Study of the Bacillus thuringiensis Cry1Ia Protein Oligomerization Promoted by Midgut Brush Border Membrane Vesicles of Lepidopteran and Coleopteran Insects, or Cultured Insect Cells

 ,
,  and
and
Abstract
:1. Introduction
2. Results
2.1. Toxicity of Cry1Ia and Cry1Ab against Lepidopteran and Coleopteran Species
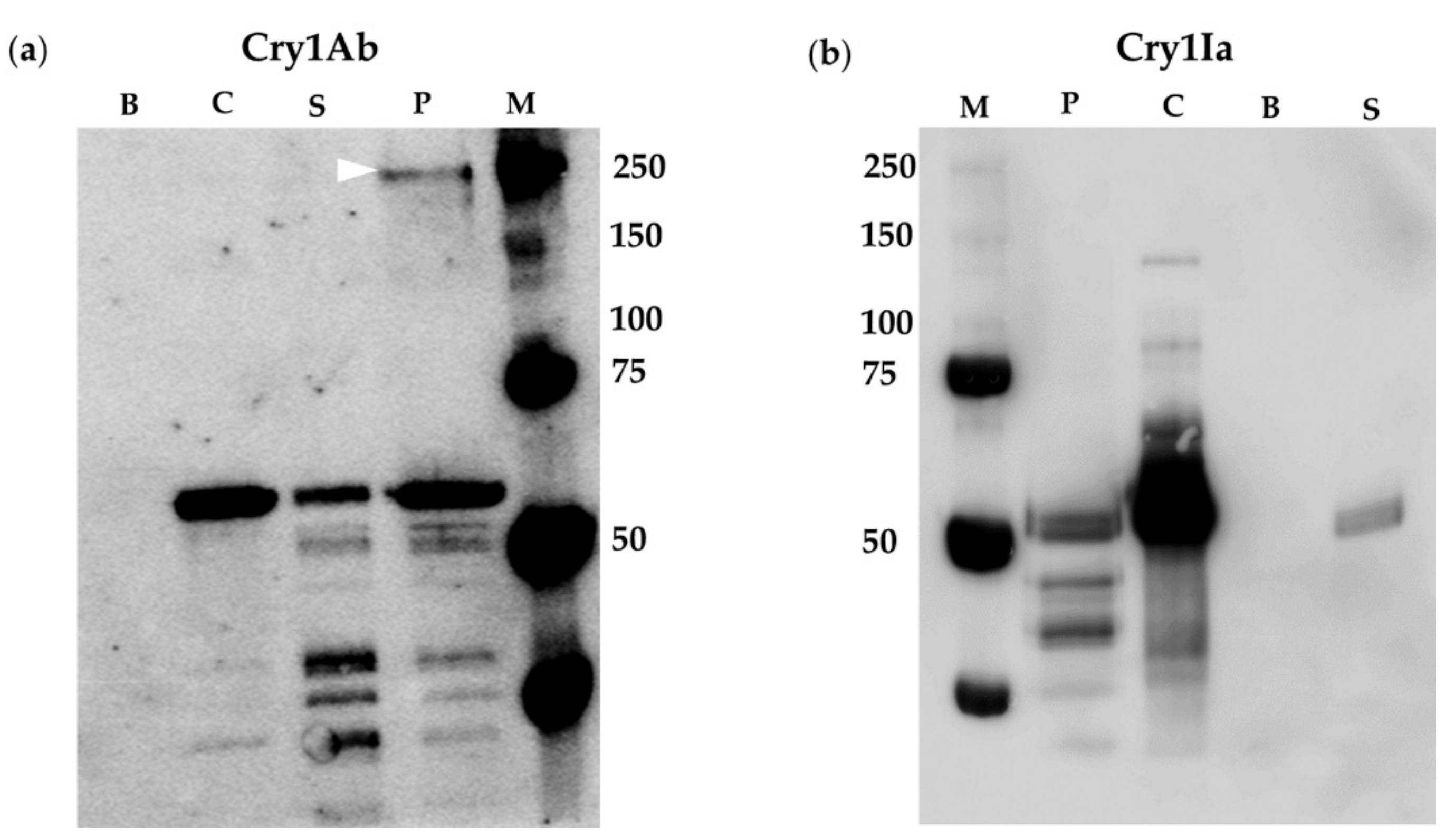
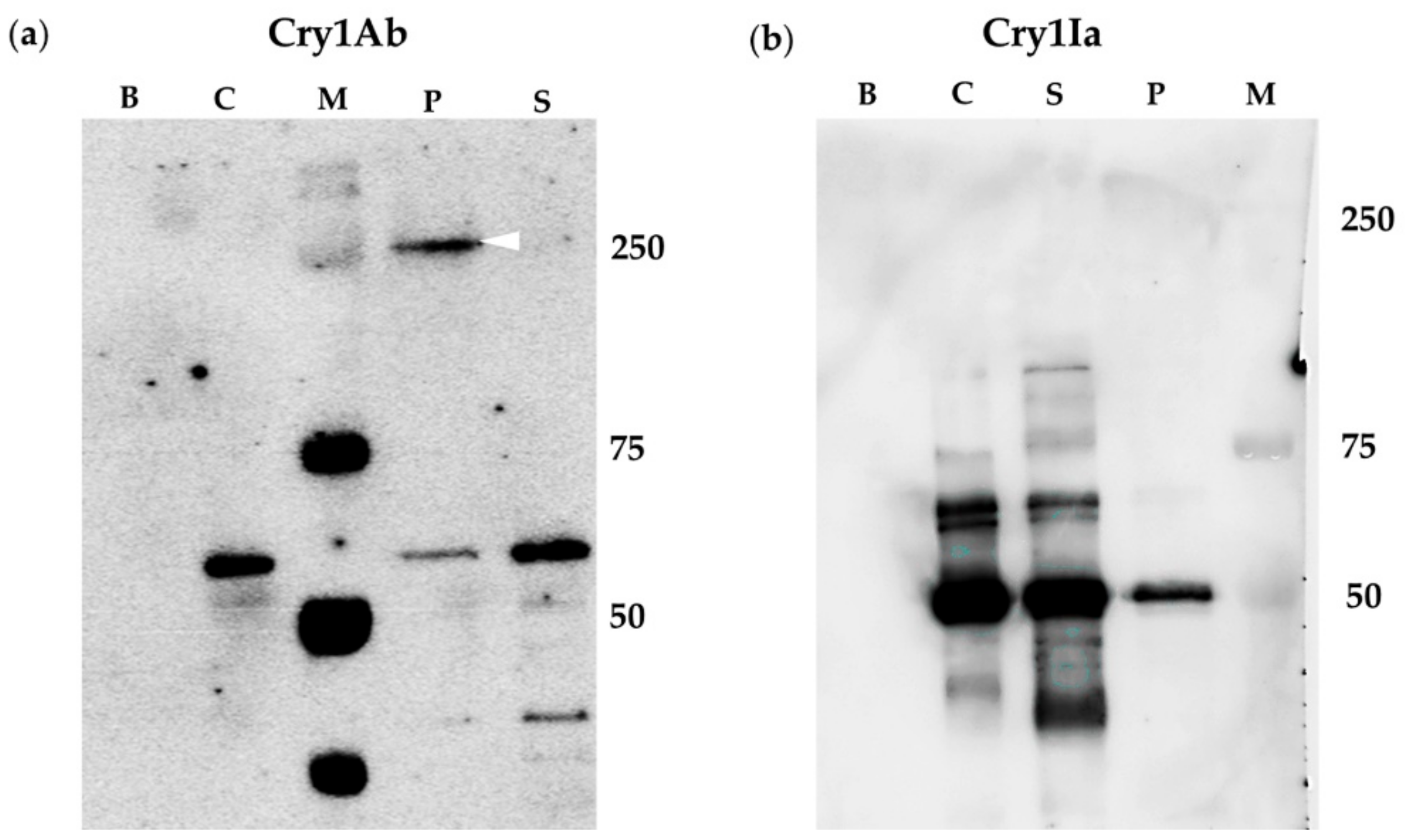
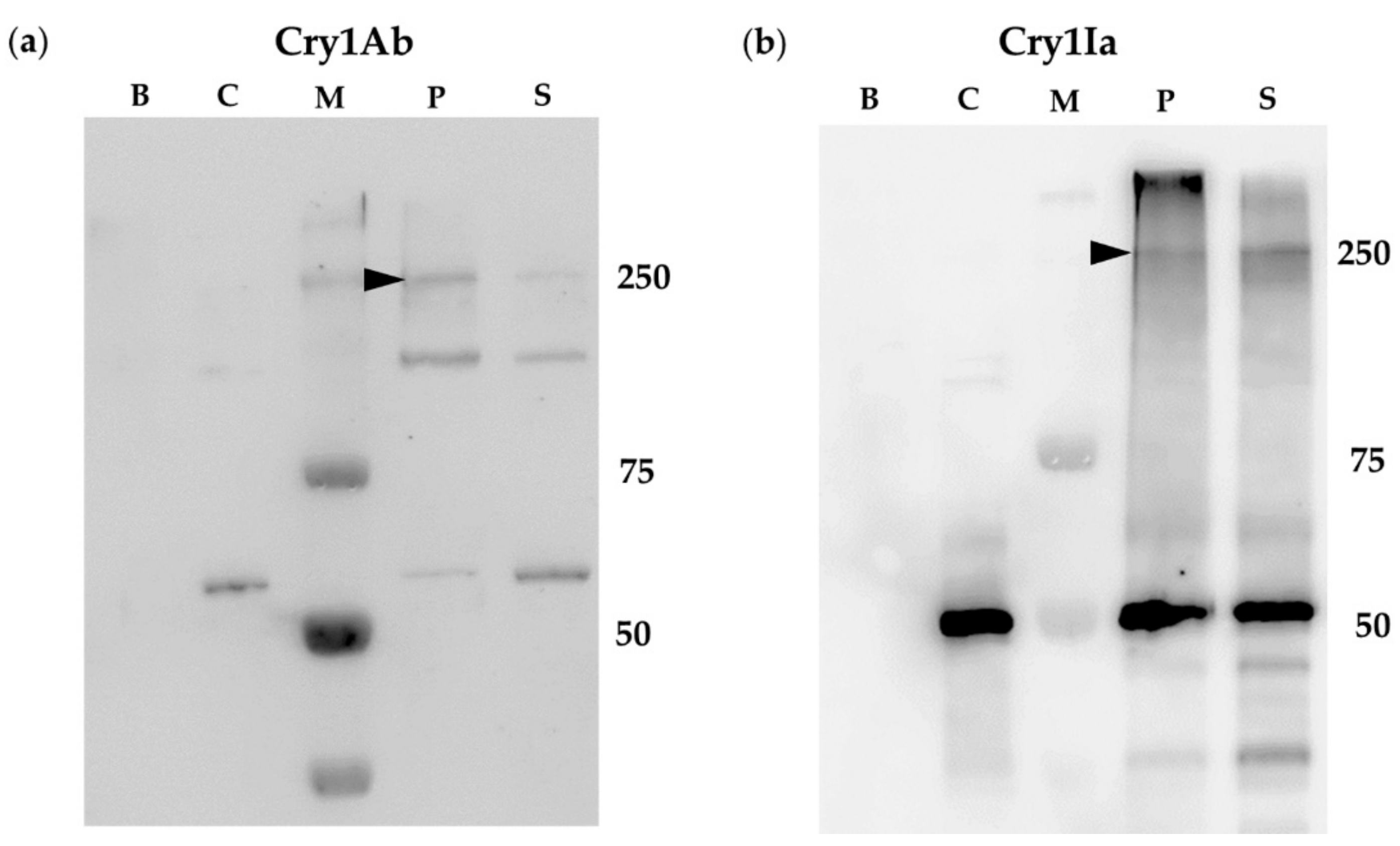
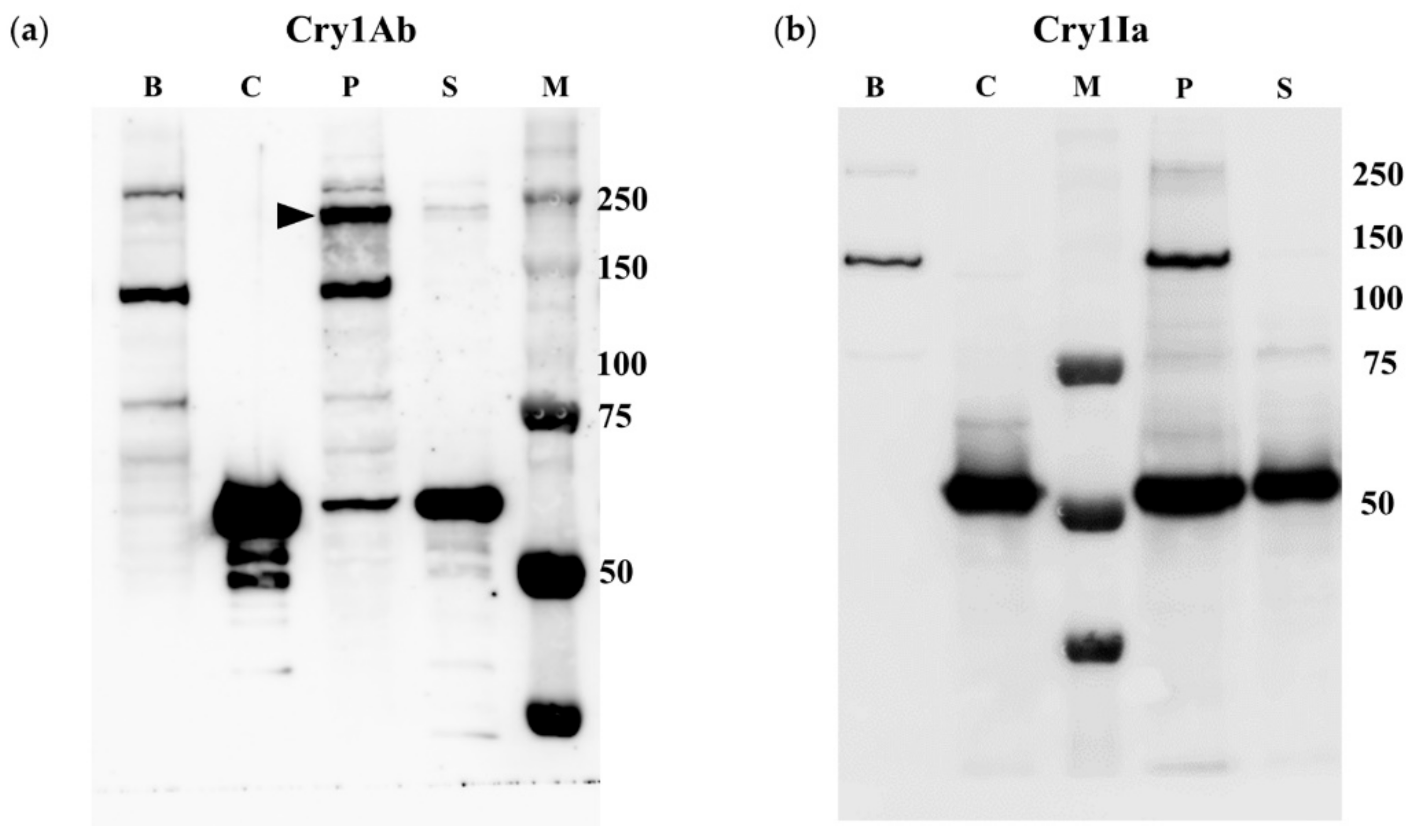
2.2. BBMV of Susceptible Lepidopteran Insects Promoted Oligomerization of Cry1Ab But Not of Cry1Ia
2.3. Oligomerization of Cry1Ia Was Promoted by BBMV from L. decemlineata
2.4. Oligomerization Promoted by Sf21 Insect Cells
3. Discussion
4. Materials and Methods
4.1. Production and Purification of Cry Proteins
4.2. Insect Rearing
4.3. Insect Cell Line
4.4. Insect Bioassays
4.5. Midgut Isolation and BBMV Preparation
4.6. Biotin Labelling
4.7. Oligomerization Assays with Sf21 Cells
4.8. Oligomerization Assays with BBMV
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Van Frankenhuyzen, K. Insecticidal activity of Bacillus thuringiensis crystal proteins. J. Invertebr. Pathol. 2009, 101, 1–16. [Google Scholar] [CrossRef]
- Ohba, M.; Mizuki, E.; Uemori, A. Parasporin, a new anticancer protein group from Bacillus thuringiensis. Anticancer Res. 2009, 29, 427–433. [Google Scholar] [PubMed]
- Palma, L.; Muñoz, D.; Berry, C.; Murillo, J.; Caballero, P. Bacillus thuringiensis toxins: An overview of their biocidal activity. Toxins 2014, 6, 3296–3325. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chakroun, M.; Banyuls, N.; Bel, Y.; Escriche, B.; Ferré, J. Bacterial vegetative insecticidal proteins (Vip) from entomophatogenic bacteria. Microbiol. Mol. Biol. Rev. 2016, 80, 329–350. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kostichka, K.; Warren, G.W.; Mullins, M.; Mullins, A.D.; Craig, J.A.; Koziel, M.G.; Estruch, A.D. Cloning of a cryV-type insecticidal protein gene from Bacillus thuringiensis: The cryV-encoded protein is expressed early in stationary phase. J. Bacteriol. 1996, 178, 2141–2144. [Google Scholar] [CrossRef] [Green Version]
- Jurat-Fuentes, J.L.; Jackson, T.A. Bacterial entomopathoges. In Insect Pathology, 2nd ed.; Vega, F.E., Kaya, H.K., Eds.; Academic Press: San Diego, CA, USA, 2012; pp. 265–349. [Google Scholar] [CrossRef]
- Hoffman, C.; Vanderbruggen, H.; Höfte, H.; Van Rie, G.; Jansens, S.; Van Mellaert, H. Specificity of Bacillus thuringiensis delta-endotoxins is correlated with the presence of high-affinity binding sites in the brush border membrane of target insect midguts. Proc. Natl. Acad. Sci. USA 1988, 85, 7844–7848. [Google Scholar] [CrossRef] [Green Version]
- Ferre, J.; VanRie, J. Biochemistry and genetics of insect resistance to Bacillus thuringiensis. Annu. Rev. Entomol. 2002, 47, 501–533. [Google Scholar] [CrossRef]
- Zhang, X.; Candas, M.; Griko, N.B.; Taussig, R.; Bulla, L.A., Jr. A mechanism of cell death involving an adenylyl cyclase/PKA signaling pathway is induced by the Cry1Ab toxin of Bacillus thuringiensis. Proc. Natl. Acad. Sci. USA 2006, 103, 9897–9902. [Google Scholar] [CrossRef] [Green Version]
- Adang, M.J.; Crickmore, N.; Jurat-Fuentes, J.L. Diversity of Bacillus thuringiensis crystal toxins and mechanism of action. Adv. Insect Physiol. 2014, 47, 39–87. [Google Scholar] [CrossRef]
- Arenas, I.; Bravo, A.; Soberón, M.; Gómez, I. Role of alkaline phosphatase from Manduca sexta in the mechanism of action of Bacillus thuringiensis Cry1Ab toxin. J. Biol. Chem. 2010, 285, 12497–12503. [Google Scholar] [CrossRef] [Green Version]
- Vachon, V.; Raynald, L.; Schwartz, J.L. Current models of the mode of action of Bacillus thuringiensis insecticidal crystal proteins: A critical review. J. Invertebr. Pathol. 2012, 111, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Pardo-López, L.; Soberón, M.; Bravo, A. Bacillus thuringiensis insecticidal three-domain Cry toxins: Mode of action, insect resistance and consequences for crop protection. FEMS Microbiol. Rev. 2013, 37, 3–22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gomez, I.; Sanchez, J.; Miranda, R.; Bravo, A.; Soberon, M. Cadherin-like receptor binding facilitates proteolytic cleavage of helix alpha-1 in domain I and oligomer pre-pore formation of Bacillus thuringiensis Cry1Ab toxin. FEBS Lett. 2002, 513, 242–246. [Google Scholar] [CrossRef] [Green Version]
- Rausell, C.; García-Robles, I.; Sánchez, J.; Muñoz-Garay, C.; Martínez-Ramírez, A.C.; Real, M.D.; Bravo, A. Role of toxin activation on binding and pore formation activity of the Bacillus thuringiensis Cry3 toxins in membranes of Leptinotarsa decemlineata (Say). Biochim. Biophys. Acta 2004, 1660, 99–105. [Google Scholar] [CrossRef] [Green Version]
- Muñoz-Garay, C.; Portugal, L.; Pardo-López, L.; Jiménez-Juárez, N.; Arenas, I.; Gómez, I.; Sánchez-López, R.; Arroyo, R.; Holzenburg, A.; Savva, C.G.; et al. Characterization of the mechanism of action of the genetically modified Cry1AbMod toxin that is active against Cry1Ab-resistant insects. Biochim. Biophys. Acta 2009, 1788, 2229–2237. [Google Scholar] [CrossRef] [Green Version]
- Tigue, N.; Jacoby, J.; Ellar, D.J. The α-helix residue, Asn135, is involved in the oligomerization of Cry1Ac1 and Cry1Ab5 Bacillus thuringiensis toxins. Appl. Environ. Microbiol. 2001, 67, 5715–5720. [Google Scholar] [CrossRef] [Green Version]
- Herrero, S.; González-Cabrera, J.; Ferré, J.; Bakker, P.L.; de Maagd, R.A. Mutations in the Bacillus thuringiensis Cry1Ca toxin demonstrate the role of domains II and III in specificity towards Spodoptera exigua larvae. Biochem. J. 2004, 384, 507–513. [Google Scholar] [CrossRef] [Green Version]
- Pérez, C.; Muñoz-Garay, C.; Portugal, L.; Sánchez, J.; Gill, S.S.; Soberón, M.; Bravo, A. Bacillus thuringiensis spp. israelensis Cyt1Aa enhances activity of Cry11Aa toxin by facilitating the formation of a pre-pore oligomeric structure. Cell Microbiol. 2007, 9, 2931–2937. [Google Scholar] [CrossRef] [Green Version]
- Fabrick, J.; Oppert, C.; Lorenzen, M.D.; Morris, K.; Oppert, B.; Jurat-Fuentes, J.L. A novel Tenebrio molitor cadherin is a functional receptor for Bacillus thuringiensis Cry3Aa toxin. J. Biol. Chem. 2009, 284, 18401–18410. [Google Scholar] [CrossRef] [Green Version]
- Groulx, N.; McGuire, H.; Laprade, R.; Schwartz, J.L.; Blunck, R. Single molecule fluorescence study of the Bacillus thuringiensis toxin Cry1Aa reveals tetramerization. J. Biol. Chem. 2011, 286, 42274–42282. [Google Scholar] [CrossRef] [Green Version]
- Carmona, D.; Rodríguez-Almazán, C.; Muñoz-Garay, C.; Portugal, L.; Pérez, C.; de Maagd, R.A.; Bakker, P.; Soberón, M.; Bravo, A. Dominant negative phenotype of Bacillus thuringiensis Cry1Ab, Cry11Aa and Cry4Ba mutants suggest hetero-oligomer formation among different Cry toxins. PLoS ONE 2011, 6, e19952. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khomkhum, N.; Leetachewa, S.; Angsuthanasombat, C.; Moonsom, S. Functional assembly of 260-kDa oligomers required for mosquito-larvicidal activity of the Bacillus thuringiensis Cry4Ba toxin. Peptides 2015, 68, 183–189. [Google Scholar] [CrossRef] [PubMed]
- Xu, L.; Pan, Z.; Zhang, J.; Niu, L.; Li, J.; Chen, Z.; Liu, B.; Zhu, Y.; Chen, Q. Exposures of helices α4 and α5 is required for insecticidal activity of Cry2Ab by prompting assembly of prepore oligomeric structure. Cell. Microbiol. 2018, e12827. [Google Scholar] [CrossRef] [PubMed]
- Abe, Y.; Inoue, H.; Ashida, H.; Maeda, Y.; Kinoshita, T.; Kitada, S. Glycan region of GPI anchored-protein is required for cytocidal oligomerization of an anticancer parasporin-2, Cry46Aa1 protein, from Bacillus thuringiensis strain A1547. J. Invertebr. Pathol. 2017, 142, 71–81. [Google Scholar] [CrossRef]
- López-Diaz, J.; Cantón, P.E.; Gill, S.S.; Soberón, M.; Bravo, A. Oligomerization is a key step in Cyt1Aa membrane insertion and toxicity but not necessary to synergize Cry11Aa toxicity in Aedes aegypti larvae. Environ. Microbiol. 2013, 15, 3030–3039. [Google Scholar] [CrossRef] [Green Version]
- Palma, L.; Scott, D.J.; Harris, G.; Din, S.; Williams, T.L.; Reoberts, O.J.; Young, M.T.; Caballero, P.; Berry, C. The Vip3Ag4 insecticidal protoxins from Bacillus thuringiensis adopts a tetrameric configuration that is maintained on proteolysis. Toxins 2017, 9, 165. [Google Scholar] [CrossRef] [PubMed]
- Guo, S.; Zhang, Y.; Song, F.; Zhang, J.; Huang, D. Protease-resistant core form of Bacillus thuringiensis Cry1Ie: Monomeric and oligomeric forms in solution. Biotechnol. Lett. 2009, 31, 1769–1774. [Google Scholar] [CrossRef]
- Tailor, R.; Tippett, J.; Gibb, G.; Pells, S.; Pike, D.; Jordan, L.; Ely, S. Identification and characterization of a novel Bacillus thuringiensis delta-endotoxin entomocidal to coleopteran and lepidopteran larvae. Mol. Microbiol. 1992, 6, 1211–1217. [Google Scholar] [CrossRef]
- Choi, S.K.; Shin, B.S.; Kong, E.M.; Rho, H.M.; Park, S.H. Cloning of a new Bacillus thuringiensis cry1I-type crystal protein gene. Curr. Microbiol. 2000, 41, 65–69. [Google Scholar] [CrossRef]
- Xu, L.; Ferry, N.; Wang, Z.; Zhang, J.; Edwards, M.G.; Gatehouse, A.M.R.; He, K. A proteomic approach to study the mechanism of tolerance to Bt toxins in Ostrinia furnacalis larvae selected for resistance to Cry1Ab. Transgenic Res. 2013, 22, 1155–1166. [Google Scholar] [CrossRef]
- Zhang, T.; He, M.; Gatehouse, A.M.R.; Wang, Z.; Edwards, M.G.; Li, Q.; He, K. Inheritance patterns, dominance and cross-resistance of Cry1Ab- and Cry1Ac-selected Ostrinia furnacalis (Guenée). Toxins 2014, 6, 2694–2707. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, L.; Wang, Z.; Zhang, J.; He, K.; Ferry, N.; Gatehouse, A.M.R. Cross-resistance of Cry1Ab-selected Asian corn borer to other Cry toxins. J. Appl. Entomol. 2010, 134, 429–438. [Google Scholar] [CrossRef]
- Guo, J.; He, K.; Hellmich, R.L.; Bai, S.; Zhang, T.; Liu, Y.; Ahmed, T.; Wang, Z. Field trials to evaluate the effects of transgenic cry1Ie maize on the community characteristics of arthropod natural enemies. Sci. Rep. 2016, 6, 22102. [Google Scholar] [CrossRef] [PubMed]
- Fan, C.; Wu, F.; Dong, J.; Wang, B.; Yin, J.; Song, X. No impact of transgenic cry1Ie maize on the diversity, abundance and composition of soil fauna in a 2-year field trial. Sci. Rep. 2019, 9, 10333. [Google Scholar] [CrossRef] [Green Version]
- Rausell, C.; Muñoz-Garay, C.; Miranda-CasoLuengo, R.; Gómez, I.; Rudiño-Piñera, E.; Soberón, M.; Bravo, A. Tryptophan spectroscopy studies and black lipid bilayer analysis indicate that the oligomeric structure of Cry1Ab toxin from Bacillus thuringiensis is the membrane-insertion intermediate. Biochem. J. 2004, 43, 166–174. [Google Scholar] [CrossRef]
- Gómez, I.; Sánchez, J.; Muñoz-Garay, C.; Matus, V.; Gill, S.S.; Soberón, M.; Bravo, A. Bacillus thuringiensis Cry1A toxins are versatile proteins with multiple modes of action: Two distinct pre-pores are involved in toxicity. Biochem. J. 2014, 459, 383–396. [Google Scholar] [CrossRef] [Green Version]
- Portugal, L.; Gringorten, J.L.; Caputo, G.F.; Soberón, M.; Muñoz-Garay, C.; Bravo, A. Toxicity and mode of action of insecticidal Cry1A proteins from Bacillus thuringiensis in an insect cell line, CF-1. Peptides 2014, 53, 292–299. [Google Scholar] [CrossRef]
- Nair, M.S.; Dean, D.H. Composition of the putative prepore complex of Bacillus thuringiensis Cry1Ab toxin. Adv. Biol. Chem. 2015, 5, 179–188. [Google Scholar] [CrossRef] [Green Version]
- Khorramnejad, A.; Bel, Y.; Hernández-Martínez, P.; Talaei-Hassanloui, R.; Escriche, B. Insecticidal activity and cytotoxicity of Bacillus thuringiensis Cry1Ia7 protein. In GMOs in Integrated Plant Protection; Meissle, M., De Schrijver, A., Smagghe, G., Eds.; IOBC-WPRS Bulletin: Ghent, Belgium, 2018; Volume 131, pp. 56–63. [Google Scholar]
- Martínez-Solis, M.; Pinos, D.; Endo, H.; Portugal, L.; Sato, R.; Ferré, F.; Herrero, S.; Hernández-Martínez, P. Role of Bacillus thuringiensis Cry1A toxins domains in the binding to the ABCC2 receptor from Spodoptera exigua. Insect. Biochem. Mol. Biol. 2018, 101, 47–56. [Google Scholar] [CrossRef]
- Crickmore, N.; Zeigler, D.R.; Feitelson, J.; Schnepf, E.; Van Rie, J.; Lereclus, D.; Baum, J.; Dean, D.H. Revision of the nomenclature for the Bacillus thuringiensis pesticidal crystal proteins. Microbiol. Mol. Biol. Rev. 1998, 62, 807–813. [Google Scholar] [CrossRef] [Green Version]
- Masson, L.; Erlandson, M.; Puzstai-Carey, M.; Brousseau, R.; Juárez-Pérez, V.; Frutos, R. A holistic approach for determining the entomopathogenic potential of Bacillus thuringiensis strains. Appl. Environ. Microbiol. 1998, 46, 4782–4788. [Google Scholar] [CrossRef] [Green Version]
- Ruiz de Escudero, I.; Estela, A.; Porcar, M.; Martínez, C.; Oguiza, J.A.; Escriche, B.; Ferré, J.; Caballero, P. Molecular and insecticidal characterization of a Cry1I protein toxic to insects of the families Noctuidae, Tortricidae, Plutellidae and Chrysomelidae. Appl. Environ. Microbiol. 2006, 72, 4796–4804. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiménez-Juárez, N.; Muñoz-Garay, C.; Gómez, I.; Saab-Rincon, G.; Damian-Almazo, J.Y.; Gill, S.S.; Soberón, M.; Bravo, A. Bacillus thuringiensis Cry1Ab mutants affecting oligomer formation are not-toxic to Manduca sexta larvae. J. Biol. Chem. 2007, 282, 21222–21229. [Google Scholar] [CrossRef] [Green Version]
- Likitvivatanavong, S.; Katzenmeier, G.; Andsuthanasombat, C. Asn183 in α5 is essential for oligomerization and toxicity of the Bacillus thuringiensis Cry4Ba toxin. Arch. Biochem. Biophys. 2005, 445, 46–55. [Google Scholar] [CrossRef]
- Rodríguez-Almazán, C.; Zavala, L.E.; Muñoz-Garay, C.; Jiménez-Juárez, N.; Pacheco, S.; Masson, L.; Soberón, M.; Bravo, A. Dominant negative mutants of Bacillus thuringiensis Cry1Ab toxin function as anti-toxins: Demonstration of the role of oligomerization in toxicity. PLoS ONE 2009, 4, e5545. [Google Scholar] [CrossRef] [PubMed]
- Jurat Fuentes, J.L.; Adang, M.J. Cry toxin mode of action in susceptible and resistant Heliothis virescens larvae. J. Invertebr. Pathol. 2006, 92, 166–171. [Google Scholar] [CrossRef]
- Sekar, V.; Held, B.; Tippet, J.; Amirhusin, B.; Robeff, P.; Wang, K.; Wilson, H.M. Biochemical and Molecular Characterization of the insecticidal fragment of CryV. Appl. Environ. Microbiol. 1997, 63, 2798–2801. [Google Scholar] [CrossRef] [Green Version]
- Crava, C.M.; Bel, Y.; Escriche, B. Bacillus thuringiensis susceptibility variation among Ostrinia nubilalis populations. In Future Research and Development in the Use of Microbial Agents and Nematodes for Biological Insect Control; Ehlers, R.U., Crickmore, N., Enkerli, J., Glazer, I., Lopez-Ferber, M., Tkaczuk, C., Eds.; IOBC-WPRS Bulletin: Pamplona, Spain, 2009; Volume 45, pp. 171–174. [Google Scholar]
- Ocelotl, J.; Sánchez, J.; Arroyo, R.; Gracía-Gómez, B.I.; Gómez, I.; Unnithan, G.C.; Tabashnik, B.E.; Bravo, A.; Soberón, M. Binding and oligomerization of modified and native Bt toxins in resistance and susceptible pink bollworm. PLoS ONE 2015, 10. [Google Scholar] [CrossRef]
- Ocelotl, J.; Sánchez, J.; Gómez, I.; Tabashnik, B.E.; Bravo, A.; Soberón, M. ABCC2 is associated with Bacillus thuringiensis Cry1Ac toxin oligomerization and membrane insertion in diamondback moth. Sci. Rep. 2017, 7, 2386. [Google Scholar] [CrossRef] [Green Version]
- Padilla, C.; Pardo-López, L.; de la Riva, G.; Gómez, I.; Sánchez, J.; Hernandez, G.; Nuñez, M.E.; Carey, M.P.; Dean, D.H.; Alzate, O.; et al. Role of tryptophan residues in toxicity of Cry1Ab toxin from Bacillus thuringiensis. Appl. Environ. Microbiol. 2006, 72, 901–907. [Google Scholar] [CrossRef] [Green Version]
- Chakroun, M.; Sellami, S.; Ferré, J.; Tounsi, S.; Rouis, S. Ephestia kuehniella tolerance to Bacillus thuringiensis Cry1Aa associated with reduced oligomer formation. Biochem. Biophys. Res. Commun. 2017, 482, 808–813. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-Almazán, C.; Reyes, E.Z.; Zúñiga-Navarrete, F.; Muñoz-Garay, C.; Gómez, I.; Evans, A.M.; Likitvivatanavong, S.; Bravo, A.; Gill, S.S.; Soberón, M. Cadherin binding is not a limiting step for Bacillus thuringiensis subsp. israelensis Cry4Ba toxicity to Aedes aegypti larvae. Biochem. J. 2012, 443, 711–717. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sayyed, A.H.; Gatsi, R.; Ibiza-Palacios, M.; Escriche, B.; Wright, D.J.; Crickmore, N. Common, but complex, mode of resistance of Plutella xylostella to Bacillus thuringiensis toxins Cry1Ab and Cry1Ac. Appl. Environ. Microbiol. 2005, 11, 6863–6868. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Crava, C.M.; Bel, Y.; Jakubowska, A.K.; Ferré, J.; Escriche, B. Midgut aminopeptidase N isoforms from Ostrinia nubilalis: Activity characterization and differential binding to Cty1Ab and Cry1Fa proteins from Bacillus thuringiensis. Insect. Biochem. Mol. Biol. 2013, 43, 924–935. [Google Scholar] [CrossRef] [PubMed]
- Stewart, G.S.; Johnstone, K.; Hagelberg, E.; Ellar, D.J. Commitment of bacterial spores to germinate. A measure of the trigger reaction. Biochem. J. 1981, 196, 101–106. [Google Scholar] [CrossRef] [PubMed]
- Bradford, M.M. A rapid and sensitive method for the quantification of microgram quantities of protein using the principle of dye-binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- MacIntosh, S.C.; Stone, T.B.; Sims, S.R.; Hunst, P.L.; Greenplate, J.T.; Marrone, P.G.; Perlak, F.J.; Fischhoff, D.A.; Fuchs, R.L. Specificity and efficacy of purified Bacillus thuringiensis proteins against agronomically important insects. J. Invertebr. Pathol. 1990, 56, 258–266. [Google Scholar] [CrossRef]
- Beegle, C.C. Bioassay methods for quantification of Bacillus thuringiensis δ-endotoxin. In Analytical Chemistry of Bacillus Thuringiensis, 1st ed.; Hickle, L.A., Fitch, W.L., Eds.; American Chemical Society: Washington, DC, USA, 1990; pp. 14–21. [Google Scholar] [CrossRef]
- Iriarte, J.; Bel, Y.; Ferrandis, M.D.; Andrew, R.; Murillo, J.; Ferré, J.; Caballero, P. Environmental distribution and diversity of Bacillus thuringiensis in Spain. Syst. Appl. Microbiol. 1998, 21, 97–106. [Google Scholar] [CrossRef]
- Finney, D.J. Probit Analysis, 3rd ed.; Cambridge University Press: Cambridge, UK, 1971. [Google Scholar] [CrossRef]
- Wolfersberger, M.G.; Luethy, P.; Maurer, A.; Parenti, P.; Sacchi, F.V.; Giordana, B.; Hanozet, G.M. Preparation and partial characterization of amino acid transporting brush border membrane vesicles from the larval midgut of the cabbage butterfly (Pieris brassicae). Comp. Biochem. Physiol. A Physiol. 1987, 86, 301–308. [Google Scholar] [CrossRef]
- Escriche, B.; Silva, F.S.; Ferré, J. Testing suitability of brush border membrane vesicles prepared from whole larvae from small insects for binding studies with Bacillus thuringiensis CrylA(b) crystal protein. J. Invertebr. Pathol. 1995, 65, 318–320. [Google Scholar] [CrossRef]
- Hernández-Martínez, P.; Vera-Velasco, N.M.; Martínez-Solís, M.; Ghislain, M.; Ferré, J.; Escriche, B. Shared binding sites for the Bacillus thuringiensis proteins Cry3Bb, Cry3Ca, and Cry7Aa in the African sweet potato pest Cylas puncticollis (Brentidae). Appl. Environ. Microbiol. 2014, 80, 7545–7550. [Google Scholar] [CrossRef] [PubMed] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Insect Species | Bt Toxins | LC50 | Fiducial Limits (95%) | Regression Line | ||
|---|---|---|---|---|---|---|
| Lower | Upper | Slope ± SE | a ± SE | |||
| L. botrana | Cry1Ab | 153 | 106 | 217 | 1.15 ± 0.13 | 2.48 ± 0.3 |
| Cry1Ia | 80 | 56 | 108 | 1.28 ± 0.13 | 2.56 ± 0.31 | |
| O. nubilalis | Cry1Ab | 69 | 47 | 97 | 1.06 ± 0.11 | 3.04 ± 0.23 |
| Cry1Ia | 273 | 88 | 1011 | 1.50 ± 0.15 | 1.34 ± 0.14 | |
| L. decemlineata | Cry1Ab | NT | - | - | - | - |
| Cry1Ia | 22 | 12 | 53 | 0.51 ± 0.07 | 4.29 ± 0.09 | |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Khorramnejad, A.; Domínguez-Arrizabalaga, M.; Caballero, P.; Escriche, B.; Bel, Y. Study of the Bacillus thuringiensis Cry1Ia Protein Oligomerization Promoted by Midgut Brush Border Membrane Vesicles of Lepidopteran and Coleopteran Insects, or Cultured Insect Cells. Toxins 2020, 12, 133. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins12020133
Khorramnejad A, Domínguez-Arrizabalaga M, Caballero P, Escriche B, Bel Y. Study of the Bacillus thuringiensis Cry1Ia Protein Oligomerization Promoted by Midgut Brush Border Membrane Vesicles of Lepidopteran and Coleopteran Insects, or Cultured Insect Cells. Toxins. 2020; 12(2):133. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins12020133
Chicago/Turabian StyleKhorramnejad, Ayda, Mikel Domínguez-Arrizabalaga, Primitivo Caballero, Baltasar Escriche, and Yolanda Bel. 2020. "Study of the Bacillus thuringiensis Cry1Ia Protein Oligomerization Promoted by Midgut Brush Border Membrane Vesicles of Lepidopteran and Coleopteran Insects, or Cultured Insect Cells" Toxins 12, no. 2: 133. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins12020133