Inflammatory Reaction Induced by Two Metalloproteinases Isolated from Bothrops atrox Venom and by Fragments Generated from the Hydrolysis of Basement Membrane Components

 , ,
, ,
Abstract
:1. Introduction
2. Results
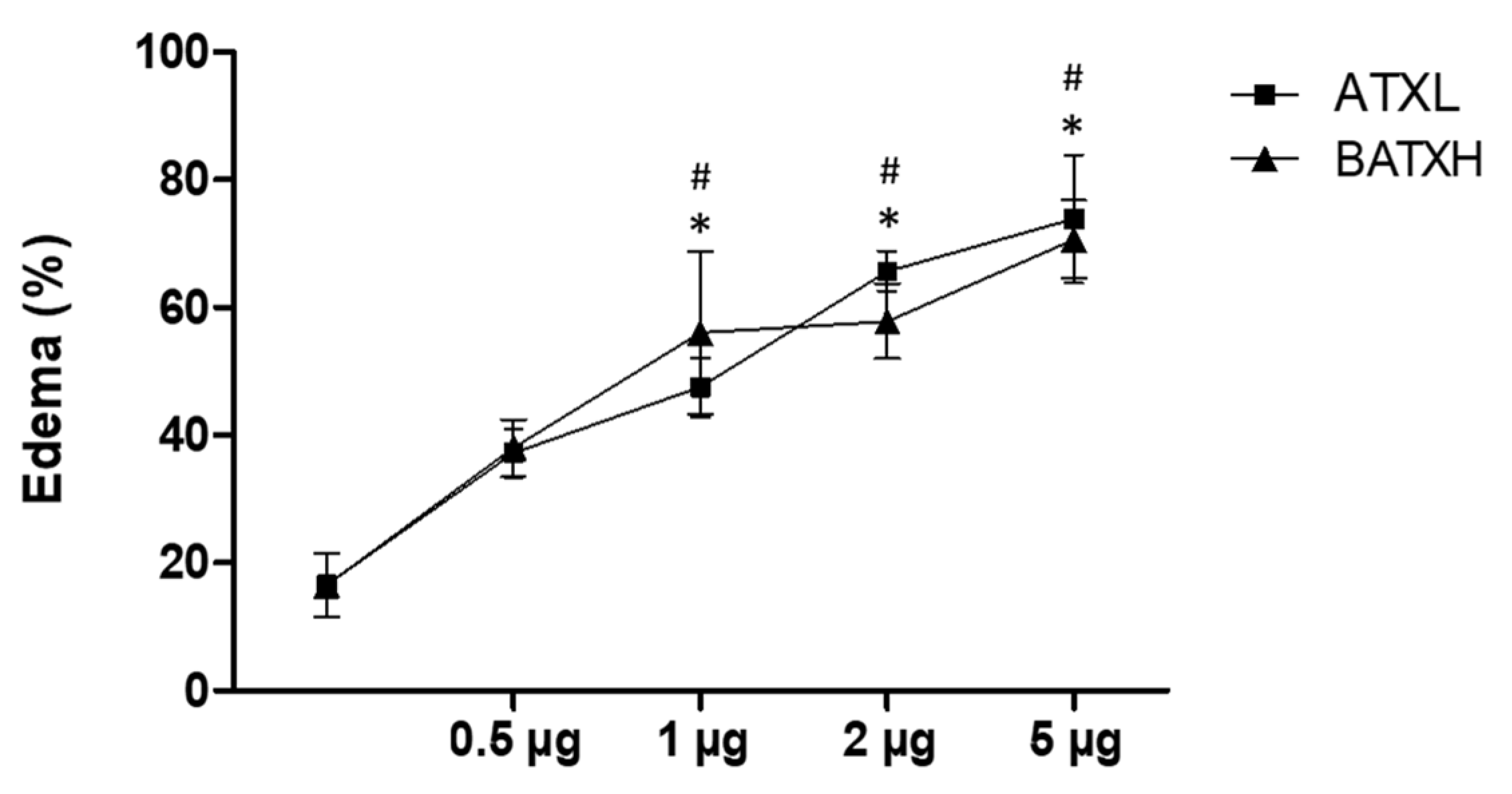
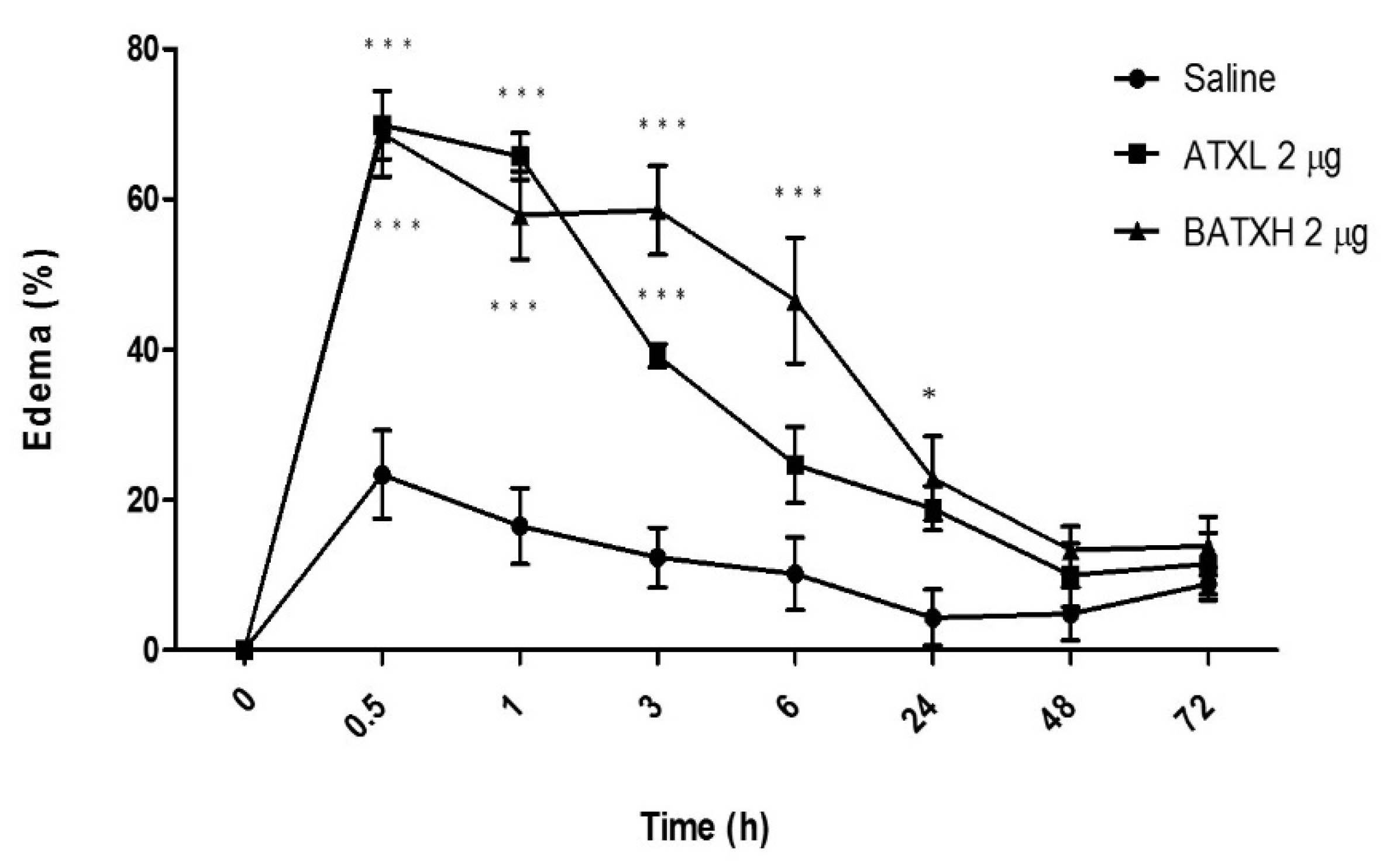
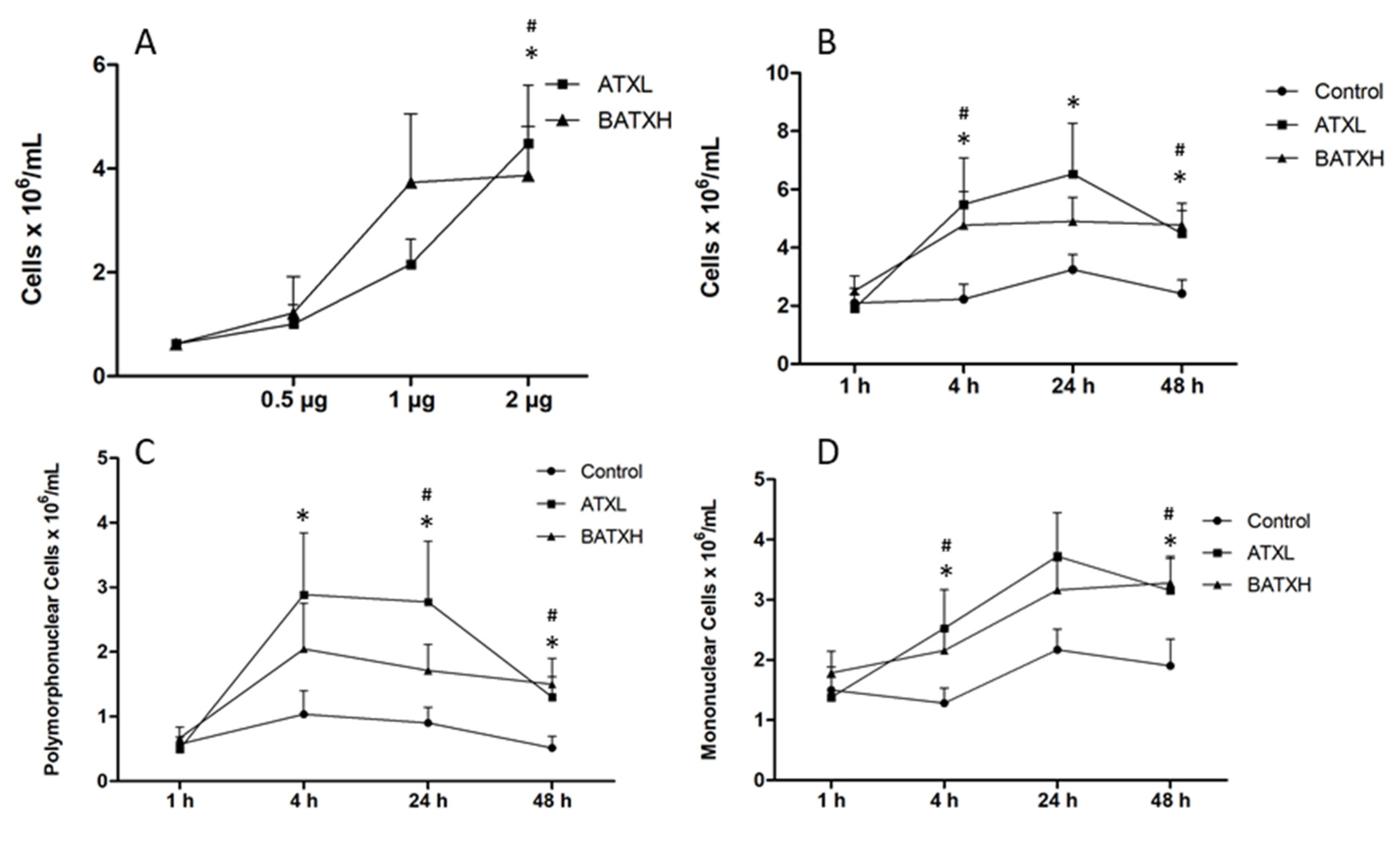
2.1. Inflammatory Reaction Induced by SVMPs
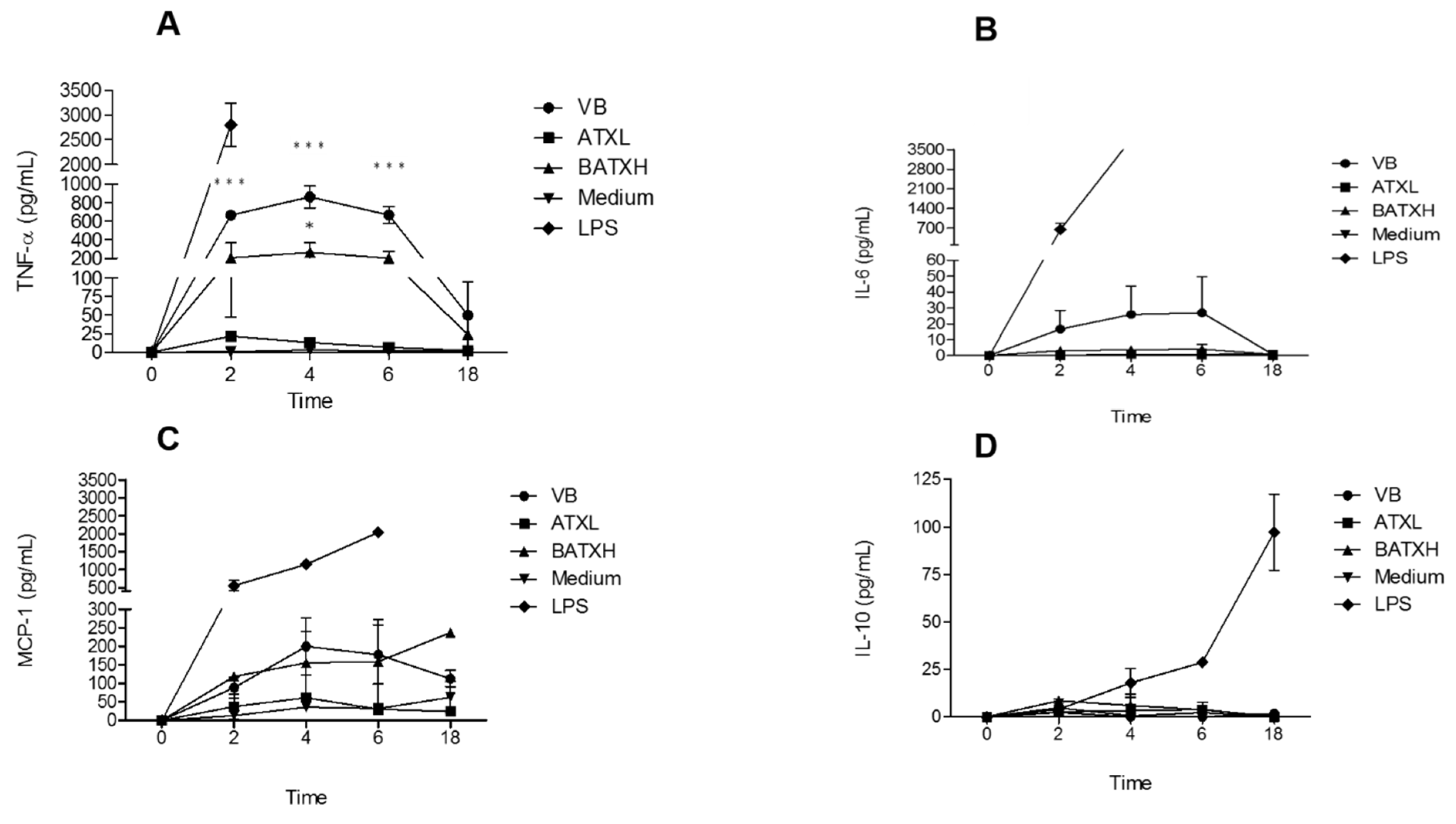
2.2. Cytokines and Chemokine Quantification
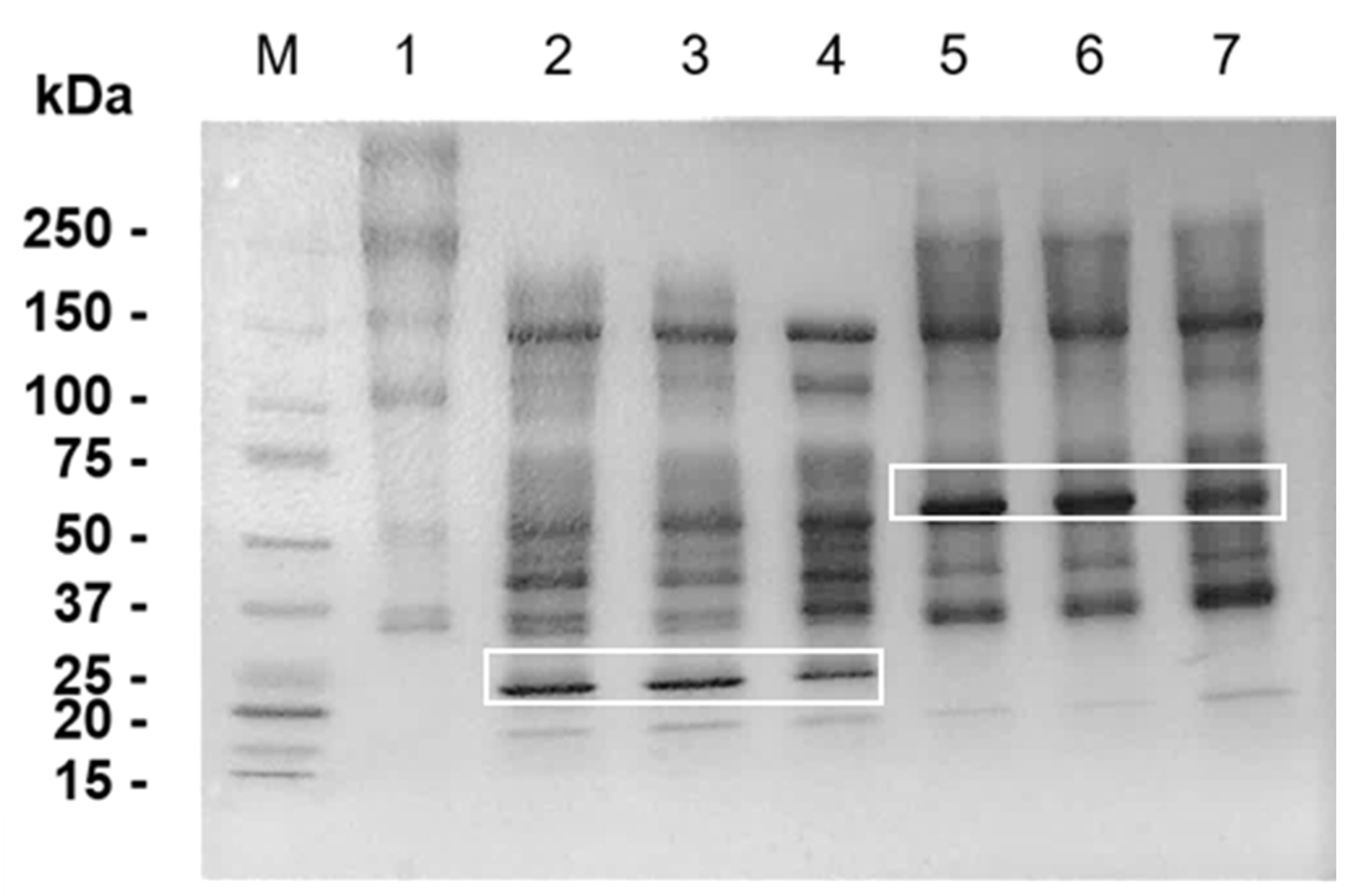
2.3. Hydrolysis of Matrigel by SVMPs
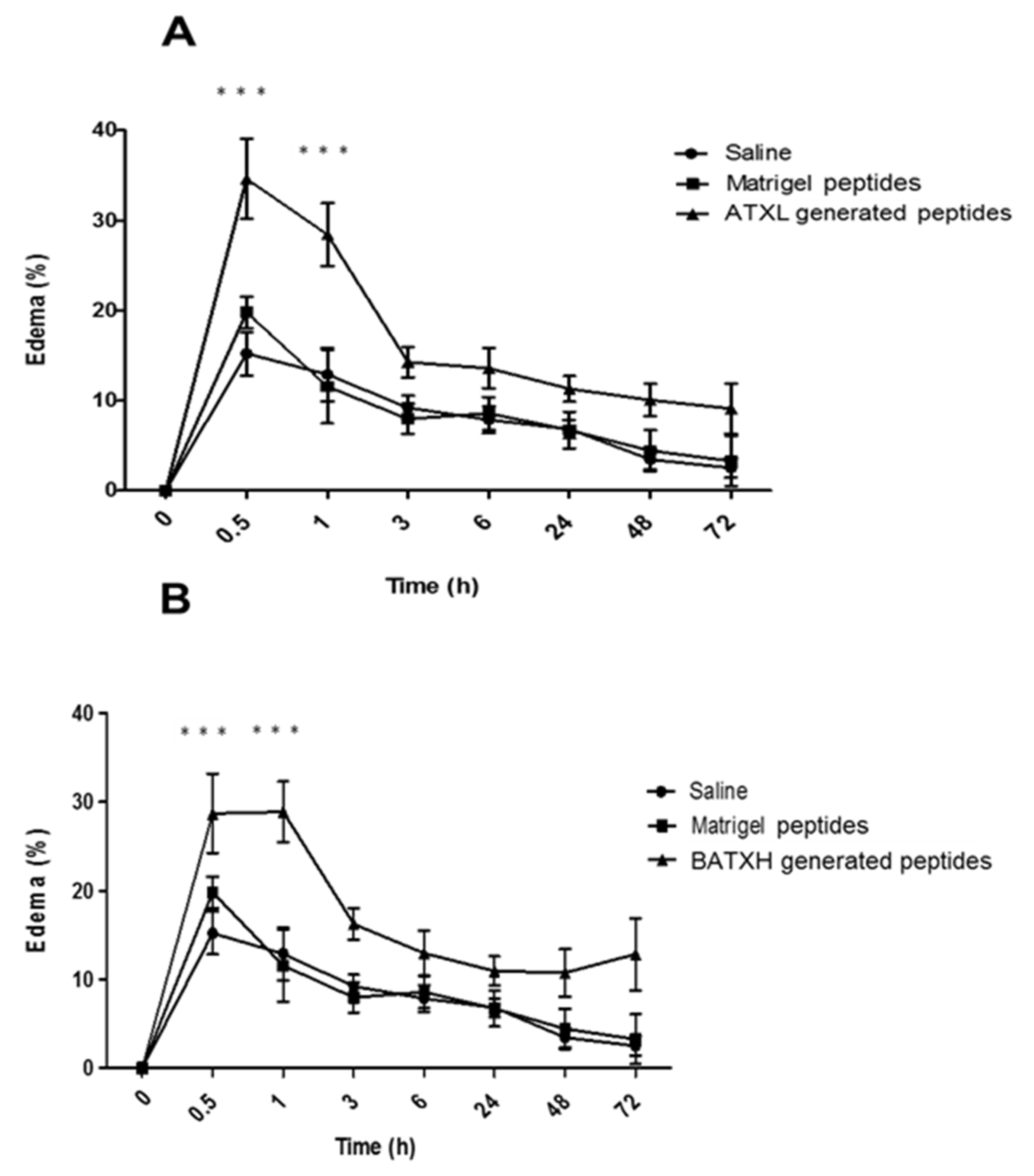
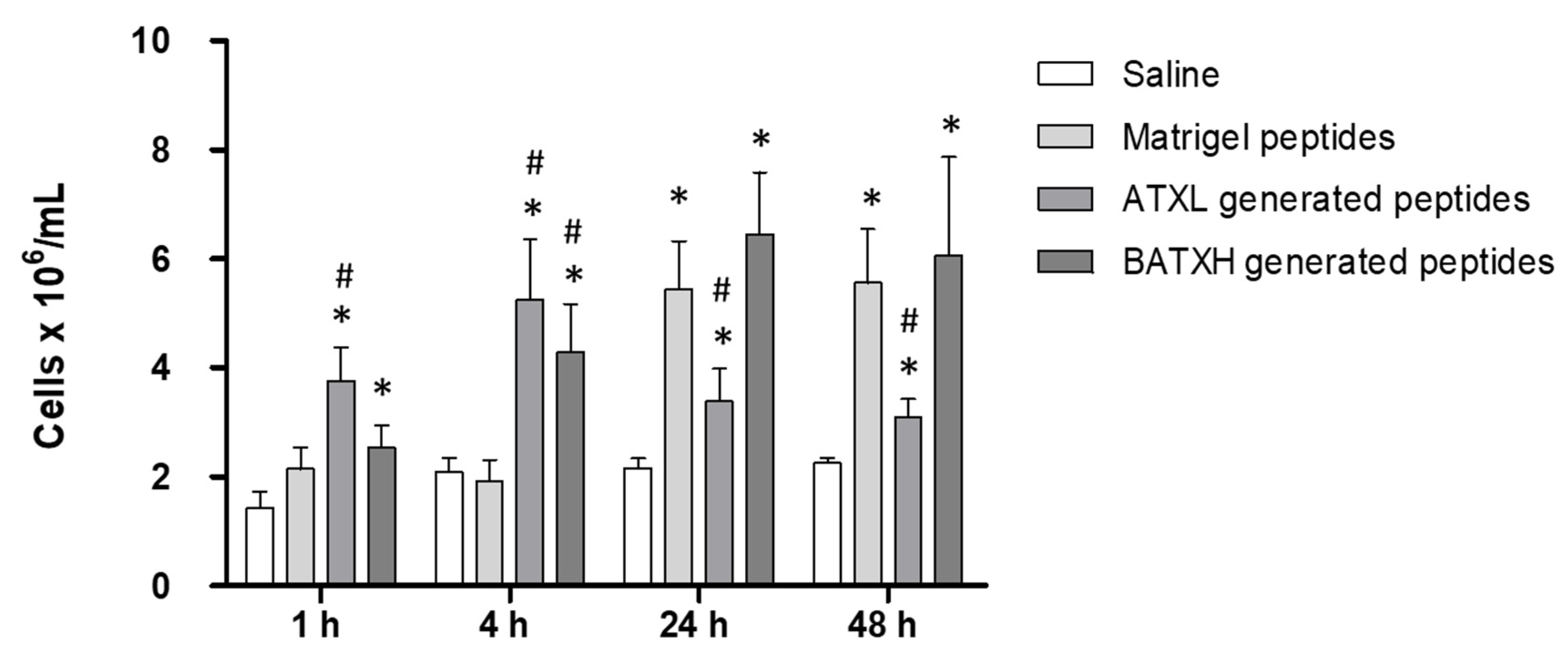
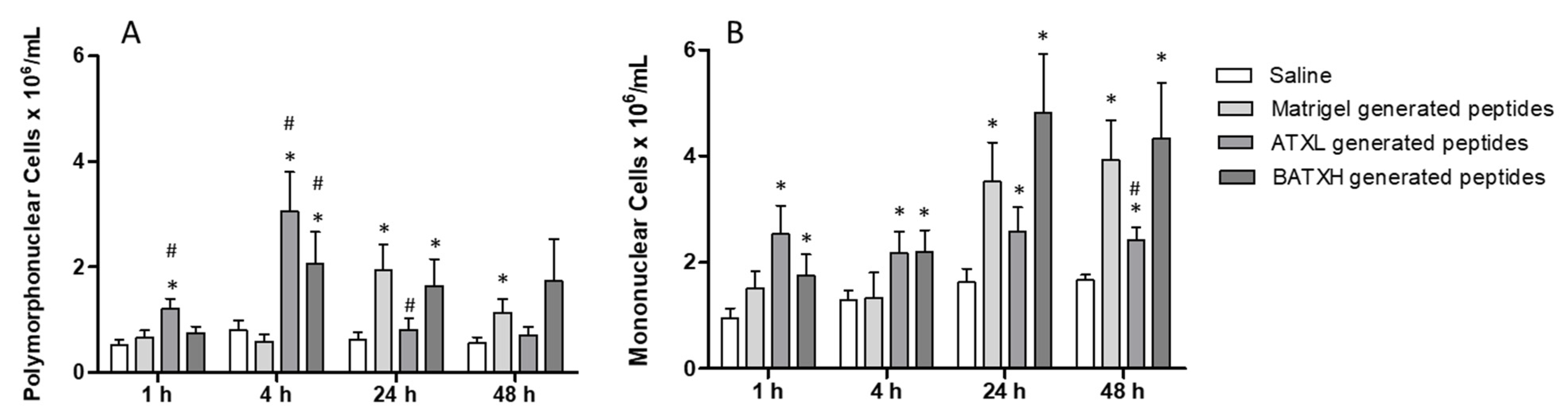
2.4. Characterization of Paw Edema and Leukocyte Accumulation Induced by Matrigel-Derived Peptides
3. Discussion
4. Materials and Methods
4.1. Toxins
4.2. Hydrolysis of Matrigel Components and Isolation of the Resulting Peptides
4.3. Gel Electrophoresis
4.4. Analysis of Generated Peptides by LC–MS/MS
4.5. Evaluation of the Inflammatory Reaction Induced by ATXL, BATXH and Matrigel Hydrolysis Products
4.5.1. Animals
4.5.2. Paw Edema
4.5.3. Leukocyte Recruitment
4.6. Production of Inflammatory Mediators
4.7. Data Analysis
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Williams, D.J.; Faiz, M.A.; Abela-Ridder, B.; Ainsworth, S.; Bulfone, T.C.; Nickerson, A.D.; Habib, A.G.; Junghanss, T.; Fan, H.W.; Turner, M.; et al. Strategy for a globally coordinated response to a priority neglected tropical disease: Snakebite envenoming. PloS Negl. Trop. Dis. 2019, 13, e0007059. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Teixeira, C.e.F.; Fernandes, C.M.; Zuliani, J.P.; Zamuner, S.F. Inflammatory effects of snake venom metalloproteinases. Mem Inst. Oswaldo Cruz 2005, 100, 181–184. [Google Scholar] [CrossRef] [PubMed]
- Zamuner, S.R.; Teixeira, C.F. Cell adhesion molecules involved in the leukocyte recruitment induced by venom of the snake Bothrops jararaca. Mediat. Inflamm. 2002, 11, 351–357. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fox, J.W.; Serrano, S.M. Timeline of key events in snake venom metalloproteinase research. J. Proteom. 2009, 72, 200–209. [Google Scholar] [CrossRef]
- Escalante, T.; Franceschi, A.; Rucavado, A.; Gutiérrez, J.M. Effectiveness of batimastat, a synthetic inhibitor of matrix metalloproteinases, in neutralizing local tissue damage induced by BaP1, a hemorrhagic metalloproteinase from the venom of the snake bothrops asper. Biochem. Pharm. 2000, 60, 269–274. [Google Scholar] [CrossRef]
- Gay, C.C.; Leiva, L.C.; Maruñak, S.; Teibler, P.; Acosta de Pérez, O. Proteolytic, edematogenic and myotoxic activities of a hemorrhagic metalloproteinase isolated from Bothrops alternatus venom. Toxicon 2005, 46, 546–554. [Google Scholar] [CrossRef]
- Baramova, E.N.; Shannon, J.D.; Bjarnason, J.B.; Fox, J.W. Degradation of extracellular matrix proteins by hemorrhagic metalloproteinases. Arch. Biochem. Biophys. 1989, 275, 63–71. [Google Scholar] [CrossRef]
- Costa, E.P.; Clissa, P.B.; Teixeira, C.F.; Moura-da-Silva, A.M. Importance of metalloproteinases and macrophages in viper snake envenomation-induced local inflammation. Inflammation 2002, 26, 13–17. [Google Scholar] [CrossRef]
- Silva, C.A.; Zuliani, J.P.; Assakura, M.T.; Mentele, R.; Camargo, A.C.; Teixeira, C.F.; Serrano, S.M. Activation of alpha(M)beta(2)-mediated phagocytosis by HF3, a P-III class metalloproteinase isolated from the venom of Bothrops jararaca. Biochem. Biophys. Res. Commun. 2004, 322, 950–956. [Google Scholar] [CrossRef]
- Rucavado, A.; Núñez, J.; Gutiérrez, J.M. Blister formation and skin damage induced by BaP1, a haemorrhagic metalloproteinase from the venom of the snake Bothrops asper. Int. J. Exp. Pathol. 1998, 79, 245–254. [Google Scholar]
- Moura-da-Silva, A.M.; Laing, G.D.; Paine, M.J.; Dennison, J.M.; Politi, V.; Crampton, J.M.; Theakston, R.D. Processing of pro-tumor necrosis factor-alpha by venom metalloproteinases: A hypothesis explaining local tissue damage following snake bite. Eur. J. Immunol. 1996, 26, 2000–2005. [Google Scholar] [CrossRef] [PubMed]
- Clissa, P.B.; Laing, G.D.; Theakston, R.D.; Mota, I.; Taylor, M.J.; Moura-da-Silva, A.M. The effect of jararhagin, a metalloproteinase from Bothrops jararaca venom, on pro-inflammatory cytokines released by murine peritoneal adherent cells. Toxicon 2001, 39, 1567–1573. [Google Scholar] [CrossRef]
- Clissa, P.B.; Lopes-Ferreira, M.; Della-Casa, M.S.; Farsky, S.H.; Moura-da-Silva, A.M. Importance of jararhagin disintegrin-like and cysteine-rich domains in the early events of local inflammatory response. Toxicon 2006, 47, 591–596. [Google Scholar] [CrossRef] [PubMed]
- Campos Borges, C.; Sadahiro, M.; dos Santos, M.C. [Epidemiological and clinical aspects of snake bites in the municipalities of the state of Amazonas, Brazil]. Rev. Soc. Bras. Med. Trop. 1999, 32, 637–646. [Google Scholar] [PubMed]
- Moreira, V.; Dos-Santos, M.C.; Nascimento, N.G.; Borges da Silva, H.; Fernandes, C.M.; D’Império Lima, M.R.; Teixeira, C. Local inflammatory events induced by Bothrops atrox snake venom and the release of distinct classes of inflammatory mediators. Toxicon 2012, 60, 12–20. [Google Scholar] [CrossRef]
- Rodrigues, F.G.; Petretski, J.H.; Kanashiro, M.M.; Lemos, L.; da Silva, W.D.; Kipnis, T.L. The complement system is involved in acute inflammation but not in the hemorrhage produced by a Bothrops atrox snake venom low molecular mass proteinase. Mol. Immunol. 2004, 40, 1149–1156. [Google Scholar] [CrossRef]
- Menaldo, D.L.; Bernardes, C.P.; Zoccal, K.F.; Jacob-Ferreira, A.L.; Costa, T.R.; Del Lama, M.P.; Naal, R.M.; Frantz, F.G.; Faccioli, L.H.; Sampaio, S.V. Immune cells and mediators involved in the inflammatory responses induced by a P-I metalloprotease and a phospholipase A. Mol. Immunol. 2017, 85, 238–247. [Google Scholar] [CrossRef]
- Paine, M.J.I.; Desmond, H.P.; Theakston, R.D.G.; Crampton, J.M. Purification, cloning, and molecular characterization of a high-molecular-weight hemorrhagic metalloprotease, jararhagin, from Bothrops jararaca venom. Insights into the disintegrin gene family. J. Biol. Chem. 1992, 267, 22869–22876. [Google Scholar]
- Moura-da-Silva, A.M.; Baldo, C. Jararhagin, A hemorrhagic snake venom metalloproteinase from Bothrops jararaca. Toxicon 2012, 60, 280–289. [Google Scholar] [CrossRef]
- Freitas-de-Sousa, L.A.; Colombini, M.; Lopes-Ferreira, M.; Serrano, S.M.T.; Moura-da-Silva, A.M. Insights into the Mechanisms Involved in Strong Hemorrhage and Dermonecrosis Induced by Atroxlysin-Ia, a PI-Class Snake Venom Metalloproteinase. Toxins 2017, 9, 239. [Google Scholar] [CrossRef] [Green Version]
- Freitas-de-Sousa, L.A.; Amazonas, D.R.; Sousa, L.F.; Sant’Anna, S.S.; Nishiyama, M.Y.; Serrano, S.M.; Junqueira-de-Azevedo, I.L.; Chalkidis, H.M.; Moura-da-Silva, A.M.; Mourão, R.H. Comparison of venoms from wild and long-term captive Bothrops atrox snakes and characterization of Batroxrhagin, the predominant class PIII metalloproteinase from the venom of this species. Biochimie 2015, 118, 60–70. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sanchez, E.F.; Schneider, F.S.; Yarleque, A.; Borges, M.H.; Richardson, M.; Figueiredo, S.G.; Evangelista, K.S.; Eble, J.A. The novel metalloproteinase atroxlysin-I from Peruvian Bothrops atrox (Jergón) snake venom acts both on blood vessel ECM and platelets. Arch. Biochem. Biophys. 2010, 496, 9–20. [Google Scholar] [CrossRef] [PubMed]
- Cintra, A.C.; De Toni, L.G.; Sartim, M.A.; Franco, J.J.; Caetano, R.C.; Murakami, M.T.; Sampaio, S.V. Batroxase, a new metalloproteinase from B. atrox snake venom with strong fibrinolytic activity. Toxicon 2012, 60, 70–82. [Google Scholar] [CrossRef] [PubMed]
- Malinda, K.M.; Kleinman, H.K. The laminins. Int. J. Biochem. Cell Biol. 1996, 28, 957–959. [Google Scholar] [CrossRef]
- Freitas, V.M.; Jaeger, R.G. The effect of laminin and its peptide SIKVAV on a human salivary gland adenoid cystic carcinoma cell line. Virchows Arch. 2002, 441, 569–576. [Google Scholar] [CrossRef]
- Siqueira, A.S.; Pinto, M.P.; Cruz, M.C.; Smuczek, B.; Cruz, K.S.; Barbuto, J.A.; Hoshino, D.; Weaver, A.M.; Freitas, V.M.; Jaeger, R.G. Laminin-111 peptide C16 regulates invadopodia activity of malignant cells through β1 integrin, Src and ERK 1/2. Oncotarget 2016, 7, 47904–47917. [Google Scholar] [CrossRef] [Green Version]
- Rucavado, A.; Nicolau, C.A.; Escalante, T.; Kim, J.; Herrera, C.; Gutiérrez, J.M.; Fox, J.W. Viperid Envenomation Wound Exudate Contributes to Increased Vascular Permeability via a DAMPs/TLR-4 Mediated Pathway. Toxins 2016, 8, 349. [Google Scholar] [CrossRef] [Green Version]
- Escalante, T.; Shannon, J.; Moura-da-Silva, A.M.; Gutiérrez, J.M.; Fox, J.W. Novel insights into capillary vessel basement membrane damage by snake venom hemorrhagic metalloproteinases: A biochemical and immunohistochemical study. Arch. Biochem. Biophys. 2006, 455, 144–153. [Google Scholar] [CrossRef]
- Paes Leme, A.F.; Sherman, N.E.; Smalley, D.M.; Sizukusa, L.O.; Oliveira, A.K.; Menezes, M.C.; Fox, J.W.; Serrano, S.M. Hemorrhagic activity of HF3, a snake venom metalloproteinase: Insights from the proteomic analysis of mouse skin and blood plasma. J. Proteome Res. 2012, 11, 279–291. [Google Scholar] [CrossRef]
- Dale, C.S.; Gonçalves, L.R.; Juliano, L.; Juliano, M.A.; da Silva, A.M.; Giorgi, R. The C-terminus of murine S100A9 inhibits hyperalgesia and edema induced by jararhagin. Peptides 2004, 25, 81–89. [Google Scholar] [CrossRef]
- Bernardes, C.P.; Menaldo, D.L.; Mamede, C.C.; Zoccal, K.F.; Cintra, A.C.; Faccioli, L.H.; Stanziola, L.; de Oliveira, F.; Sampaio, S.V. Evaluation of the local inflammatory events induced by BpirMP, a metalloproteinase from Bothrops pirajai venom. Mol. Immunol. 2015, 68, 456–464. [Google Scholar] [CrossRef] [PubMed]
- Kimura, L.F.; Prezotto-Neto, J.P.; Távora, B.C.; Faquim-Mauro, E.L.; Pereira, N.A.; Antoniazzi, M.M.; Jared, S.G.; Teixeira, C.F.; Santoro, M.L.; Barbaro, K.C. Mast cells and histamine play an important role in edema and leukocyte recruitment induced by Potamotrygon motoro stingray venom in mice. Toxicon 2015, 103, 65–73. [Google Scholar] [CrossRef] [PubMed]
- Ricciotti, E.; FitzGerald, G.A. Prostaglandins and inflammation. Arter. Thromb. Vasc. Biol. 2011, 31, 986–1000. [Google Scholar] [CrossRef]
- De Toni, L.G.; Menaldo, D.L.; Cintra, A.C.; Figueiredo, M.J.; de Souza, A.R.; Maximiano, W.M.; Jamur, M.C.; Souza, G.E.; Sampaio, S.V. Inflammatory mediators involved in the paw edema and hyperalgesia induced by Batroxase, a metalloproteinase isolated from Bothrops atrox snake venom. Int. Immunopharmacol. 2015, 28, 199–207. [Google Scholar] [CrossRef]
- Fernandes, C.M.; Zamuner, S.R.; Zuliani, J.P.; Rucavado, A.; Gutiérrez, J.M.; Teixeira, C.e.F. Inflammatory effects of BaP1 a metalloproteinase isolated from Bothrops asper snake venom: Leukocyte recruitment and release of cytokines. Toxicon 2006, 47, 549–559. [Google Scholar] [CrossRef]
- Farsky, S.H.; Gonçalves, L.R.; Gutiérrez, J.M.; Correa, A.P.; Rucavado, A.; Gasque, P.; Tambourgi, D.V. Bothrops asper snake venom and its metalloproteinase BaP-1 activate the complement system. Role in leucocyte recruitment. Mediat. Inflamm. 2000, 9, 213–221. [Google Scholar] [CrossRef] [Green Version]
- Preciado, L.M.; Pereañez, J.A.; Comer, J. Potential of Matrix Metalloproteinase Inhibitors for the Treatment of Local Tissue Damage Induced by a Type P-I Snake Venom Metalloproteinase. Toxins 2019, 12, 8. [Google Scholar] [CrossRef] [Green Version]
- Moura-da-Silva, A.M.; Ramos, O.H.P.; Baldo, C.; Niland, S.; Hansen, U.; Ventura, J.S.; Furlan, S.; Butera, D.; Della-Casa, M.S.; Tanjoni, I.; et al. Collagen binding is a key factor for the hemorrhagic activity of snake venom metalloproteinases. Biochimie 2008, 90, 484–492. [Google Scholar] [CrossRef]
- Menezes, M.C.; Leme, A.F.P.; Melo, R.L.; Silva, C.A.; Della Casa, M.; Bruni, F.M.; Lima, C.; Lopes-Ferreira, M.; Camargo, A.C.M.; Fox, J.W.; et al. Activation of leukocyte rolling by the cysteine-rich domain and the hyper-variable region of HF3, a snake venom hemorrhagic metalloproteinase. Febs Lett. 2008, 582, 3915–3921. [Google Scholar] [CrossRef] [Green Version]
- Moreira, V.; Teixeira, C.; da Silva, H.B.; Lima, M.R.D.; Dos-Santos, M.C. The role of TLR2 in the acute inflammatory response induced by Bothrops atrox snake venom. Toxicon 2016, 118, 121–128. [Google Scholar] [CrossRef]
- Delgado-Rizo, V.; Martínez-Guzmán, M.A.; Iñiguez-Gutierrez, L.; García-Orozco, A.; Alvarado-Navarro, A.; Fafutis-Morris, M. Neutrophil Extracellular Traps and Its Implications in Inflammation: An Overview. Front. Immunol. 2017, 8, 81. [Google Scholar] [CrossRef] [Green Version]
- Rucavado, A.; Escalante, T.; Teixeira, C.F.; Fernándes, C.M.; Diaz, C.; Gutiérrez, J.M. Increments in cytokines and matrix metalloproteinases in skeletal muscle after injection of tissue-damaging toxins from the venom of the snake Bothrops asper. Mediat. Inflamm 2002, 11, 121–128. [Google Scholar] [CrossRef] [Green Version]
- Fasshauer, M.; Klein, J.; Lossner, U.; Paschke, R. Interleukin (IL)-6 mRNA expression is stimulated by insulin, isoproterenol, tumour necrosis factor alpha, growth hormone, and IL-6 in 3T3-L1 adipocytes. Horm. Metab. Res. 2003, 35, 147–152. [Google Scholar] [CrossRef]
- Tanaka, T.; Narazaki, M.; Kishimoto, T. IL-6 in inflammation, immunity, and disease. Cold Spring Harb Perspect. Biol. 2014, 6, a016295. [Google Scholar] [CrossRef]
- Lopes, D.S.; Baldo, C.; Oliveira, C.e.F.; de Alcântara, T.M.; Oliveira, J.D.; Gourlart, L.R.; Hamaguchi, A.; Homsi-Brandeburgo, M.I.; Moura-da-Silva, A.M.; Clissa, P.B.; et al. Characterization of inflammatory reaction induced by neuwiedase, a P-I metalloproteinase isolated from Bothrops neuwiedi venom. Toxicon 2009, 54, 42–49. [Google Scholar] [CrossRef]
- Gutiérrez, J.M.; Escalante, T.; Rucavado, A.; Herrera, C.; Fox, J.W. A Comprehensive View of the Structural and Functional Alterations of Extracellular Matrix by Snake Venom Metalloproteinases (SVMPs): Novel Perspectives on the Pathophysiology of Envenoming. Toxins 2016, 8, 304. [Google Scholar] [CrossRef] [Green Version]
- Baldo, C.; Jamora, C.; Yamanouye, N.; Zorn, T.M.; Moura-da-Silva, A.M. Mechanisms of vascular damage by hemorrhagic snake venom metalloproteinases: Tissue distribution and in situ hydrolysis. PloS Negl. Trop. Dis. 2010, 4, e727. [Google Scholar] [CrossRef] [Green Version]
- Baldo, C.; Lopes, D.S.; Faquim-Mauro, E.L.; Jacysyn, J.F.; Niland, S.; Eble, J.A.; Clissa, P.B.; Moura-da-Silva, A.M. Jararhagin disruption of endothelial cell anchorage is enhanced in collagen enriched matrices. Toxicon 2015, 108, 240–248. [Google Scholar] [CrossRef] [Green Version]
- Taylor, K.R.; Gallo, R.L. Glycosaminoglycans and their proteoglycans: Host-associated molecular patterns for initiation and modulation of inflammation. Faseb. J. 2006, 20, 9–22. [Google Scholar] [CrossRef]
- Parish, C.R. Heparan sulfate and inflammation. Nat. Immunol. 2005, 6, 861–862. [Google Scholar] [CrossRef]
- Frevert, C.W.; Felgenhauer, J.; Wygrecka, M.; Nastase, M.V.; Schaefer, L. Danger-Associated Molecular Patterns Derived From the Extracellular Matrix Provide Temporal Control of Innate Immunity. J. Histochem. Cytochem. 2018, 66, 213–227. [Google Scholar] [CrossRef] [PubMed]
- Newton, K.; Dixit, V.M. Signaling in innate immunity and inflammation. Cold Spring Harb. Perspect. Biol. 2012, 4. [Google Scholar] [CrossRef] [PubMed]
- Anders, H.J.; Schaefer, L. Beyond tissue injury-damage-associated molecular patterns, toll-like receptors, and inflammasomes also drive regeneration and fibrosis. J. Am. Soc. Nephrol. 2014, 25, 1387–1400. [Google Scholar] [CrossRef] [PubMed]
- Sorokin, L. The impact of the extracellular matrix on inflammation. Nat. Rev. Immunol. 2010, 10, 712–723. [Google Scholar] [CrossRef] [PubMed]
- Hughes, C.S.; Postovit, L.M.; Lajoie, G.A. Matrigel: A complex protein mixture required for optimal growth of cell culture. Proteomics 2010, 10, 1886–1890. [Google Scholar] [CrossRef] [PubMed]
- Kleinman, H.K.; Martin, G.R. Matrigel: Basement membrane matrix with biological activity. Semin. Cancer Biol. 2005, 15, 378–386. [Google Scholar] [CrossRef]
- Talbot, N.C.; Caperna, T.J. Proteome array identification of bioactive soluble proteins/peptides in Matrigel: Relevance to stem cell responses. Cytotechnology 2015, 67, 873–883. [Google Scholar] [CrossRef] [Green Version]
- Herrera, C.; Macêdo, J.K.; Feoli, A.; Escalante, T.; Rucavado, A.; Gutiérrez, J.M.; Fox, J.W. Muscle Tissue Damage Induced by the Venom of Bothrops asper: Identification of Early and Late Pathological Events through Proteomic Analysis. Plos Negl. Trop. Dis. 2016, 10, e0004599. [Google Scholar] [CrossRef] [Green Version]
- Laemmli, U.K. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 1970, 227, 680–685. [Google Scholar] [CrossRef]
- Távora, B.C.; Kimura, L.F.; Antoniazzi, M.M.; Chiariello, T.M.; Faquim-Mauro, E.L.; Barbaro, K.C. Involvement of mast cells and histamine in edema induced in mice by Scolopendra viridicornis centipede venom. Toxicon 2016, 121, 51–60. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Proteins * | Uniprot Entry | Identified Peptides * | |
|---|---|---|---|
| ATXL | BATXH | ||
| Laminin subunit alpha-1 | P19137 | HADIIIKGNG | ALLHAPTGS |
| IRSQQDVLGGHRQ | LWDLGSGSTR | ||
| LVEHVPGRPVR | LINGRPSADDPSP | ||
| LINGRPSADDPSP | |||
| Laminin subunit beta-1 | P02469 | AIKQADEDIQGTQN | |
| Laminin subunit gamma-1 | P02468 | IRNTIEETGI | |
| Tubulin beta-4B chain | P68372 | HSLGGGTGSGMGT | |
| Vimentin | P20152 | ANYQDTIGR | |
| Actin, cytoplasmic 2 | P63260 | TVLSGGTTMYPGIAD | |
| QVITIGNER | |||
| Fibrinogen beta chain | Q8K0E8 | LRPAPPPISGGGY | |
| 60 kDa heat shock protein, mitochondrial | P63038 | VGGTSDVEVNEK | |
| 60S ribosomal protein L30 | P62889 | IIDPGDSDIIR | |
| Glyceraldehyde-3-phosphate dehydrogenase | P16858 | HSSTFDAGAGIA | IFQERDPTNIK |
| ITIFQERDPTNIK | |||
| Heterogeneous nuclear ribonucleoprotein F | Q9Z2X1 | SVQRPGPYDRPGTA | |
| Prolyl 3-hydroxylase 1 | Q3V1T4 | FSSGTENPHGVKA | |
| Transcription intermediary factor 1-beta | Q62318 | LTEGPGAEGPR | |
| 40S ribosomal protein S3 | P62908 | IGPKKPLPDHVS | |
| 40S ribosomal protein S4, X isoform | P62702 | TIRYPDPLI | |
| 78 kDa glucose-regulated protein | P20029 | VAFTPEGER | |
| Hemoglobin subunit beta-1 | P02088 | LVVYPWTQR | |
| Protein disulfide-isomerase | P09103 | ITSNSGVFSK | |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Almeida, M.T.d.; Freitas-de-Sousa, L.A.; Colombini, M.; Gimenes, S.N.C.; Kitano, E.S.; Faquim-Mauro, E.L.; Serrano, S.M.T.; Moura-da-Silva, A.M. Inflammatory Reaction Induced by Two Metalloproteinases Isolated from Bothrops atrox Venom and by Fragments Generated from the Hydrolysis of Basement Membrane Components. Toxins 2020, 12, 96. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins12020096
Almeida MTd, Freitas-de-Sousa LA, Colombini M, Gimenes SNC, Kitano ES, Faquim-Mauro EL, Serrano SMT, Moura-da-Silva AM. Inflammatory Reaction Induced by Two Metalloproteinases Isolated from Bothrops atrox Venom and by Fragments Generated from the Hydrolysis of Basement Membrane Components. Toxins. 2020; 12(2):96. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins12020096
Chicago/Turabian StyleAlmeida, Michelle Teixeira de, Luciana Aparecida Freitas-de-Sousa, Monica Colombini, Sarah N. C. Gimenes, Eduardo S. Kitano, Eliana L. Faquim-Mauro, Solange M. T. Serrano, and Ana Maria Moura-da-Silva. 2020. "Inflammatory Reaction Induced by Two Metalloproteinases Isolated from Bothrops atrox Venom and by Fragments Generated from the Hydrolysis of Basement Membrane Components" Toxins 12, no. 2: 96. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins12020096