2.7. Discussion
The first strategy of application assumed that the EXT added to the FN may increase the lethality of
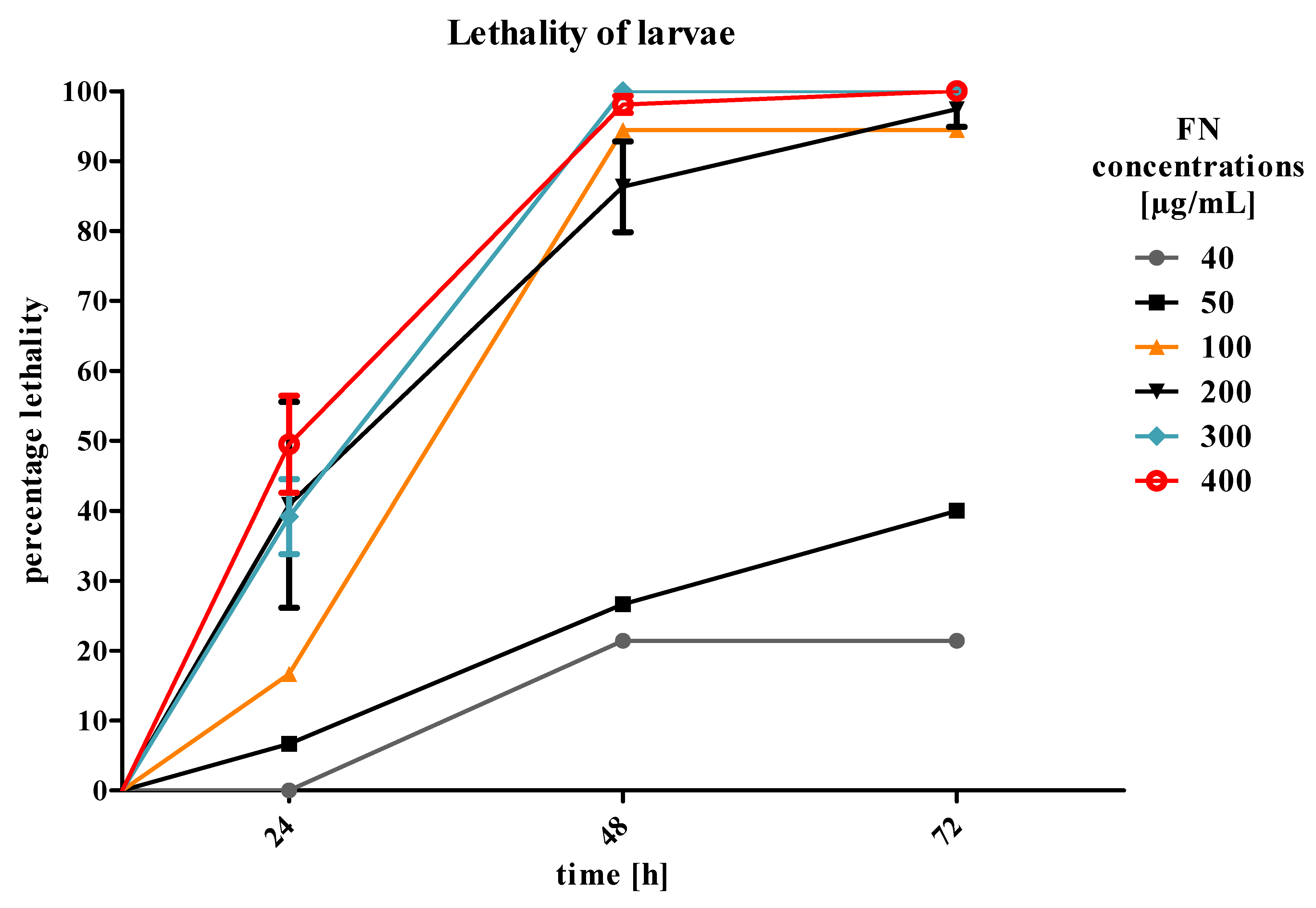
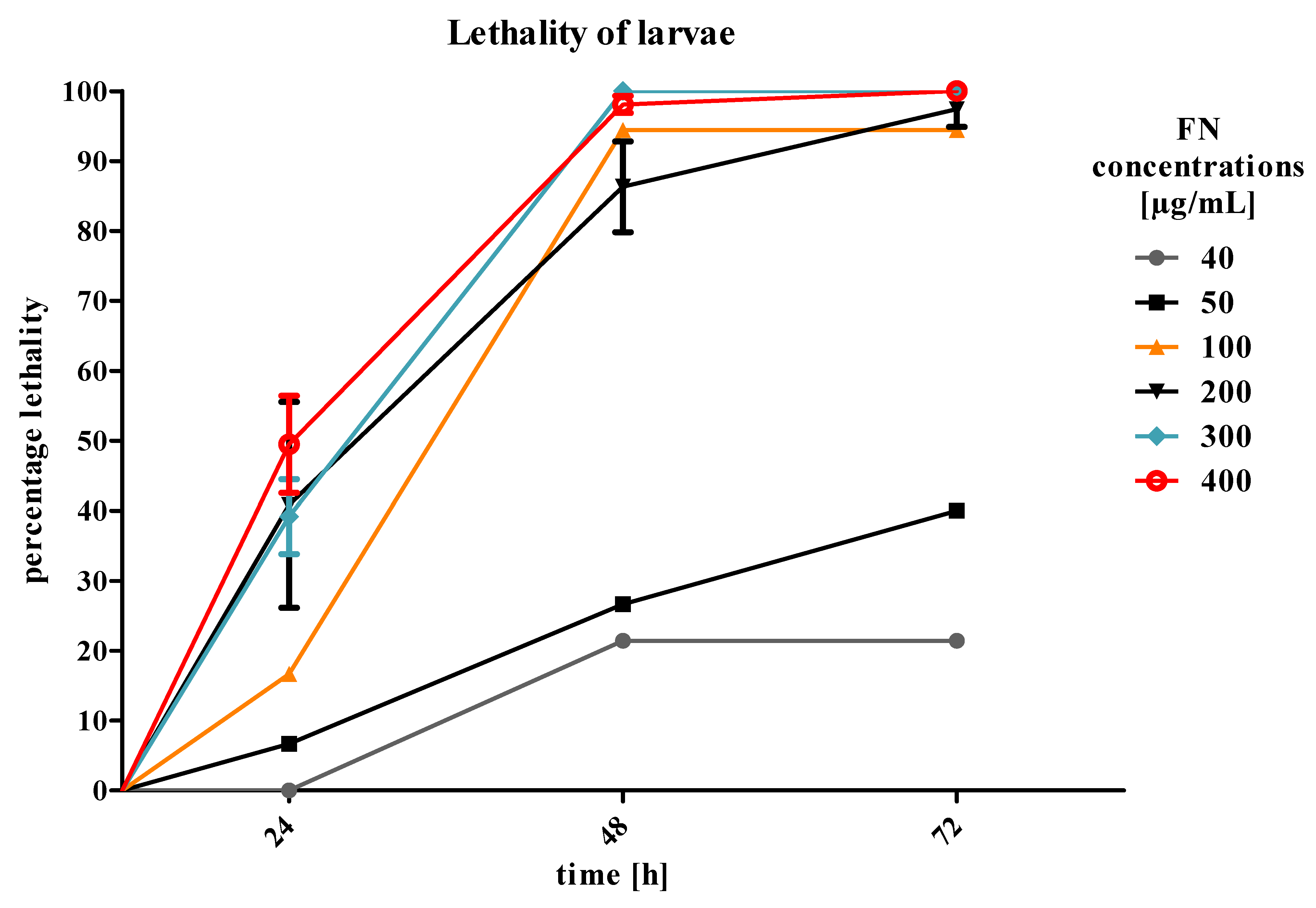
T. molitor larvae by additive action on the tissues crucial for the absorption and detoxification processes, such as fat body and midgut. What is more, we planned to mimic agrochemical application that would not last for more than two-to-three days. Therefore, we focused on the first results and acute toxicity effects. However, the larvae did not show any significant increase in mortality after 24 h of treatment. In addition, in the highest concentration of EXT (10%) mixed with FN, a slight decrease in mortality was observed (
Figure 2). Due to the observed effects, the strategy of application has been changed and the EXT in four concentrations was given to the nourishment for 24 h before application of FN in the concentration of 300 μg/mL, which was a concentration causing 40% of lethality of
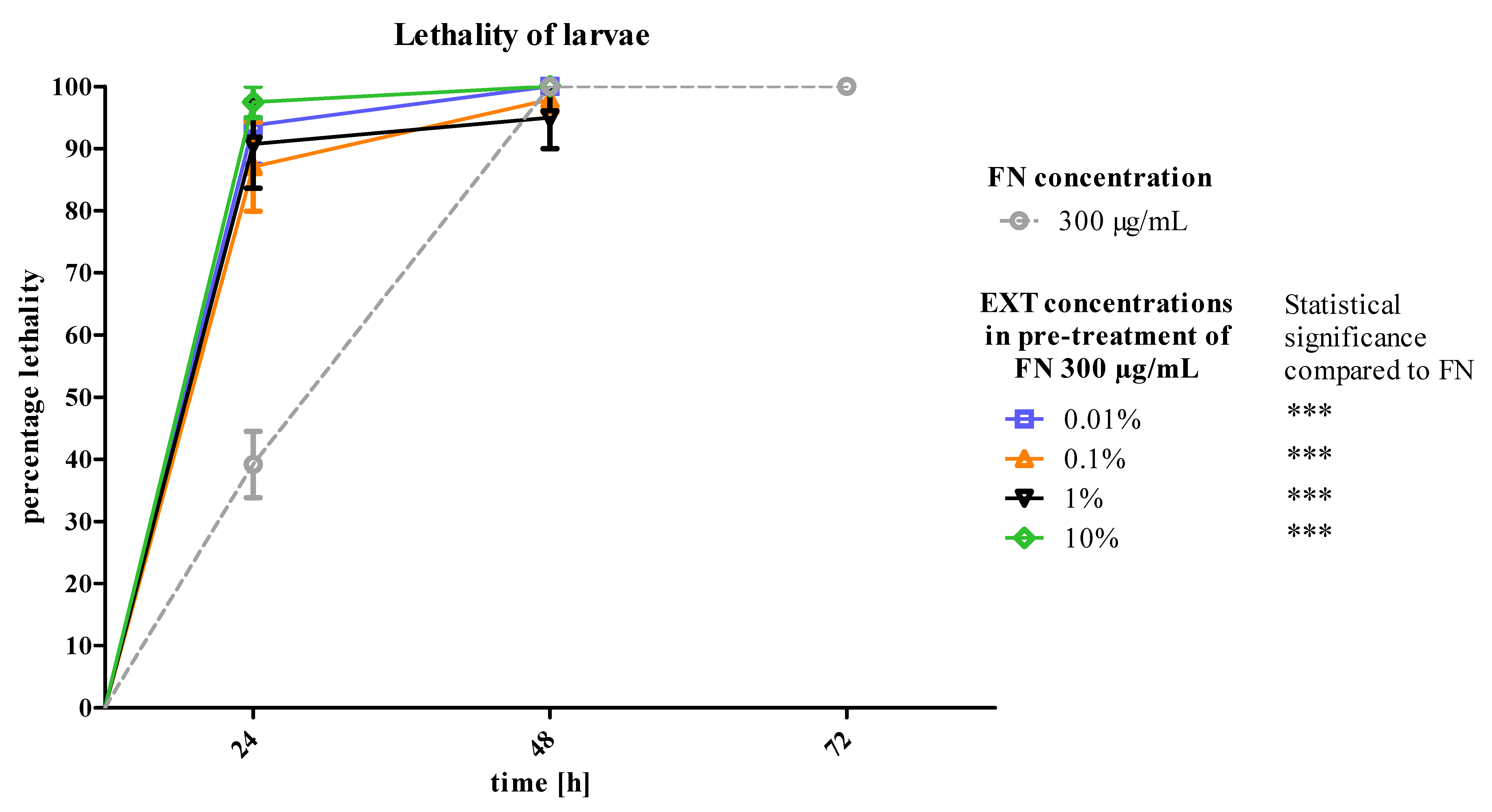
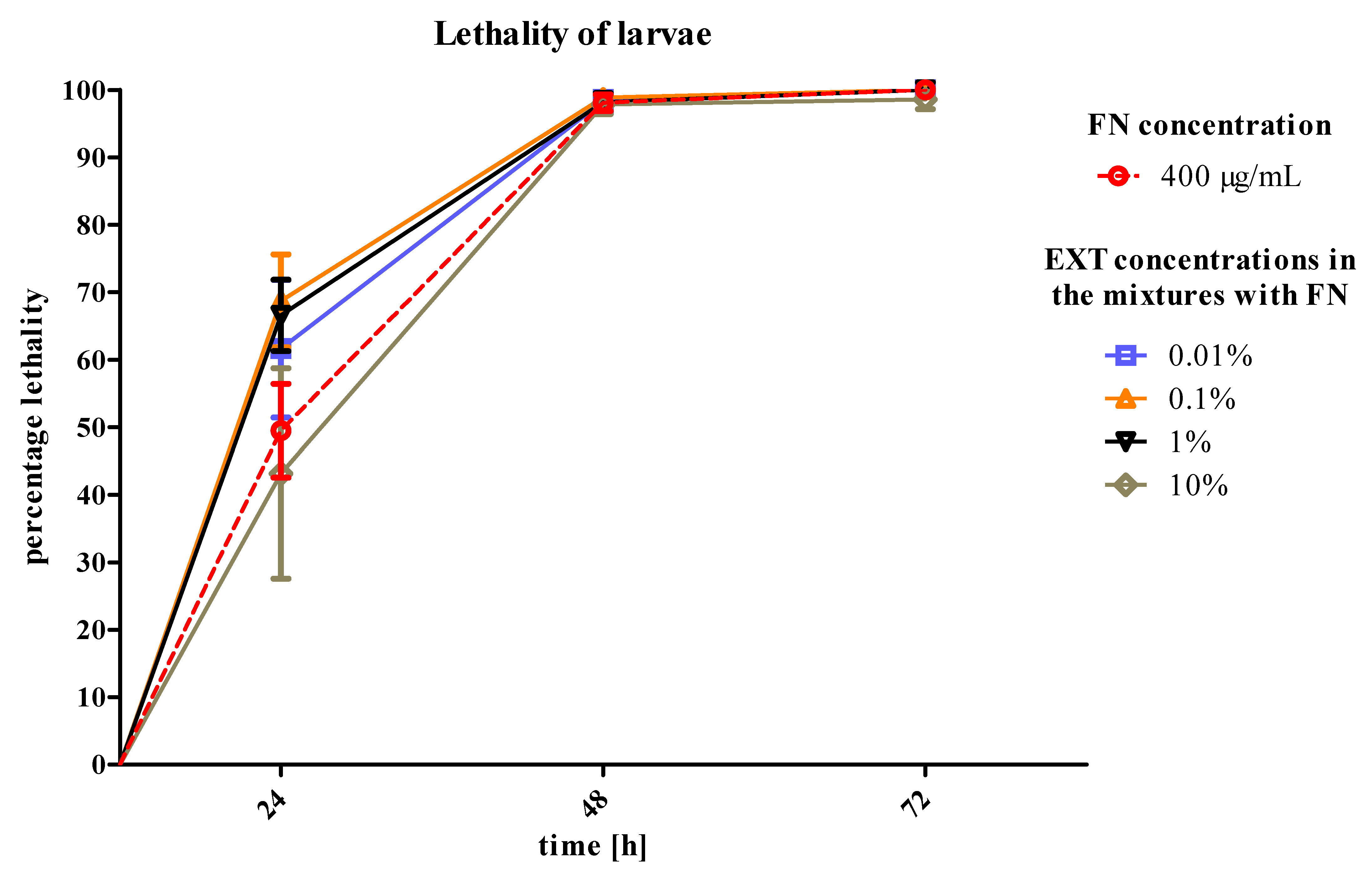
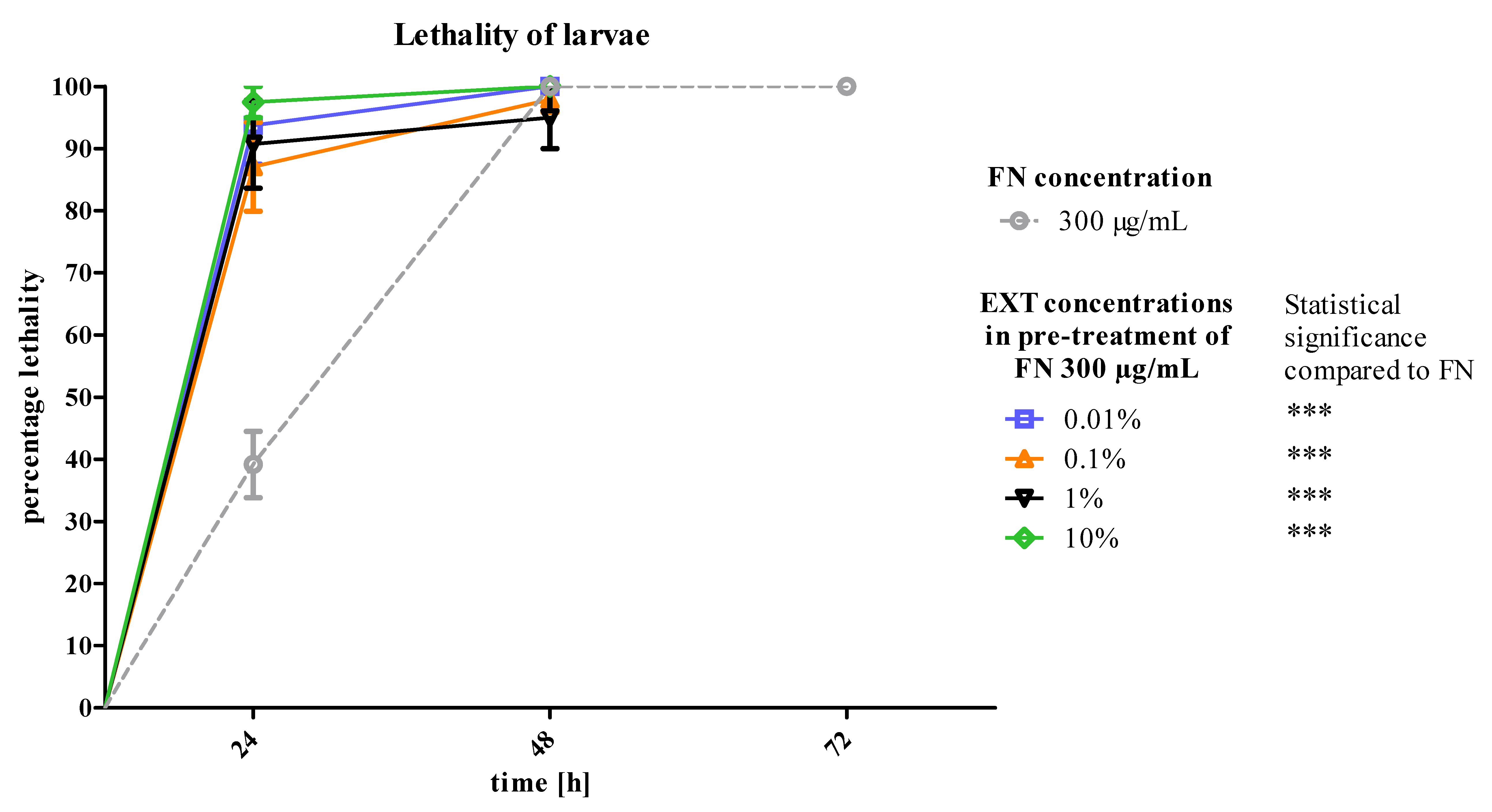
T. molitor larvae after 24 h of treatment. The second variant of the experiment showed that the mortality of larvae increased significantly after application of FN, when the EXT was applied 24 h before (
Figure 6). That may prove the increase of the sensitivity of larvae to synthetic insecticide under the influence of the EXT. In case of this strategy, the potentiation of the lethal toxicity of FN by EXT was observed.
The loss of the body mass of larvae treated with this mixture was significantly lower from the loss of the body mass of starving larvae (
Table 1) which suggests high energy expenditure for the detoxification process [
32,
33]. Also, this proves that the lethal effect was not caused by deprived feeding. The loss of the weight by the larvae could be the result of water loss through open spiracles as a consequence of the paralysis and an increase in the metabolic rate after the insecticide exposure [
34]. Similar results appeared in the second variant of the experiment.
Sublethal effects on the fat body of
T. molitor larvae caused by
S. nigrum extract were previously described [
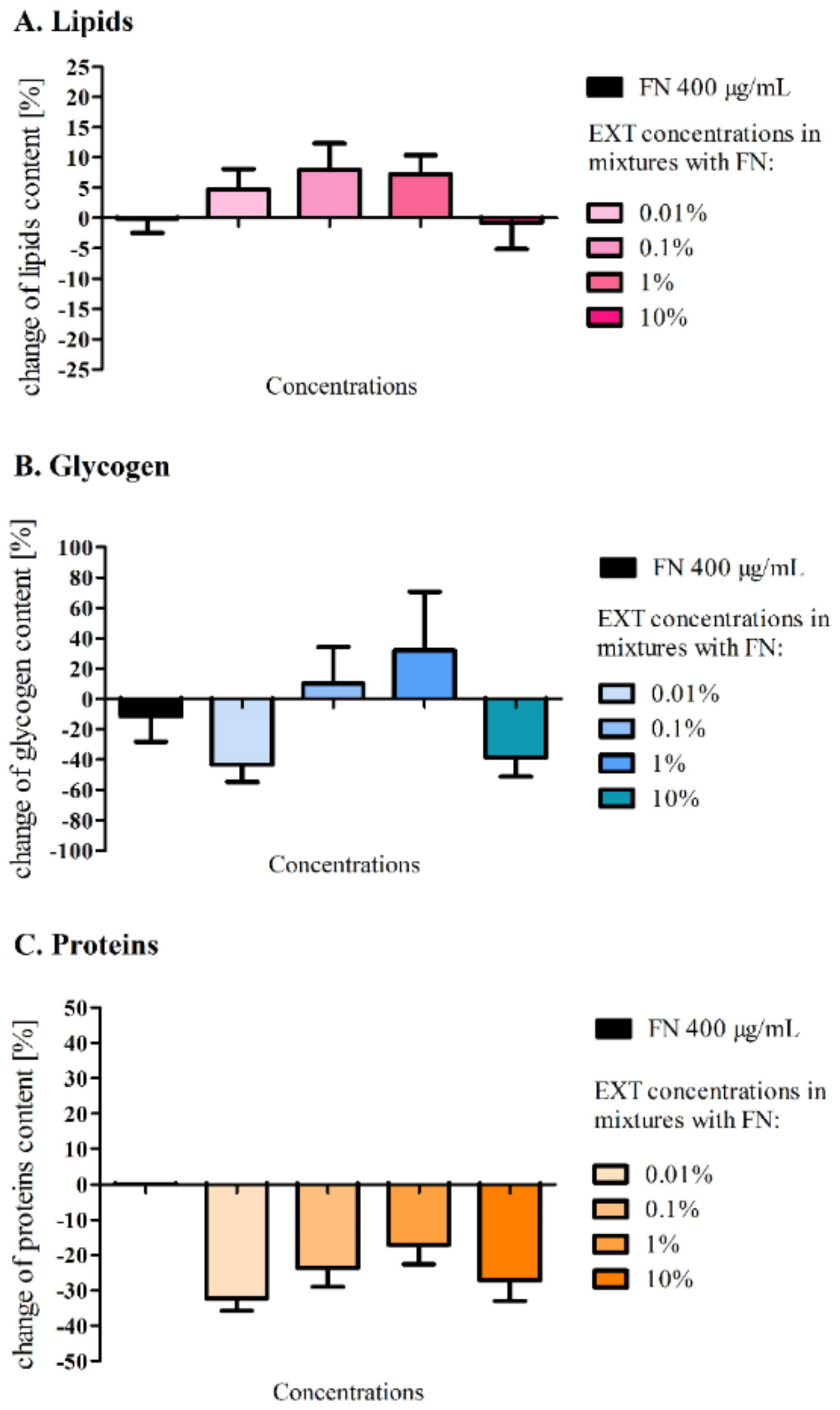
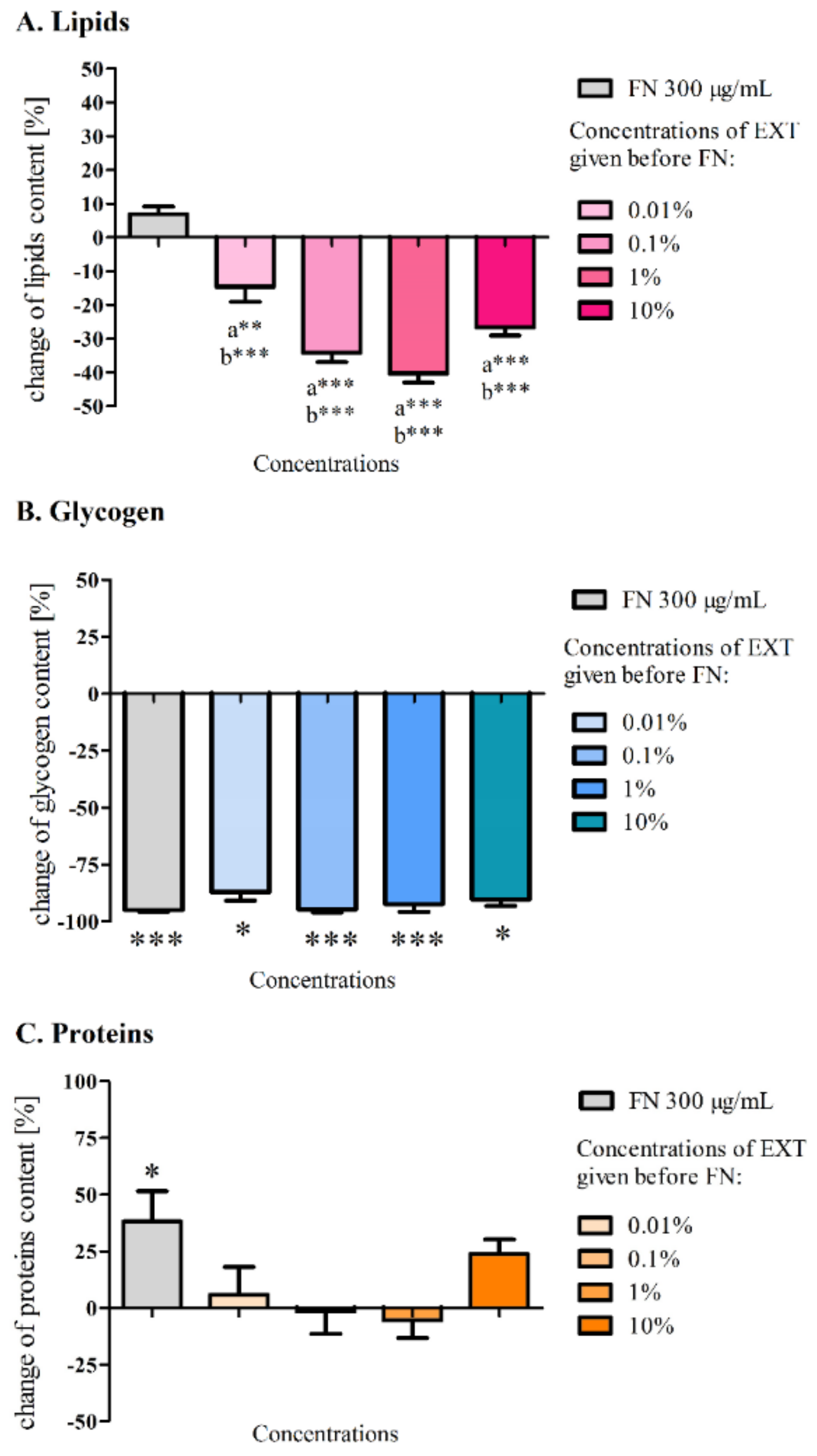
12]. The EXT caused a significant decrease in the glycogen level at a concentration of 0.1% and decrease of lipids content at a concentration 1% in the fat body cells. Further biochemical analyses of the levels of glycogen, lipids and proteins (
Figure 3) in the first variant of the experiment did not show any significant changes compared to the control, which may be a confirmation of a short and strong neurotoxic, lethal effect caused by FN in the chosen concentration and its mixtures with the EXT, rather than other metabolic effects. It can also indicate slight changes of quantity or quality of particular compounds among tested substances that might have not been visible, due to the short period of exposure. Perhaps, the changes would be significant after longer exposure. However, such a strategy of application would not mimic the mode of agrochemical treatments we wanted to follow. This is confirmed by the work of Wojciechowska et al. [
35], where the composition of the fat body changed in terms of the quality of compounds such as amino acids, fatty acids, cholesterol and carboxylic acids, in larvae treated with various types of insecticides. However, in our studies, particular fractions of lipids were not analysed. In the second variant of the experiment, the comparison of the amount of lipids after application of FN (300 μg/mL) and FN (300 μg/mL) with pre-treatment of EXT gave significant differences. The additive effect can be confirmed by the previous observations of lipid content in the fat body after the EXT application [
12]. In this case, the acute toxic effect was postponed, and some detoxification mechanisms could appear. Additionally, lipids are used as the first energetic substrates [
32] in cases such as intoxication.
Glycogen levels varied widely in the first variant of the experiment, which can be explained not only by the different state of food intake by each larvae [
36], but also by the acute toxicity, which was tested in our experiment, did not allow for the development of the extensive detoxifying strategy. Perhaps, that is the reason why the observed alterations were not statistically significant. Possibly, larvae might have used energy for detoxification from another source, such as trehalose from hemolymph [
37,
38], which level was not examined in our studies. In the case of the second variant of our experiment, each used concentration decreased glycogen level significantly. FN in the sublethal and lethal doses given in the diet of the silkworm
Bombyx mori was proven to decrease the glycogen level in the fat body [
37] which confirms that this compound can change the energetic metabolism in insects.
Protein level in the fat body increased notably after feeding the larvae with FN (300 μg/mL). Other research reports decrease of protein level after application of FN to
B. mori [
39] and increase when insecticides from other groups and Deodar oil were applied to
T. molitor [
10]. These insects may use different compensatory mechanisms in response to stress, also, different used substances and strategies of their application may develop different reaction.
In our previous study, the EXT in the concentration of 0.01% given for 3 days to larvae, did not show any visible changes in the midgut cells ultrastructure. The concentration of 1% was responsible for the disruption of nuclear membranes and 10% EXT additionally for the appearance of glycogen vacuoles, a sign of changes in its metabolism in the midgut cells [
12]. The above-mentioned observations were neither visible after the application of FN and FN–EXT blends nor in the midgut cells from larvae pre-treated with the EXT for 24 h and then FN for 48 h. Possibly, the exposure on the EXT was to short and as a result of an acute toxic action of FN, EXT did not penetrate the cells. However, this proposal is difficult to agree with, because the effects of FN–EXT blends were present in the fat body, which points that both glycoalkaloids and FN or their metabolites had to cross the midgut epithelium and reach the fat body causing malformations. Similar results were observed by Büyükgüzel et al. [
40], where boric acid caused much more significant alterations of trophocytes of the fat body, than of the epithelial cells of the midgut.
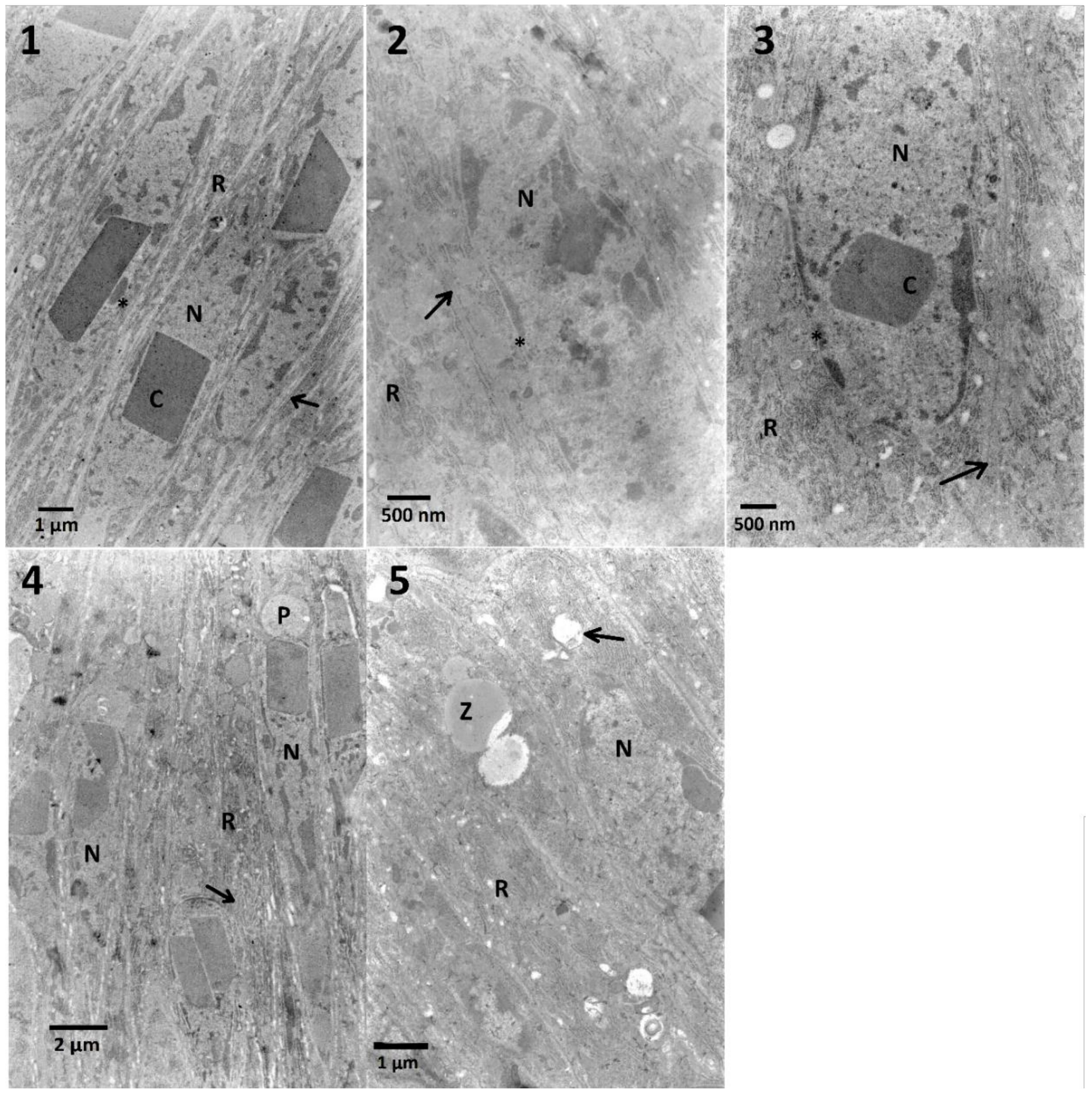
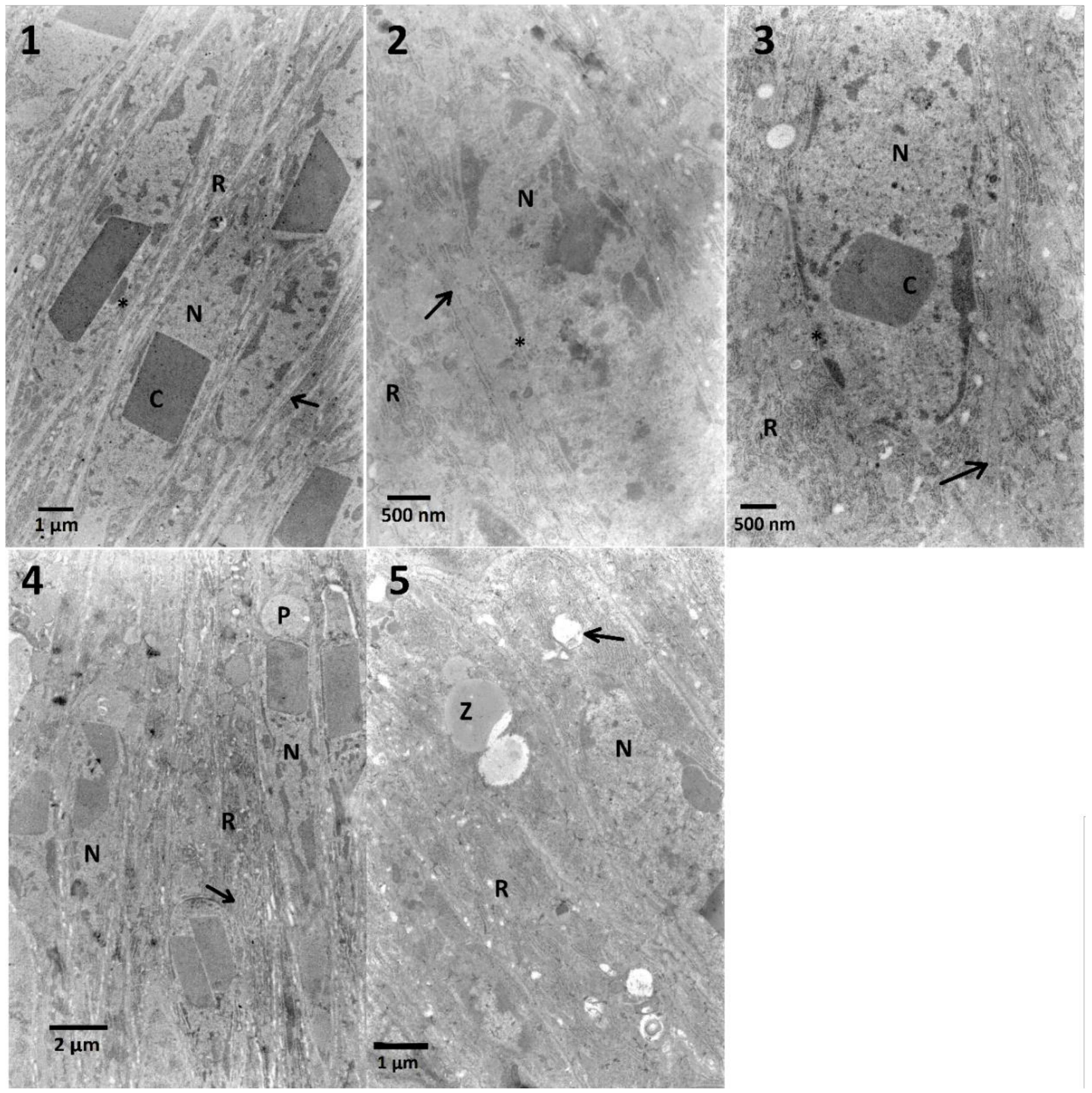
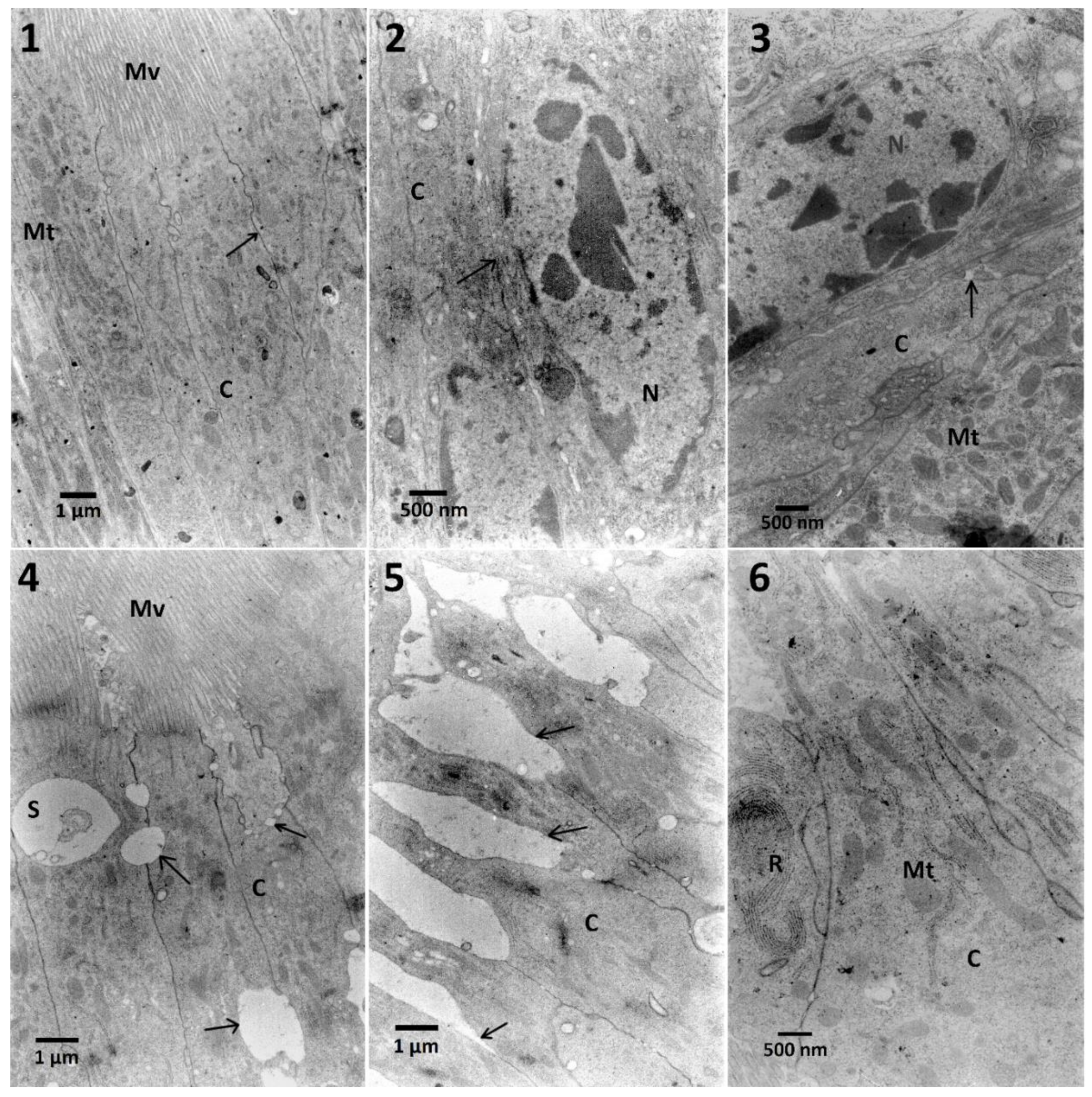
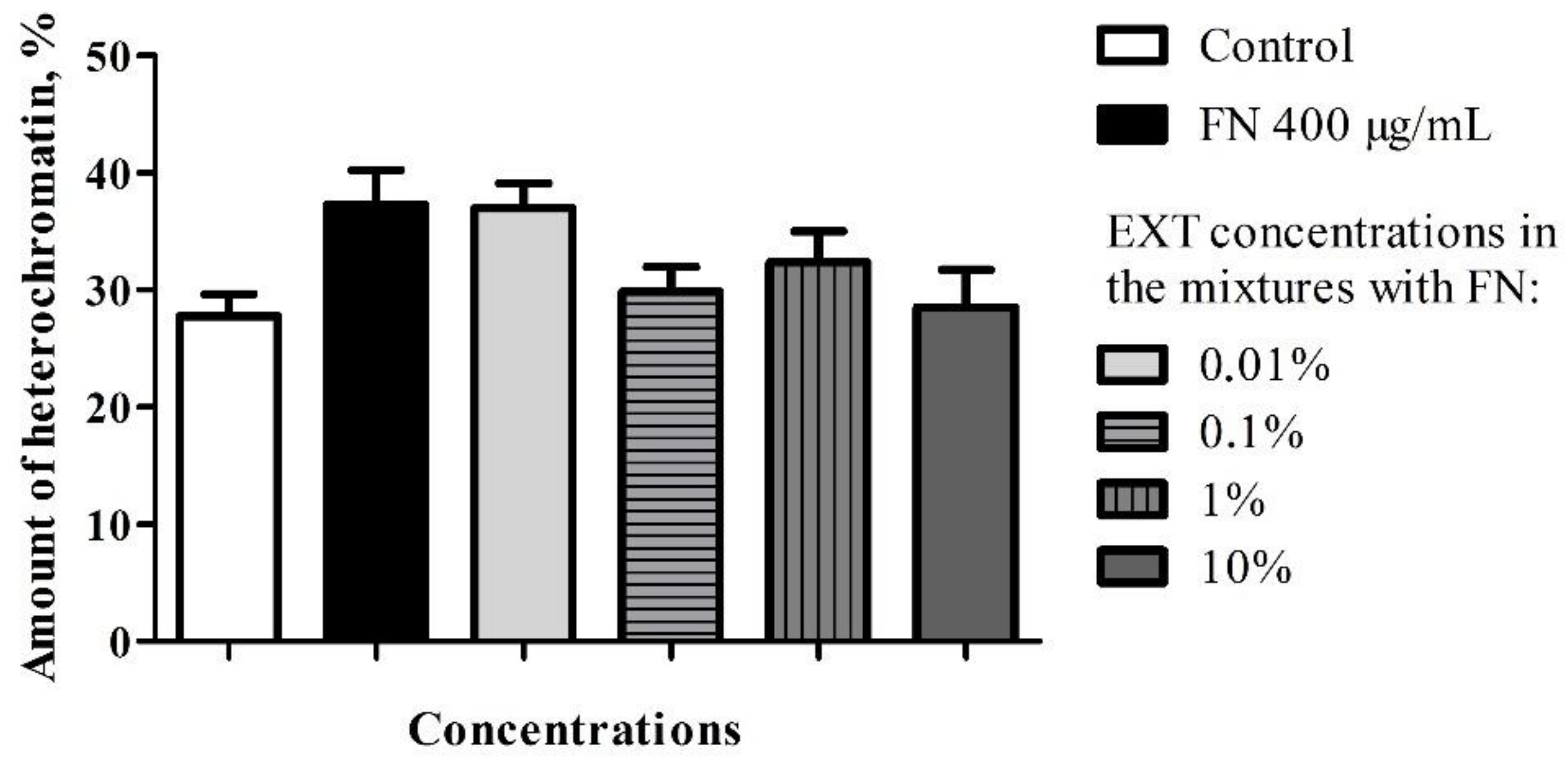
In the case of midgut cells in the first variant of the experiment, neither synergy, nor additive effect between FN and EXT were observed (
Table A1). The calculation of the correlation coefficient in the midgut cells for determination of chromatin density (
Figure A1) showed moderate negative correlation which suggests the possible inhibition of FN activity by EXT in these cells. Midgut cells also exhibited electron-dense cytoplasm and abundance of RER, where in the two highest concentrations its swelling was observed (
Table A1,
Figure 4). However, in the second variant, different effects appeared, such as cellular membrane disruption. Cells were also rich in RER in all tested concentrations, and the swelling of ER was present (
Table A3). This can be explained by the fact, that synthetic insecticides can cause disturbance of ER membranes and cell membranes in the midgut cells as well as mentioned chromatin condensation [
41]. Both synthetic and bio-insecticides are responsible for oxidative and nitrosative stress [
42], which can result in observed effects.
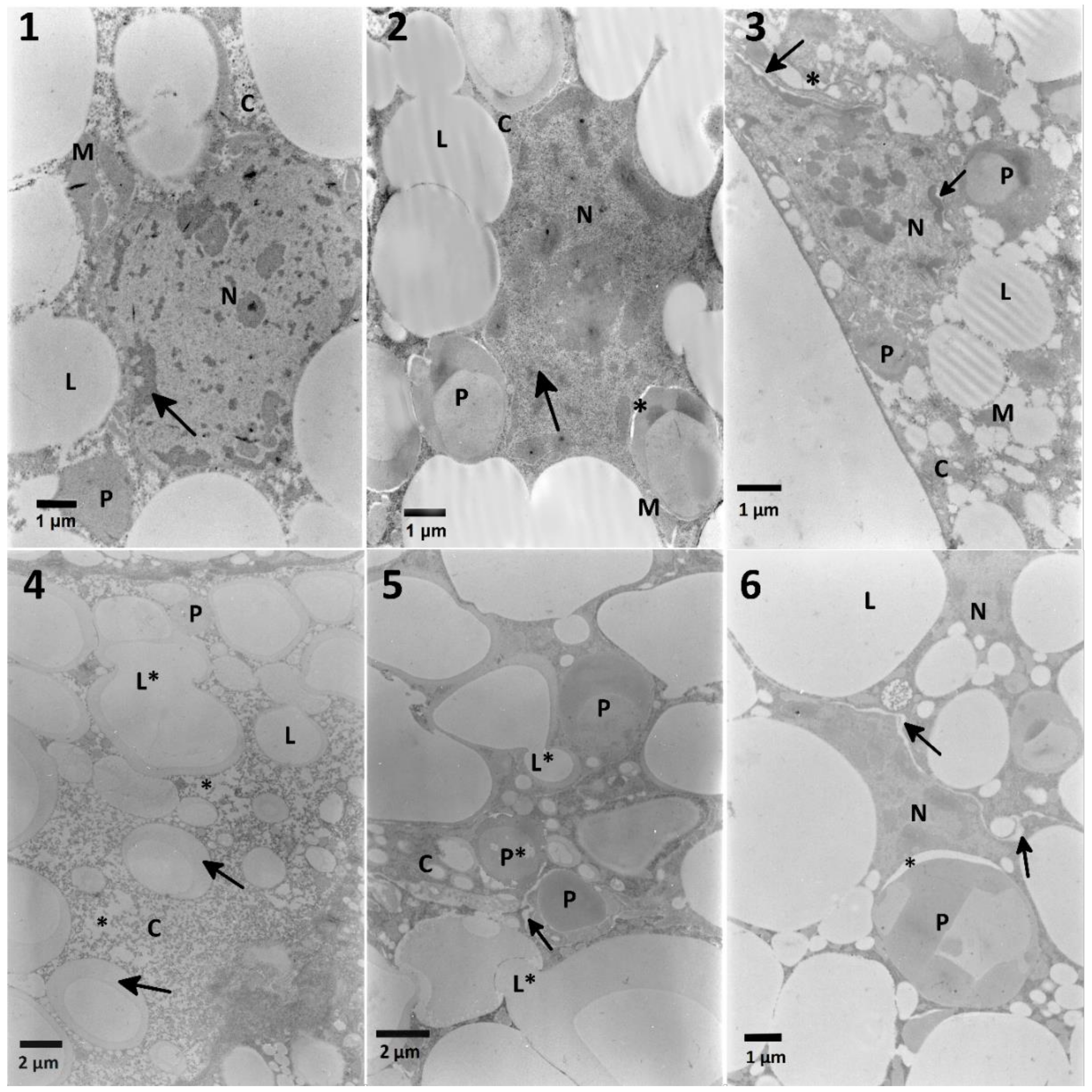
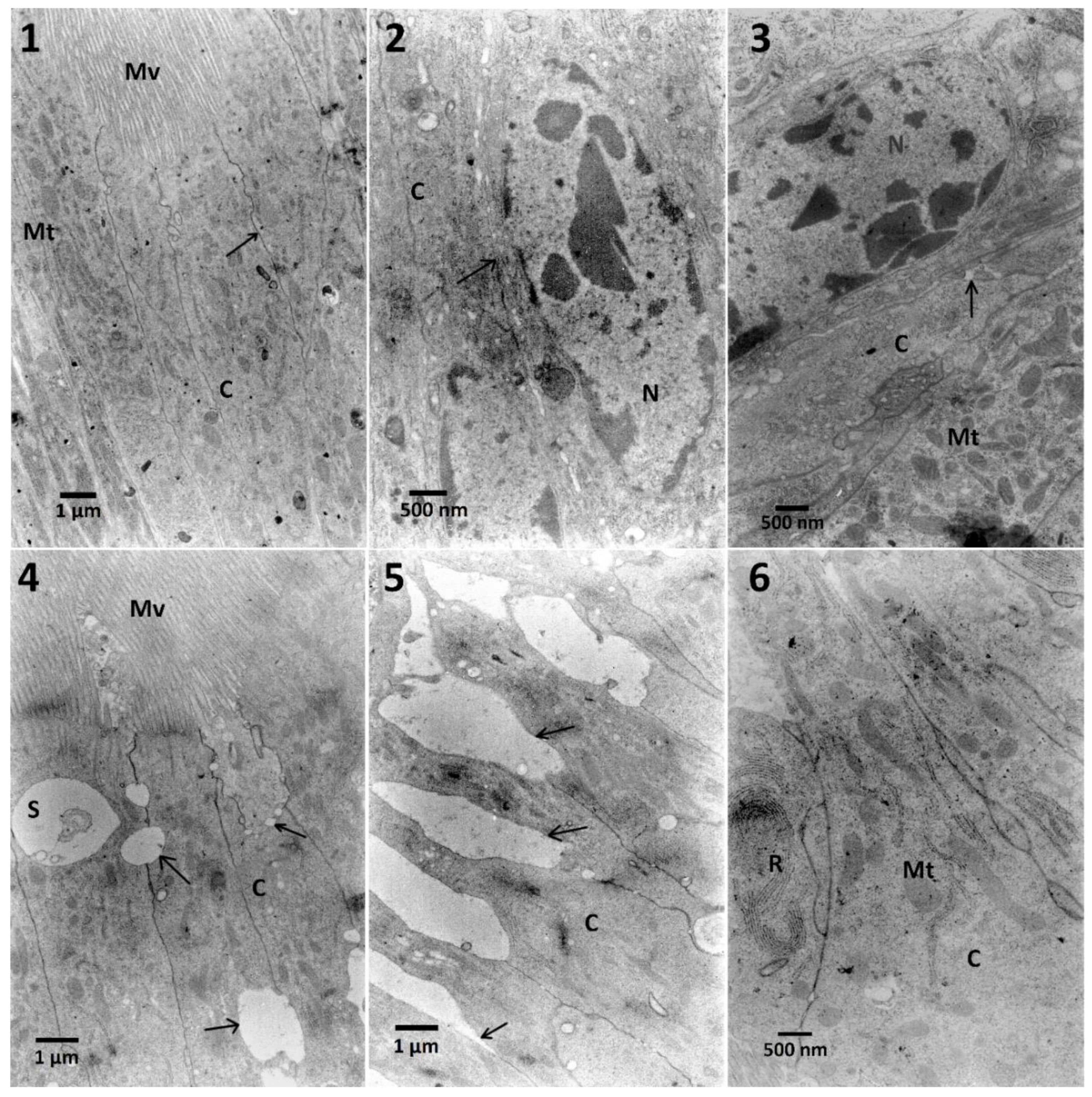
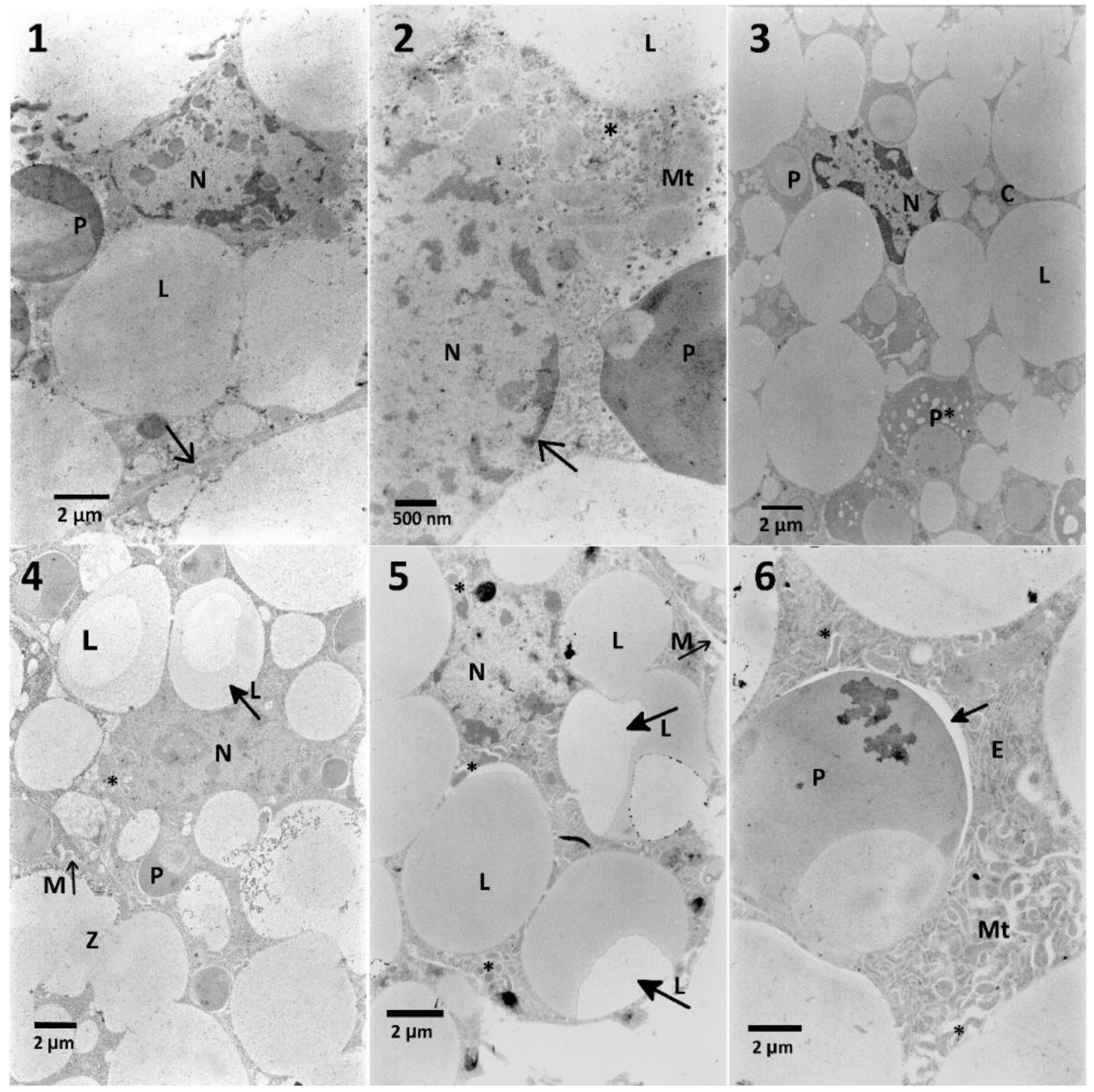
Effects observed in the fat body cells obtained from insects treated only with the
S. nigrum extract were limited to the decreased cytoplasmic density and the homogeneity of lipid droplets and an increased electron-density of nuclei. Additionally, solasonine and solamargine were responsible for disturbance of lipid droplets and stored proteins [
12]. The effects of cell disruption were observed in the ultrastructure of the fat body (
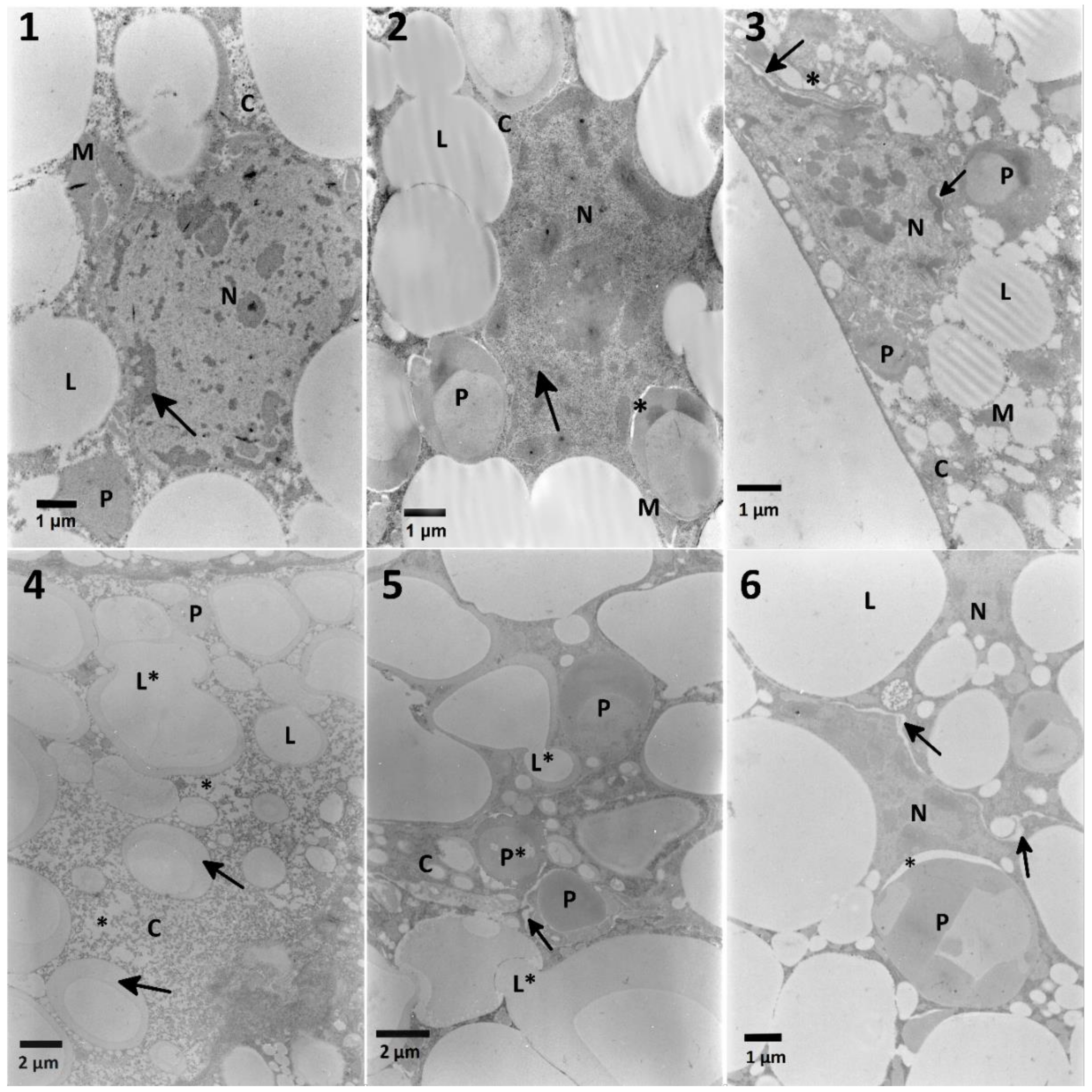
Figure 5,
Table A2) in both variants of experiment. The mixtures of
S. nigrum extracts caused additional effects such as fusion and loss of homogeneity of lipid droplets (
Figure 5, No. 4,5) suggesting some additive effect. What is interesting, these changes were not noted in the lowest and the highest used concentrations of the extract in mixtures. The fat body cells of larvae treated with EXT and then FN (
Table A4;
Figure 9) showed different signs of intoxication, such as vacuolization of cytoplasm around mitochondria, stored proteins, and endoplasmic reticulum, which can be symptoms of disruption of cellular membranes caused by the EXT.
Most found nuclei in the fat body cells, and also midgut cells from larvae treated with FN mixed with the EXT featured with electron-dense chromatin. Observed effects can be a sign of the response of insects to contact with xenobiotics, and may indicate the early beginning of pyknotic processes, which are characterized by shrunken and electron-dense nuclei [
43,
44]. The ultrastructural malformations of fat droplets are most probably due to high lipophilic properties of FN. Besides, lipids are the first reserves, used before glycogen or other energy-rich substrates [
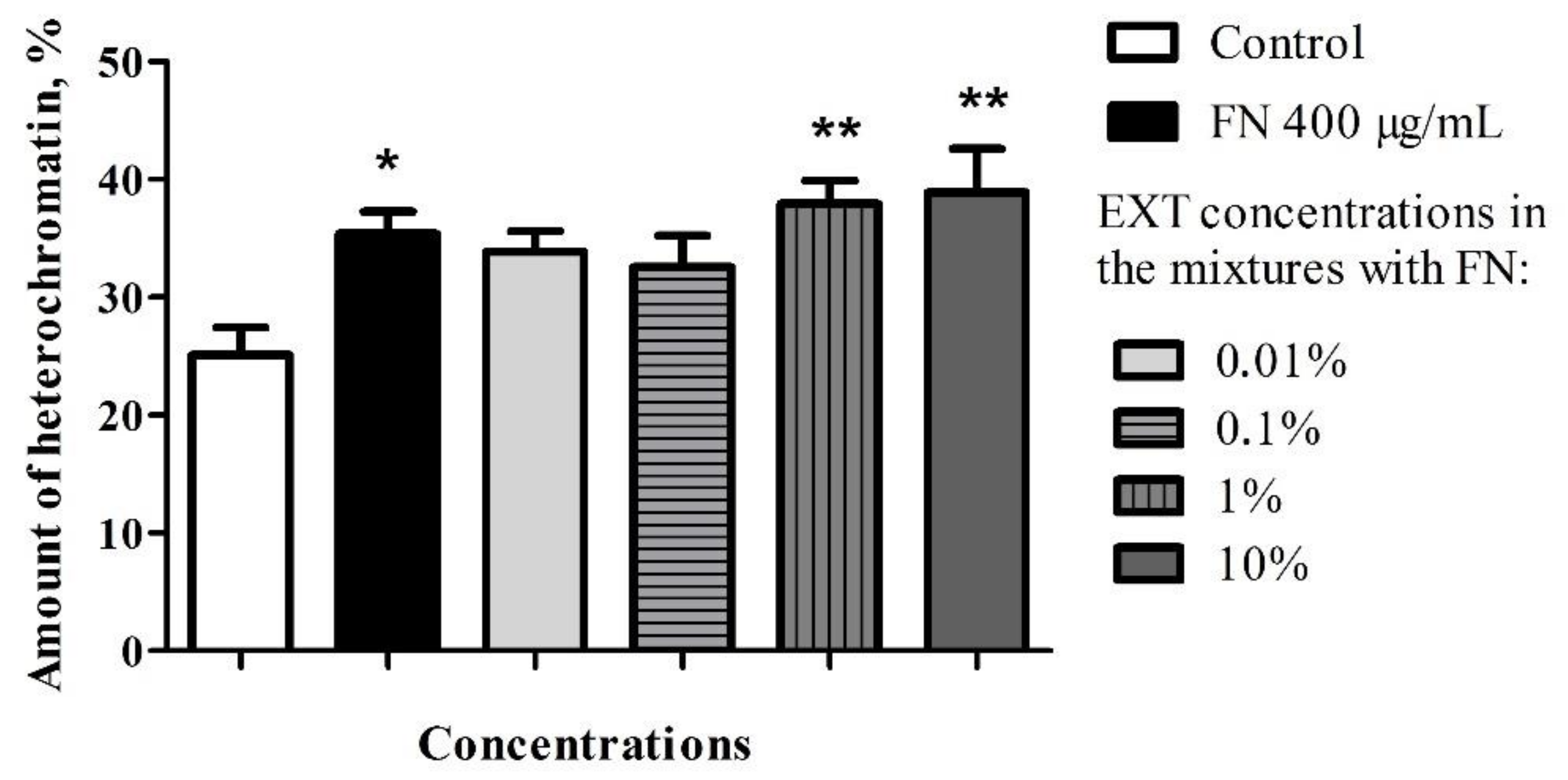
32]. Strong positive correlation obtained in the chromatin density of fat body cells treated with FN–EXT blends, suggest the increase of FN toxicity caused by increasing concentration of the EXT, especially well visible in the highest used concentration of EXT in blends (
Figure 5, No. 6).
The malformation of biological membranes caused by glycoalkaloids (solasonine and solamargine) has been previously described [
45]. They cause easier penetration of toxic substances to the cell [
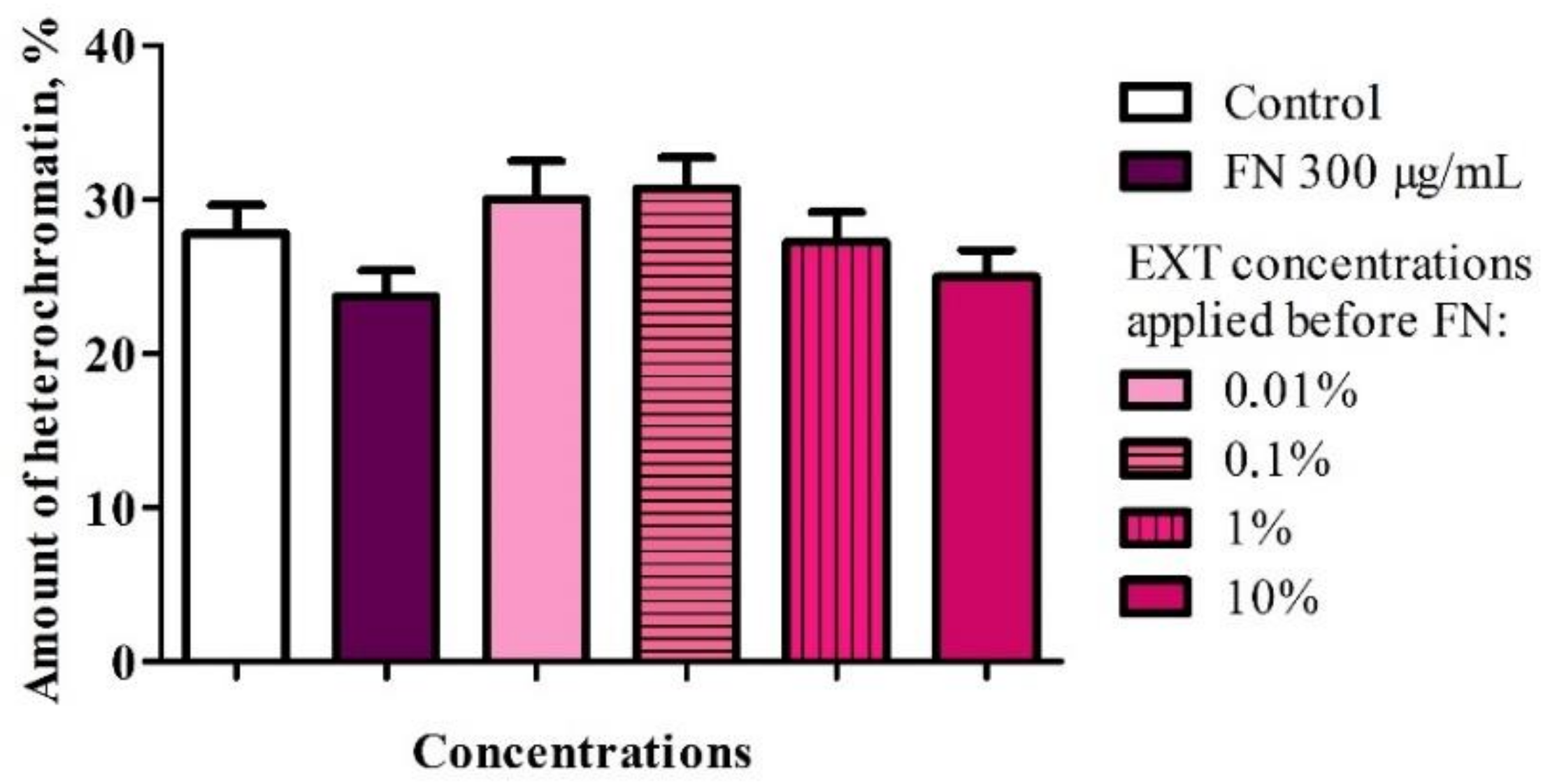
46], which could be an additive factor to increase FN toxicity. The changes in the chromatin condensation in the fat body after application of FN and with pre-treatment of EXT also suggests a potentiation effect (
Figure A3), confirmed by calculated correlation coefficient, which showed strong positive correlation. Also, some pure glycoalkaloids induce liposome disruption and hemolysis [
47]. Therefore, the observed malformations of cellular membranes and lipid droplets could be observed in our research. Next, FN may increase polarity and lead to enhanced water penetration [
48]. In consequence, osmolarity of cytoplasm, shrinkage or swelling of intramembranous space of membrane-bound organelles may be observed.
Abovementioned observations suggest (1) relatively fast and unharmful transfer of toxic substances such as FN that is a small, non-polar molecule and is able to migrate through the cell membrane or by the paracellular pathway [
49], and (2) their longer persistence in the fat body, which is the main detoxifying organ in larvae. Besides, due to high lipid content, it enables a high accumulation of toxins within fat body cells. This manifests in the disturbance of cellular and nuclear membranes, and an increase of the chromatin and cytoplasm electro-density, which next can lead to further changes in metabolism [
50].
In several studies, plant-derived substances were used in binary mixtures with synthetic insecticides on insects due to increase of toxic properties, mainly by extension of mode of action on different targets obtaining synergistic effects. In many studies, the application of both plant-derived and synthetic insecticide with obtained synergism or additivity, were performed without creating their mixtures before the application [
51,
52]. First, we focused on a more practical aspect, where the mixtures were created to avoid multiplication of the field application. A similar strategy was used in studies of Maurya et al. [
53], where imidacloprid mixed with crude petroleum ether
Ocimum basilicum leave extract in ratio 1:1 showed synergistic properties against
Anopheles stephensi than imidacloprid applied alone. Fenitrothion blended in ratio 1:1 with
Callitris glaucophylla, Daucus carota or
Khaya senegalensis extract was effective against
Culex annulirostris [
54]. Note that in both cases, FN and LC
25 extracts were calculated and the final summary of effectiveness of both used substances could be predicted and compared with results. In our studies, we used EXT, which did not cause lethal effects [
12], which could result in no increase in lethality in the first variant of the experiment. However, other studies of Maliszewska and Tęgowska [
55] shows that capsaicin which does not have insecticidal properties, added to an organophosphorus insecticide, methidathion, increased its toxicity. Therefore, the effects may be alkaloid-specific. We decided to perform the second variant of the experiment and avoid blending used chemicals, which resulted in a massive increase of larvae lethality, proving the potentiation—the effect when non-lethal substance increases lethality of a toxic compound. A similar strategy was used on a mosquito (
Aedes aegypti) with 4 h exposition on essential oils vapors before deltamethrin topical application, which also resulted in toxicity growth [
56]. The effects prove that the strategy of application may significantly affect results of the agrochemical strategies.
To sum up our experiment results, we can agree that increased lethality, significant changes in the levels of glycogen and lipids and ultrastructure malformations prove that the second variant of the experiment is more effective. We assume that the mode of action of the extract depends on its sublethal effects [
12], such as the membranes malformations that increase the FN permeation. Second, the detoxication of previously applied EXT may decrease the energy resources necessary for FN detoxication. And last, observed changes in the lipids stored in the fat body may decrease the ability to store the lipophilic xenobiotics (such as FN) in the fat body and their spreading in other tissues, i.e., nerve tissue.

 ,
, 


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}