Staphylococcal Panton–Valentine Leucocidin and Gamma Haemolysin Target and Lyse Mature Bone Marrow Leucocytes

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
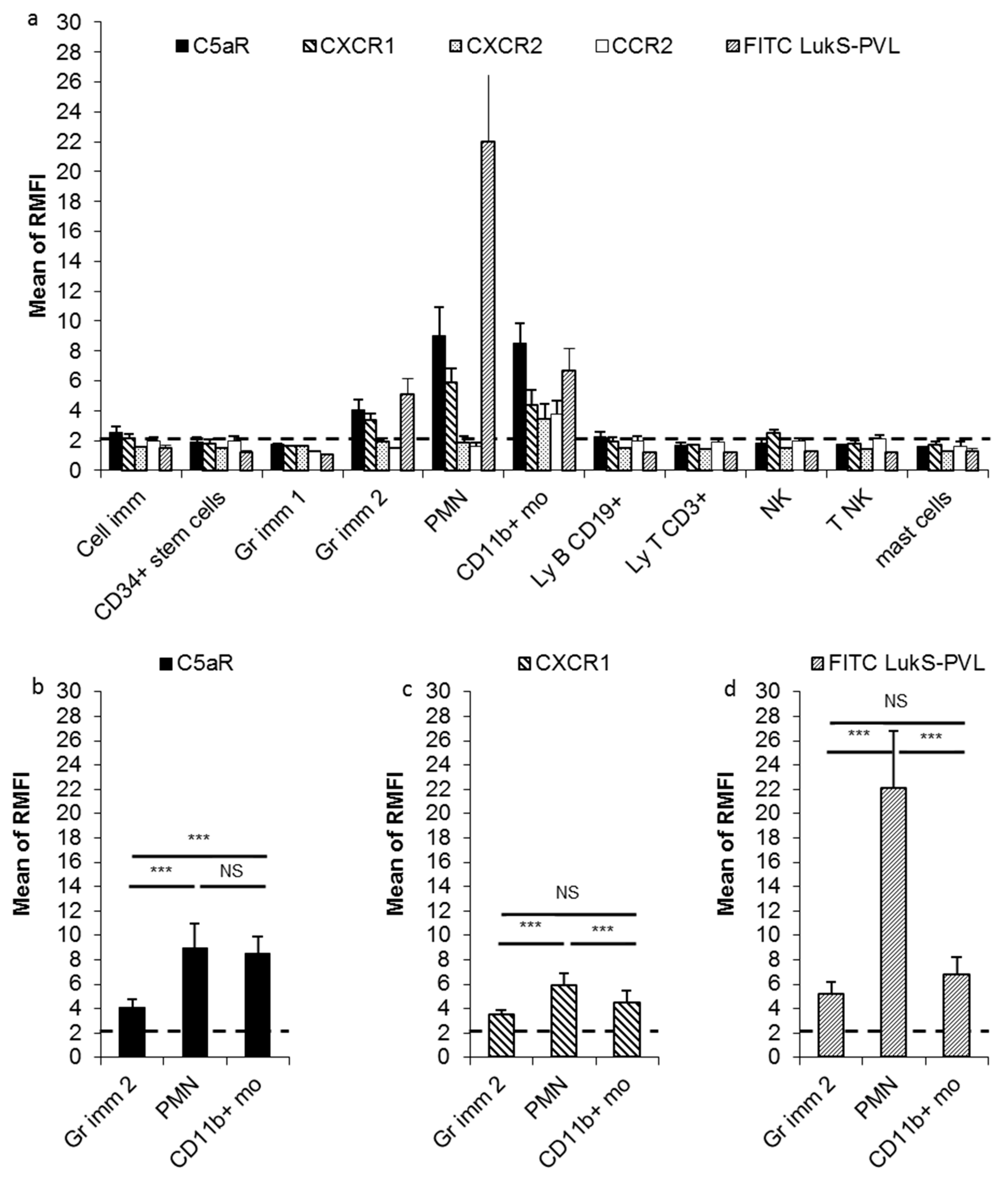
2.1. CXCR1, CXCR2, CCR2, and C5aR Expression on Different Bone Marrow Leucocyte Subpopulation
2.2. Fluorescein Isothiocyanate (FITC) LukS-PVL Binding to Different Bone Marrow Leucocyte Populations
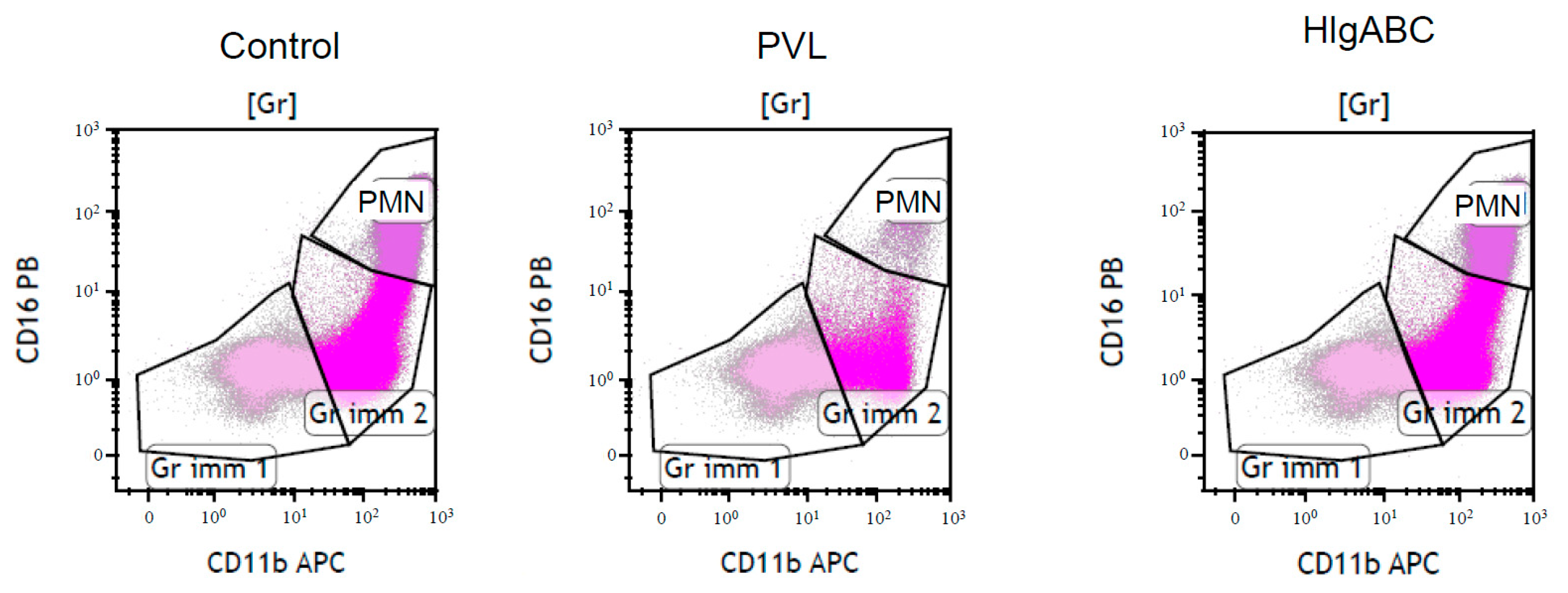
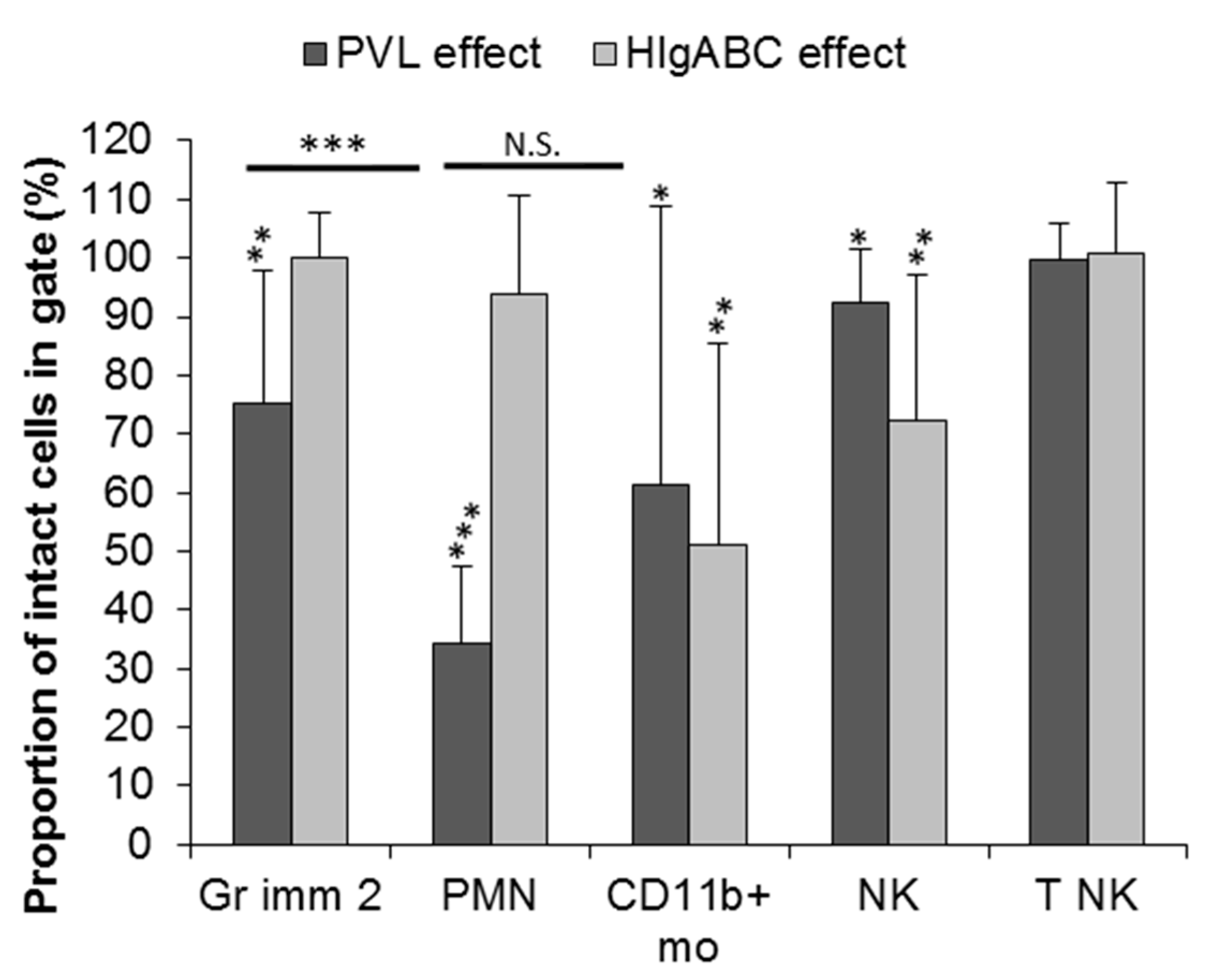
2.3. Effect of PVL and HlgABC on Bone Marrow Leucocyte Sub-Population
2.3.1. Effect of PVL
2.3.2. Effect of HlgABC
3. Discussion
4. Materials and Methods
4.1. Bone Marrow Selection
4.2. S. aureus Leucocidins
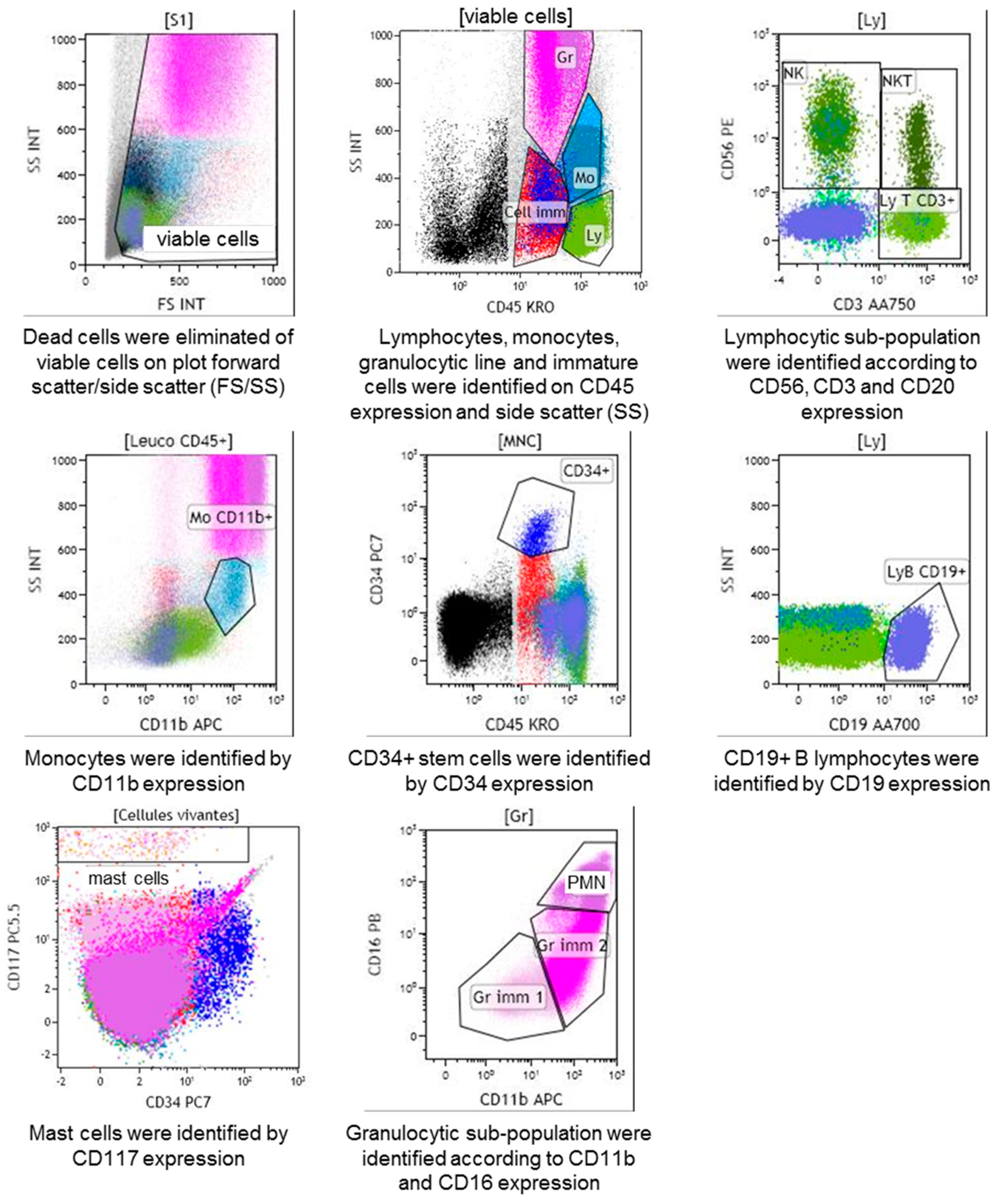
4.3. Expression of Bone Marrow Leucocytes Receptors
4.4. Staphylococcal Leucocidins Effect on Leucocyte Populations
4.5. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Lowy, F.D. Staphylococcus aureus infections. N. Engl. J. Med. 1998, 339, 520–532. [Google Scholar] [CrossRef] [PubMed]
- Grumann, D.; Nübel, U.; Bröker, B.M. Staphylococcus aureus toxins—Their functions and genetics. Infect. Genet. Evol. J. Mol. Epidemiol. Evol. Genet. Infect. Dis. 2013. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spaan, A.N.; Henry, T.; van Rooijen, W.J.M.; Perret, M.; Badiou, C.; Aerts, P.C.; Kemmink, J.; de Haas, C.J.C.; van Kessel, K.P.M.; Vandenesch, F.; et al. The staphylococcal toxin Panton-Valentine Leukocidin targets human C5a receptors. Cell Host Microbe 2013, 13, 584–594. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spaan, A.N.; Vrieling, M.; Wallet, P.; Badiou, C.; Reyes-Robles, T.; Ohneck, E.A.; Benito, Y.; de Haas, C.J.C.; Day, C.J.; Jennings, M.P.; et al. The staphylococcal toxins γ-haemolysin AB and CB differentially target phagocytes by employing specific chemokine receptors. Nat. Commun. 2014, 5, 5438. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tromp, A.T.; Van Gent, M.; Abrial, P.; Martin, A.; Jansen, J.P.; De Haas, C.J.C.; Van Kessel, K.P.M.; Bardoel, B.W.; Kruse, E.; Bourdonnay, E.; et al. Human CD45 is an F-component-specific receptor for the staphylococcal toxin Panton-Valentine leukocidin. Nat. Microbiol. 2018, 3, 708–717. [Google Scholar] [CrossRef] [PubMed]
- Gillet, Y.; Issartel, B.; Vanhems, P.; Fournet, J.-C.; Lina, G.; Bes, M.; Vandenesch, F.; Piémont, Y.; Brousse, N.; Floret, D.; et al. Association between Staphylococcus aureus strains carrying gene for Panton-Valentine leukocidin and highly lethal necrotising pneumonia in young immunocompetent patients. Lancet 2002, 359, 753–759. [Google Scholar] [CrossRef]
- Gillet, Y.; Vanhems, P.; Lina, G.; Bes, M.; Vandenesch, F.; Floret, D.; Etienne, J. Factors predicting mortality in necrotizing community-acquired pneumonia caused by Staphylococcus aureus containing Panton-Valentine leukocidin. Clin. Infect. Dis. Off. Publ. Infect. Dis. Soc. Am. 2007, 45, 315–321. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Diep, B.A.; Chan, L.; Tattevin, P.; Kajikawa, O.; Martin, T.R.; Basuino, L.; Mai, T.T.; Marbach, H.; Braughton, K.R.; Whitney, A.R.; et al. Polymorphonuclear leukocytes mediate Staphylococcus aureus Panton-Valentine leukocidin-induced lung inflammation and injury. Proc. Natl. Acad. Sci. USA 2010, 107, 5587–5592. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Szmigielski, S.; Jeljaszewicz, J.; Wilczynski, J.; Korbecki, M. Reaction of rabbit leucocytes to staphylococcal (Panton-Valentine) leucocidin in vivo. J. Pathol. Bacteriol. 1966, 91, 599–604. [Google Scholar] [CrossRef] [PubMed]
- Werfel, T.; Oppermann, M.; Schulze, M.; Krieger, G.; Weber, M.; Götze, O. Binding of fluorescein-labeled anaphylatoxin C5a to human peripheral blood, spleen, and bone marrow leukocytes. Blood 1992, 79, 152–160. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Löffler, B.; Hussain, M.; Grundmeier, M.; Brück, M.; Holzinger, D.; Varga, G.; Roth, J.; Kahl, B.C.; Proctor, R.A.; Peters, G. Staphylococcus aureus panton-valentine leukocidin is a very potent cytotoxic factor for human neutrophils. PLoS Pathog. 2010, 6, e1000715. [Google Scholar] [CrossRef] [PubMed]
- Holzinger, D.; Gieldon, L.; Mysore, V.; Nippe, N.; Taxman, D.J.; Duncan, J.A.; Broglie, P.M.; Marketon, K.; Austermann, J.; Vogl, T.; et al. Staphylococcus aureus Panton-Valentine leukocidin induces an inflammatory response in human phagocytes via the NLRP3 inflammasome. J. Leukoc. Biol. 2012, 92, 1069–1081. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, C.; Li, Z.; Sheng, W.; Fu, R.; Li, L.; Zhang, T.; Wu, Y.; Xing, L.; Song, J.; Wang, H.; et al. Abnormalities of quantities and functions of natural killer cells in severe aplastic anemia. Immunol. Investig. 2014, 43, 491–503. [Google Scholar] [CrossRef] [PubMed]
- Haynes, D.R.; Harkin, D.G.; Bignold, L.P.; Hutchens, M.J.; Taylor, S.M.; Fairlie, D.P. Inhibition of C5a-induced neutrophil chemotaxis and macrophage cytokine production in vitro by a new C5a receptor antagonist. Biochem. Pharmacol. 2000, 60, 729–733. [Google Scholar] [CrossRef]
- Vergunst, C.E.; Gerlag, D.M.; Dinant, H.; Schulz, L.; Vinkenoog, M.; Smeets, T.J.M.; Sanders, M.E.; Reedquist, K.A.; Tak, P.P. Blocking the receptor for C5a in patients with rheumatoid arthritis does not reduce synovial inflammation. Rheumatol. Oxf. Engl. 2007, 46, 1773–1778. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choudhry, N.; Li, K.; Zhang, T.; Wu, K.-Y.; Song, Y.; Farrar, C.A.; Wang, N.; Liu, C.-F.; Peng, Q.; Wu, W.; et al. The complement factor 5a receptor 1 has a pathogenic role in chronic inflammation and renal fibrosis in a murine model of chronic pyelonephritis. Kidney Int. 2016, 90, 540–554. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tseng, C.W.; Biancotti, J.C.; Berg, B.L.; Gate, D.; Kolar, S.L.; Müller, S.; Rodriguez, M.D.; Rezai-Zadeh, K.; Fan, X.; Beenhouwer, D.O.; et al. Increased Susceptibility of Humanized NSG Mice to Panton-Valentine Leukocidin and Staphylococcus aureus Skin Infection. PLoS Pathog. 2015, 11, e1005292. [Google Scholar] [CrossRef] [PubMed]
- Perret, M.; Badiou, C.; Lina, G.; Burbaud, S.; Benito, Y.; Bes, M.; Cottin, V.; Couzon, F.; Juruj, C.; Dauwalder, O.; et al. Cross-talk between Staphylococcus aureus leukocidins-intoxicated macrophages and lung epithelial cells triggers chemokine secretion in an inflammasome-dependent manner. Cell. Microbiol. 2012, 14, 1019–1036. [Google Scholar] [CrossRef] [PubMed]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hodille, E.; Plesa, A.; Bourrelly, E.; Belmont, L.; Badiou, C.; Lina, G.; Dumitrescu, O. Staphylococcal Panton–Valentine Leucocidin and Gamma Haemolysin Target and Lyse Mature Bone Marrow Leucocytes. Toxins 2020, 12, 725. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins12110725
Hodille E, Plesa A, Bourrelly E, Belmont L, Badiou C, Lina G, Dumitrescu O. Staphylococcal Panton–Valentine Leucocidin and Gamma Haemolysin Target and Lyse Mature Bone Marrow Leucocytes. Toxins. 2020; 12(11):725. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins12110725
Chicago/Turabian StyleHodille, Elisabeth, Adriana Plesa, Eve Bourrelly, Lucie Belmont, Cédric Badiou, Gerard Lina, and Oana Dumitrescu. 2020. "Staphylococcal Panton–Valentine Leucocidin and Gamma Haemolysin Target and Lyse Mature Bone Marrow Leucocytes" Toxins 12, no. 11: 725. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins12110725