Boronic Acids of Pharmaceutical Importance Affect the Growth and Photosynthetic Apparatus of Cyanobacteria in a Dose-Dependent Manner


Abstract
:
1. Introduction
2. Results
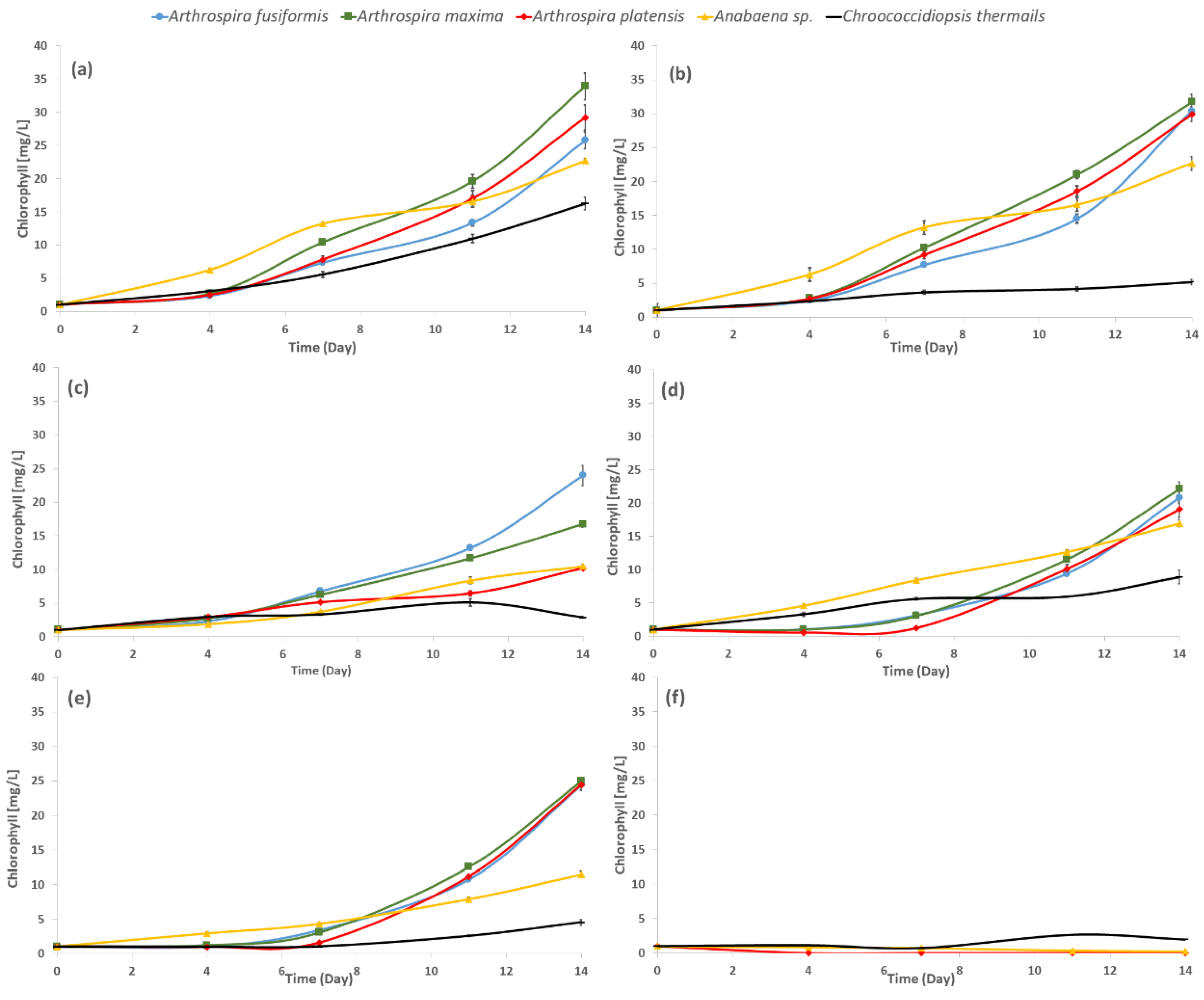
2.1. Growth of Cyanobacteria
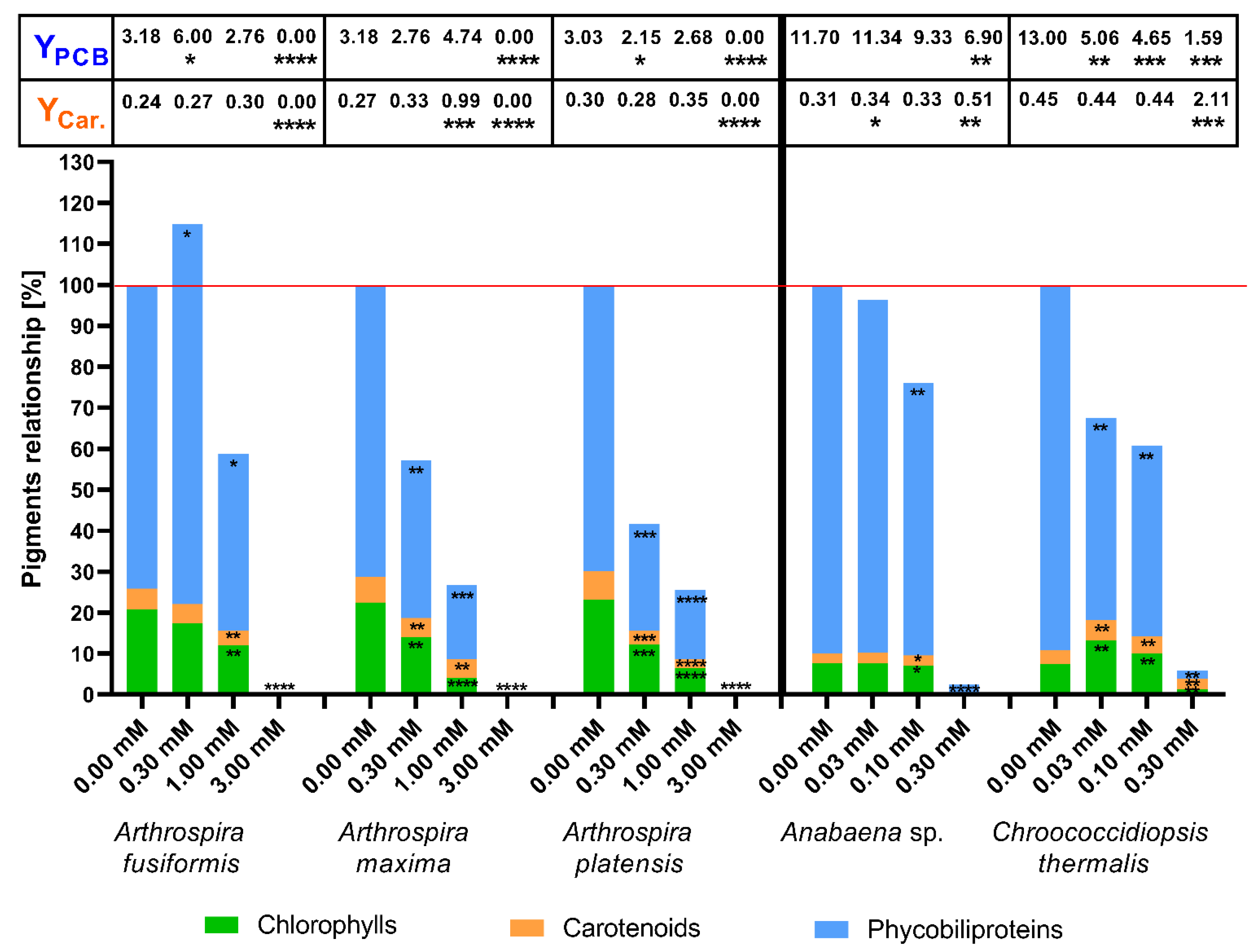
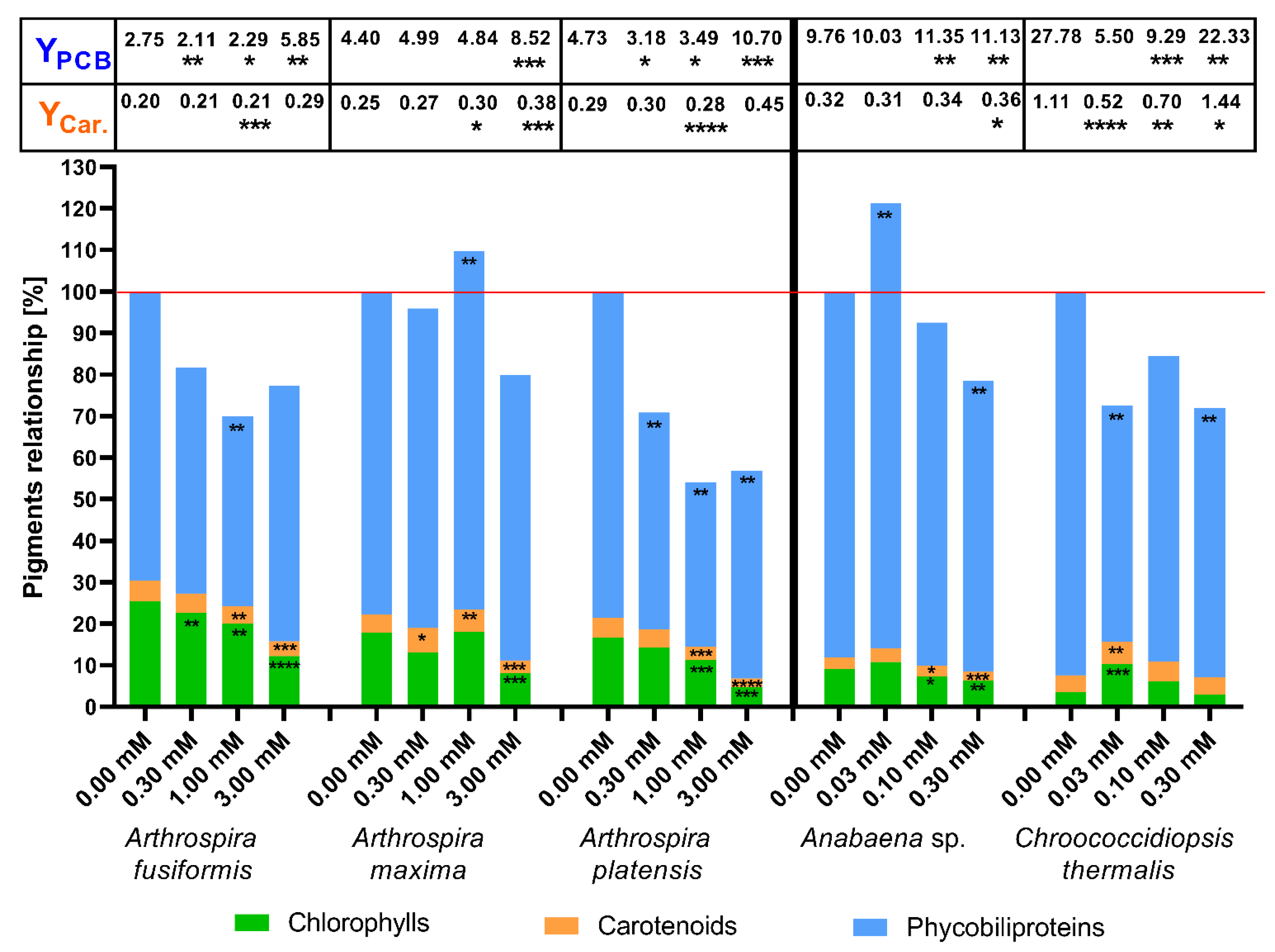
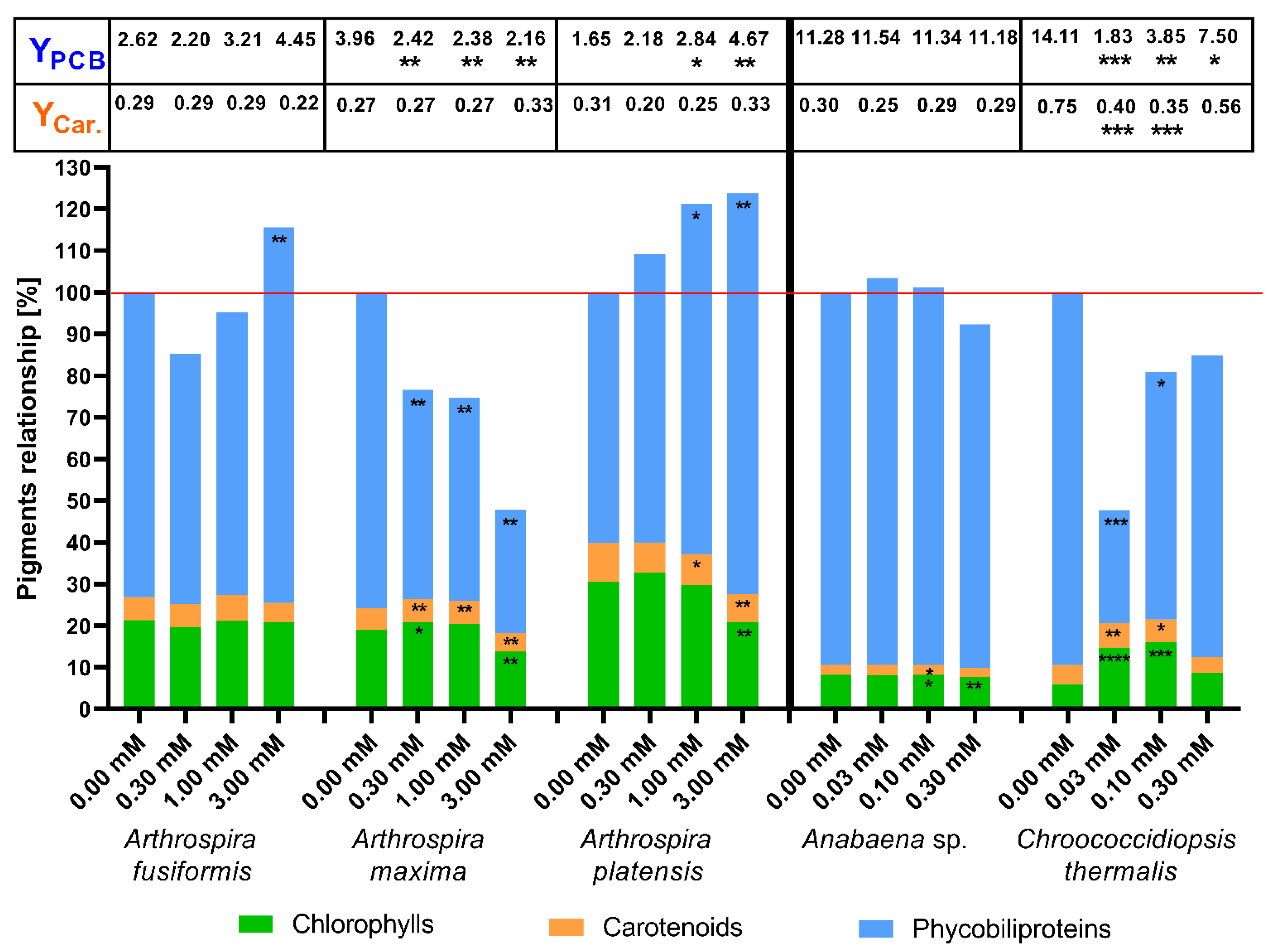
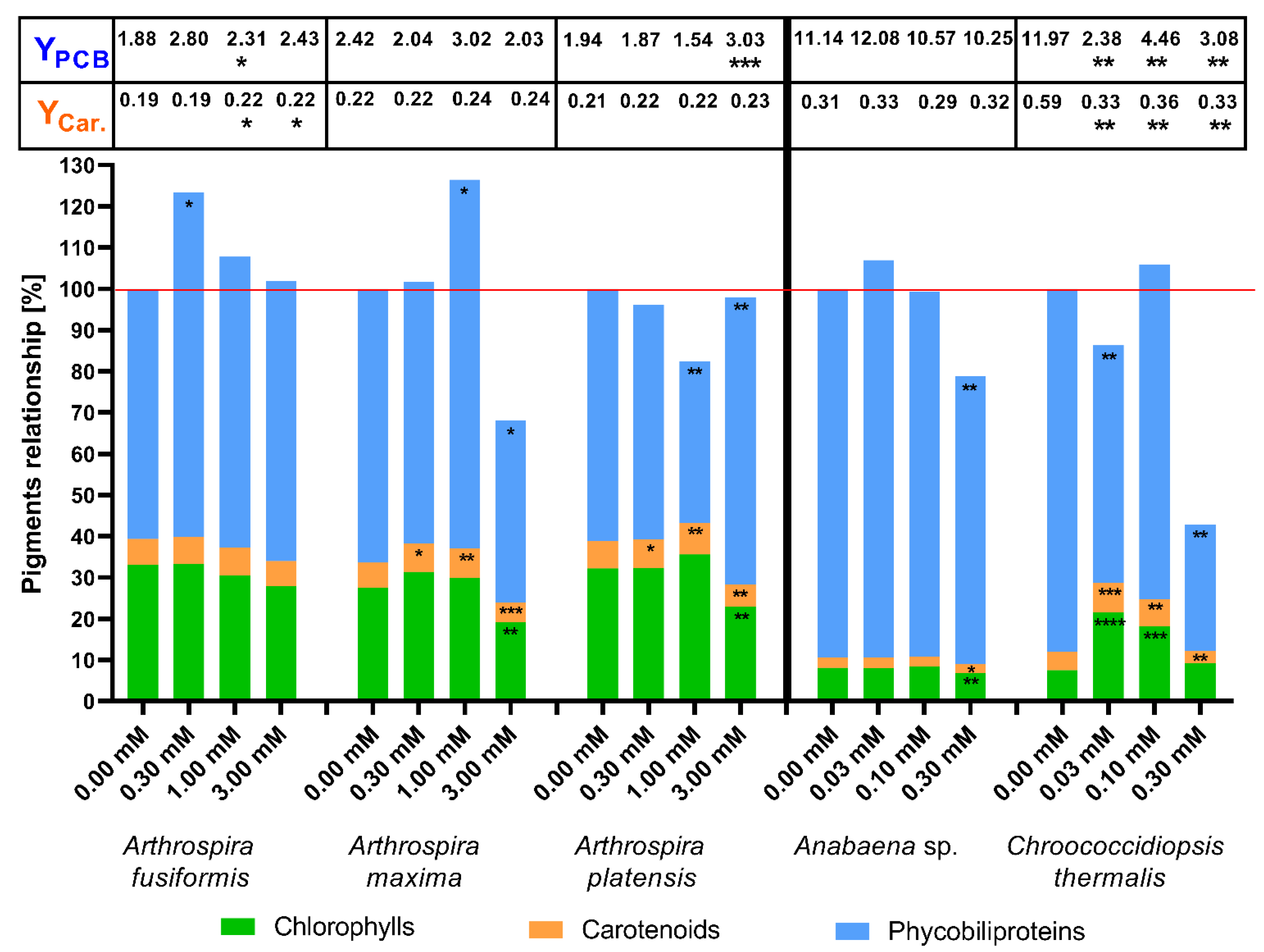
2.2. The Concentration of Photosynthetic Pigments as a Factor of Change in Photosynthetic Apparatus
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Tested Substances and Other Chemicals
5.2. Experimental Cultivations
5.3. Measurements of the Photosynthetic Pigments Chlorophyll, Phycobiliproteins and Carotenoids
5.4. Determination of the Growth Rate Ratio
5.5. Statistical Analysis
Author Contributions
Funding
Conflicts of Interest
References
- Bolanos, L.; Lukaszewski, K.; Bonilla, I.; Blevins, D. Why boron? Plant Physiol. Biochem. 2004, 40, 907–912. [Google Scholar] [CrossRef]
- Howe, P.D. A Review of Boron Effects in the Environment. Biol. Trace Element Res. 1998, 66, 153–166. [Google Scholar] [CrossRef]
- Parks, J.F.; Edwards, M. Boron in the Environment. Environ. Sci. Technol. 2005, 35, 81–114. [Google Scholar] [CrossRef]
- Geffen, N.; Semiat, R.; Eisen, M.S.; Balazs, Y.; Katz, I.; Dosoretz, C.G. Boron removal from water by complexation to polyol compounds. J. Membrane Sci. 2006, 286, 45–51; [Google Scholar] [CrossRef]
- Dembitsky, V.; Smoum, R.; Al Aziz Quntar, A.; Abu Ali, H.; Pergment, I.; Srebnik, M. Natural occurrence of boron-containing compounds in plants, algae and microorganisms. Plant Sci. 2002, 163, 931–942. [Google Scholar] [CrossRef]
- Rock, F.L.; Mao, W.; Yaremchuk, A.; Tukalo, M.; Crépin, T.; Zhou, H.; Zhang, Y.K.; Hernandez, V.; Akama, T.; Baker, S.J.; et al. An Antifungal Agent Inhibits an Aminoacyl-tRNA Synthetase by Trapping tRNA in the Editing Site. Science 2007, 316, 1759. [Google Scholar] [CrossRef]
- Trippier, P.C.; McGuigan, C. Boronic acids in medicinal chemistry: Anticancer, antibacterial and antiviral applications. MedChemComm 2010, 1, 183–198. [Google Scholar] [CrossRef]
- Zhang, J.; Zhu, M.Y.; Lin, Y.N.; Zhou, H.C. The synthesis of benzoxaboroles and their applications in medicinal chemistry. Sci. China Chem. 2013, 56, 1372–1381. [Google Scholar] [CrossRef]
- Nocentini, A.; Supuran, C.T.; Winum, J.Y. Benzoxaborole compounds for therapeutic uses: A patent review (2010–2018). Expert Opin. Ther. Patents 2018, 28, 493–504. [Google Scholar] [CrossRef]
- Das, B.C.; Thapa, P.; Karki, R.; Schinke, C.; Das, S.; Kambhampati, S.; Banerjee, S.K.; Veldhuizen, P.V.; Verma, A.; Weiss, L.M.; et al. Boron chemicals in diagnosis and therapeutics. Future Med. Chem. 2013, 5, 653–676. [Google Scholar] [CrossRef] [Green Version]
- Sonoiki, E.; Palencia, A.; Guo, D.; Ahyong, V.; Dong, C.; Li, X.; Hernandez, V.S.; Zhang, Y.-K.; Choi, W.; Gut, J.; et al. Antimalarial benzoxaboroles target Plasmodium falciparum leucyl-tRNA synthetase. Antimicrob. Agents Chemother. 2016, 60, 4886–4895. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Manhas, R.; Tandon, S.; Sen, S.S.; Tiwari, N.; Munde, M.; Madhubala, R. Leishmania donovani parasites are inhibited by the benzoxaborole AN2690 targeting leucyl-tRNA synthetase. Antimicrob. Agents Chemother. 2018, 62, e00079-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wall, R.J.; Rico, E.; Lukac, I.; Zuccotto, F.; Elg, S.; Glibert, I.H.; Freund, Y.; Alley, M.R.K.; Field, M.C.; Wyllie, S.; et al. Clinical and veterinary trypanocidal benzoxaboroles target CPSF3. Proc. Natl. Acad. Sci. USA 2018, 115, 9616–9621. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wieczorek, D.; Lipok, J.; Borys, K.M.; Adamczyk-Woźniak, A.; Sporzyński, A. Investigation of fungicidal activity of 3-piperazine-bis(benzoxaborole) and its boronic acid analogue. Appl. Organometal. Chem. 2014, 28, 347–350. [Google Scholar] [CrossRef]
- Drzyzga, D.; Lipok, J. Glyphosate dose modulates the uptake of inorganic phosphate by freshwater cyanobacteria. J. Appl. Phycol. 2018, 30, 299–309. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adamczyk-Woźniak, A.; Borys, K.M.; Sporzyński, A. Recent developments in the chemistry and biological applications of benzoxaboroles. Chem. Rev. 2015, 115, 5224–5247. [Google Scholar] [CrossRef]
- Fontaine, F.; Hequet, A.; Voisin-Chiret, A.S.; Bouillon, A.; Lesnard, A.; Cresteil, T.; Jolivalt, C.; Rault, S. Boronic species as promising inhibitors of the Staphylococcus aureus NorA efflux pump: Study of 6-substituted pyridine-3-boronic acid derivatives. Eur. J. Med. Chem. 2015, 95, 185–198. [Google Scholar] [CrossRef]
- Benkovic, S.J.; Baker, S.J.; Alley, M.R.K.; Woo, Y.H.; Zhang, Y.K.; Akama, T.; Mao, W.; Baboval, J.; Rajagopalan, P.T.R.; Wall, M.; et al. Identification of borinic esters as inhibitors of bacterial cell growth and bacterial methyltransferases. J. Med. Chem. 2005, 48, 7468–7476. [Google Scholar] [CrossRef]
- Korkegian, A.; O’Malley, T.; Xia, Y.; Zhou, Y.; Carter, D.S.; Sunde, B.; Flint, L.; Thompson, D.; Ioerger, T.R.; Sacchettini, J.; et al. The 7-phenylbenzoxaborole series is active against Mycobacterium tuberculosis. Tuberculosis 2018, 108, 96–98. [Google Scholar] [CrossRef]
- Lipok, J.; Niemczyk, E. Fikobiliproteiny Mikroalg–Właściwości, Racjonalne Pozyskiwanie I Zastosowanie” W „Na Pograniczu Chemii I Biologii; Koroniak, H., Ed.; Wydawnictwo UAM: Poznań, Poland, 2017; Volume 24. [Google Scholar]
- Tomitani, A.; Okada, K.; Miyashia, H.; Matthijs, H.C.P.; Ohno, T.; Tanaka, A. Chlorophyll b and phycobilins in the common ancestor of cyanobacteria and chloroplasts. Nature 1999, 400, 159–162. [Google Scholar] [CrossRef]
- Stolarzewicz, I.; Kapturowska, A.; Białecka-Florjanczyk, E. Mikrobiologiczne źródła barwników w technologii żywności. Post. Mikrobiol. 2012, 51, 167–176. [Google Scholar]
- Khatoon, H.; Leong, L.K.; Rahman, N.A.; Mian, S.; Begum, H.; Banerjee, S.; Endut, A. Effects of different light source and media on growth and production of phycobiliprotein from freshwater cyanobacteria. Bioresour. Technol. 2018, 249, 652–658. [Google Scholar] [CrossRef] [PubMed]
- Rizwan, M.; Mujtaba, G.; Memon, S.A.; Lee, K.; Rashaid, N. Exploring the potential of microalgae for new biotechnology applications and beyond: A review. Renew. Sustain. Energy Rev. 2018, 92, 394–404. [Google Scholar] [CrossRef]
- Encyclopedia of Astrobiology; Gargaud, M. (Ed.) Springer: Berlin/Heidelberg, Germany, 2011. [Google Scholar] [CrossRef] [Green Version]
- Drzyzga, D.; Forlani, G.; Niemczyk, E.; Lipok, J. The aminophosphonate glyphosine enhances phycobiliprotein yields from selected cyanobacterial cultures. J. Appl. Phycol. 2017, 30. [Google Scholar] [CrossRef]
- Franklin, N.M.; Adams, M.S.; Stauber, J.L.; Lim, R.P. Develoment of an Improved Rapid Enzyme Inhibition Bioassay with Marine and Freshwater Microalgae Using Flow Cytometry; Arch. Environ. Contam. Toxicol. 2001, 40, 469–480. [Google Scholar] [CrossRef]
- Forlani, G.; Pavan, M.; Gramek, M.; Kafarski, P.; Lipok, J. Biochemical Bases for a Widespread Tolerance of Cyanobacteria to the Phosphonate Herbicide Glyphosate. Plant Cell Physiol. 2008, 49, 443–456. [Google Scholar] [CrossRef]
- Żyszka-Haberecht, B.; Poliwoda, A.; Lipok, J. Biocatalytic hydrogenation of the C=C bond in the enone unit of hydroxylated chalcones—process arising from cyanobacterial adaptations. Appl. Microbiol. Biotechnol. 2018, 102, 7097–7111. [Google Scholar] [CrossRef] [Green Version]
- Eide-Haugmo, I.; Brakstad, O.G.; Hoff, K.A.; Sørheim, K.R.; da Silva, E.F.; Svendsen, H.F. Environmental impact of amines. Energy Proc. 2009, 1, 1297–1304. [Google Scholar] [CrossRef] [Green Version]
- Pogrzeba, J.R. The influence of selected aminophosphonate and boronic compounds on growth and metabolism of cyanobacteria. Master Thesis, University of Opole, Opole, Poland, 9 July 2020. [Google Scholar]
- Adamczyk-Woźniak, A.; Komarovska-Porokhnyavets, O.; Misterkiewicz, B.; Novkov, V.P.; Sporzyński, A. Biological activity of selected boronic acids and their derivatives. Appl. Organometal. Chem. 2012, 26, 390–393. [Google Scholar] [CrossRef]
- Bhatnagar, M.; Bhatnagar, A. Algal and cyanobacterial responses to fluoride. Fluoride 2000, 33, 55–65. [Google Scholar]
- Rezayian, M.; Niknam, V.; Ebrahimzadeh, H. Response to different stresses in cyanobacteria. Iran. J. Plant Physiol. 2016, 9, 2773–2787. [Google Scholar] [CrossRef]
- Fernandus, T.A.; Iyer, V.; Apte, S.K. Differential Responses of Nitrogen-Fixing Cyanobacteria to Salinity and Osmotic Stresses. Mol. Biol. Agric. Div. 1992, 59, 899–904. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Kreij, C.; Başar, H. Leaf tip yellowing in eggplant is caused by boron deficiency. J. Plant Nutr. 1997, 20, 47–53. [Google Scholar] [CrossRef]
- Reed, H.S. A physiological study of boron deficiency on plants. J. Agric. Sc. 1947, 17, 11. [Google Scholar] [CrossRef]
- Bonilla, I.; Garcia-González, M.; Mateo, P. Boron Requirement in Cyanobacteria. Plant Physiol. 1990, 94, 1554–1560. [Google Scholar] [CrossRef] [Green Version]
- Zarzeczańska, D.; Adamczyk-Woźniak, A.; Kulpa, A.; Ossowski, T.; Sporzyński, A. Fluorinated Boronic Acids: Acidity and Hydrolytic Stability of Fluorinated Phenylboronic Acids. Eur. J. Inorg. Chem. 2017, 4493–4498. [Google Scholar] [CrossRef] [Green Version]
- Mackinney, G. Absorption of light by chlorophyll solutions. J. Biol. Chem. 1945, 140, 315–322. [Google Scholar]
- Chamivitz, D.; Sandman, G. Molecular and biochemical characterization of herbicide resistant mutants of cyanobacteria reveals that phytoene desaturation is a rate-limiting step in carotenoid biosynthesis. J. Biol. Chem. 1993, 265, 17348–17353. [Google Scholar]
- Bennett, A.; Bogorad, L. Complimentary chromatic adaptation in a filamentous blue-green alga. J. Cell Biol. 1973, 58, 419–435. [Google Scholar] [CrossRef]







{kind=link}
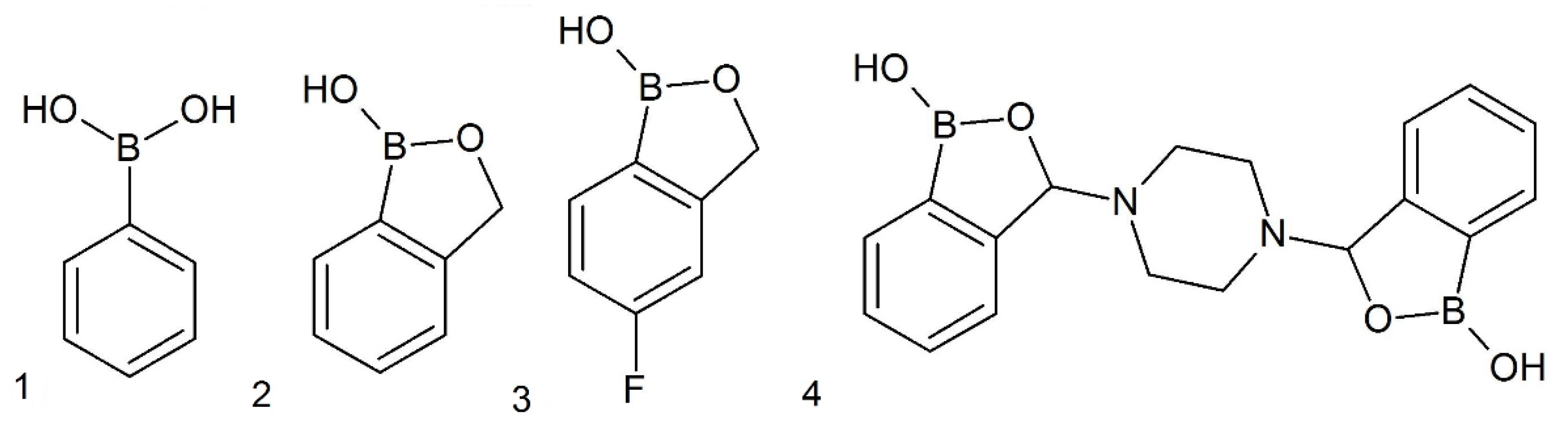{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Substance | Concentration | Arthrospira fusiformis | Arthrospira maxima | Arthrospira platensis | Concentration | Anabaena sp. | Chroococcidiopsis thermalis |
|---|---|---|---|---|---|---|---|
| Phenylboronic acid (1) | 0.00 mM | 100.0 ± 1.7 | 100.0 ± 3.8 | 100.0 ± 3.9 | 0.00 mM | 100.0 ± 3.2 | 100.0 ± 8.4 |
| 0.30 mM | 92.2 ± 2.4 ** | 75.8 ± 15.2* | 89.1 ± 3.8 ** | 0.03 mM | 122.7 ± 7.3 ** | 406.9 ± 2.1 **** | |
| 1.00 mM | 83.1 ± 3.8 *** | 95.9 ± 6.7 | 64.5 ± 3.8 **** | 0.10 mM | 81.4 ± 6.2 ** | 224.9 ± 7.1 **** | |
| 3.00 mM | 53.0 ± 5.1 **** | 42.4 ± 2.4 **** | 25.3 ± 3.2 **** | 0.30 mM | 70.1 ± 3.5 **** | 97.9 ± 9.8 | |
| Benzoxaborole (2) | 0.00 mM | 100.0 ± 3.6 | 100.0 ± 2.3 | 100.0 ± 5.6 | 0.00 mM | 100.0 ± 0.9 | 100.0 ± 0.9 |
| 0.30 mM | 98.9 ± 4.5 | 105.8 ± 1.2 **** | 103.8 ± 7.3 | 0.03 mM | 98.8 ± 6.3 | 297.1 ± 3.0 **** | |
| 1.00 mM | 107.2 ± 4.4 ** | 101.5 ± 2.2 | 93.3 ± 2.4 | 0.10 mM | 100.0 ± 2.2 | 330.3 ± 4.9 **** | |
| 3.00 mM | 91.5 ± 3.7 ** | 68.5 ± 3.9 **** | 65.4 ± 6.4 *** | 0.30 mM | 93.5 ± 2.3 ** | 157.9 ± 8.9 **** | |
| 5-fluoro-substituted benzoxaborole (3) | 0.00 mM | 100.0 ± 2.9 | 100.0 ± 4.0 | 100.0 ± 1.8 | 0.00 mM | 100.0 ± 6.9 | 100.0 ± 14.0 |
| 0.30 mM | 102.3 ± 2.3 | 114.7 ± 3.4 ** | 103.7 ± 4.7 | 0.03 mM | 97.2 ± 0.6 | 338.9 ± 4.1 **** | |
| 1.00 mM | 92.8 ± 3.0 | 110.7 ± 7.9 * | 110.6 ± 5.5 * | 0.10 mM | 102.5 ± 2.8 | 280.5 ± 4.0 **** | |
| 3.00 mM | 80.8 ± 2.8 *** | 69.5 ± 1.7 **** | 69.5 ± 0.9 **** | 0.30 mM | 84.5 ± 2.9 ** | 127.1 ± 7.5 ** | |
| 3-piperazine-bis(benzoxaborole) (4) | 0.00 mM | 100.0 ± 4.1 | 100.0 ± 2.1 | 100.0 ± 4.4 | 0.00 mM | 100.0 ± 1.3 | 100.0 ± 16.8 |
| 0.30 mM | 79.8 ± 5.5 ** | 57.6 ± 9.3 *** | 47.7 ± 7.3 **** | 0.03 mM | 98.17 ± 3.6 | 200.9 ± 8.4 **** | |
| 1.00 mM | 51.5 ± 11.9 *** | 16.1 ± 0.2 **** | 20.3 ± 6.3 **** | 0.10 mM | 94.2 ± 3.1 | 142.3 ± 2.6 *** | |
| 3.00 mM | 0.0 **** | 0.0 **** | 0.0 **** | 0.30 mM | 0.0 **** | 23.3 ± 4.2 **** |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Niemczyk, E.; Pogrzeba, J.; Adamczyk-Woźniak, A.; Lipok, J. Boronic Acids of Pharmaceutical Importance Affect the Growth and Photosynthetic Apparatus of Cyanobacteria in a Dose-Dependent Manner. Toxins 2020, 12, 793. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins12120793
Niemczyk E, Pogrzeba J, Adamczyk-Woźniak A, Lipok J. Boronic Acids of Pharmaceutical Importance Affect the Growth and Photosynthetic Apparatus of Cyanobacteria in a Dose-Dependent Manner. Toxins. 2020; 12(12):793. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins12120793
Chicago/Turabian StyleNiemczyk, Emilia, Jerzy Pogrzeba, Agnieszka Adamczyk-Woźniak, and Jacek Lipok. 2020. "Boronic Acids of Pharmaceutical Importance Affect the Growth and Photosynthetic Apparatus of Cyanobacteria in a Dose-Dependent Manner" Toxins 12, no. 12: 793. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins12120793