Snake C-Type Lectins Potentially Contribute to the Prey Immobilization in Protobothrops mucrosquamatus and Trimeresurus stejnegeri Venoms


Abstract
:1. Introduction
2. Results
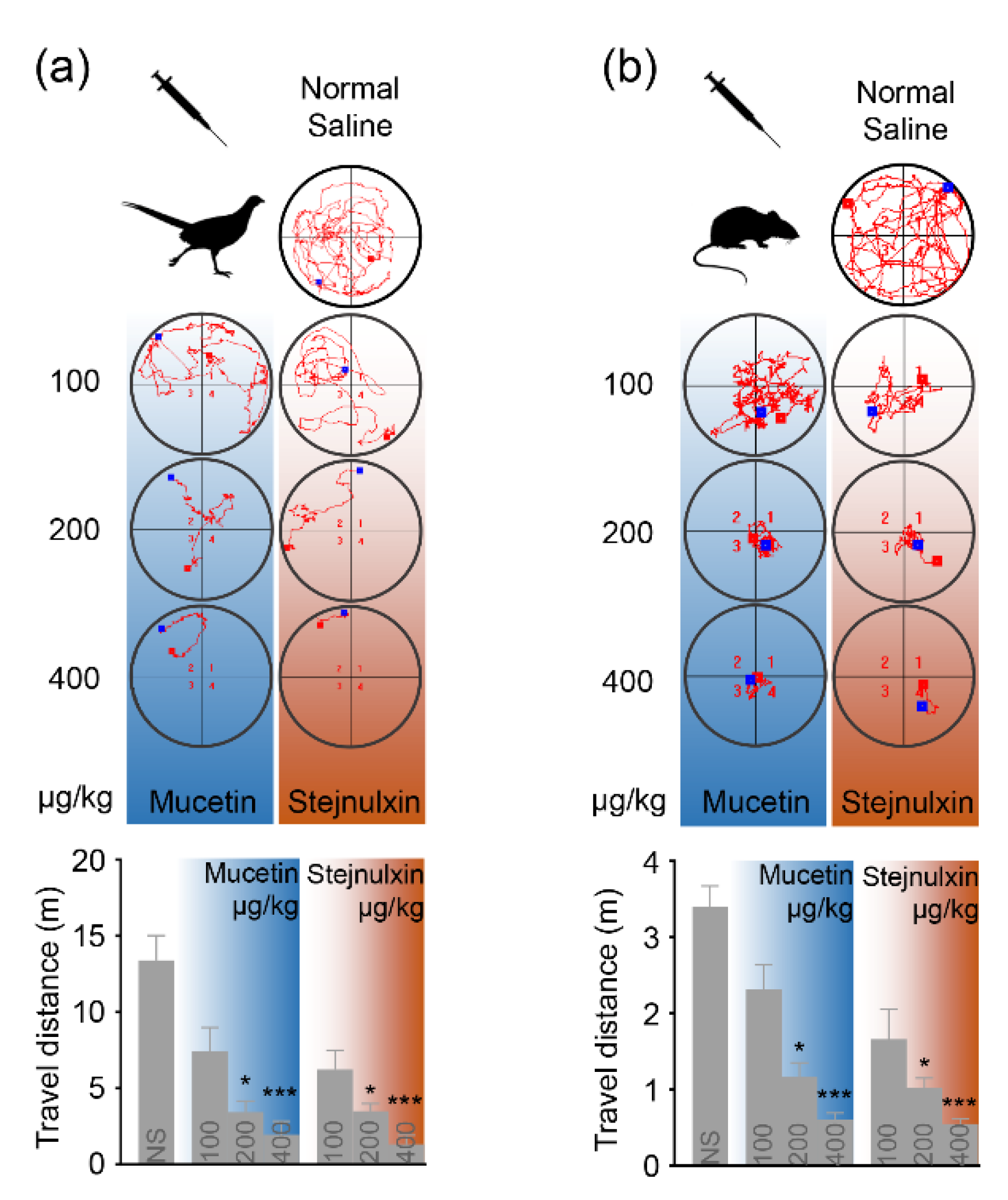
2.1. Snaclecs Can Rapidly Immobilize Prey
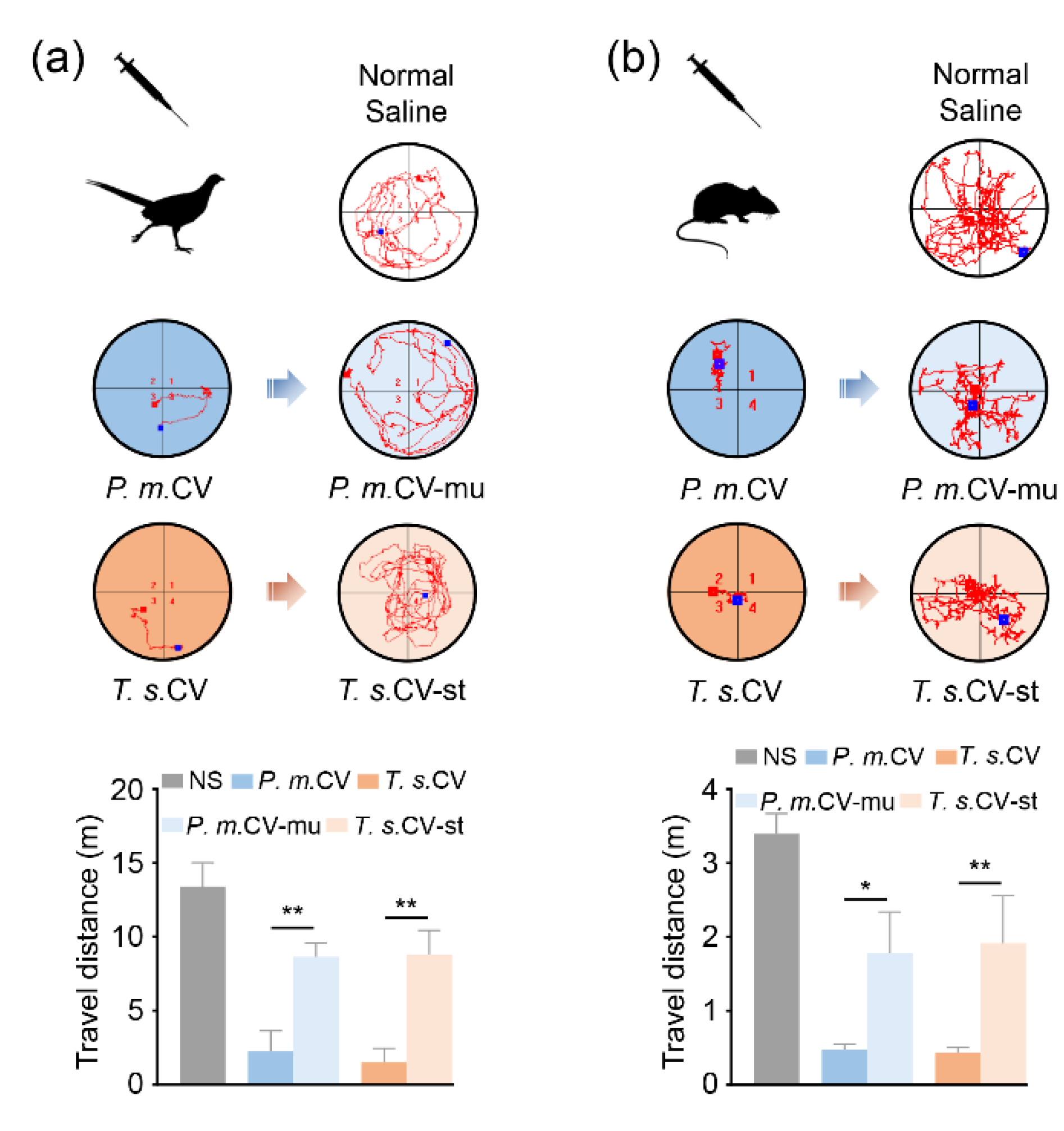
2.2. Snaclecs are Critical for Viper Venom Induced Prey Paralysis
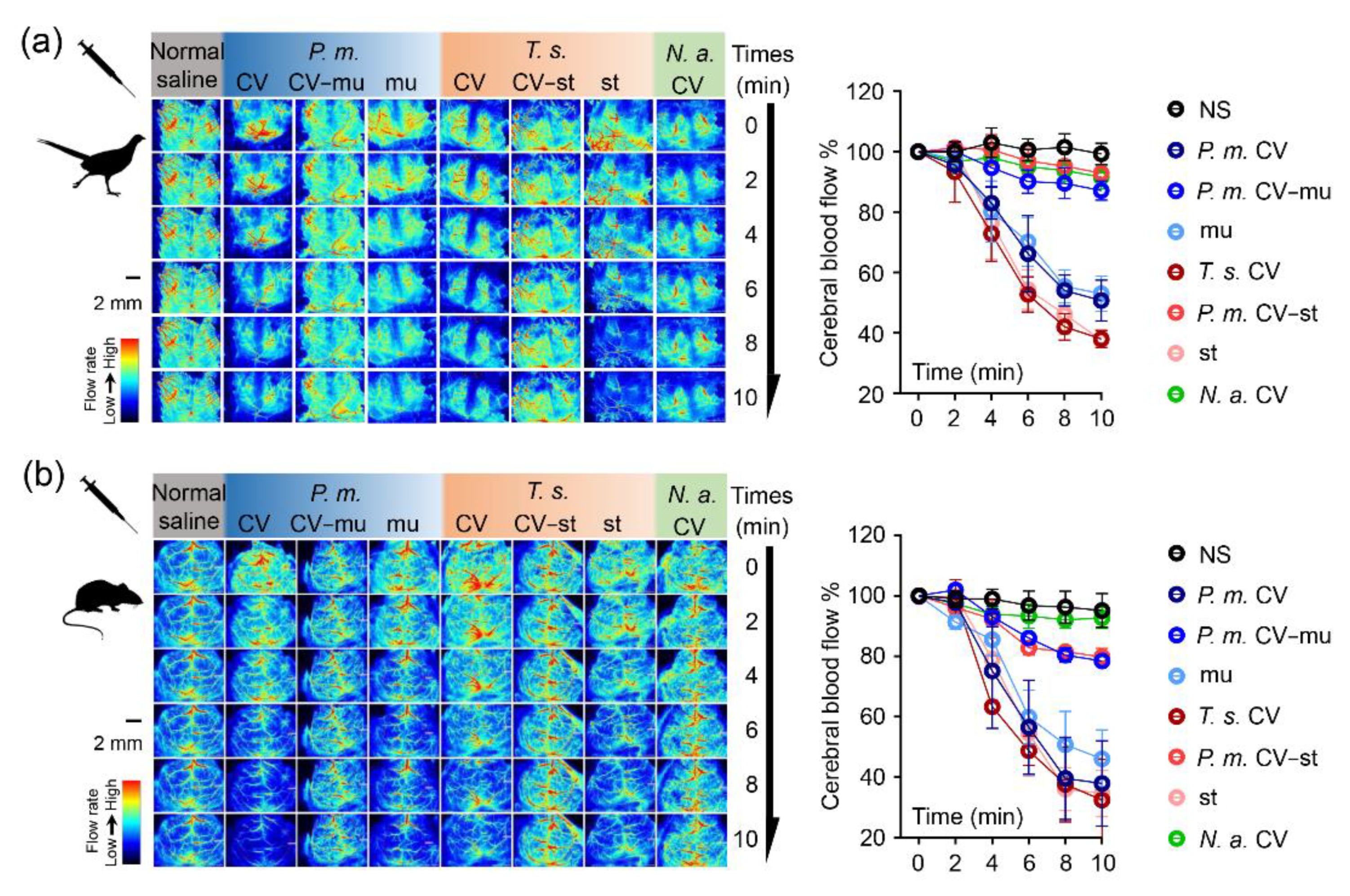
2.3. Snaclecs Induce Cerebral Ischemia
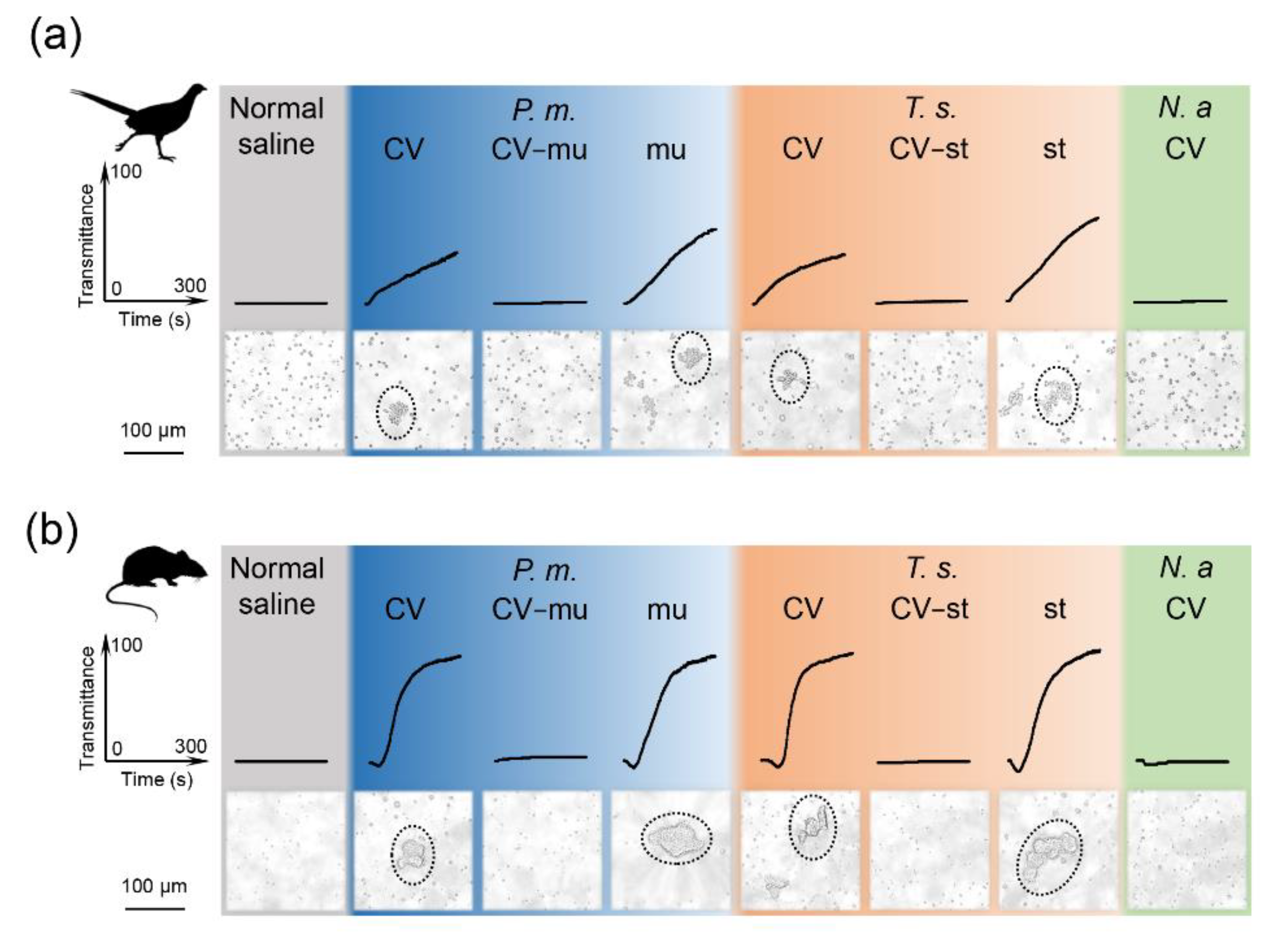
2.4. Snaclecs Activate Thrombocytes or Platelets
3. Discussion
4. Material and Methods
4.1. Venom Collection and Toxins Purification
4.2. The Locomotor Activity in an Open Field Test
4.3. Continuous Measurement of Cerebral Cortex Blood Flow
4.4. Platelet/Thrombocyte Isolation and Stimulation
4.5. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Barlow, A.; Pook, C.E.; Harrison, R.A.; Wuster, W. Coevolution of diet and prey-specific venom activity supports the role of selection in snake venom evolution. Proc. R. Soc. B Biol. Sci. 2009, 276, 2443–2449. [Google Scholar] [CrossRef] [Green Version]
- Lewis, R.L.; Gutmann, L. Snake venoms and the neuromuscular junction. Semin. Neurol. 2004, 24, 175–179. [Google Scholar] [CrossRef]
- Tasoulis, T.; Isbister, G.K. A Review and Database of Snake Venom Proteomes. Toxins 2017, 9, 290. [Google Scholar] [CrossRef] [Green Version]
- Del Brutto, O.H.; Del Brutto, V.J. Neurological complications of venomous snake bites: A review. Acta Neurol. Scand. 2012, 125, 363–372. [Google Scholar] [CrossRef]
- Fry, B.G.; Wuster, W. Assembling an arsenal: Origin and evolution of the snake venom proteome inferred from phylogenetic analysis of toxin sequences. Mol. Biol. Evol. 2004, 21, 870–883. [Google Scholar] [CrossRef] [Green Version]
- Modahl, C.M.; Mrinalini; Frietze, S.; Mackessy, S.P. Adaptive evolution of distinct prey-specific toxin genes in rear-fanged snake venom. P Roy. Soc. B-Biol. Sci. 2018, 285. [Google Scholar] [CrossRef]
- Charles, W.; Radcliffe, T.P.; Feiler, F.; Warnoch, N.; Byers, T.; Radcliffe, A.; A., D. Immobilization of mice following envenomation by cobras. Bull. Psychon. Soc. 1983, 21, 243–246. [Google Scholar]
- Tan, C.H.; Tan, K.Y.; Ng, T.S.; Quah, E.S.H.; Ismail, A.K.; Khomvilai, S.; Sitprija, V.; Tan, N.H. Venomics of Trimeresurus (Popeia) nebularis, the Cameron Highlands Pit Viper from Malaysia: Insights into Venom Proteome, Toxicity and Neutralization of Antivenom. Toxins 2019, 11, 95. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Patra, A.; Kalita, B.; Chanda, A.; Mukherjee, A.K. Proteomics and antivenomics of Echis carinatus carinatus venom: Correlation with pharmacological properties and pathophysiology of envenomation. Sci. Rep. 2017, 7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abidin, S.A.Z.; Rajadurai, P.; Chowdhury, M.E.H.; Rusmili, M.R.A.; Othman, I.; Naidu, R. Proteomic Characterization and Comparison of Malaysian Tropidolaemus wagleri and Cryptelytrops purpureomaculatus Venom Using Shotgun-Proteomics. Toxins 2016, 8, 299. [Google Scholar] [CrossRef] [Green Version]
- Tan, K.Y.; Tan, N.H.; Tan, C.H. Venom proteomics and antivenom neutralization for the Chinese eastern Russell’s viper, Daboia siamensis from Guangxi and Taiwan. Sci. Rep. 2018, 8. [Google Scholar] [CrossRef] [PubMed]
- Kalita, B.; Singh, S.; Patra, A.; Mukherjee, A.K. Quantitative proteomic analysis and antivenom study revealing that neurotoxic phospholipase A (2) enzymes, the major toxin class of Russell’s viper venom from southern India, shows the least immuno-recognition and neutralization by commercial polyvalent antivenom. Int. J. Biol. Macromol. 2018, 118, 375–385. [Google Scholar] [CrossRef] [PubMed]
- Lu, Q.M.; Navdaev, A.; Clemetson, J.M.; Clemetson, K.J. Snake venom C-type lectins interacting with platelet receptors. Structure-function relationships and effects on haemostasis. Toxicon 2005, 45, 1089–1098. [Google Scholar] [CrossRef] [PubMed]
- Clemetson, K.J. Snaclecs (snake C-type lectins) that inhibit or activate platelets by binding to receptors. Toxicon 2010, 56, 1236–1246. [Google Scholar] [CrossRef]
- Wei, Q.; Lu, Q.M.; Jin, Y.; Li, R.; Wei, J.F.; Wang, W.Y.; Xiong, Y.L. Purification and cloning of a novel C-type lectin-like protein with platelet aggregation activity from Trimeresurus mucrosquamatus venom. Toxicon 2002, 40, 1331–1338. [Google Scholar] [CrossRef]
- Lee, W.H.; Du, X.Y.; Lu, Q.M.; Clemetson, K.J.; Zhang, Y. Stejnulxin, a novel snake C-type lectin-like protein from Trimeresurus stejnegeri venom is a potent platelet agonist acting specifically via GPVl. Thromb. Haemost. 2003, 90, 662–671. [Google Scholar] [CrossRef] [Green Version]
- Lacosteeleaume, A.S.; Bleux, C.; Quere, P.; Cordert, F.; Corbel, C.; Kanellopouloslangevin, C. Biochemical and Functional-Characterization of an Avian Homolog of the Integrin Gpiib-Iiia Present on Chicken Thrombocytes. Exp. Cell Res. 1994, 213, 198–209. [Google Scholar] [CrossRef]
- Abumiya, T.; Fitridge, R.; Mazur, C.; Copeland, B.R.; Koziol, J.A.; Tschopp, J.F.; Pierschbacher, M.D.; del Zoppo, G.J. Integrin alpha (IIb) beta (3) inhibitor preserves microvascular patency in experimental acute focal cerebral ischemia. Stroke 2000, 31, 1402–1409. [Google Scholar] [CrossRef] [Green Version]
- Xu, X.R.; Carrim, N.; Neves, M.A.; McKeown, T.; Stratton, T.W.; Coelho, R.M.; Lei, X.; Chen, P.; Xu, J.; Dai, X.; et al. Platelets and platelet adhesion molecules: Novel mechanisms of thrombosis and anti-thrombotic therapies. Thromb. J. 2016, 14, 29. [Google Scholar] [CrossRef] [Green Version]
- Tai, H.; Wei, Q.; Jin, Y.; Su, M.; Song, J.X.; Zhou, X.D.; Ouyang, H.M.; Wang, W.Y.; Xiong, Y.L.; Zhang, Y. TMVA, a snake C-type lectin-like protein from Trimeresurus mucrosquamatus venom, activates platelet via GPIb. Toxicon 2004, 44, 649–656. [Google Scholar] [CrossRef]
- Lu, Q.M.; Navdaev, A.; Clemetson, J.M.; Clemetson, K.J. GPIb is involved in platelet aggregation induced by mucetin, a snake C-type lectin protein from Chinese habu (Trimeresurus mucrosquamatus) venom. Thromb. Haemost. 2004, 91, 1168–1176. [Google Scholar] [CrossRef] [PubMed]
- Tsai, I.H.; Wang, Y.M. Effect of site directed mutagenesis on the activity of recombinant trimucrotoxin, a neurotoxic phospholipase from Trimeresurus mucrosquamatus venom. Toxicon 1998, 36, 1591–1597. [Google Scholar] [CrossRef]
- Tsai, I.H.; Wang, Y.M.; Chen, Y.H.; Tsai, T.S.; Tu, M.C. Venom phospholipases A2 of bamboo viper (Trimeresurus stejnegeri): Molecular characterization, geographic variations and evidence of multiple ancestries. Biochem. J. 2004, 377, 215–223. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harris, J.B. Phospholipases in Snake-Venoms and Their Effects on Nerve and Muscle. Pharm. Ther. 1985, 31, 79–102. [Google Scholar] [CrossRef]
- Montecucco, C.; Rossetto, O. How do presynaptic PLA (2) neurotoxins block nerve terminals? Trends Biochem. Sci. 2000, 25, 266–270. [Google Scholar] [CrossRef]
- Rigoni, M.; Caccin, P.; Gschmeissner, S.; Koster, G.; Postle, A.D.; Rossetto, O.; Schiavo, G.; Montecucco, C. Equivalent effects of snake PLA2 neurotoxins and lysophospholipid-fatty acid mixtures. Science 2005, 310, 1678–1680. [Google Scholar] [CrossRef] [PubMed]
- Gutierrez, J.M.; Escalante, T.; Rucavado, A.; Herrera, C.; Fox, J.W. A Comprehensive View of the Structural and Functional Alterations of Extracellular Matrix by Snake Venom Metalloproteinases (SVMPs): Novel Perspectives on the Pathophysiology of Envenoming. Toxins 2016, 8, 304. [Google Scholar] [CrossRef] [Green Version]
- Kakumanu, R.; Kemp-Harper, B.K.; Silva, A.; Kuruppu, S.; Isbister, G.K.; Hodgson, W.C. An in vivo examination of the differences between rapid cardiovascular collapse and prolonged hypotension induced by snake venom. Sci. Rep. 2019, 9, 1–9. [Google Scholar] [CrossRef]
- Koh, D.C.I.; Armugam, A.; Jeyaseelan, K. Snake venom components and their applications in biomedicine. Cell. Mol. Life Sci. 2006, 63, 3030–3041. [Google Scholar] [CrossRef]
- Clemetson, K.J.; Lu, Q.M.; Clemetson, J.M. Snake venom proteins affecting platelets and their applications to anti-thrombotic research. Curr. Pharm. Des. 2007, 13, 2887–2892. [Google Scholar] [CrossRef]
- Morgenstern, D.; King, G.F. The venom optimization hypothesis revisited. Toxicon 2013, 63, 120–128. [Google Scholar] [CrossRef] [PubMed]
- Shine, R. Habitats, Diets, and Sympatry in Snakes—Study from Australia. Can. J. Zool. 1977, 55, 1118–1128. [Google Scholar] [CrossRef]
- Colston, T.J.; Costa, G.C.; Vitt, L.J. Snake diets and the deep history hypothesis. Biol. J. Linn. Soc. 2010, 101, 476–486. [Google Scholar] [CrossRef]
- Zeng, X.; Hu, J.; Liang, X.; Wu, Y.; Yan, M.; Zhu, M.; Fu, Y. Acute cerebral infarction following a Trimeresurus stejnegeri snakebite: A case report. Medicine 2019, 98, e15684. [Google Scholar] [CrossRef] [PubMed]
- Boviatsis, E.J.; Kouyialis, A.T.; Papatheodorou, G.; Gavra, M.; Korfias, S.; Sakas, D.E. Multiple hemorrhagic brain infarcts after viper envenomation. Am. J. Trop. Med. Hyg. 2003, 68, 253–257. [Google Scholar] [CrossRef] [PubMed]
- Murthy, J.M.K.; Kishore, L.T.; Naidu, K.S. Cerebral infarction after envenomation by viper. J. Comput. Assist. Tomogr. 1997, 21, 35–37. [Google Scholar] [CrossRef]
- Jeevagan, V.; Chang, T.; Gnanathasan, C.A. Acute ischemic stroke following Hump-nosed viper envenoming; first authenticated case. Thromb. J. 2012, 10, 21. [Google Scholar] [CrossRef] [Green Version]
- Panicker, J.N.; Madhusudanan, S. Cerebral infarction in a young male following viper envenomation. J. Assoc. Physicians India 2000, 48, 744–745. [Google Scholar]
- Malbranque, S.; Piercecchi-Marti, M.D.; Thomas, L.; Barbey, C.; Courcier, D.; Bucher, B.; Ridarch, A.; Smadja, D.; Warrell, D.A. Fatal diffuse thrombotic microangiopathy after a bite by the “Fer-de-Lance” pit viper (Bothrops lanceolatus) of Martinique. Am. J. Trop. Med. Hyg. 2008, 78, 856–861. [Google Scholar] [CrossRef] [Green Version]
- Walsh, R.N.; Cummins, R.A. The Open-Field Test: A critical review. Psychol. Bull. 1976, 83, 482–504. [Google Scholar] [CrossRef]
- Duan, T.T.; Tan, J.W.; Yuan, Q.; Cao, J.; Zhou, Q.X.; Xu, L. Acute ketamine induces hippocampal synaptic depression and spatial memory impairment through dopamine D1/D5 receptors. Psychopharmacology 2013, 228, 451–461. [Google Scholar] [CrossRef] [PubMed]
- Scott, T.; Owens, M.D. Thrombocytes respond to lipopolysaccharide through toll-like receptor-4, and map kinase and NF-kappa b pathways leading to expression of interleukin-6 and cyclooxygenase-2 with production of prostaglandin E2. Mol. Immunol. 2008, 45, 1001–1008. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Prey Species | Crude Venom | Percentage to Lose Righting Reflex (%) | ||
|---|---|---|---|---|
| (2 mg/kg) | 2.5 min | 5 min | 10 min | |
| Pheasant chicks | P. mucrosquamatus | 5% | 30% | 75% |
| (P. colchicus) | T. stejnegeri | 10% | 40% | 90% |
| N = 20 per group | N.atra | 5% | 40% | 85% |
| Adult mice | P. mucrosquamatus | 30% | 70% | 100% |
| (M. musculus) | T. stejnegeri | 40% | 85% | 100% |
| N = 20 per group | N. atra | 35% | 80% | 100% |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tian, H.; Liu, M.; Li, J.; Xu, R.; Long, C.; Li, H.; Mwangi, J.; Lu, Q.; Lai, R.; Shen, C. Snake C-Type Lectins Potentially Contribute to the Prey Immobilization in Protobothrops mucrosquamatus and Trimeresurus stejnegeri Venoms. Toxins 2020, 12, 105. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins12020105
Tian H, Liu M, Li J, Xu R, Long C, Li H, Mwangi J, Lu Q, Lai R, Shen C. Snake C-Type Lectins Potentially Contribute to the Prey Immobilization in Protobothrops mucrosquamatus and Trimeresurus stejnegeri Venoms. Toxins. 2020; 12(2):105. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins12020105
Chicago/Turabian StyleTian, Huiwen, Ming Liu, Jiameng Li, Runjia Xu, Chengbo Long, Hao Li, James Mwangi, Qiumin Lu, Ren Lai, and Chuanbin Shen. 2020. "Snake C-Type Lectins Potentially Contribute to the Prey Immobilization in Protobothrops mucrosquamatus and Trimeresurus stejnegeri Venoms" Toxins 12, no. 2: 105. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins12020105