and date of the ethics committee: 2019040 (27 September 2019) and 290015 (28 September 2017)
4.2. Generation of Mouse Enteroids
Enteroids from the mouse jejunum were generated as previously described [
16,
17]. Briefly, the isolated mouse jejunum was incubated with rocking at 4 °C for 30 min in 2.5 mM ethylenediaminetetraacetic acid (EDTA) in Dulbecco’s phosphate-buffered saline (DPBS; Nacalai Tesque, Inc., Kyoto, Japan) without calcium or magnesium. The tissue was then placed in an ice-cold dissociation buffer (43.3 mM sucrose (FUJIFILM Wako Pure Chemical Corporation, Osaka, Japan) and 54.9 mM D-sorbitol (Nacalai Tesque, Inc., Kyoto, Japan) in DPBS) and was forcefully shaken by hand to dissociate individual crypts. The dissociated crypts were centrifuged at 400×
g for 4 min at 4 °C, and the pellet was resuspended in 60% Matrigel (Corning Incorporated, Corning, NY, USA) in ice-cold DPBS. The crypts suspended in Matrigel were added to a 24-well cell culture plate (NIPPON Genetics Co., Ltd., Tokyo, Japan). After Matrigel polymerization at 37 °C, enteroid culture medium (advanced Dulbecco’s modified Eagle medium/Ham’s F-12 (DMEM/F-12; Thermo Fisher Scientific Inc., Waltham, MA, USA) supplemented with 2 mM L-glutamine (Nacalai Tesque, Inc., Kyoto, Japan), 10 mM 2-[4-(2-hydroxyethyl)piperazin-1-yl]ethanesulfonic acid (HEPES; Nacalai Tesque, Inc., Kyoto, Japan), 1× penicillin–streptomycin (Nacalai Tesque, Inc., Kyoto, Japan), 1× B27 supplements (Thermo Fisher Scientific Inc., Waltham, MA, USA), 1 mM N-acetyl-L-cysteine (Sigma–Aldrich Co., LLC, St. Louis, MO, USA), 10% R-spondin conditioned medium, 5% Noggin conditioned medium, and 50 ng/mL epidermal growth factor (EGF; PeproTech Inc., Rocky Hill, NJ, USA)) was added to the wells. Enteroids were cultured in a 5% CO
2 incubator at 37 °C until day 3. The enteroid culture medium was replaced every two days. For the experiment, enteroids were collected and resuspended in ice-cold 100% Matrigel and transferred into a 35-mm cell imaging dish (Eppendorf AG, Hamburg, Germany). After Matrigel polymerization at 37 °C, the enteroid culture medium was added into each dish. The enteroids were prepared from the control mice or mice orally treated with DON using the aforementioned protocol for assessing the enteroids from orally DON-treated mice. In this experiment, we used two C57/BL6 male mice for the control and three male mice for the DON-treated group. Enteroid-forming efficiency was calculated as the ratio of the number of enteroids at day 4 to the number of crypts seeded at day 0. To calculate enteroid-forming efficiency, crypts in four wells per one mouse, that is, 1.5–6.3 × 10
2 crypts/well (eight wells for the control and 12 wells for the DON-treated group;
n = 8–12) were counted at day 0. After four days, grown enteroids were counted in the same wells of 12-well cell culture plate.
4.3. DON Exposure of Enteroids
For basolateral DON exposure, DON (final conc. 1 µM; Sigma–Aldrich Co. LLC, St. Louis, MO, USA) was added to the enteroid culture medium. In contrast, luminal DON exposure was conducted through DON microinjection (28.3 µM in DPBS) into enteroids. Enteroid lumen microinjection and volume measurement of enteroid lumen and microinjection were done as previously described [
20]. The final concentration was equivalent to 1 µM in microinjected enteroids (
Table S1). Briefly, the tip of the needle (Femtotips II; Eppendorf AG, Hamburg, Germany) was broken off and its inner diameter was adjusted to 3–4 µm. The 28.3-µM DON solution (final conc. 1 µM) was loaded into the needle using a microloader tip (Eppendorf AG, Hamburg, Germany). Microinjection was performed using a microinjector (FemtoJet
® 4i; Eppendorf AG) and manipulators (MM-92 and MMO-202ND; Narishige Scientific Instrument Lab, Tokyo, Japan) under a microscope (Olympus Co., Tokyo, Japan). Enteroid lumen was stained with 1 µM rhodamine123 (Rh123) for 3 h (
Figure S1B,C) to measure its volume. Likewise, DPBS was microinjected into the enteroids to test the effects of microinjection itself (described as “PBS injection” in the figures). For the experiment to test dextran permeability, DPBS was microinjected into the enteroids in the basolateral DON exposure group before exposure in view of the effects of microinjection itself.
4.4. Long-Term Live Imaging of Enteroids
Live imaging was performed on the enteroid culture medium at 37 °C under 5% CO2 (incubation chamber conditions) on a confocal microscope (Fv10i; Olympus Co., Tokyo, Japan) and imaged with a 40× objective lens. To assess intestinal barrier functions, 10 μM Alexa Fluor® 647–10K Dextran (excitation: 633 nm, emission: 650–700 nm; Thermo Fisher Scientific Inc., Waltham, MA, USA) was added to the enteroid culture medium for 96 h. To test the effects of DON on intestinal stem cells, the fluorescence of Lgr5-EGFP in the enteroids was monitored for 24 h (excitation: 488 nm, emission: 500–550 nm). Five to seven enteroids in each group were imaged for each experiment using one mouse.
4.5. Whole-Mount Immunostaining
Enteroids with or without DON exposure for 72 h were fixed with 4% cold paraformaldehyde (Nacalai Tesque, Inc., Kyoto, Japan) for 30 min at room temperature. The enteroids were then blocked with 5% goat serum (Jackson ImmunoResearch Laboratories, Inc, West Grove, PA, USA) for 1 h at room temperature before incubating with ZO-1 (1:100; Cat #33-9100; Thermo Fisher Scientific Inc., Waltham, MA, USA), E-cadherin (1:400; Cat #AF748-SP; R&D systems, Inc., Minneapolis, MN, USA), claudin-2 (1:250; Cat #351-6100; Thermo Fisher Scientific Inc., Waltham, MA, USA), or occludin (1:250; Cat #33-1500; Thermo Fisher Scientific Inc., Waltham, MA, USA) antibodies overnight at 4 °C. The enteroids were incubated with Alexa Flour® 488, 647, or 555 (1:1000; Thermo Fisher Scientific Inc., Waltham, MA, USA) secondary antibodies overnight at 4 °C after washing all day with 0.1% Triton-X in PBS. Additionally, the enteroids were incubated with 10 μg/mL Hoechst 33342 (Thermo Fisher Scientific Inc., Waltham, MA, USA) for 20 min. Finally, the enteroids (five to seven enteroids in each group per one mouse) were imaged using a confocal microscope (Fv10i; Olympus Co., Tokyo, Japan).
4.6. EdU Assay
Enteroids with or without the DON exposure for 24 h were incubated with 10 μM EdU (Thermo Fisher Scientific Inc., Waltham, MA, USA) for 3 h. Then, 10 μg/mL Hoechst 33342 (Thermo Fisher Scientific Inc., Waltham, MA, USA) was added to the enteroid culture medium for 30 min prior to fixation with 4% cold paraformaldehyde (Nacalai Tesque, Inc., Kyoto, Japan) for 30 min at room temperature. The enteroids were stained using a Click-iT™ EdU Cell Proliferation Kit for Imaging (Cat# C10339; Thermo Fisher Scientific Inc., Waltham, MA, USA), according to the manufacturer’s protocol. Briefly, the enteroids were permeabilized with 0.5% Triton X-100 in PBS for 20 min and incubated with Click-iT detection cocktail for 30 min to detect EdU+ cells. The enteroids (5–7 enteroids in each group per one mouse) were imaged using a confocal microscope (Fv10i; Olympus Co., Tokyo, Japan). EdU assay was conducted, similarly to the aforementioned protocol, to investigate enteroids from mice orally treated with DON.
4.7. Image Analysis
For the dextran permeability experiment, Alexa Fluor
® 647-10K Dextran intensity in enteroid lumen was measured every hour until 96 h. For quantification, data were collected from 17 enteroids and 18 organoids in the PBS injection or the luminal DON exposure groups and in the basolateral DON exposure group, respectively, in total from three different mice (
Figure 2B,C;
n = 17–18). The fluorescence intensity ratio was calculated as follows: The fluorescence intensity in the enteroid lumen at various timepoints was normalized with that at 0 h (set to 1), and the values were relatively described as the fluorescence intensity ratio.
For the live imaging of Lgr5-EGFP enteroids, Lgr5
+ cells in the enteroids were counted every hour until 24 h. Data were collected from eight enteroids in the control group, 12 enteroids in the basolateral DON exposure group, 13 enteroids in the PBS injection group, and 16 enteroids in the luminal DON exposure group in total from three different mice
(Figure 4C;
n = 8–12). The Lgr5
+ stem cell number was normalized to the number of Lgr5
+ cell at 0 h after each treatment, which was relatively expressed as the Lgr5
+ cell number ratio.
For the EdU proliferation assay, the percentage of the EdU
+ cell number to total cell number was calculated and represented as EdU/total cells (%). Data were collected from 16 enteroids in the control group, 19 enteroids in the PBS injection group, 20 enteroids in the luminal DON exposure group, and 22 enteroids in the basolateral DON exposure group in total from three different mice (
Figure 4D;
n = 16–22). Similarly, data were collected from 12 enteroids in control group and 18 enteroids in DON-treated group in total from 2–3 different mouse (
Figure 5E;
n = 12–18).
4.8. RNA-Seq Analysis
The total RNA was isolated from the control or DON-exposed enteroids using ISOGEN II (Nippon Gene Co., Ltd., Tokyo, Japan). The isolated total RNA was treated with DNase to eliminate DNA contamination. The RNA library was prepared by TruSeq
® Stranded mRNA Library Prep. Kit (Illumina, Inc., San Diego, CA, USA), according to the manufacturer’s protocol. The cDNA was amplified, and the obtained cDNA pool was subjected to high-throughput sequencing using the NovaSeq 6000 system. Raw read data quality was checked using the FastQC v0.11.7 software (
http://www.bioinformatics.babraham.ac.uk/projects/fastqc/). Based on the scores, each read was cleaned through trimming adapter sequence and filtering low-quality sequence using the Trimmomatic 0.38 software (
http://www.usadellab.org/cms/?page=trimmomatic). The cleaned read data were aligned using the HISAT2 software (
https://daehwankimlab.github.io/hisat2/) version 2.1.0. Gene expression value was quantified using StringTie version 1.3.4d (
https://0-ccb-jhu-edu.brum.beds.ac.uk/software/stringtie/). The statistical test for comparing between two groups was performed using the R package, edgeR (
https://bioconductor.org/packages/release/bioc/html/edgeR.html). Significant DEGs were identified based on the conditions of |fold change| > = 2 and raw
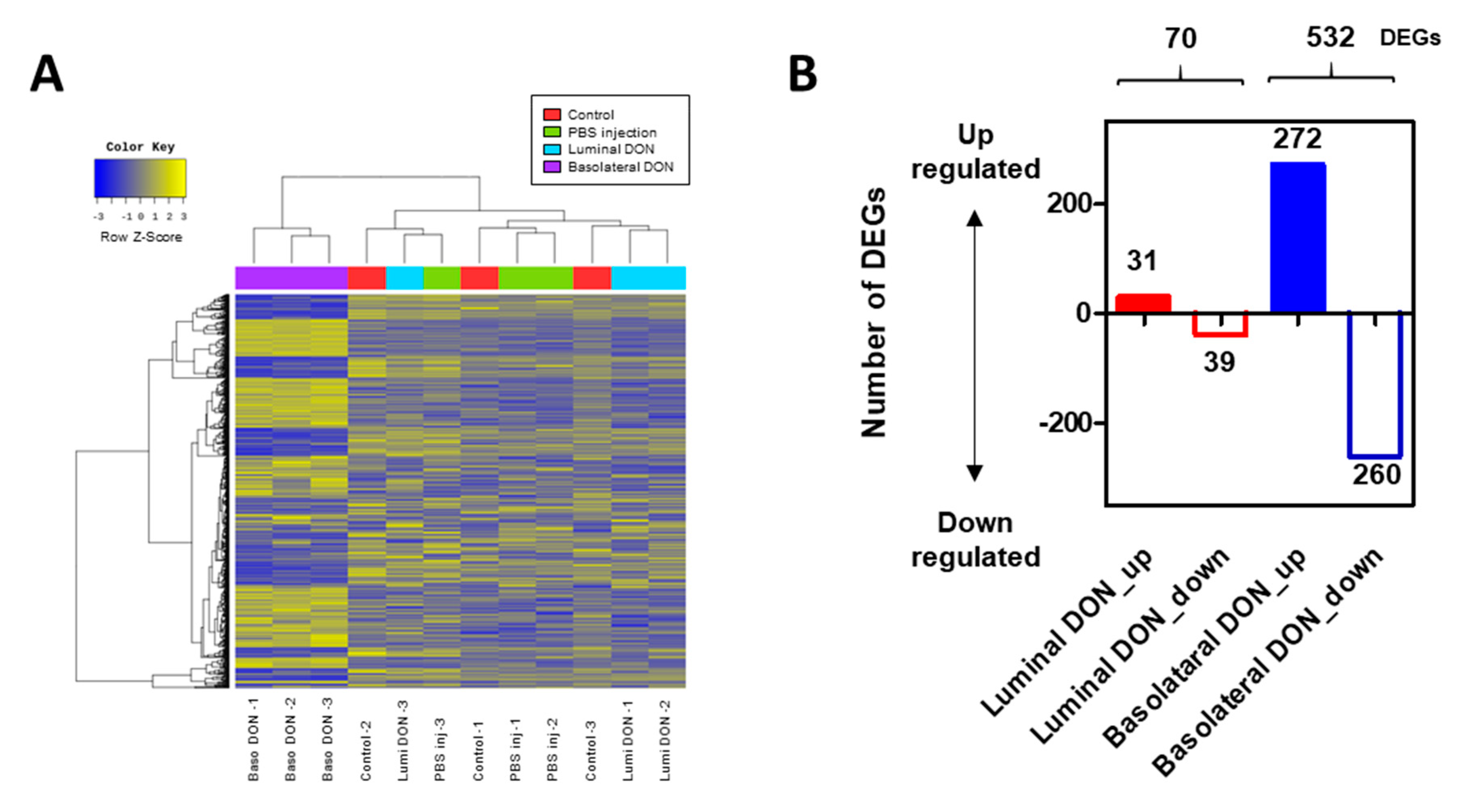
p-value < 0.05 in at least one comparison group. A functional annotation and gene ontology enrichment analysis were performed based on 539 DEGs between enteroid groups after luminal versus basolateral DON exposures using DAVID Bioinformatics Resources (
https://david.ncifcrf.gov/).
4.9. Statistical Analysis
All values were reported from representative experiments as the mean ± standard error of the mean (SEM) from multiple experiments using each enteroid derived from at least three animals. We determined the statistical significance using unpaired Student’s t-test, or one-way ANOVA with Tukey′s multiple comparison post hoc test. A p-value <0.05 was considered significant.

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}






