Cross-Reactive Carbohydrate Determinant in Apis mellifera, Solenopsis invicta and Polybia paulista Venoms: Identification of Allergic Sensitization and Cross-Reactivity

 and
and
Abstract
:1. Introduction
2. Results
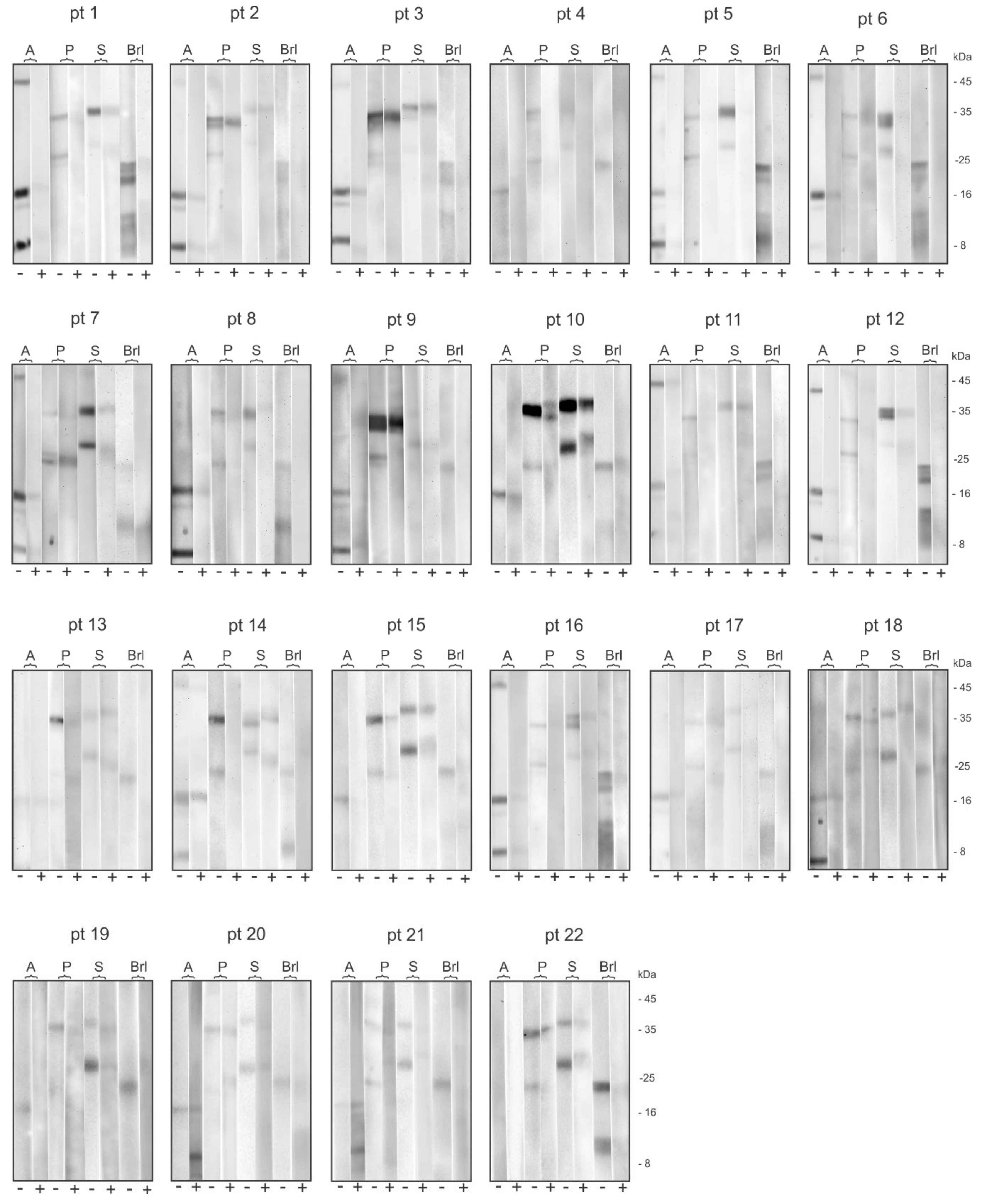
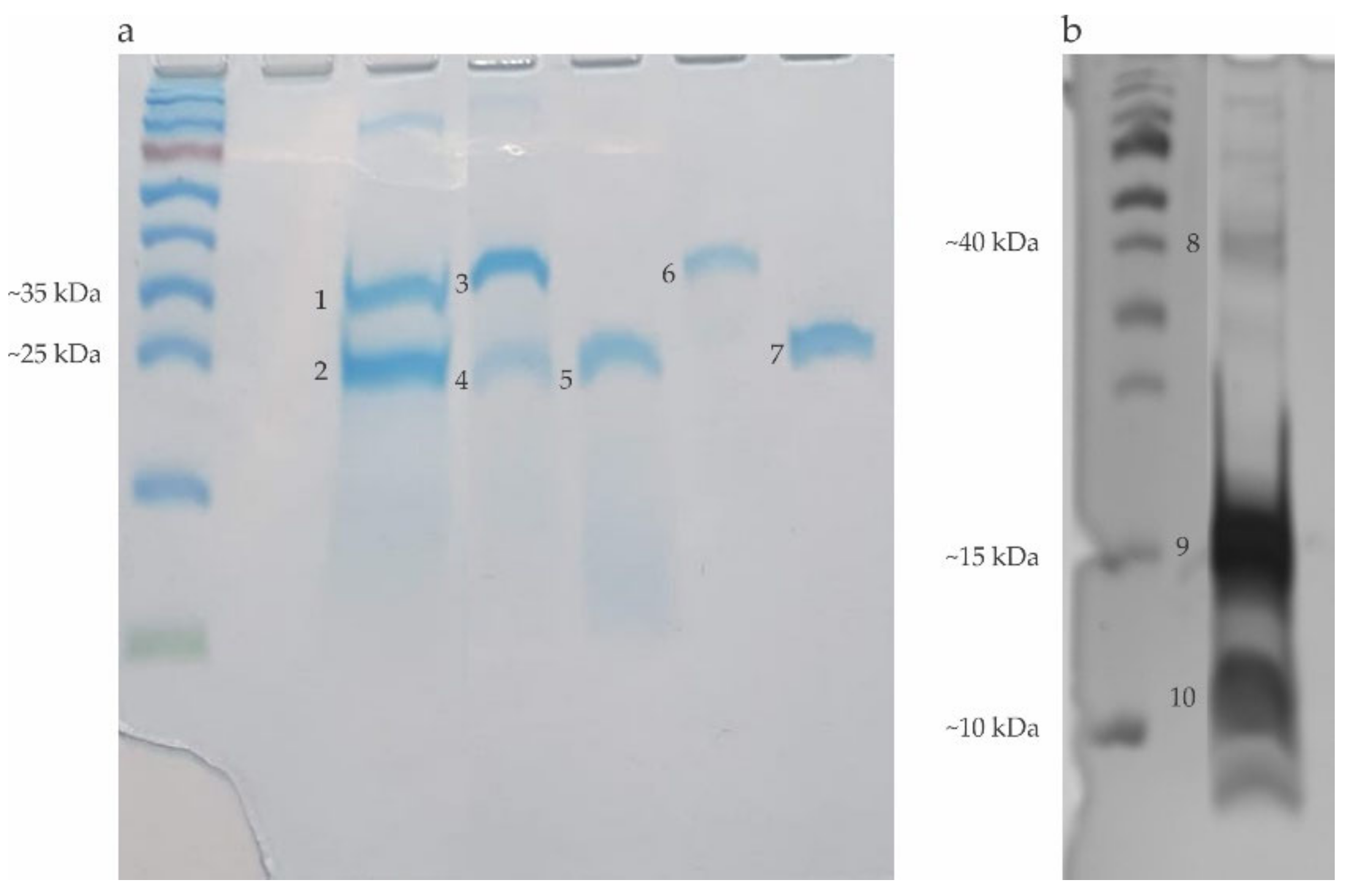
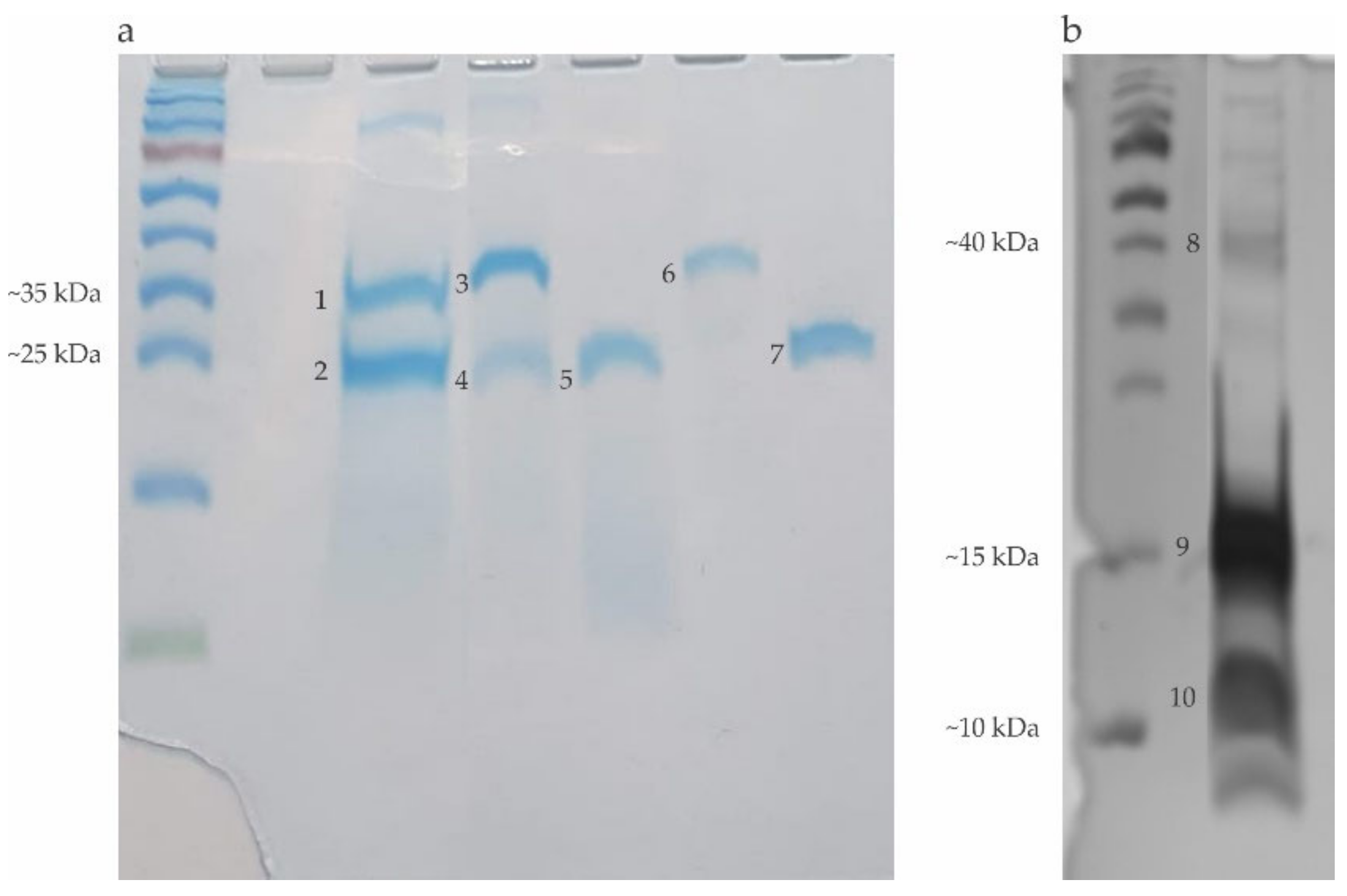
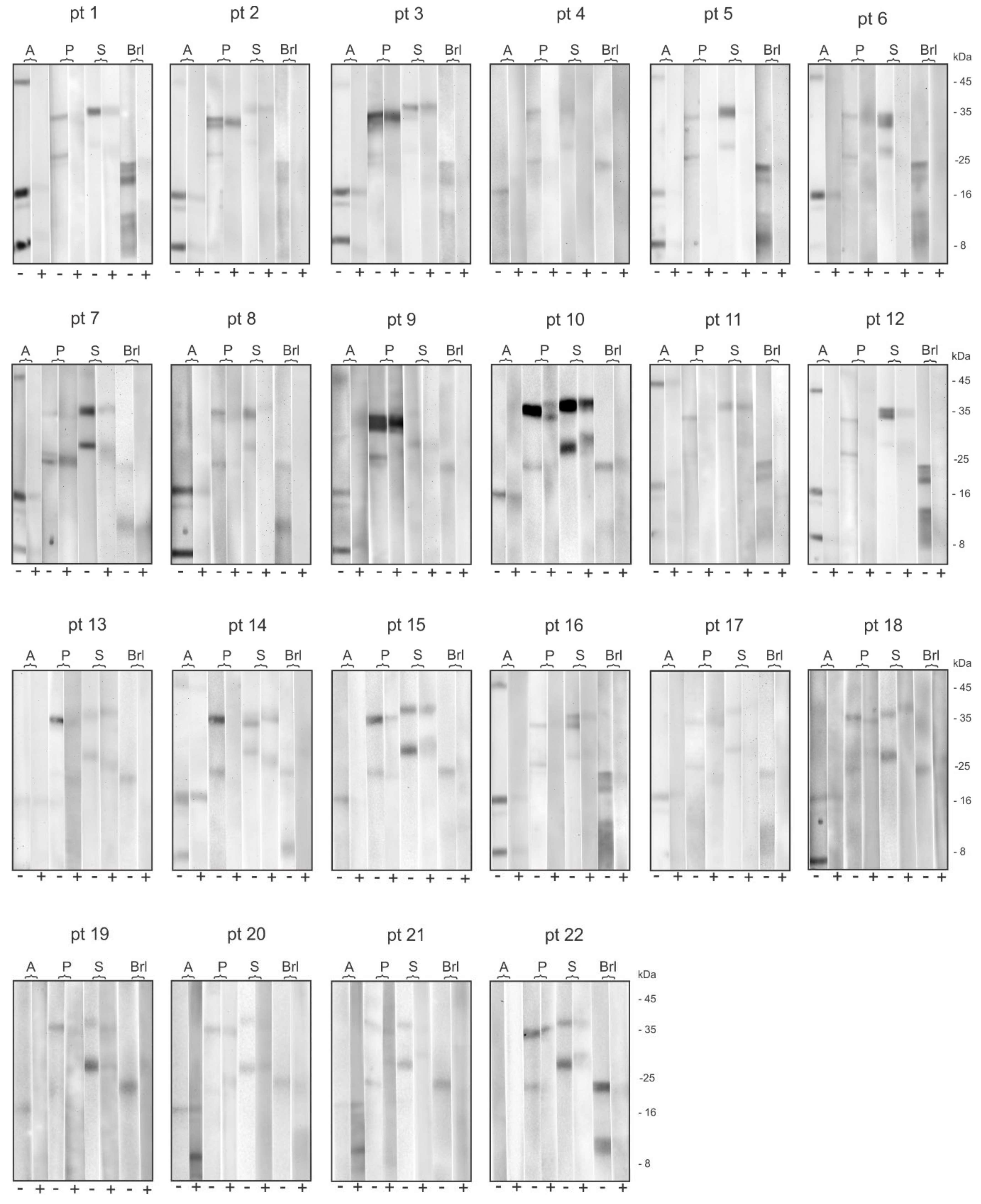
2.1. Electrophoretic Profile of the Allergens Obtained in the Hymenoptera Venoms Extracts and Bromelain
2.2. Patient’s Specific IgE Reactivity Profile
2.3. CCDs sIgE Reactivity Profile
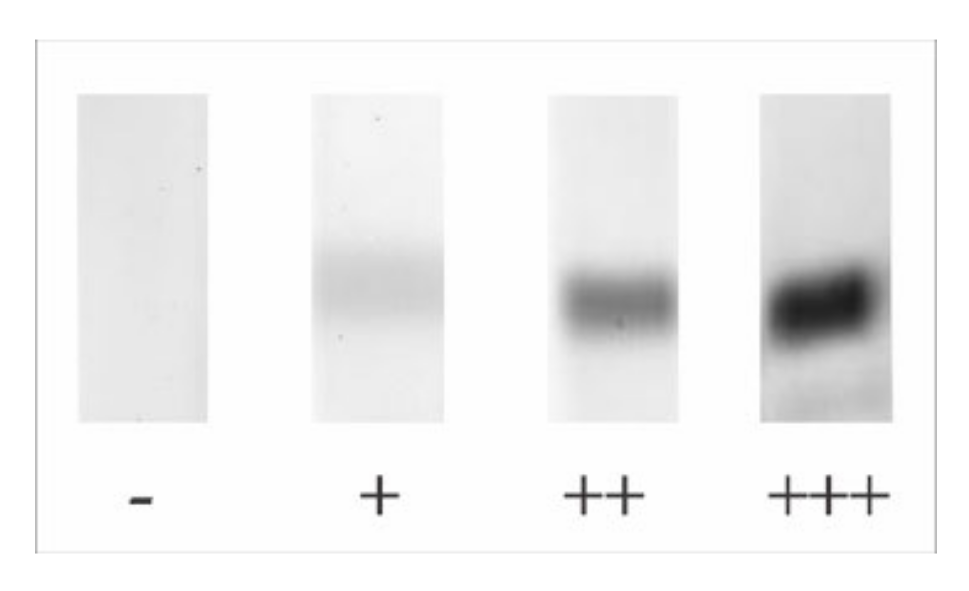
2.4. Bromelain Adsorption Effect on Patient’s Sera
2.5. IgE-Specific Reactivity to Non-Glycosylated Recombinant Antigens (rPoly p1, rPoly p5) from Polybia Paulista Wasp Venom
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Patients
5.2. Venoms Extracts and Bromelain Solution Preparation
5.3. Recombinant Allergens (rPoly P1 and rPoly p5) Obtention
5.4. Adsorption Tests with Bromelain and Serum
5.5. Electrophoresis and Immunoblot Assay
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A

References
- Tracy, J.M. Insect allergy. Mt. Sinai J. Med. 2011, 78, 773–783. [Google Scholar] [CrossRef] [PubMed]
- Bilò, B.M.; Bonifazi, F. Epidemiology of insect-venom anaphylaxis. Curr. Opin. Allergy Clin. Immunol. 2008, 8, 330–337. [Google Scholar] [CrossRef] [PubMed]
- Biló, B.M.; Rueff, F.; Mosbech, H.; Bonifazi, F.; Oude-Elberink, J.N.G.; Birnbaum, J.; Bucher, C.; Forster, J.; Hemmer, W.; Incorvaia, C.; et al. Diagnosis of Hymenoptera venom allergy. Allergy Eur. J. Allergy Clin. Immunol. 2005, 60, 1339–1349. [Google Scholar] [CrossRef] [PubMed]
- Fitzgerald, K.T.; Flood, A.A. Hymenoptera Stings. Clin. Tech. Small Anim. Pract. 2006, 21, 194–204. [Google Scholar] [CrossRef]
- Moreno, M.; Giralt, E. Three valuable peptides from bee and wasp venoms for therapeutic and biotechnological use: Melittin, apamin and mastoparan. Toxins 2015, 7, 1126–1150. [Google Scholar] [CrossRef] [Green Version]
- Antonicelli, L.; Bilò, M.B.; Bonifazi, F. Epidemiology of hymenoptera allergy. Curr. Opin. Allergy Clin. Immunol. 2002, 2, 341–346. [Google Scholar] [CrossRef]
- Krishna, M.T.; Ewan, P.W.; Diwakar, L.; Durham, S.R.; Frew, A.J.; Leech, S.C.; Nasser, S.M. Diagnosis and management of hymenoptera venom allergy: British Society for Allergy and Clinical Immunology (BSACI) guidelines. Clin. Exp. Allergy 2011, 41, 1201–1220. [Google Scholar] [CrossRef]
- Perez-Riverol, A.; Justo-Jacomini, D.L.; de Lima-Zollner, R.; Brochetto-Braga, M.R. Facing hymenoptera venom allergy: From natural to recombinant allergens. Toxins 2015, 7, 2551–2570. [Google Scholar] [CrossRef]
- Brehler, R.; Grundmann, S.; Stöcker, B. Cross-reacting carbohydrate determinants and Hymenoptera venom allergy. Curr. Opin. Allergy Clin. Immunol. 2013, 13, 360–364. [Google Scholar] [CrossRef]
- Ebo, D.G.; Hagendorens, M.M.; Bridts, C.H.; de Clerck, L.S.; Stevens, W.J. Sensitization to cross-reactive carbohydrate determinants and the ubiquitous protein profilin: Mimickers of allergy. Clin. Exp. Allergy 2004, 34, 137–144. [Google Scholar] [CrossRef]
- Hemmer, W. Cross-reactivity to honeybee and wasp venom. Hautarzt 2008, 59, 194. [Google Scholar] [CrossRef]
- Altmann, F. The role of protein glycosylation in allergy. Int. Arch. Allergy Immunol. 2007, 142, 99–115. [Google Scholar] [CrossRef] [PubMed]
- Matricardi, P.M.; Kleine-Tebbe, J.; Hoffmann, H.J.; Valenta, R.; Hilger, C.; Hofmaier, S.; Aalberse, R.C.; Agache, I.; Asero, R.; Ballmer-Weber, B.; et al. EAACI Molecular Allergology User’s Guide. Pediatr. Allergy Immunol. 2016, 27, 1–250. [Google Scholar] [CrossRef] [PubMed]
- Sturm, G.J.; Jin, C.; Kranzelbinder, B.; Hemmer, W.; Sturm, E.M.; Griesbacher, A.; Heinemann, A.; Vollmann, J.; Altmann, F.; Crailsheim, K.; et al. Inconsistent results of diagnostic tools hamper the differentiation between bee and vespid venom allergy. PLoS ONE 2011, 6, e20842. [Google Scholar] [CrossRef] [PubMed]
- Holzweber, F.; Svehla, E.; Fellner, W.; Dalik, T.; Stubler, S.; Hemmer, W.; Altmann, F. Inhibition of IgE binding to cross-reactive carbohydrate determinants enhances diagnostic selectivity. Allergy Eur. J. Allergy Clin. Immunol. 2013, 73, 1964–1978. [Google Scholar] [CrossRef] [Green Version]
- Yokoi, H.; Yoshitake, H.; Matsumoto, Y.; Kawada, M.; Takato, Y.; Shinagawa, K.; Sakurai, H.; Saito, K. Involvement of cross-reactive carbohydrate determinants-specific IgE in pollen allergy testing. Asia Pac. Allergy 2017, 7, 29–36. [Google Scholar] [CrossRef] [PubMed]
- Kubelka, V.; Altmann, F.; März, L. The asparagine-linked carbohydrate of honeybee venom hyaluronidase. Glycoconj. J. 1995, 12, 77–83. [Google Scholar] [CrossRef]
- Tretter, V.; Altmann, F.; Kubelka, V.; März, L.; Becker, W.M. Fucose α1,3-linked to the core region of glycoprotein n-glycans creates an important epitope for ige from honeybee venom allergic individuals. Int. Arch. Allergy Immunol. 1993, 102, 259–266. [Google Scholar] [CrossRef]
- Hoffman, D.R.; Dove, D.E.; Moffitt, J.E.; Stafford, C.T. Allergens in Hymenoptera venom. XXI. Cross-reactivity and multiple reactivity between fire ant venom and bee and wasp venoms. J. Allergy Clin. Immunol. 1988, 82, 828–834. [Google Scholar] [CrossRef]
- dos Santos-Pinto, J.R.A.; Perez-Riverol, A.; Lasa, A.M.; Palma, M.S. Diversity of peptidic and proteinaceous toxins from social Hymenoptera venoms. Toxicon 2018, 148, 172–196. [Google Scholar] [CrossRef] [Green Version]
- Perez-Riverol, A.; Lasa, A.M.; dos Santos-Pinto, J.R.A.; Palma, M.S. Insect venom phospholipases A1 and A2: Roles in the envenoming process and allergy. Insect Biochem. Mol. Biol. 2019, 105, 10–24. [Google Scholar] [CrossRef] [PubMed]
- Maurer, H.R. Bromelain: Biochemistry, pharmacology and medical use. Cell. Mol. Life Sci. 2001, 58, 1234–1245. [Google Scholar] [CrossRef] [PubMed]
- Nwagu, T.N.; Ugwuodo, C.J. Stabilizing bromelain for therapeutic applications by adsorption immobilization on spores of probiotic Bacillus. Int. J. Biol. Macromol. 2019, 127, 406–414. [Google Scholar] [CrossRef] [PubMed]
- Mertens, M.; Brehler, R. Suitability of different glycoproteins and test systems for detecting cross-reactive carbohydrate determinant-specific IgE in Hymenoptera venom-allergic patients. Int. Arch. Allergy Immunol. 2011, 156, 43–50. [Google Scholar] [CrossRef] [PubMed]
- de Lencastre-Novaes, L.C.; Jozala, A.F.; Lopes, A.M.; de Carvalho-Santos-Ebinuma, V.; Mazzola, P.G.; Pessoa-Junior, A. Stability, purification, and applications of bromelain: A review. Biotechnol. Prog. 2016, 32, 5–13. [Google Scholar] [CrossRef]
- Müller, U.R.; Johansen, N.; Petersen, A.B.; Fromberg-Nielsen, J.; Haeberli, G. Hymenoptera venom allergy: Analysis of double positivity to honey bee and Vespula venom by estimation of IgE antibodies to species-specific major allergens Api m1 and Ves v5. Allergy Eur. J. Allergy Clin. Immunol. 2009, 64, 543–548. [Google Scholar] [CrossRef]
- Aalberse, R.C.; Koshte, V.; Clemens, J.G.J. Immunoglobulin E antibodies that cross-react with vegetable foods, pollen, and Hymenoptera venom. J. Allergy Clin. Immunol. 1981, 68, 356–364. [Google Scholar] [CrossRef]
- Hemmer, W.; Focke, M.; Kolarich, D.; Wilson, I.B.H.; Altmann, F.; Wöhrl, S.; Gotz, M.; Jarisch, R. Antibody binding to venom carbohydrates is a frequent cause for double positivity to honeybee and yellow jacket venom in patients with stinging-insect allergy. J. Allergy Clin. Immunol. 2001, 108, 1045–1052. [Google Scholar] [CrossRef]
- Perez-Riverol, A.; Campos-Pereira, F.D.; Musacchio-Lasa, A.; Romani-Fernandes, L.G.; Santos-Pinto, J.R.A.; Justo-Jacomini, D.L.; Oliveira de Azevedo, G.; Bazon, M.L.; Palma, M.S.; Zollner, R.L.; et al. Molecular cloning, expression and IgE-immunoreactivity of phospholipase A1, a major allergen from Polybia paulista (Hymenoptera: Vespidae) venom. Toxicon 2016, 124, 44–52. [Google Scholar] [CrossRef]
- Bazon, M.L.; Perez-Riverol, A.; dos Santos-Pinto, J.R.A.; Fernandes, L.G.R.; Lasa, A.M.; Justo-Jacomini, D.L.; Palma, M.S.; de Lima-Zollner, R.; Brochetto-Braga, M.R. Heterologous expression, purification and immunoreactivity of the antigen 5 from Polybia paulista wasp venom. Toxins 2017, 9, 124. [Google Scholar] [CrossRef] [Green Version]
- Mosbech, H.; Nielsen, N.H.; Dirksen, A.; Launbjerg, J.; Biering, I.; Søborg, M. Comparison between specific IgE measured by RAST, two chemiluminescent assays and skin prick test. Allergol. Immunopathol. 1992, 20, 220–224. [Google Scholar] [PubMed]
- Eržen, R.; Korošec, P.; Šilar, M.; Mušič, E.; Košnik, M. Carbohydrate epitopes as a cause of cross-reactivity in patients allergic to Hymenoptera venom. Wien. Klin. Wochenschr. 2009, 121, 349–352. [Google Scholar] [CrossRef] [PubMed]
- Jappe, U.; Raulf-Heimsoth, M.; Hoffmann, M.; Burow, G.; Hübsch-Müller, C.; Enk, A. In vitro hymenoptera venom allergy diagnosis: Improved by screening for cross-reactive carbohydrate determinants and reciprocal inhibition. Allergy Eur. J. Allergy Clin. Immunol. 2006, 61, 1220–1229. [Google Scholar] [CrossRef]
- Hemmer, W.; Focke, M.; Kolarich, D.; Dalik, I.; Götz, M.; Jarisch, R. Identification by immunoblot of venom glycoproteins displaying immunoglobulin E-binding N-glycans as cross-reactive allergens in honeybee and yellow jacket venom. Clin. Exp. Allergy 2004, 34, 460–469. [Google Scholar] [CrossRef]
- Rodríguez-Pérez, R.; Monsalve, R.I.; Galán, A.; Perez-Piñar, T.; Umpierrez, A.; Lluch-Bernal, M.; Polo, F.; Caballero, M.L. Cross-reactivity between Anisakis spp. and Wasp venom allergens. Int. Arch. Allergy Immunol. 2014, 163, 179–184. [Google Scholar] [CrossRef]
- Perez-Riverol, A.; Miehe, M.; Jabs, F.; Seismman, H.; Romani-Fernandes, L.G.; de Lima-Zollner, R.; Jakob, T.; Brochetto-Braga, M.R.; Spillner, E. Venoms of Neotropical wasps lack cross-reactive carbohydrate determinants enabling reliable protein-based specific IgE determination. J. Allergy Clin. Immunol. 2018, 141, 1917–1919. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Calabozo, B.; Barber, D.; Polo, F. Studies on the carbohydrate moiety of Pla l 1 allergen. Identification of a major N-glycan and significance for the immunoglobulin E-binding activity. Clin. Exp. Allergy 2002, 32, 1628–1634. [Google Scholar] [CrossRef]
- Gonçalves-Paterson-Fox, E.; Russ-Solis, D.; Delazari dos Santos, L.; Aparecido dos Santos-Pinto, J.R.; Ribeiro da Silva-Menegasso, A.; Cardoso-Maciel-Costa-Silva, R.; Sergio-Palma, M.; Correa-Bueno, O.; de Alcântara-Machado, E. A simple, rapid method for the extraction of whole fire ant venom (Insecta: Formicidae: Solenopsis). Toxicon 2013, 65, 5–8. [Google Scholar] [CrossRef] [Green Version]
- Hartree, E.F. Determination of protein: A modification of the Lowry method that gives a linear photometric response. Anal. Biochem. 1972, 193, 265–275. [Google Scholar] [CrossRef]
- Sedmak, J.J.; Grossberg, S.E. A rapid, sensitive, and versatile assay for protein using Coomassie brilliant blue G250. Anal. Biochem. 1977, 79, 544–552. [Google Scholar] [CrossRef]
- Laemmli, U.K. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 1970, 13, 5–6. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Patient | Age (Years) | Sex | Total IgE | RAST Bee | RAST Wasp | RAST Ant |
|---|---|---|---|---|---|---|
| 1 | 28 | m | 76 | 12.5 | 5.4 | 1.6 |
| 2 | 3 | m | 1228 | 10.8 | 1.2 | 15.9 |
| 3 | 9 | m | 1878 | 19.6 | 16.8 | 11.9 |
| 4 | 8 | m | 841 | 1.8 | 1.0 | 24.0 |
| 5 | 7 | m | 433 | 7.4 | 2.1 | 4.1 |
| 6 | 7 | f | 277 | 27.0 | 15.2 | 4.7 |
| 7 | 32 | m | 2979 | 13.9 | 42.2 | 61.6 |
| 8 | 7 | m | 136 | 2.3 | 6.0 | 7.0 |
| 9 | 20 | m | 462 | 4.9 | 2.7 | 6.4 |
| 10 | 26 | m | 485 | 24.0 | 4.8 | 7.6 |
| 11 | 24 | f | 210 | 10.9 | 15.0 | neg |
| 12 | 8 | f | n.d. | 3.3 | neg | 2.2 |
| 13 | 52 | f | n.d. | 24.7 | 1.5 | neg |
| 14 | 33 | m | n.d. | 24.8 | 3.5 | neg |
| 15 | 40 | f | 500 | neg | 12.3 | neg |
| 16 | 10 | m | 113 | 4.7 | neg | neg |
| 17 | 7 | f | n.d. | neg | neg | 7.7 |
| 18 | 8 | m | 1023 | neg | neg | 8,9 |
| 19 | 7 | m | 264 | neg | 1.0 | neg |
| 20 | 6 | f | 13 | neg | neg | 2.7 |
| 21 | 7 | f | n.d. | neg | 1.3 | neg |
| 22 | 38 | m | 35 | 1.1 | neg | neg |
| 23 | 55 | m | n.d. | 18.9 | 11.2 | 24.9 |
| 24 | 55 | m | n.d. | 6.1 | 4.7 | 1.0 |
| 25 | 4 | m | 1589 | >100 | 4.7 | 31.0 |
| 26 | 42 | f | 183 | 3.8 | 1.0 | 5.9 |
| 27 | 8 | m | n.d. | 19,3 | 2,4 | 11.3 |
| 28 | 8 | m | 1459 | 5.7 | 2.1 | 7.9 |
| 29 | 17 | m | 764 | 2.4 | 11.5 | 1.0 |
| 30 | 17 | m | 207 | 5.0 | 10.7 | 10.8 |
| 31 | 63 | f | n.d. | 1.0 | 3.2 | neg |
| 32 | 52 | m | 62 | 5.3 | 6.1 | neg |
| 33 | 4 | m | 116 | 9.1 | neg | 3.2 |
| 34 | 9 | f | n.d. | 9.7 | neg | >100 |
| 35 | 46 | m | n.d. | 2.9 | 4.0 | neg |
| 36 | 12 | m | 653 | 3.8 | neg | 1.4 |
| 37 | 52 | m | n.d. | 2.5 | neg | neg |
| 38 | 51 | m | n.d. | 1.0 | neg | neg |
| 39 | 30 | m | n.d. | 9.2 | neg | neg |
| 40 | 45 | m | 149 | 68.4 | neg | neg |
| 41 | 44 | m | n.d. | 1.7 | neg | neg |
| 42 | 22 | f | 106 | neg | neg | 3.1 |
| 43 | 10 | m | 372 | neg | neg | 24.5 |
| 44 | 7 | m | n.d. | neg | neg | 36.1 |
| 45 | 20 | f | 344 | neg | neg | 44.9 |
| 46 | 5 | m | 424 | neg | neg | 26.9 |
| 47 | 34 | m | 217 | neg | 1.1 | neg |
| 48 | 69 | f | 40 | neg | 1.1 | neg |
| 49 | 57 | m | n.d. | neg | 1.6 | neg |
| 50 | 19 | f | n.d. | 12.8 | neg | neg |
| 51 | 9 | m | 487 | neg | neg | 23.1 |
| Apis mellifera | Polybia paulista | Solenopsis invicta | ||||||
|---|---|---|---|---|---|---|---|---|
| Patient | Hyal. | PLA2 | Api m6 | PLA1 | Ag 5 | PLA1 | Ag 5 | |
| Pre-adsorption | 1 | ++ | +++ | +++ | ++ | ++ | ++ | - |
| Post-adsorption | - | - | - | - | - | + | - | |
| 2 | - | ++ | ++ | ++ | + | + | - | |
| - | + | + | ++ | - | + | - | ||
| 3 | + | ++ | ++ | +++ | + | + | - | |
| - | + | + | +++ | - | + | - | ||
| 4 | - | + | - | + | + | - | - | |
| - | - | - | - | - | - | - | ||
| 5 | + | ++ | ++ | ++ | ++ | +++ | ++ | |
| - | - | - | - | - | - | - | ||
| 6 | ++ | ++ | + | + | + | +++ | ++ | |
| - | + | - | - | - | - | - | ||
| 7 | ++ | ++ | + | + | ++ | +++ | +++ | |
| - | + | - | + | ++ | ++ | + | ||
| 8 | - | ++ | ++ | + | + | ++ | + | |
| - | + | - | - | - | - | - | ||
| 9 | - | ++ | +++ | +++ | ++ | - | + | |
| - | - | - | +++ | - | - | + | ||
| 10 | - | ++ | - | +++ | + | +++ | +++ | |
| - | + | - | ++ | - | ++ | ++ | ||
| 11 | ++ | + | - | ++ | - | ++ | - | |
| + | - | - | - | - | ++ | - | ||
| 12 | ++ | ++ | ++ | ++ | ++ | +++ | + | |
| - | + | + | - | - | ++ | - | ||
| 13 | - | + | + | ++ | - | + | + | |
| - | + | - | - | - | + | + | ||
| 14 | + | ++ | ++ | +++ | + | + | + | |
| + | ++ | - | - | - | + | + | ||
| 15 | - | + | - | ++ | + | ++ | ++ | |
| - | - | - | + | + | + | + | ||
| 16 | ++ | ++ | ++ | + | + | +++ | + | |
| - | - | - | - | - | + | - | ||
| 17 | - | ++ | - | + | + | ++ | ++ | |
| - | - | - | + | + | - | - | ||
| 18 | - | + | ++ | ++ | - | ++ | ++ | |
| - | + | - | + | - | + | - | ||
| 19 | - | + | - | ++ | - | ++ | +++ | |
| - | - | - | - | - | + | + | ||
| 20 | - | + | - | ++ | + | + | ++ | |
| - | + | + | ++ | + | - | - | ||
| 21 | - | + | - | + | ++ | ++ | ++ | |
| - | + | + | + | + | - | - | ||
| 22 | - | - | - | ++ | + | ++ | ++ | |
| - | - | - | + | - | + | + | ||
| Apis mellifera | Polybia paulista | Solenopsis invicta | ||||||
|---|---|---|---|---|---|---|---|---|
| Patient | Hyal. | PLA2 | Api m 6 | PLA1 | Ag 5 | PLA1 | Ag 5 | |
| Pre-adsorption | 23 | - | ++ | + | ++ | - | - | - |
| Post-adsorption | - | + | - | ++ | - | - | - | |
| 24 | + | ++ | + | ++ | ++ | + | + | |
| - | + | + | - | - | + | - | ||
| 25 | + | ++ | ++ | + | + | + | - | |
| - | - | - | - | - | + | - | ||
| 26 | - | + | + | +++ | + | + | + | |
| - | - | - | +++ | - | - | - | ||
| 27 | + | ++ | - | + | + | ++ | ++ | |
| - | + | - | - | - | + | - | ||
| 28 | - | ++ | ++ | + | - | + | +++ | |
| - | + | - | - | - | ++ | +++ | ||
| 29 | - | ++ | ++ | - | - | +++ | + | |
| - | ++ | - | - | - | ++ | + | ||
| 30 | - | + | ++ | ++ | - | + | ++ | |
| - | + | - | + | - | + | ++ | ||
| 31 | - | ++ | ++ | ++ | ++ | + | + | |
| - | + | - | - | - | + | - | ||
| 32 | - | ++ | ++ | ++ | ++ | + | - | |
| - | - | - | + | - | - | - | ||
| 33 | - | ++ | ++ | - | + | +++ | + | |
| - | + | - | + | + | +++ | + | ||
| 34 | - | + | + | + | - | + | + | |
| - | + | - | - | - | + | + | ||
| 35 | - | + | + | +++ | + | + | + | |
| - | + | - | - | - | + | + | ||
| 36 | - | + | + | ++ | + | + | + | |
| - | + | - | - | + | + | + | ||
| 37 | - | ++ | ++ | ++ | ++ | + | + | |
| - | + | - | - | - | + | + | ||
| 38 | - | ++ | +++ | - | - | - | - | |
| - | - | - | - | - | - | - | ||
| 39 | - | ++ | + | ++ | ++ | + | + | |
| - | + | + | - | - | + | + | ||
| 40 | - | ++ | +++ | ++ | ++ | + | + | |
| - | + | + | - | - | + | - | ||
| 41 | - | ++ | - | + | + | ++ | ++ | |
| - | ++ | - | - | - | - | - | ||
| 42 | - | ++ | - | + | + | + | ++ | |
| - | + | - | - | - | - | - | ||
| 43 | - | ++ | ++ | ++ | - | ++ | +++ | |
| - | - | - | - | - | - | - | ||
| 44 | - | ++ | ++ | - | + | ++ | ++ | |
| - | ++ | - | + | + | - | - | ||
| 45 | - | ++ | +++ | - | + | + | + | |
| - | + | - | - | + | - | + | ||
| 46 | - | +++ | +++ | - | - | ++ | ++ | |
| - | + | - | - | - | - | - | ||
| 47 | - | ++ | - | ++ | - | + | ++ | |
| - | + | - | - | - | - | - | ||
| 48 | - | ++ | ++ | - | + | + | + | |
| - | + | - | + | - | + | + | ||
| 49 | - | + | ++ | ++ | ++ | + | + | |
| - | + | - | - | - | + | + | ||
| 50 | - | + | + | ++ | + | + | + | |
| - | + | - | - | + | + | + | ||
| 51 | - | + | + | +++ | + | + | ++ | |
| - | + | - | - | - | + | ++ | ||
| Bromelain (CCD) | |||||
|---|---|---|---|---|---|
| Patient | Relative Intensity on Immunoblotting Reaction | sIgE (RAST) | Patient | Relative Intensity on Immunoblotting Reaction | sIgE (RAST) |
| 1 | +++ | i1, i3, i70 | 27 | - | i1, i3, i70 |
| 2 | + | i1, i3, i70 | 28 | - | i1, i3, i70 |
| 3 | + | i1, i3, i70 | 29 | - | i1, i3, i70 |
| 4 | + | i1, i3, i70 | 30 | - | i1, i3, i70 |
| 5 | +++ | i1, i3, i70 | 31 | - | i1, i3 |
| 6 | ++ | i1, i3, i70 | 32 | - | i1, i3 |
| 7 | + | i1, i3, i70 | 33 | - | i1, i70 |
| 8 | + | i1, i3, i70 | 34 | - | i1, i70 |
| 9 | + | i1, i3, i70 | 35 | - | i1, i3 |
| 10 | + | i1, i3, i70 | 36 | - | i1, i70 |
| 11 | ++ | i1, i3 | 37 | - | i1 |
| 12 | +++ | i1, i70 | 38 | - | i1 |
| 13 | + | i1, i3 | 39 | - | i1 |
| 14 | + | i1, i3 | 40 | - | i1 |
| 15 | + | i3 | 41 | - | i1 |
| 16 | +++ | i1 | 42 | - | i70 |
| 17 | + | i70 | 43 | - | i70 |
| 18 | + | i70 | 44 | - | i70 |
| 19 | + | i3 | 45 | - | i70 |
| 20 | + | i70 | 46 | - | i70 |
| 21 | ++ | i3 | 47 | - | i3 |
| 22 | ++ | i1 | 48 | - | i3 |
| 23 | - | i1, i3, i70 | 49 | - | i3 |
| 24 | - | i1, i3, i70 | 50 | - | i1 |
| 25 | - | i1, i3, i70 | 51 | - | i70 |
| 26 | - | i1, i3, i70 | |||
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Abram, D.M.; Fernandes, L.G.R.; Perez-Riverol, A.; Regina Brochetto-Braga, M.; Zollner, R.d.L. Cross-Reactive Carbohydrate Determinant in Apis mellifera, Solenopsis invicta and Polybia paulista Venoms: Identification of Allergic Sensitization and Cross-Reactivity. Toxins 2020, 12, 649. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins12100649
Abram DM, Fernandes LGR, Perez-Riverol A, Regina Brochetto-Braga M, Zollner RdL. Cross-Reactive Carbohydrate Determinant in Apis mellifera, Solenopsis invicta and Polybia paulista Venoms: Identification of Allergic Sensitization and Cross-Reactivity. Toxins. 2020; 12(10):649. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins12100649
Chicago/Turabian StyleAbram, Débora Moitinho, Luís Gustavo Romani Fernandes, Amilcar Perez-Riverol, Márcia Regina Brochetto-Braga, and Ricardo de Lima Zollner. 2020. "Cross-Reactive Carbohydrate Determinant in Apis mellifera, Solenopsis invicta and Polybia paulista Venoms: Identification of Allergic Sensitization and Cross-Reactivity" Toxins 12, no. 10: 649. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins12100649