A Lateral Flow Strip Based on a Truncated Aptamer-Complementary Strand for Detection of Type-B Aflatoxins in Nuts and Dried Figs


Abstract
:1. Introduction
2. Results and Discussion
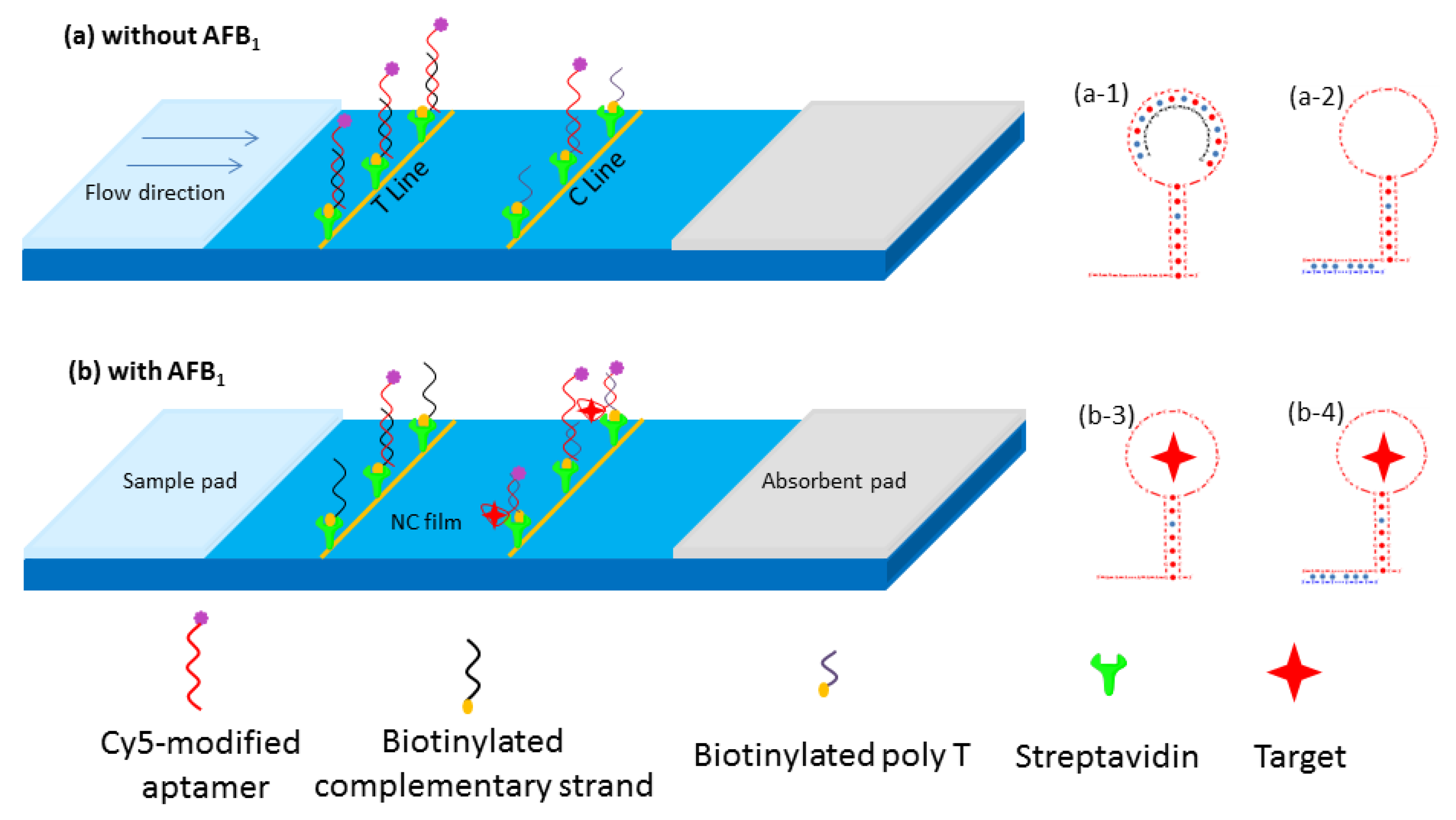
2.1. Principles for the Aptamer-Complementary Strand-Based Lateral Flow Test Strip
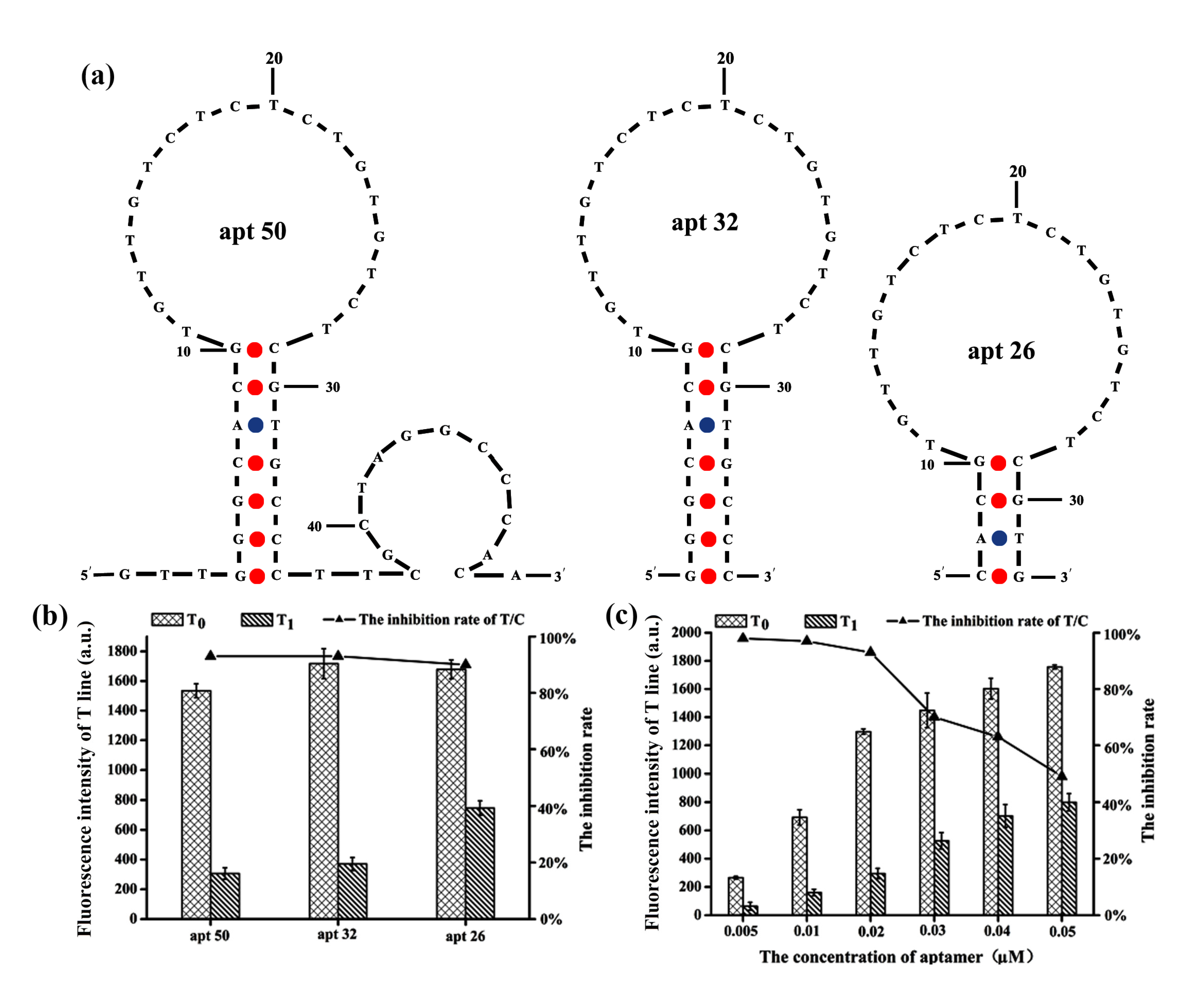
2.2. Optimization of the Aptamer Length and Concentration
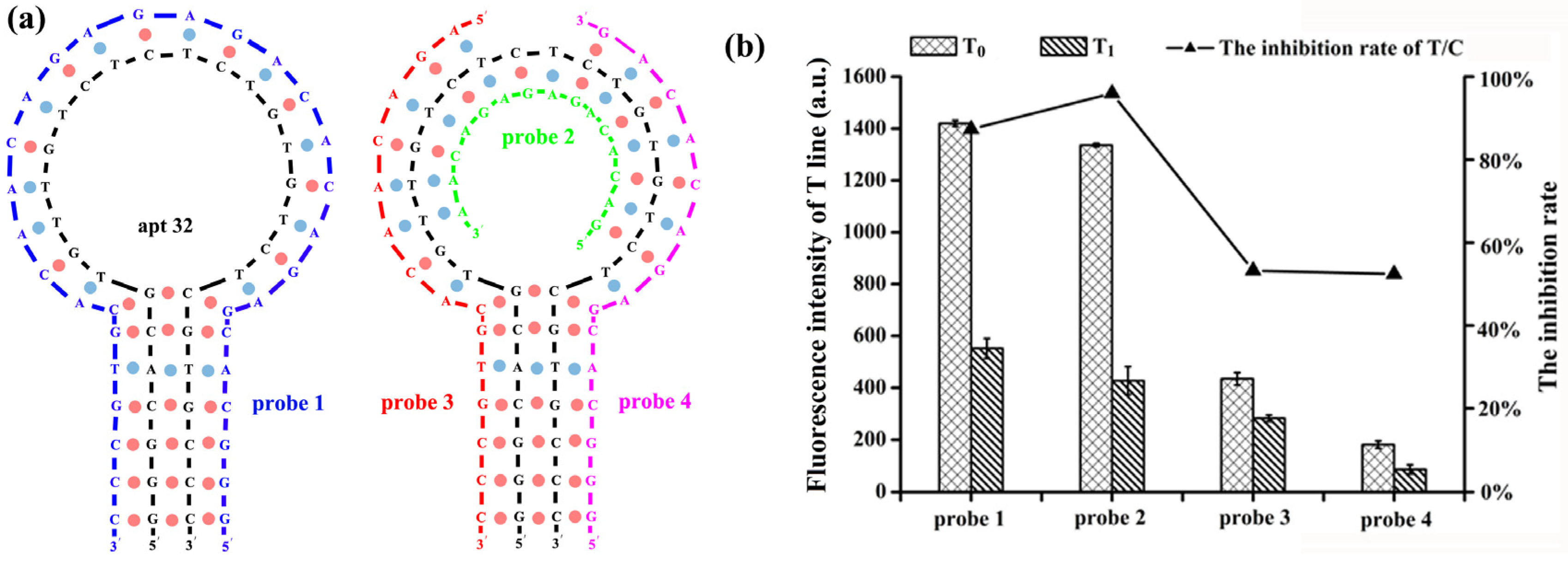
2.3. Optimization of the Complementary Strand on the T-Line
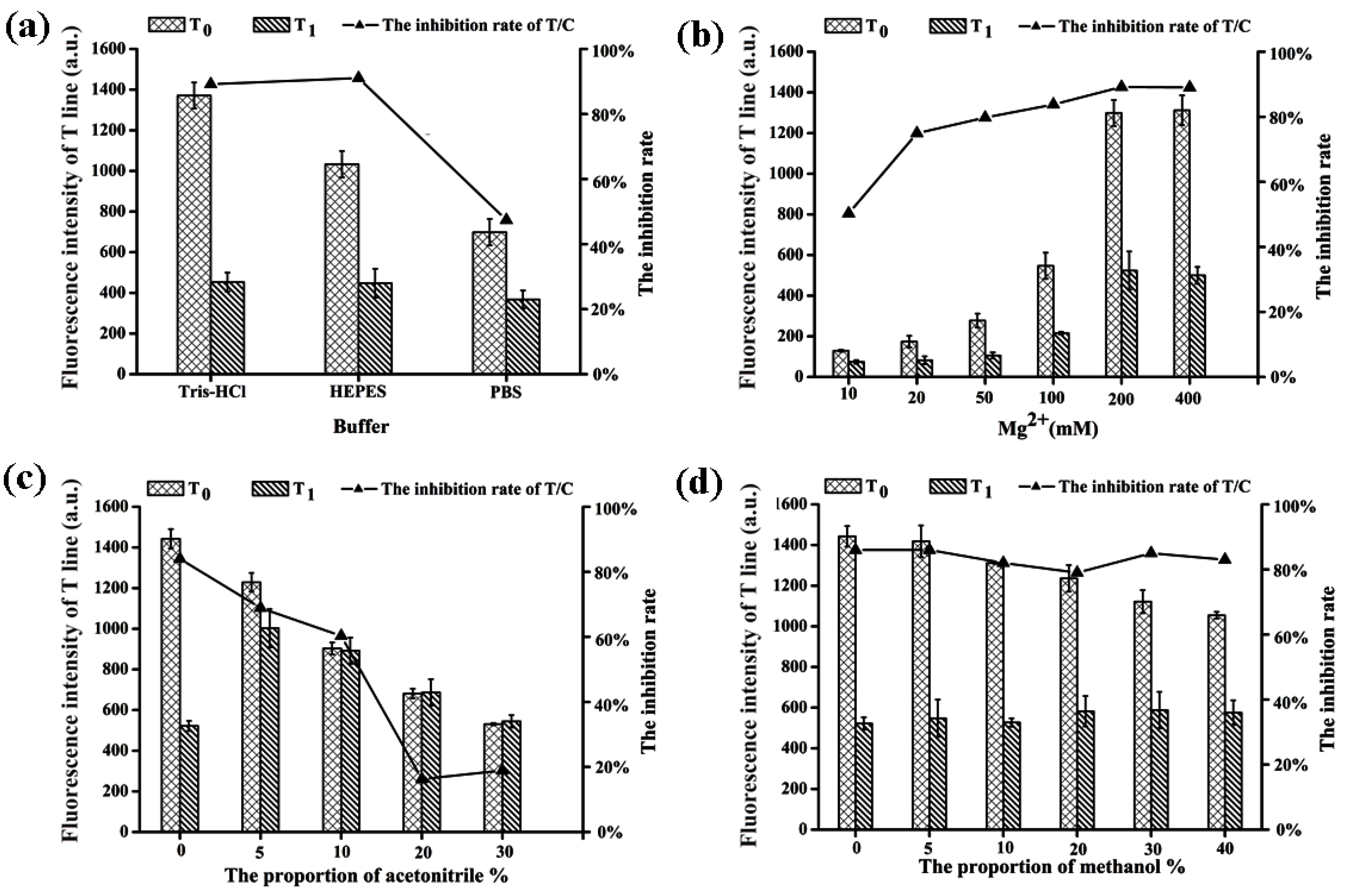
2.4. Optimization of Reaction Conditions between Aptamer and AFB1
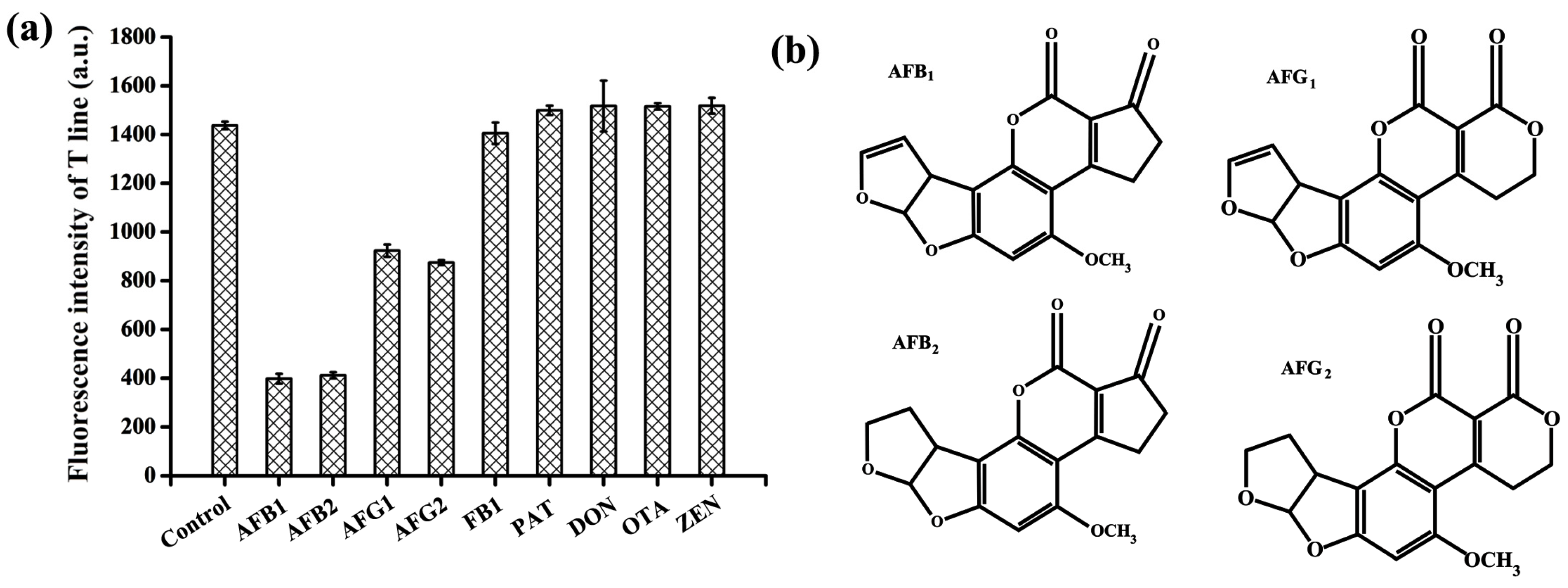
2.5. Specificity Test
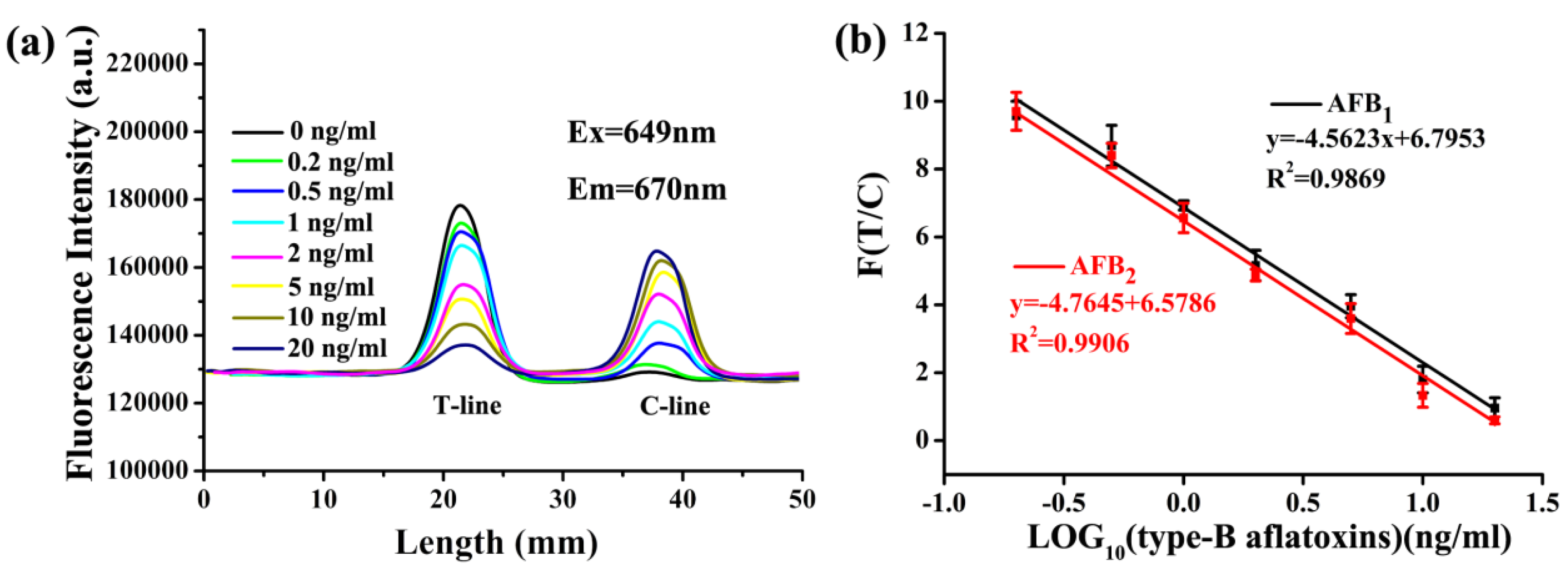
2.6. Quantitative Analysis of Type-B Aflatoxins
2.7. Analysis of Type-B Aflatoxins in Foods
3. Conclusions
4. Materials and Methods
4.1. Reagents and Instruments
4.2. Preparation of ALFs
4.3. Type-B Aflatoxins Analysis
4.4. Optimization of Test Conditions
4.5. Analysis of Food Samples
Author Contributions
Funding
Conflicts of Interest
References
- Wu, J.H.; Zeng, L.W.; Li, N.L.; Liu, C.S.; Chen, J. A wash-free and label-free colorimetric biosensor for naked-eye detection of aflatoxin B1 using G-quadruplex as the signal reporter. Food Chem. 2019, 298, 125034. [Google Scholar] [CrossRef] [PubMed]
- Xue, Z.H.; Zhang, Y.X.; Yu, W.C.; Zhang, J.C.; Wang, J.Y.; Wan, F.; Kim, Y.; Liu, Y.D.; Kou, X.H. Recent advances in aflatoxin B1 detection based on nanotechnology and nanomaterials-A review. Anal. Chim. Acta 2019, 1069, 1–27. [Google Scholar] [CrossRef] [PubMed]
- Wu, F. Perspective: Time to face the fungal threat. Nature 2014, 516, S7. [Google Scholar] [CrossRef] [PubMed]
- Van de Perre, E.; Jacxsens, L.; Lachat, C.; El Tahan, F.; De Meulenaer, B. Impact of maximum levels in European legislation on exposure of mycotoxins in dried products: Case of aflatoxin B1 and ochratoxin A in nuts and dried fruits. Food Chem. Toxicol. 2015, 75, 112–117. [Google Scholar] [CrossRef]
- Hidalgo-Ruiz, J.L.; Romero-González, R.; Martínez Vidal, J.L.; Garrido Frenich, A. Determination of mycotoxins in nuts by ultra high-performance liquid chromatography-tandem mass spectrometry: Looking for a representative matrix. J. Food Compos. Anal. 2019, 82, 103228. [Google Scholar] [CrossRef]
- Wang, Y.J.; Nie, J.Y.; Yan, Z.; Li, Z.X.; Cheng, Y.; Farooq, S. Multi-mycotoxin exposure and risk assessments for Chinese consumption of nuts and dried fruits. J. Integr. Agr. 2018, 17, 1676–1690. [Google Scholar] [CrossRef]
- Campone, L.; Piccinelli, A.L.; Aliberti, L.; Rastrelli, L. Application of pressurized liquid extraction in the analysis of aflatoxins B1, B2, G1 and G2 in nuts. J. Sep. Sci. 2009, 32, 3837–3844. [Google Scholar] [CrossRef]
- Masood, M.; Iqbal, S.Z.; Asi, M.R.; Malik, N. Natural occurrence of aflatoxins in dry fruits and edible nuts. Food Control. 2015, 55, 62–65. [Google Scholar] [CrossRef]
- Cheng, N.; Song, Y.; Shi, Q.; Du, D.; Liu, D.; Luo, Y.; Xu, W.; Lin, Y. Au@Pd nanopopcorn and aptamer nanoflower assisted lateral flow strip for thermal detection of exosomes. Anal. Chem. 2019, 91, 13986–13993. [Google Scholar] [CrossRef]
- Chen, Y.; Cheng, N.; Xu, Y.; Huang, K.; Luo, Y.; Xu, W. Point-of-care and visual detection of P. aeruginosa and its toxin genes by multiple LAMP and lateral flow nucleic acid biosensor. Biosens. Bioelectron. 2016, 81, 317–323. [Google Scholar] [CrossRef]
- Zhang, L.; Chen, Y.; Cheng, N.; Xu, Y.; Huang, K.; Luo, Y.; Wang, P.; Duan, D.; Xu, W. Ultrasensitive detection of viable enterobacter sakazakii by a continual cascade nanozyme biosensor. Anal. Chem. 2017, 89, 10194–10200. [Google Scholar] [CrossRef] [PubMed]
- Tuerk, C.; Gold, L. Systematic evolution of ligands by exponential enrichment RNA ligands to bacteriophage T4 DNA polymerase. Science 1990, 249, 505–510. [Google Scholar] [CrossRef] [PubMed]
- Famulok, M.; Hartig, J.S.; Mayer, G. Functional aptamers and aptazymes in biotechnology, diagnostics, and therapy. Chem. Rev. 2007, 107, 3715–3743. [Google Scholar] [CrossRef] [PubMed]
- Xiao, X.; Zhu, L.; He, W.; Luo, Y.; Xu, W. Functional nucleic acids tailoring and its application. Trac Trends Anal. Chem. 2019, 118, 138–157. [Google Scholar] [CrossRef]
- Shao, X.; Zhu, L.; Feng, Y.; Zhang, Y.; Luo, Y.; Huang, K.; Xu, W. Detachable nanoladders: A new method for signal identification and their application in the detection of ochratoxin A (OTA). Anal. Chim. Acta 2019, 1087, 113–120. [Google Scholar] [CrossRef] [PubMed]
- Wu, S.J.; Liu, L.H.; Duan, N.; Wang, W.Y.; Yu, Q.R.; Wang, Z.P. A test strip for ochratoxin A based on the use of aptamer-modified fluorescence upconversion nanoparticles. Microchim. Acta 2018, 185, 497. [Google Scholar] [CrossRef] [PubMed]
- Zhou, W.L.; Kong, W.J.; Dou, X.W.; Zhao, M.; Ouyang, Z.; Yang, M.H. An aptamer based lateral flow strip for on-site rapid detection of ochratoxin A in Astragalus membranaceus. J. Chromatogr. B 2016, 1022, 102–108. [Google Scholar] [CrossRef]
- Zhang, G.L.; Zhu, C.; Huang, Y.F.; Yan, J.; Chen, A.L. A lateral flow strip based aptasensor for detection of ochratoxin A in corn samples. Molecules 2018, 23, 291. [Google Scholar] [CrossRef] [Green Version]
- Wu, S.J.; Liu, L.H.; Duan, N.; Li, Q.; Zhou, Y.; Wang, Z.P. Aptamer-based lateral flow test strip for rapid detection of zearalenone in corn samples. J. Agr. Food Chem. 2018, 66, 1949–1954. [Google Scholar] [CrossRef]
- Zhu, C.; Zhang, G.L.; Huang, Y.F.; Yang, S.M.; Ren, S.Y.; Gao, Z.X.; Chen, A.L. Dual-competitive lateral flow aptasensor for detection of aflatoxin B1 in food and feedstuffs. J. Hazard. Mater. 2018, 344, 249–257. [Google Scholar] [CrossRef]
- Sun, L.L.; Zhao, Q. Direct fluorescence anisotropy approach for aflatoxin B1 detection and affinity binding study by using single tetramethylrhodamine labeled aptamer. Talanta 2018, 189, 442–450. [Google Scholar] [CrossRef] [PubMed]
- Nutiu, R.; Li, Y. Structure-Switching Signaling Aptamers. J. Am. Chem. Soc. 2003, 125, 4771–4778. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Fang, Z.; Liu, J.; Zeng, L. A simple and rapid biosensor for ochratoxin A based on a structure-switching signaling aptamer. Food Control. 2012, 25, 555–560. [Google Scholar] [CrossRef]
- Le, L.C.; Cruz-Aguado, J.A.; Penner, G.A. DNA ligands for aflatoxin and zearalenone. U.S. Patent Application U.S. PCT/CA2010/001292, 6 September 2012. [Google Scholar]
- Shim, W.B.; Kim, M.J.; Mun, H.; Kim, M.G. An aptamer-based dipstick assay for the rapid and simple detection of aflatoxin B1. Biosens. Bioelectron. 2014, 62, 288–294. [Google Scholar] [CrossRef]
- Li, Y.P.; Sun, L.L.; Zhao, Q. Aptamer-structure switch coupled with horseradish peroxidase labeling on a microplate for the sensitive detection of small molecules. Anal. Chem. 2019, 91, 2615–2619. [Google Scholar] [CrossRef]
- Sun, L.L.; Zhao, Q. Competitive horseradish peroxidase-linked aptamer assay for sensitive detection of aflatoxin B1. Talanta 2018, 179, 344–349. [Google Scholar] [CrossRef]
- Liu, H.; Luan, Y.; Lu, A.; Li, B.; Yang, M.; Wang, J. An oligosorbent-based aptamer affinity column for selective extraction of aflatoxin B2 prior to HPLC with fluorometric detection. Mikrochim. Acta 2017, 185, 71. [Google Scholar] [CrossRef]
- Wang, D.; Zhang, Z.; Li, P.; Zhang, Q.; Zhang, W. Time-resolved fluorescent immunochromatography of aflatoxin B1 in soybean sauce: A rapid and sensitive quantitative analysis. Sensors 2016, 16, 1094. [Google Scholar] [CrossRef] [Green Version]
- Yu, L.; Li, P.W.; Ding, X.X.; Zhang, Q. Graphene oxide and carboxylated graphene oxide: Viable two-dimensional nanolabels for lateral flow immunoassays. Talanta 2017, 165, 167–175. [Google Scholar] [CrossRef]
- Hu, S.R.; Dou, X.W.; Zhang, L.; Xie, Y.J.; Yang, S.H.; Yang, M.H. Rapid detection of aflatoxin B1 in medicinal materials of radix and rhizome by gold immunochromatographic assay. Toxicon 2018, 150, 144–150. [Google Scholar] [CrossRef]
- Wang, M.; Jiang, N.; Xian, H.; Wei, D.; Shi, L.; Feng, X. A single-step solid phase extraction for the simultaneous determination of 8 mycotoxins in fruits by ultra-high performance liquid chromatography tandem mass spectrometry. J. Chromatogr. A 2016, 1429, 22–29. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Sequence (5’–3’) |
|---|---|
| apt 50 | Cy5-A12-GTTGGGCACGTGTTGTCTCTCTGTGTCTCGTGCCCTTCGCTAGGCCCACA |
| apt 32 | Cy5-A12-GGGCACGTGTTGTCTCTCTGTGTCTCGTGCCC |
| apt 26 | Cy5-A12-CACGTGTTGTCTCTCTGTGTCTCGTG |
| probe 1 | biotin-GGGCACGAGACACAGAGAGACAACACGTGCCC |
| probe 2 | biotin-GACACAGAGAGACAA |
| probe 3 | biotin-GGGCACGAGACACAG |
| probe 4 | biotin-AGACAACACGTGCCC |
| poly T | biotin-TTTTTTTTTTTT |
| Method | Test Line | Range (μg/kg) | LOD1 (μg/kg) | Sample | Reference |
|---|---|---|---|---|---|
| Antibody-strip | AFB1−BSA | 0.3–10 | 0.1 | Soybean sauce | [29] |
| Antibody-strip | AFB1−BSA | 0.3–1 | 0.3 | Agricultural products | [30] |
| Antibody-strip | AFB1−BSA | – | 0.1 | medicinal materials | [31] |
| Aptamer-strip | AFB1−BSA | 0.1–1000 | 0.1 | food and feedstuffs | [20] |
| Aptamer-strip | DNA single strand | 0.2–20 | 0.16 | nuts and dried figs | present |
| Sample | Mycotoxins | Spiked (μg/kg) | HPLC-MS/MS Detected (μg/kg) | RSD1 (%) | Proposed Method Detected (μg/kg) | Recovery (%) | RSD (%) |
|---|---|---|---|---|---|---|---|
| Peanut | AFB1 | 3 | 2.9 | 4.1 | 3.1 | 103.7 | 2.2 |
| 10 | 10.1 | 2.9 | 9.6 | 96.5 | 6.5 | ||
| AFB2 | 3 | 3.1 | 1.4 | 3.0 | 101.0 | 3.3 | |
| 10 | 10.5 | 0.8 | 10.9 | 108.7 | 6.9 | ||
| Dried figs | AFB1 | 3 | 2.9 | 3.6 | 3.3 | 109.4 | 2.0 |
| 10 | 9.5 | 3.2 | 10.2 | 102.2 | 2.2 | ||
| AFB2 | 3 | 3.1 | 0.47 | 2.8 | 93.3 | 5.9 | |
| 10 | 10.5 | 0.57 | 11.4 | 112.0 | 2.7 | ||
| Almond | AFB1 | 3 | 2.9 | 0.4 | 3.1 | 102.2 | 2.2 |
| 10 | 10.0 | 2.0 | 9.8 | 97.6 | 4.9 | ||
| AFB2 | 3 | 2.7 | 1.2 | 2.8 | 93.7 | 4.1 | |
| 10 | 9.89 | 0.6 | 9.2 | 93.5 | 7.9 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhao, Z.; Wang, H.; Zhai, W.; Feng, X.; Fan, X.; Chen, A.; Wang, M. A Lateral Flow Strip Based on a Truncated Aptamer-Complementary Strand for Detection of Type-B Aflatoxins in Nuts and Dried Figs. Toxins 2020, 12, 136. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins12020136
Zhao Z, Wang H, Zhai W, Feng X, Fan X, Chen A, Wang M. A Lateral Flow Strip Based on a Truncated Aptamer-Complementary Strand for Detection of Type-B Aflatoxins in Nuts and Dried Figs. Toxins. 2020; 12(2):136. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins12020136
Chicago/Turabian StyleZhao, Zhilei, He Wang, Wenlei Zhai, Xiaoyuan Feng, Xia Fan, Ailiang Chen, and Meng Wang. 2020. "A Lateral Flow Strip Based on a Truncated Aptamer-Complementary Strand for Detection of Type-B Aflatoxins in Nuts and Dried Figs" Toxins 12, no. 2: 136. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins12020136