Further Advance of Gambierdiscus Species in the Canary Islands, with the First Report of Gambierdiscus belizeanus

 , , ,
, , ,  and
and
Abstract
:1. Introduction
2. Results and Discussion
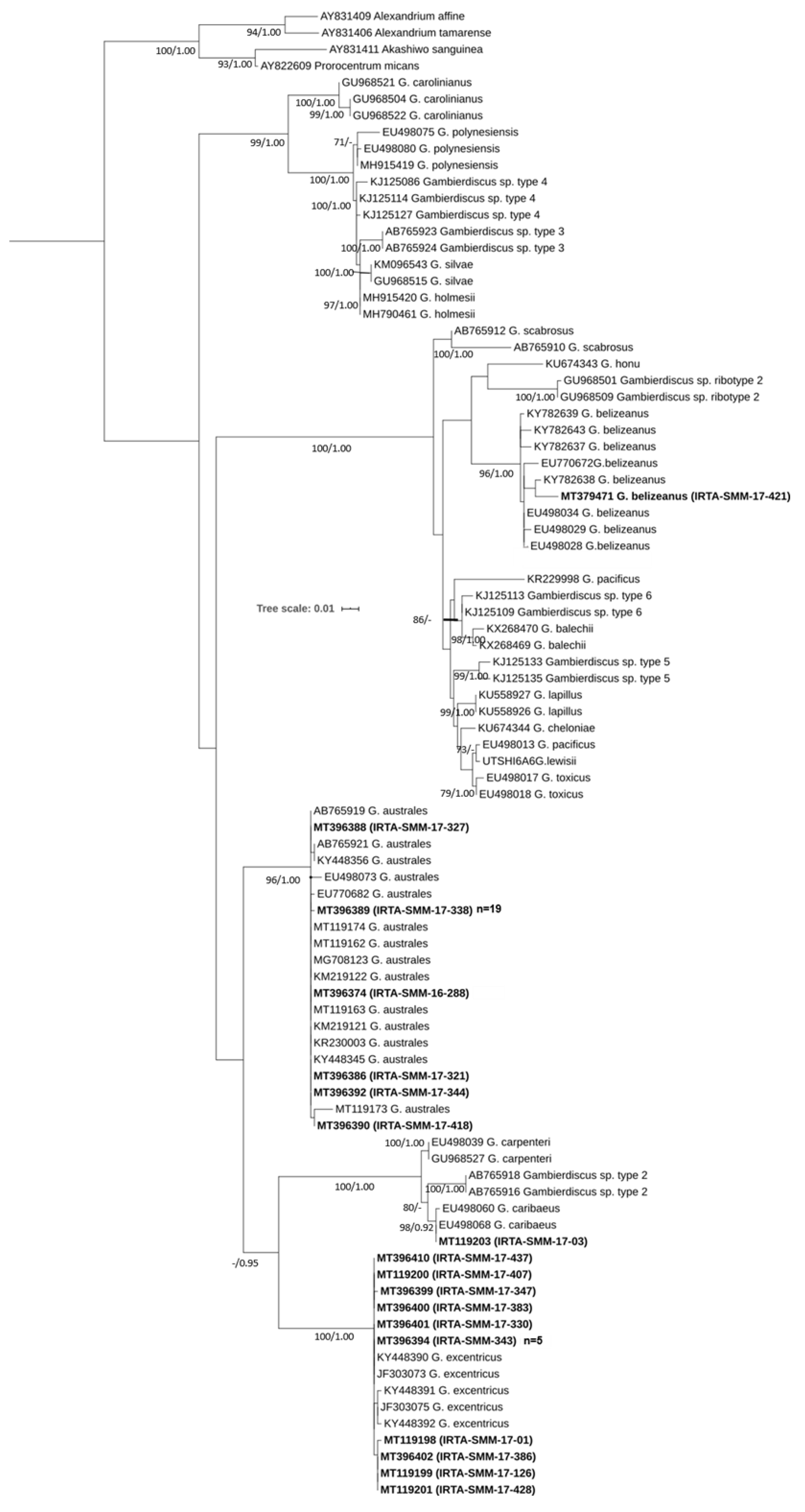
2.1. Molecular Identification
2.2. Morphological Characterization
Morphological Characterization of G. belizeanus
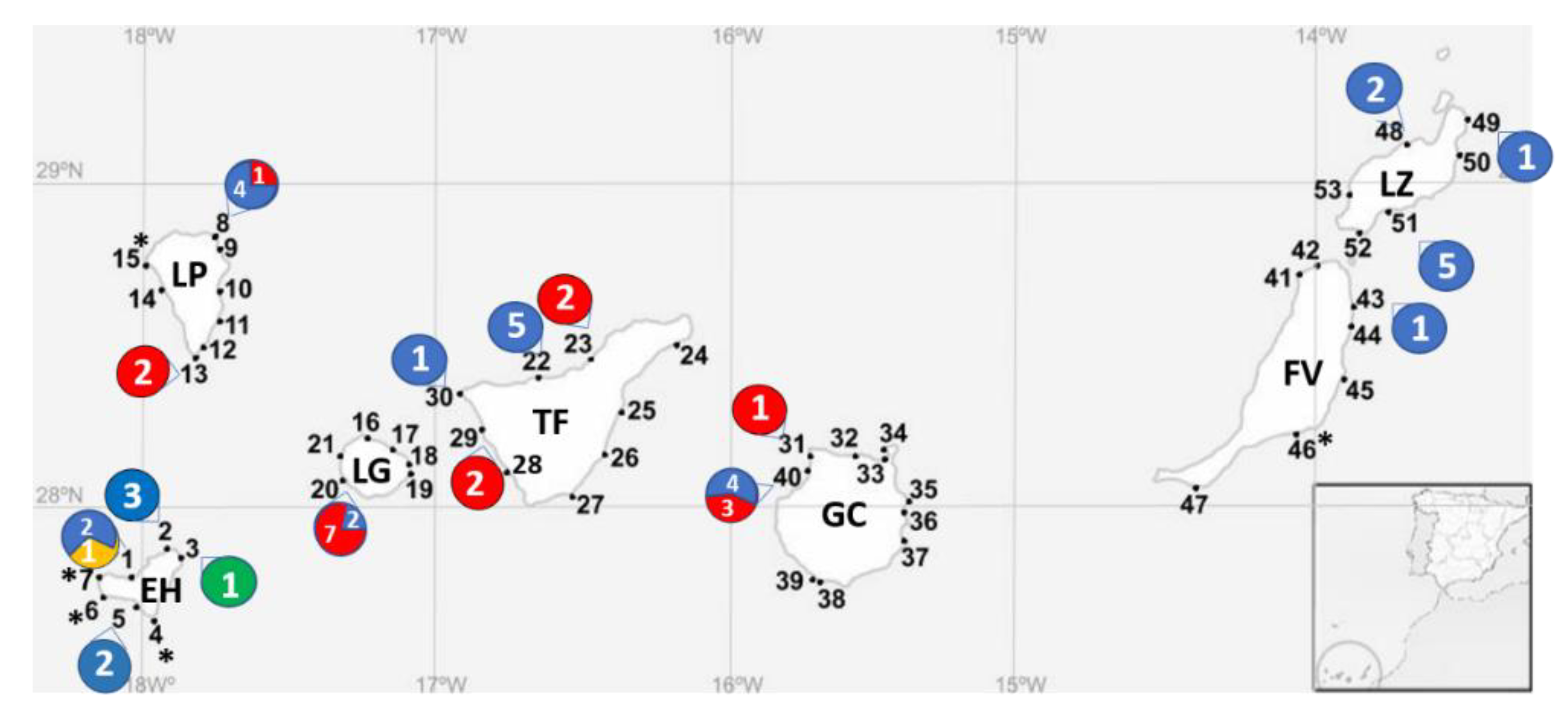
2.3. Distribution of the Gambierdiscus Species in the Canary Islands
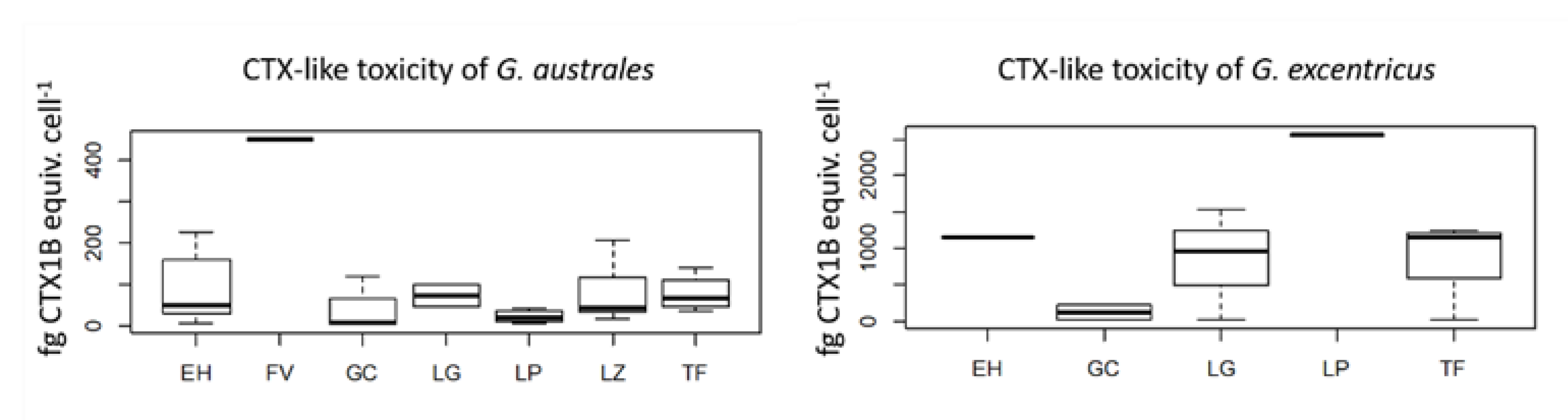
2.4. Evaluation of CTX-Like Toxicity with the Neuro-2a CBA
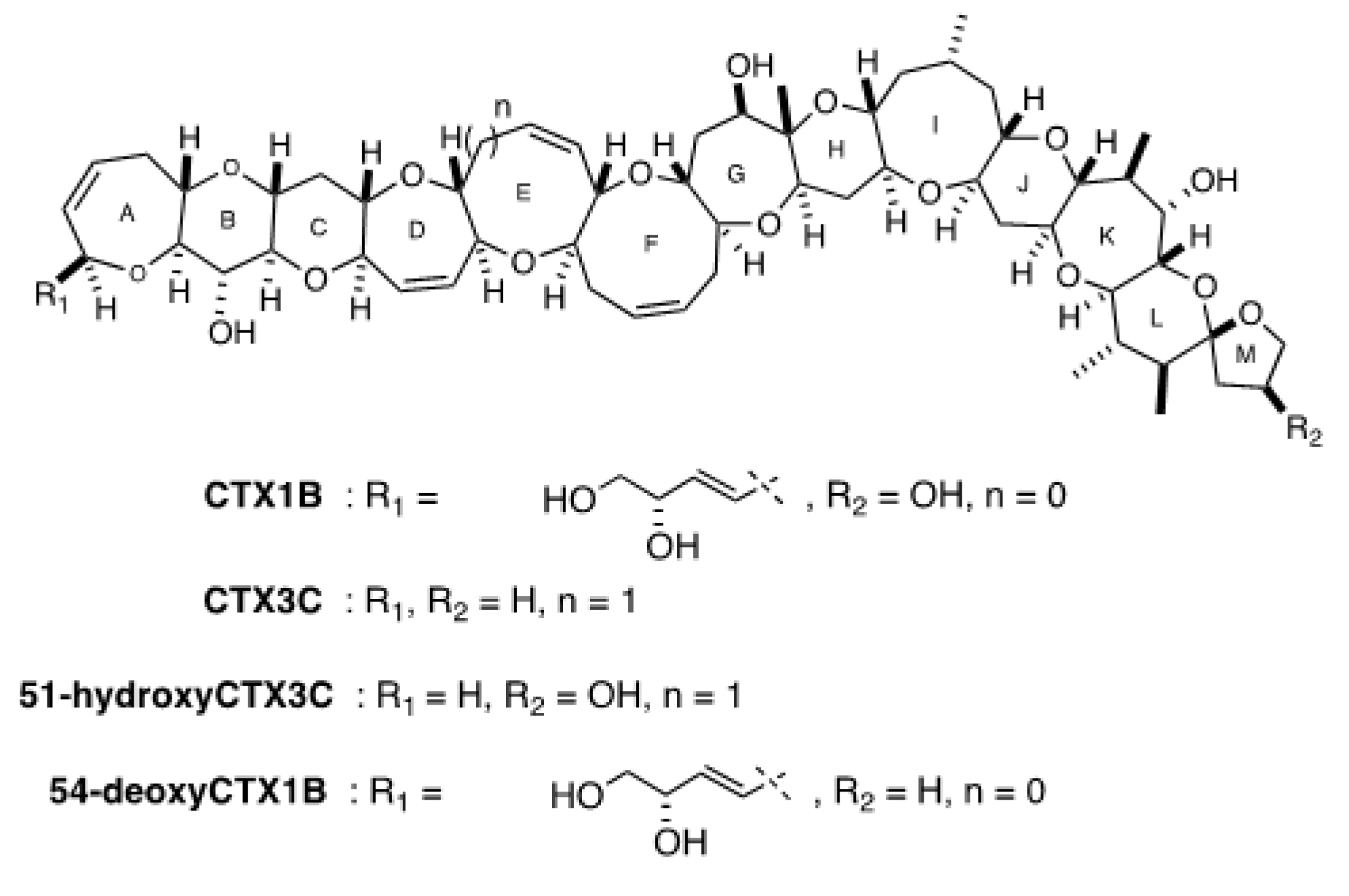
2.5. Evaluation of the Presence of Two Series of CTX Congener Equivalents (CTX1B and CTX3C) in G. belizeanus with a Colorimetric Immunoassay and an Electrochemical Immunosensor
2.6. Future Research Strategies to Understand CP in the Canary Islands
3. Conclusions
4. Methods
4.1. Reagents and Equipment
4.2. Sampling Area and the Strategy
4.3. Isolation and Culturing
4.4. Molecular Identification
4.5. Morphological Characterization
4.6. Production of Microalgal Extracts for Toxin Evaluation
4.7. Evaluation of CTX-Like Toxicity with the Neuroblastoma Cell-Based Assay
4.8. Evaluation of the Presence of Two Series of CTX Congeners (CTX1B and CTX3C) in G. belizeanus with a Colorimetric Immunoassay and an Electrochemical Immunosensor
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Adachi, R.; Fukuyo, Y. The thecal structure of a marine toxic dinoflagellate Gambierdiscus toxicus gen. et spec. nov. collected in a ciguatera-endemic area. Bull. Jpn. Soc. Sci. Fish. 1979, 45, 67–71. [Google Scholar] [CrossRef] [Green Version]
- Murata, M.; Ishibashi, Y.; Fukui, M.; Yasumoto, T.; Legrand, A.M. Structures and Configurations of Ciguatoxin from the Moray Eel Gymnothorax javanicus and Its Likely Precursor from the Dinoflagellate Gambierdiscus toxicus. J. Am. Chem. Soc. 1990, 112, 4380–4386. [Google Scholar] [CrossRef]
- Parsons, M.L.; Settlemier, C.J.; Ballauer, J.M. An examination of the epiphytic nature of Gambierdiscus toxicus, a dinoflagellate involved in ciguatera fish poisoning. Harmful Algae 2011, 10, 598–605. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- FAO; WHO. Report of the Expert Meeting on Ciguatera Poisoning; Food Safety and Quality: Rome, Italy, 2020; ISBN 9789240006294. [Google Scholar]
- Molgó, J.; Shimahara, T.; Legrand, A.M. Ciguatoxin, extracted from poisonous morays eels, causes sodium-dependent calcium mobilization in NG108-15 neuroblastoma × glioma hybrid cells. Neurosci. Lett. 1993, 158, 147–150. [Google Scholar] [CrossRef]
- Nicholson, G.M.; Lewis, R.J. Ciguatoxins: Cyclic polyether modulators of voltage-gated ion channel function. Mar. Drugs 2006, 4, 82–118. [Google Scholar] [CrossRef] [Green Version]
- Gillespie, N.C.; Lewis, R.J.; Pearn, J.H.; Bourke, A.T.; Holmes, M.J.; Bourke, J.B.; Shields, W.J. Ciguatera in Australia. Occurrence, clinical features, pathophysiology and management. Med. J. Aust. 1986, 145, 584–590. [Google Scholar] [CrossRef]
- Friedman, M.A.; Fernandez, M.; Backer, L.C.; Dickey, R.W.; Bernstein, J.; Schrank, K.; Kibler, S.; Stephan, W.; Gribble, M.O.; Bienfang, P.; et al. An updated review of ciguatera fish poisoning: Clinical, epidemiological, environmental, and public health management. Mar. Drugs 2017, 15, 72. [Google Scholar] [CrossRef]
- Skinner, M.P.; Brewer, T.D.; Johnstone, R.; Fleming, L.E.; Lewis, R.J. Ciguatera fish poisoning in the pacific islands (1998 to 2008). PLoS Negl. Trop. Dis. 2011, 5, 1–7. [Google Scholar] [CrossRef]
- Llewellyn, L.E. Revisiting the association between sea surface temperature and the epidemiology of fish poisoning in the South Pacific: Reassessing the link between ciguatera and climate change. Toxicon 2010, 56, 691–697. [Google Scholar] [CrossRef]
- Tester, P.A.; Feldman, R.L.; Nau, A.W.; Kibler, S.R.; Wayne Litaker, R. Ciguatera fish poisoning and sea surface temperatures in the Caribbean Sea and the West Indies. Toxicon 2010, 56, 698–710. [Google Scholar] [CrossRef]
- Farrell, H.; Edwards, A.; Zammit, A. Four recent ciguatera fish poisoning incidents in New South Wales, Australia linked to imported fish. Commun. Dis. Intell. 2019, 43. [Google Scholar] [CrossRef]
- Bravo, J.; Suarez, F.C.; Ramirez, A.S.; Acosta, F. Ciguatera, an Emerging Human Poisoning in Europe. J. Aquac. Mar. Biol. 2015, 3, 1–6. [Google Scholar]
- Chinain, M.; Gatti, C.M.; Roué, M.; Darius, H.T. Ciguatera poisoning in French Polynesia: Insights into the novel trends of an ancient disease. New Microbes New Infect. 2019, 31, 100565. [Google Scholar] [CrossRef] [PubMed]
- Nishimura, T.; Sato, S.; Tawong, W.; Sakanari, H.; Uehara, K.; Shah, M.M.R.; Suda, S.; Yasumoto, T.; Taira, Y.; Yamaguchi, H.; et al. Genetic Diversity and Distribution of the Ciguatera-Causing Dinoflagellate Gambierdiscus spp. (Dinophyceae) in Coastal Areas of Japan. PLoS ONE 2013, 8, e60882. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tester, P.A.; Vandersea, M.W.; Buckel, C.A.; Kibler, S.R.; Holland, W.C.; Davenport, E.D.; Clark, R.D.; Edwards, K.F.; Taylor, J.C.; Pluym, J.L.V.; et al. Gambierdiscus (Dinophyceae) species diversity in the flower garden banks national marine sanctuary, Northern Gulf of Mexico, USA. Harmful Algae 2013, 29, 1–9. [Google Scholar] [CrossRef]
- Pérez-Arellano, J.L.; Luzardo, O.P.; Brito, A.P.; Cabrera, M.H.; Zumbado, M.; Carranza, C.; Angel-Moreno, A.; Dickey, R.W.; Boada, L.D. Ciguatera fish poisoning, Canary Islands. Emerg. Infect. Dis. 2005, 11, 1981–1982. [Google Scholar] [CrossRef] [Green Version]
- Gouveia, N.N.; Vale, P.; Gouveia, N.; Delgado, J. Primeiro Registo da Ocorrência de Episódios do Tipo Ciguatérico no Arquipélago da Madeira. In Algas Toxicas e Biotoxinas nas Águas da Península Ibérica; IPIMAR: Lisboa, Portugal, 2010; pp. 152–157. [Google Scholar]
- Falcón García, I. Epidemiología de la Intoxicación Alimentaria por Ciguatoxinas en Canarias; Hospital Universitario Nuestra Señora de Candelaria: Santa Cruz de Tenerife, Spain, 2018. [Google Scholar]
- Caillaud, A.; Eixarch, H.; de la Iglesia, P.; Rodriguez, M.; Dominguez, L.; Andree, K.B.; Diogène, J. Towards the standardisation of the neuroblastoma (neuro-2a) cell-based assay for ciguatoxin-like toxicity detection in fish: Application to fish caught in the Canary Islands. Food Addit. Contam. Part A Chem. Anal. Control. Expo. Risk Assess. 2012, 29, 1000–1010. [Google Scholar] [CrossRef]
- Sanchez-Henao, J.A.; García-Álvarez, N.; Fernández, A.; Saavedra, P.; Silva Sergent, F.; Padilla, D.; Acosta-Hernández, B.; Martel Suárez, M.; Diogène, J.; Real, F. Predictive score and probability of CTX-like toxicity in fish samples from the official control of ciguatera in the Canary Islands. Sci. Total Environ. 2019, 673, 576–584. [Google Scholar] [CrossRef]
- Munday, R.; Murray, S.; Rhodes, L.; Larsson, M.; Harwood, D. Ciguatoxins and Maitotoxins in Extracts of Sixteen Gambierdiscus Isolates and One Fukuyoa Isolate from the South Pacific and Their Toxicity to Mice by Intraperitoneal and Oral Administration. Mar. Drugs 2017, 15, 208. [Google Scholar] [CrossRef] [Green Version]
- Rhodes, L.; Harwood, T.; Smith, K.; Argyle, P.; Munday, R. Production of ciguatoxin and maitotoxin by strains of Gambierdiscus australes, G. pacificus and G. polynesiensis (Dinophyceae) isolated from Rarotonga, Cook Islands. Harmful Algae 2014, 39, 185–190. [Google Scholar] [CrossRef]
- Pisapia, F.; Holland, W.C.; Hardison, D.R.; Litaker, R.W.; Fraga, S.; Nishimura, T.; Adachi, M.; Nguyen-Ngoc, L.; Séchet, V.; Amzil, Z.; et al. Toxicity screening of 13 Gambierdiscus strains using neuro-2a and erythrocyte lysis bioassays. Harmful Algae 2017, 63, 173–183. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rossignoli, A.E.; Tudó, A.; Bravo, I.; Díaz, P.A.; Diogène, J.; Riobó, P. Toxicity characterisation of Gambierdiscus species from the canary Islands. Toxins (Basel) 2020, 12, 134. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tester, P.A.; Litaker, R.W.; Berdalet, E. Climate change and harmful benthic microalgae. Harmful Algae 2020, 91, 101655. [Google Scholar] [CrossRef]
- Navarro-Pérez, E.; Barton, E.D. Seasonal and interannual variability of the Canary Current. Sci. Mar. 2001, 65, 205–213. [Google Scholar] [CrossRef]
- Davenport, R.; Neuer, S.; Helmke, P.; Perez-Marrero, J.; Llinas, O. Primary productivity in the northern Canary Islands region as inferred from SeaWiFS imagery. Deep. Res. Part II Top. Stud. Oceanogr. 2002, 49, 3481–3496. [Google Scholar] [CrossRef]
- Fraga, S.; Riobó, P.; Diogène, J.; Paz, B.; Franco, J.M. Toxic and potenially toxic benthic dinoflagellates observed in Macaronesia (NE Atlantic Archipelago). In Proceedings of the Abstract Book of the 11th International Conference on Harmful Algae, Capetown, South Africa, 15–19 November 2004; p. 115. [Google Scholar]
- Fraga, S.; Rodríguez, F.; Caillaud, A.; Diogène, J.; Raho, N.; Zapata, M. Gambierdiscus excentricus sp. nov. (Dinophyceae), a benthic toxic dinoflagellate from the Canary Islands (NE Atlantic Ocean). Harmful Algae 2011, 11, 10–22. [Google Scholar] [CrossRef] [Green Version]
- Fraga, S.; Rodríguez, F. Genus Gambierdiscus in the Canary Islands (NE Atlantic Ocean) with Description of Gambierdiscus silvae sp. nov., a New Potentially Toxic Epiphytic Benthic Dinoflagellate. Protist 2014, 165, 839–853. [Google Scholar] [CrossRef]
- Bravo, I.; Rodriguez, F.; Ramilo, I.; Rial, P.; Fraga, S. Ciguatera-causing dinoflagellate Gambierdiscus spp. (Dinophyceae) in a subtropical region of North Atlantic Ocean (Canary Islands): Morphological characterization and biogeography. Toxins (Basel) 2019, 11, 423. [Google Scholar] [CrossRef] [Green Version]
- Rodríguez, F.; Fraga, S.; Ramilo, I.; Rial, P.; Figueroa, R.I.; Riobó, P.; Bravo, I. “Canary Islands (NE Atlantic) as a biodiversity ‘hotspot’ of Gambierdiscus: Implications for future trends of ciguatera in the area”. Harmful Algae 2017, 67, 131–143. [Google Scholar]
- Faust, M.A. Observation of sand-dwelling toxic dinoflagellates (Dinophyceae) from widely differing sites, including two new species. J. Phycol. 1995, 31, 996–1003. [Google Scholar] [CrossRef]
- Díaz-Asencio, L.; Clausing, R.J.; Vandersea, M.; Chamero-Lago, D.; Gómez-Batista, M.; Hernández-Albernas, J.I.; Chomérat, N.; Rojas-Abrahantes, G.; Litaker, R.W.; Tester, P.; et al. Ciguatoxin occurrence in food-web components of a cuban coral reef ecosystem: Risk-assessment implications. Toxins (Basel) 2019, 11, 722. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Richlen, M.L.; Morton, S.L.; Barber, P.H.; Lobel, P.S. Phylogeography, morphological variation and taxonomy of the toxic dinoflagellate Gambierdiscus toxicus (Dinophyceae). Harmful Algae 2008, 7, 614–629. [Google Scholar] [CrossRef]
- Litaker, R.W.; Holland, W.C.; Hardison, D.R.; Pisapia, F.; Hess, P.; Kibler, S.R.; Tester, P.A. Ciguatoxicity of Gambierdiscus and Fukuyoa species from the Caribbean and Gulf of Mexico. PLoS ONE 2017, 12, 1–19. [Google Scholar] [CrossRef]
- Catania, D.; Richlen, M.L.; Mak, Y.L.; Morton, S.L.; Laban, E.H.; Xu, Y.; Anderson, D.M.; Chan, L.L.; Berumen, M.L. The prevalence of benthic dinoflagellates associated with ciguatera fish poisoning in the central Red Sea. Harmful Algae 2017, 68, 206–216. [Google Scholar] [CrossRef] [PubMed]
- Murray, S.; Momigliano, P.; Heimann, K.; Blair, D. Molecular phylogenetics and morphology of Gambierdiscus yasumotoi from tropical eastern Australia. Harmful Algae 2014, 39, 242–252. [Google Scholar] [CrossRef]
- Leaw, C.P.; Lim, P.T.; Tan, T.H.; Tuan-Halim, T.N.; Cheng, K.W.; Ng, B.K.; Usup, G. First report of the benthic dinoflagellate, Gambierdiscus belizeanus (Gonyaulacales: Dinophyceae) for the east coast of Sabah, Malaysian Borneo. Phycol. Res. 2011, 59, 143–146. [Google Scholar] [CrossRef]
- Xu, Y.; Richlen, M.L.; Morton, S.L.; Mak, Y.L.; Chan, L.L.; Tekiau, A.; Anderson, D.M. Distribution, abundance and diversity of Gambierdiscus spp. from a ciguatera-endemic area in Marakei, Republic of Kiribati. Harmful Algae 2014, 34, 56–68. [Google Scholar] [CrossRef] [Green Version]
- Bravo, I.; Figueroa, R.I.; Fraga, S. Cellular and nuclear morphological variability within a single species of the toxigenic dinoflagellate genus Gambierdiscus: Relationship to life-cycle processes. Harmful Algae 2014, 40, 1–8. [Google Scholar] [CrossRef]
- Chinain, M.; Faust, M.A.; Pauillac, S. Morphology and molecular analyses of three toxic species of Gambierdiscus (Dinophyceae): G. pacificus, sp nov., G. australes, sp nov., and G. polynesiensis, sp nov. J. Phycol. 1999, 35, 1282–1296. [Google Scholar] [CrossRef]
- Litaker, R.W.; Vandersea, M.W.; Faust, M.A.; Kibler, S.R.; Chinain, M.; Holmes, M.J.; Holland, W.C.; Tester, P.A. Taxonomy of Gambierdiscus including four new species, Gambierdiscus caribaeus, Gambierdiscus carolinianus, Gambierdiscus carpenteri and Gambierdiscus ruetzleri (Gonyaulacales, Dinophyceae). Phycologia 2009, 48, 344–390. [Google Scholar] [CrossRef]
- Hoppenrath, M.; Kretzschmar, A.L.; Kaufmann, M.J.; Murray, S.A. Morphological and molecular phylogenetic identification and record verification of Gambierdiscus excentricus (Dinophyceae) from Madeira Island (NE Atlantic Ocean). Mar. Biodivers. Rec. 2019, 12, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Aligizaki, K.; Nikolaidis, G.; Fraga, S. Is Gambierdiscus expanding to new areas? Harmful Algae News 2008, 36, 6–7. [Google Scholar]
- Kibler, S.R.; Litaker, R.W.; Holland, W.C.; Vandersea, M.W.; Tester, P.A. Growth of eight Gambierdiscus (Dinophyceae) species: Effects of temperature, salinity and irradiance. Harmful Algae 2012, 19, 1–14. [Google Scholar] [CrossRef]
- Xu, Y.; Richlen, M.L.; Liefer, J.D.; Robertson, A.; Kulis, D.; Smith, T.B.; Parsons, M.L.; Anderson, D.M. Influence of Environmental Variables on Gambierdiscus spp. (Dinophyceae) Growth and Distribution. PLoS ONE 2016, 11, e0153197. [Google Scholar] [CrossRef] [PubMed]
- Espino, F.; Tuya, F.; Rosario, A.; Bosch, N.E.; Coca, J.; Gonz, A.J.; Rosario, F.; Otero-ferrer, F.J.; Moreno, Á.C.; Haroun, R. Geographical Range Extension of the Spotfin burrfish, Chilomycterus reticulatus (L. 1758), in the Canary Islands: A Response to Ocean Warming? Diversity 2019, 11, 230. [Google Scholar] [CrossRef] [Green Version]
- PLOCAN Oceanic Platform of the Canary Islands (PLOCAN). Available online: http://obsplatforms.plocan.eu/climatology (accessed on 3 June 2020).
- Yoshimatsu, T.; Yamaguchi, H.; Iwamoto, H.; Nishimura, T.; Adachi, M. Effects of temperature, salinity and their interaction on growth of Japanese Gambierdiscus spp. (Dinophyceae). Harmful Algae 2014, 35, 29–37. [Google Scholar] [CrossRef]
- Litaker, R.W.; Tester, P.A.; Vandersea, M.W. Species-specific PCR assays for Gambierdiscus excentricus and Gambierdiscus silvae (Gonyaulacales, Dinophyceae). J. Phycol. 2019, 55, 730–732. [Google Scholar] [CrossRef]
- Tawong, W.; Yoshimatsu, T.; Yamaguchi, H.; Adachi, M. Temperature and salinity effects and toxicity of <i>Gambierdiscus caribaeus (Dinophyceae) from Thailand. Phycologia 2016, 55, 274–278. [Google Scholar] [CrossRef]
- Tuya, F.; Haroun, R.J. Spatial patterns and response to wave exposure of shallow water algal assemblages across the Canarian Archipelago: A multi-scaled approach. Mar. Ecol. Prog. Ser. 2006, 311, 15–28. [Google Scholar] [CrossRef]
- Soler-Onis, E.; Fernández-Zabala, J.; Ojeda-Rodriguez, A.; Amorim, A. Bloom of Gambierdiscus caribaeus in the temperate-subtropical waters of El Hierro, Canary Islands (North East Atlantic). Harmful Algae News 2016, 55, 14–16. [Google Scholar]
- Vélez-Belchí, P.; Gonzalez-Carballo, M.; Perez-Hernández, M.D.; Hernández-Guerra, A. Open ocean temperature and salinity trends. Oceanogr. Biol. Featur. Canar. Curr. Large Mar. Ecosyst. 2015, 13, 299–308. [Google Scholar]
- Kibler, S.R.; Tester, P.A.; Kunkel, K.E.; Moore, S.K.; Litaker, R.W. Effects of ocean warming on growth and distribution of dinoflagellates associated with ciguatera fish poisoning in the Caribbean. Ecol. Modell. 2015, 316, 194–210. [Google Scholar] [CrossRef] [Green Version]
- Ikehara, T.; Kuniyoshi, K.; Oshiro, N.; Yasumoto, T. Biooxidation of ciguatoxins leads to species-specific toxin profiles. Toxins (Basel) 2017, 9, 205. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yogi, K.; Sakugawa, S.; Oshiro, N.; Ikehara, T.; Sugiyama, K.; Yasumoto, T. Determination of toxins involved in ciguatera fish poisoning in the Pacific by LC/MS. J. AOAC Int. 2014, 97, 398–402. [Google Scholar] [CrossRef] [PubMed]
- Lewis, R.J.; Sellin, M.; Poli, M.A.; Norton, R.S.; MacLeod, J.K.; Sheil, M.M. Purification and characterization of ciguatoxins from moray eel (Lycodontis javanicus, Muraenidae). Toxicon 1991, 29, 1115–1127. [Google Scholar] [CrossRef]
- Reverté, L.; Toldrà, A.; Andree, K.B.; Fraga, S.; de Falco, G.; Campàs, M.; Diogène, J. Assessment of cytotoxicity in ten strains of Gambierdiscus australes from Macaronesian Islands by neuro-2a cell-based assays. J. Appl. Phycol. 2018, 30, 2447–2461. [Google Scholar] [CrossRef]
- Paz, B.; Riobó, P.; Franco, J.M. Preliminary study for rapid determination of phycotoxins in microalgae whole cells using matrix-assisted laser desorption/ionization time-of-flight mass spectrometry. Rapid Commun. Mass Spectrom. 2011, 25, 3627–3639. [Google Scholar] [CrossRef] [Green Version]
- Pisapia, F.; Sibat, M.L.; Herrenknecht, C.; Lhaute, K.; Gaiani, G.; Ferron, P.J.; Fessard, V.; Fraga, S.; Nascimento, S.M.; Litaker, R.W.; et al. Maitotoxin-4, a novel MTX analog produced by Gambierdiscus excentricus. Mar. Drugs 2017, 15, 220. [Google Scholar] [CrossRef]
- Estevez, P.; Sibat, M.; Leao, J.M.; Tudó, A.; Rambla-Alegre, M.; Aligizak, K.; Gago-Martinez, A.; Diogène, J.; Hess, P. Use of Mass Spectrometry to determine the Diversity of Toxins Produced by Gambierdiscus and Fukuyoa Species from Balearic Islands and Crete (Mediterranean Sea) and the Canary Islands (Northeast Atlantic). Toxins (Basel) 2020, 12, 305. [Google Scholar] [CrossRef]
- Caillaud, A.; Yasumoto, T.; Diogène, J. Detection and quantification of maitotoxin-like compounds using a neuroblastoma (Neuro-2a) cell based assay. Application to the screening of maitotoxin-like compounds in Gambierdiscus spp. Toxicon 2010, 56, 36–44. [Google Scholar] [CrossRef]
- Lewis, R.J.; Inserra, M.; Vetter, I.; Holland, W.C.; Hardison, D.R.; Tester, P.A.; Litaker, R.W. Rapid extraction and identification of maitotoxin and ciguatoxin-like toxins from Caribbean and pacific Gambierdiscus using a new functional bioassay. PLoS ONE 2016, 11, e0160006. [Google Scholar] [CrossRef] [PubMed]
- De Haro, L.; Pommier, P.; Valli, M. Emergence of Imported Ciguatera in Europe: Report of 18 Cases at the Poison Control Centre of Marseille. J. Toxicol. Clin. Toxicol. 2003, 41, 927–930. [Google Scholar] [CrossRef] [PubMed]
- Gaiani, G.; Leonardo, S.; Tudó, À.; Toldrà, A.; Rey, M.; Andree, K.B.; Tsumuraya, T.; Hirama, M.; Diogène, J.; O’Sullivan, C.K.; et al. Rapid extraction of ciguatoxins from Gambierdiscus and Fukuyoa and detection with immunosensing tools. Ecotoxicol. Environ. Saf. 2020, 204, 111004. [Google Scholar] [CrossRef]
- Sanchez-Henao, A.; García-álvarez, N.; Silva, F.; Estévez, P.; Gago-martínez, A.; Martín, F.; Ramos-sosa, M.; Fernández, A.; Diogène, J.; Real, F. Presence of CTXs in moray eels and dusky groupers in the marine environment of the Canary Islands. Aquat. Toxicol. 2020, 221, 105427. [Google Scholar] [CrossRef] [PubMed]
- Tibiriçá, C.E.J.A.; Sibat, M.; Fernandes, L.F.; Bilien, G.; Chomérat, N.; Hess, P.; Mafra, L.L., Jr. Diversity and Toxicity of the Genus Coolia Meunier in Brazil, and Detection of 44-methyl Gambierone in Coolia tropicalis. Toxins 2020, 12, 327. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez, I.; Genta-Jouve, G.; Alfonso, C.; Calabro, K.; Alonso, E.; Sánchez, J.A.; Alfonso, A.; Thomas, O.P.; Botana, L.M. Gambierone, a Ladder-Shaped Polyether from the Dinoflagellate Gambierdiscus belizeanus. Org. Lett. 2015, 17, 2392–2395. [Google Scholar] [CrossRef]
- Boente-Juncal, A.; Álvarez, M.; Antelo, Á.; Rodríguez, I.; Calabro, K.; Vale, C.; Thomas, O.P.; Botana, L.M. Structure elucidation and biological evaluation of maitotoxin-3, a homologue of gambierone, from Gambierdiscus belizeanus. Toxins (Basel) 2019, 11, 79. [Google Scholar] [CrossRef] [Green Version]
- Murray, J.S.; Nishimura, T.; Finch, S.C.; Rhodes, L.L.; Puddick, J.; Harwood, D.T.; Larsson, M.E.; Doblin, M.A.; Leung, P.; Yan, M.; et al. The role of 44-methylgambierone in ciguatera fish poisoning: Acute toxicity, production by marine microalgae and its potential as a biomarker for Gambierdiscus spp. Harmful Algae 2020, 97, 101853. [Google Scholar] [CrossRef]
- Reverté, L.; Soliño, L.; Carnicer, O.; Diogène, J.; Campàs, M. Alternative methods for the detection of emerging marine toxins: Biosensors, biochemical assays and cell-based assays. Mar. Drugs 2014, 12, 5719–5963. [Google Scholar] [CrossRef] [Green Version]
- Caillaud, A.; De La Iglesia, P.; Darius, H.T.; Pauillac, S.; Aligizaki, K.; Fraga, S.; Chinain, M.; Diogène, J. Update on methodologies available for ciguatoxin determination: Perspectives to confront the onset of ciguatera fish poisoning in Europe. Mar. Drugs 2010, 8, 1838–1907. [Google Scholar] [CrossRef]
- Kretzschmar, A.L.; Verma, A.; Kohli, G.; Murray, S. Development of a quantitative PCR assay for the detection and enumeration of a potentially ciguatoxin-producing dinoflagellate, Gambierdiscus lapillus (Gonyaulacales, Dinophyceae). PLoS ONE 2019, 14, 1–18. [Google Scholar] [CrossRef] [Green Version]
- Smith, K.F.; Biessy, L.; Argyle, P.A.; Trnski, T.; Halafihi, T.; Rhodes, L.L. Molecular identification of Gambierdiscus and Fukuyoa (Dinophyceae) from environmental samples. Mar. Drugs 2017, 15, 243. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fernández-Zabala, J.; Tuya, F.; Amorim, A.; Soler-Onís, E. Benthic dinoflagellates: Testing the reliability of the artificial substrate method in the Macaronesian region. Harmful Algae 2019, 87, 101634. [Google Scholar] [CrossRef] [PubMed]
- Nagumo, Y.; Oguri, H.; Tsumoto, K.; Shindo, Y.; Hirama, M.; Tsumuraya, T.; Fujii, I.; Tomioka, Y.; Mizugaki, M.; Kumagai, I. Phage-display selection of antibodies to the left end of CTX3C using synthetic fragments. J. Immunol. Methods 2004, 289, 137–146. [Google Scholar] [CrossRef]
- Nagumo, Y.; Oguri, H.; Shindo, Y.; Sasaki, S.-Y.; Oishi, T.; Hirama, M.; Tomioka, Y.; Mizugaki, M.; Tsumuraya, T. Concise synthesis of ciguatoxin ABC-ring fragments and surface plasmon resonance study of the interaction of their BSA conjugates with monoclonal antibodies. Bioorg. Med. Chem. Lett. 2001, 11, 2037–2040. [Google Scholar] [CrossRef]
- Oguri, H.; Hirama, M.; Tsumuraya, T.; Fujii, I.; Maruyama, M.; Uehara, H.; Nagumo, Y. Synthesis-based approach toward direct sandwich immunoassay for ciguatoxin CTX3C. J. Am. Chem. Soc. 2003, 125, 7608–7612. [Google Scholar] [CrossRef]
- Oguri, H.; Tanaka, S.-I.; Hishiyama, S.; Oishi, T.; Hirama, M.; Tumuraya, T.; Tomioka, Y.; Mizugaki, M. Designed hapten aimed at anti-ciguatoxin monoclonal antibody: Synthesis, immunization and discrimination of the C2 configuration. Syntesis 1999, S1, 1431–1436. [Google Scholar] [CrossRef]
- Tsumuraya, T.; Takeuchi, K.; Yamashita, S.; Fujii, I.; Hirama, M. Development of a monoclonal antibody against the left wing of ciguatoxin CTX1B: Thiol strategy and detection using a sandwich ELISA. Toxicon 2012, 60, 348–357. [Google Scholar] [CrossRef]
- Tsumuraya, T.; Fujii, I.; Hirama, M. Production of monoclonal antibodies for sandwich immunoassay detection of Pacific ciguatoxins. Toxicon 2010, 56, 797–803. [Google Scholar] [CrossRef]
- Tsumuraya, T.; Fujii, I.; Inoue, M.; Tatami, A.; Miyazaki, K.; Hirama, M. Production of monoclonal antibodies for sandwich immunoassay detection of ciguatoxin 51-hydroxyCTX3C. Toxicon 2006, 48, 287–294. [Google Scholar] [CrossRef]
- Hoshaw, R.W.; Rosowski, J.R. Handbook of Phycological Methods: Culture Methods and Growth Measurements; Stein, J.R., Ed.; Cambridge University Press: London, UK; New York, NY, USA, 1973; p. 448. [Google Scholar]
- Provasoli, L. Media and prospects of the cultivation of marine algae. In Culure and Collection of Algae Proceedings; Japanese Society of Plant Physiology: Hakone, Japan, 1968; pp. 63–75. [Google Scholar]
- Toldrà, A.; Andree, K.B.; Fernández-Tejedor, M.; Diogène, J.; Campàs, M. Dual quantitative PCR assay for identification and enumeration of Karlodinium veneficum and Karlodinium armiger combined with a simple and rapid DNA extraction method. J. Appl. Phycol. 2018, 30, 2435–2445. [Google Scholar] [CrossRef]
- Rozewicki, J.; Li, S.; Amada, K.M.; Standley, D.M.; Katoh, K. MAFFT-DASH: Integrated protein sequence and structural alignment. Nucleic Acids Res. 2019, 47, W5–W10. [Google Scholar] [CrossRef]
- Stamatakis, A. RAxML version 8: A tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics 2014, 30, 1312–1313. [Google Scholar] [CrossRef] [PubMed]
- Huelsenbeck, J.P.; Ronquist, F. MRBAYES: Bayesian inference of phylogenetic trees. Bioinformatics 2001, 17, 754–755. [Google Scholar] [CrossRef] [Green Version]
- Darriba, D.; Taboada, G.L.; Doallo, R.; Posada, D. jModelTest 2: More models, new heuristics and high-performance computing Europe PMC Funders Group. Nat. Methods 2012, 9, 772. [Google Scholar] [CrossRef] [Green Version]
- Rambaut, A.; Drummond, A.J.; Xie, D.; Baele, G.; Suchard, M.A. Posterior summarization in Bayesian phylogenetics using Tracer 1.7. Syst. Biol. 2018, 67, 901. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bomber, J.W.; Tindall, D.R.; Miller, D.M. Genetic variability in toxin potencies among seventeen clones of Gambierdiscus toxicus (Dinophyceae). J. Phycol. 1989, 25, 617–625. [Google Scholar] [CrossRef]
- Fritz, L.; Triemer, R.E. A Rapid simple technique utilizing calcofluor white M2R for the visualization of dinofalgellate thecal plates. J. Phycol. 1985, 21, 662–664. [Google Scholar] [CrossRef]
- Catterall, W.A.; Nirenberg, M. Sodium uptake associated with activation of action potential ionophores of cultured neuroblastoma and muscle cells. Proc. Natl. Acad. Sci. USA 1973, 70, 3759–3763. [Google Scholar] [CrossRef] [Green Version]
- Catterall, W.A. Molecular Properties of Voltage-Sensitive Sodium Channels. In New Insights into Cell and Membrane Transport Processes; Springer: Boston, MA, USA, 1986; pp. 3–20. [Google Scholar]
- Manger, R.L.; Leja, L.S.; Lee, S.Y.; Hungerforg, J.M.; Wekell, M.M. Tetrazolium-based cell bioassay for neurotoxins active on voltage-sensitive sodium channels: Semiautomated assay for saxitoxins, brevetoxins, and ciguatoxins. Anal. Biochem. 1993, 214, 190–194. [Google Scholar] [CrossRef]
- Caillaud, A.; Cañete, E.; de la Iglesia, P.; Giménez, G.; Diogène, J. Cell-based assay coupled with chromatographic fractioning: A strategy for marine toxins detection in natural samples. Toxicol. Vitr. 2009, 23, 1591–1596. [Google Scholar] [CrossRef] [PubMed]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2017. [Google Scholar]
- Tsumuraya, T.; Sato, T.; Hirama, M.; Fujii, I. Highly Sensitive and Practical Fluorescent Sandwich ELISA for Ciguatoxins. Anal. Chem. 2018, 90, 7318–7324. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Depth (µm) | Width (µm) |
|---|---|---|
| G. australes | 71.13 ± 7.06 (60.6–98.4) | 65.40 ± 6.54 (53.4–82.1) |
| G. belizeanus | 64.12 ± 5.28 (52.8–76.2) | 59.64 ± 5.95 (46.5–76.0) |
| G. caribaeus | 87.20 ± 11.19 (61.2–116.5) | 86.45 ± 11.44 (63.17–119.7) |
| G. excentricus | 90.25 ± 8.90 (72.2–109.7) | 82.97 ± 9.06 (67.8–106.7) |
| Species | Islands | Date of Sampling (References) |
|---|---|---|
| G. australes | LZ, FV, GC | September–October 2015 [33] October 2016 (this study) |
| EH | September–October 2015 [33] October 2017 (this study) | |
| TF | 2013 [31] September–October 2015 [33] October 2017 (this study) | |
| LG | October 2017 ([33] and this study) | |
| LP | October 2017 (this study) | |
| G. belizeanus | EH | October 2017 (This study) |
| G. caribaeus | EH | September–October 2015 [33] October 2015 [55] October 2017 (this study) |
| LG | October 2017 [32] | |
| G. carolinianus | TF | September–October 2015 [33] |
| G. excentricus | TF | March 2004 [30] 2013 [31] September–October 2015 [33] October 2017 (This study) |
| LZ | September–October 2015 [33] | |
| FV | September–October 2015 [33] | |
| GC | September–October 2015 [33] April 2017, October 2017 (this study) | |
| LP | March 2004 [30] October 2017 ([33] and this study) | |
| LG | March 2004 [30] October 2017 ([33] and this study) | |
| G. silvae | TF | 2013 [31] September–October 2015 [33] |
| GC | Winter 2010 [31] | |
| LG | October 2017 [32] |
| Strain Code | CTX-Like Toxicity | Island | Station | Strain Code | CTX-Like Toxicity | Island | Station |
|---|---|---|---|---|---|---|---|
| G. australes | G. australes | ||||||
| IRTA-SMM-17-004 | 205.1 ± 34.5 | LZ | 48 | IRTA-SMM-17-316 | 51.5 ± 6.9 | TF | 22 |
| IRTA-SMM-17-006 | 127.7 ± 85.0 | LZ | 51 | IRTA-SMM-17-307 | 37.3 ± 12.6 | TF | 30 |
| IRTA-SMM-16-288 | 106.1 ± 75.3 | LZ | 51 | IRTA-SMM-17-436 | 98.6 ± 25.4 | LG | 20 |
| IRTA-SMM-16-290 | 46.1 ± 22.2 | LZ | 51 | IRTA-SMM-17-393 | 44.6 ± 11.5 | LG | 20 |
| IRTA-SMM-16-292 | 39.7 ± 10.5 | LZ | 49 | IRTA-SMM-17-344 | 41.2 ± 0.1 | LP | 8 |
| IRTA-SMM-16-286 | 33.6 ± 6.5 | LZ | 51 | IRTA-SMM-17-335 | 29.1 ± 8.6 | LP | 8 |
| IRTA-SMM-16-293 | 32.7 ± 10.0 | LZ | 51 | IRTA-SMM-17-287 | 11.3 ± 2.3 | LP | 8 |
| IRTA-SMM-17-007 | 15.8 ± 1.7 | LZ | 48 | IRTA-SMM-17-288 | 5.7 ± 3.8 | LP | 8 |
| IRTA-SMM-17-002 | 452.6 ± 23.2 | FV | 43 | IRTA-SMM-17-389 | 226.3 ± 24.5 | EH | 1 |
| IRTA-SMM-17-103 | 118.7 ± 30.3 | GC | 40 | IRTA-SMM-17-324 | 160.4 ± 17.2 | EH | 5 |
| IRTA-SMM-17-107 | 12.2 ± 2.1 | GC | 40 | IRTA-SMM-17-418 | 68.3 ± 9.5 | EH | 2 |
| IRTA-SMM-17-112 | 1.9 ± 0.6 | GC | 40 | IRTA-SMM-17-321 | 31.9 ± 15.0 | EH | 5 |
| IRTA-SMM-17-106 | 1.7 ± 0.1 | GC | 40 | IRTA-SMM-17-425 | 27.8 ± 3.3 | EH | 2 |
| IRTA-SMM-17-358 | 138.9 ± 17.7 | TF | 22 | IRTA-SMM-17-327 | 7.2 ± 0.3 | EH | 2 |
| IRTA-SMM-17-291 | 82.8 ± 22.2 | TF | 22 | ||||
| G. belizeanus | G. caribaeus | ||||||
| IRTA-SMM-17-421 | 5.6 ± 0.1 | EH | 1 | IRTA-SMM-17-003 | Neg. LOD < 0.42 | EH | 3 |
| G. excentricus | G. excentricus | ||||||
| IRTA-SMM-17-001 | 1149.3 ± 212.3 | EH | 1 | IRTA-SMM-17-386 | 12.8 ± 2.8 | TF | 23 |
| IRTA-SMM-17-126 | 226.7 ± 22.1 | GC | 40 | IRTA-SMM-17-429 | 1525.9 ± 634.1 | LG | 20 |
| IRTA-SMM-17-128 | 9.5 ± 2.6 | GC | 40 | IRTA-SMM-17-432 | 962.1 ± 154.7 | LG | 20 |
| IRTA-SMM-17-404 | 1257.64.8 ± 319.3 | TF | 29 | IRTA-SMM-17-413 | 18.1 ± 5.7 | LG | 20 |
| IRTA-SMM-17-405 | 1153.4 ± 238.8 | TF | 29 | IRTA-SMM-17-330 | 2566.7 ± 333.3 | LP | 13 |
| Ref. | Island | Location | Ref. | Island | Location |
|---|---|---|---|---|---|
| 1 | El Hierro | Charco Azul | 27 | Tenerife | La Tejita |
| 2 | El Hierro | Charco Manso | 28 | Tenerife | La Caleta |
| 3 | El Hierro | Tamaduste | 29 | Tenerife | El Pto. de Santiago |
| 4 | El Hierro | La Restinga | 30 | Tenerife | Punta de Teno |
| 5 | El Hierro | Tacoron | 31 | Gran Canaria | Punta Sardina |
| 6 | El Hierro | Orchilla | 32 | Gran Canaria | El Puertillo, Bañaderos |
| 7 | El Hierro | Verodal | 33 | Gran Canaria | Las Canteras |
| 8 | La Palma | La Fajana | 34 | Gran Canaria | El Confital |
| 9 | La Palma | El Puerto Espíndola | 35 | Gran Canaria | Melenara |
| 10 | La Palma | Los Cancajos | 36 | Gran Canaria | Playa Tufia |
| 11 | La Palma | Salemera | 37 | Gran Canaria | Agüimes, Playa El Cabrón |
| 12 | La Palma | El Puerto de Trigo | 38 | Gran Canaria | Arguineguín El Pajar |
| 13 | La Palma | El Faro Fuencaliente | 39 | Gran Canaria | Arguineguín Sta. Águeda |
| 14 | La Palma | Tazacorte | 40 | Gran Canaria | Las Charcas de Agaete |
| 15 | La Palma | Puntagorda | 41 | Fuerteventura | Caleta del Río, El Cotillo |
| 16 | La Gomera | Vallehermoso | 42 | Fuerteventura | Majanicho |
| 17 | La Gomera | La Caleta | 43 | Fuerteventura | Playa Jabalito |
| 18 | La Gomera | Playa de Ávalos | 44 | Fuerteventura | Puerto Lajas |
| 19 | La Gomera | Playa de la Cueva | 45 | Fuerteventura | Puerto Caleta del Fuste |
| 20 | La Gomera | Playa de Vueltas | 46 | Fuerteventura | Gran Tarajal |
| 21 | La Gomera | Alojera | 47 | Fuerteventura | Morro Jable |
| 22 | Tenerife | Charca del Viento | 48 | Lanzarote | Caleta Caballo |
| 23 | Tenerife | Puerto del Sauzal | 49 | Lanzarote | Las Cocinitas |
| 24 | Tenerife | Playa las Teresitas | 50 | Lanzarote | Charco del Palo |
| 25 | Tenerife | El Puertito | 51 | Lanzarote | Puerto Calero |
| 26 | Tenerife | Punta de Abona | 52 | Lanzarote | Playa Mujeres |
| 53 | Lanzarote | El Golfo |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tudó, À.; Gaiani, G.; Rey Varela, M.; Tsumuraya, T.; Andree, K.B.; Fernández-Tejedor, M.; Campàs, M.; Diogène, J. Further Advance of Gambierdiscus Species in the Canary Islands, with the First Report of Gambierdiscus belizeanus. Toxins 2020, 12, 692. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins12110692
Tudó À, Gaiani G, Rey Varela M, Tsumuraya T, Andree KB, Fernández-Tejedor M, Campàs M, Diogène J. Further Advance of Gambierdiscus Species in the Canary Islands, with the First Report of Gambierdiscus belizeanus. Toxins. 2020; 12(11):692. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins12110692
Chicago/Turabian StyleTudó, Àngels, Greta Gaiani, Maria Rey Varela, Takeshi Tsumuraya, Karl B. Andree, Margarita Fernández-Tejedor, Mònica Campàs, and Jorge Diogène. 2020. "Further Advance of Gambierdiscus Species in the Canary Islands, with the First Report of Gambierdiscus belizeanus" Toxins 12, no. 11: 692. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins12110692