Effects of Nutrient Limitation on the Synthesis of N-Rich Phytoplankton Toxins: A Meta-Analysis

 , ,
, ,
Abstract
:1. Introduction
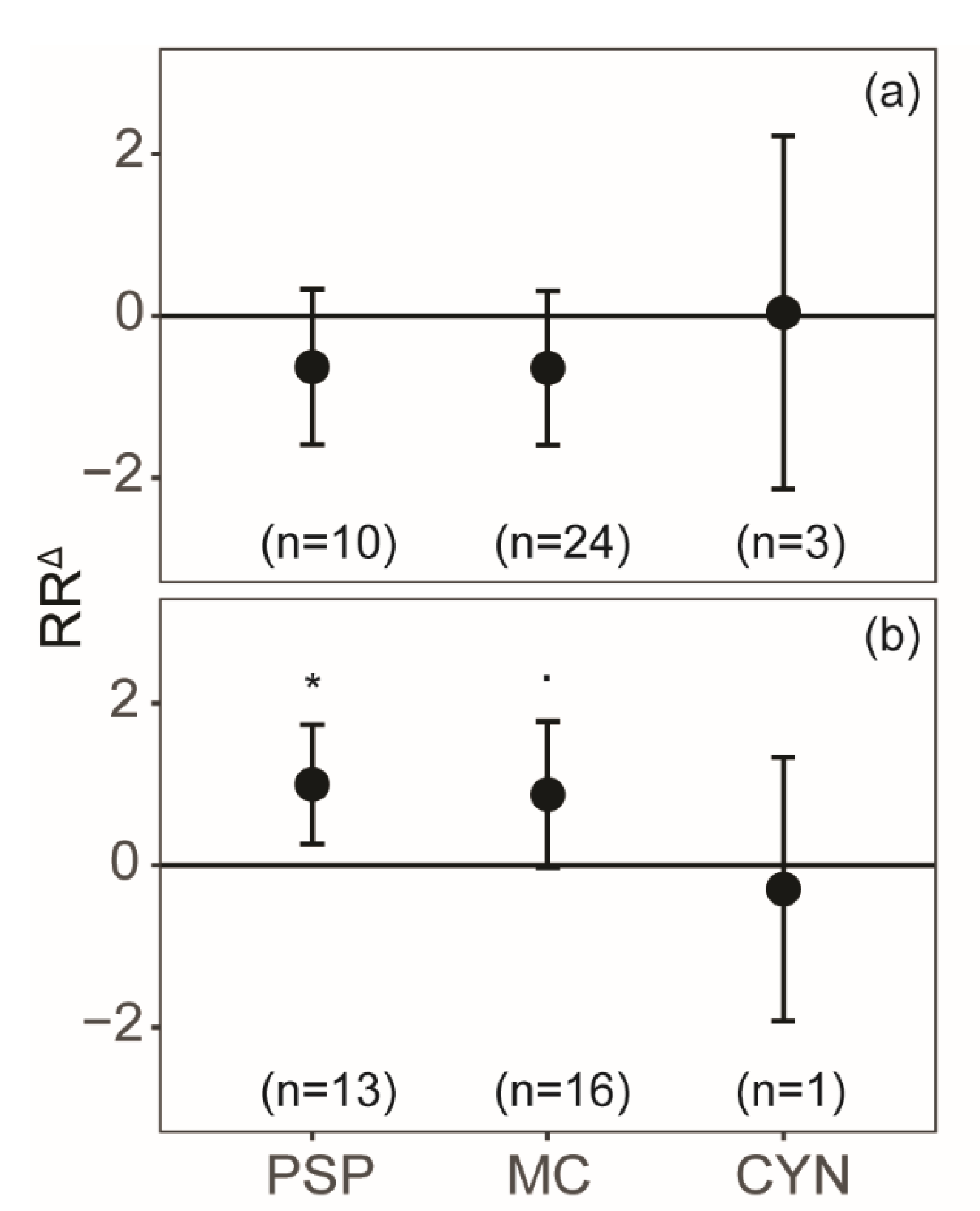
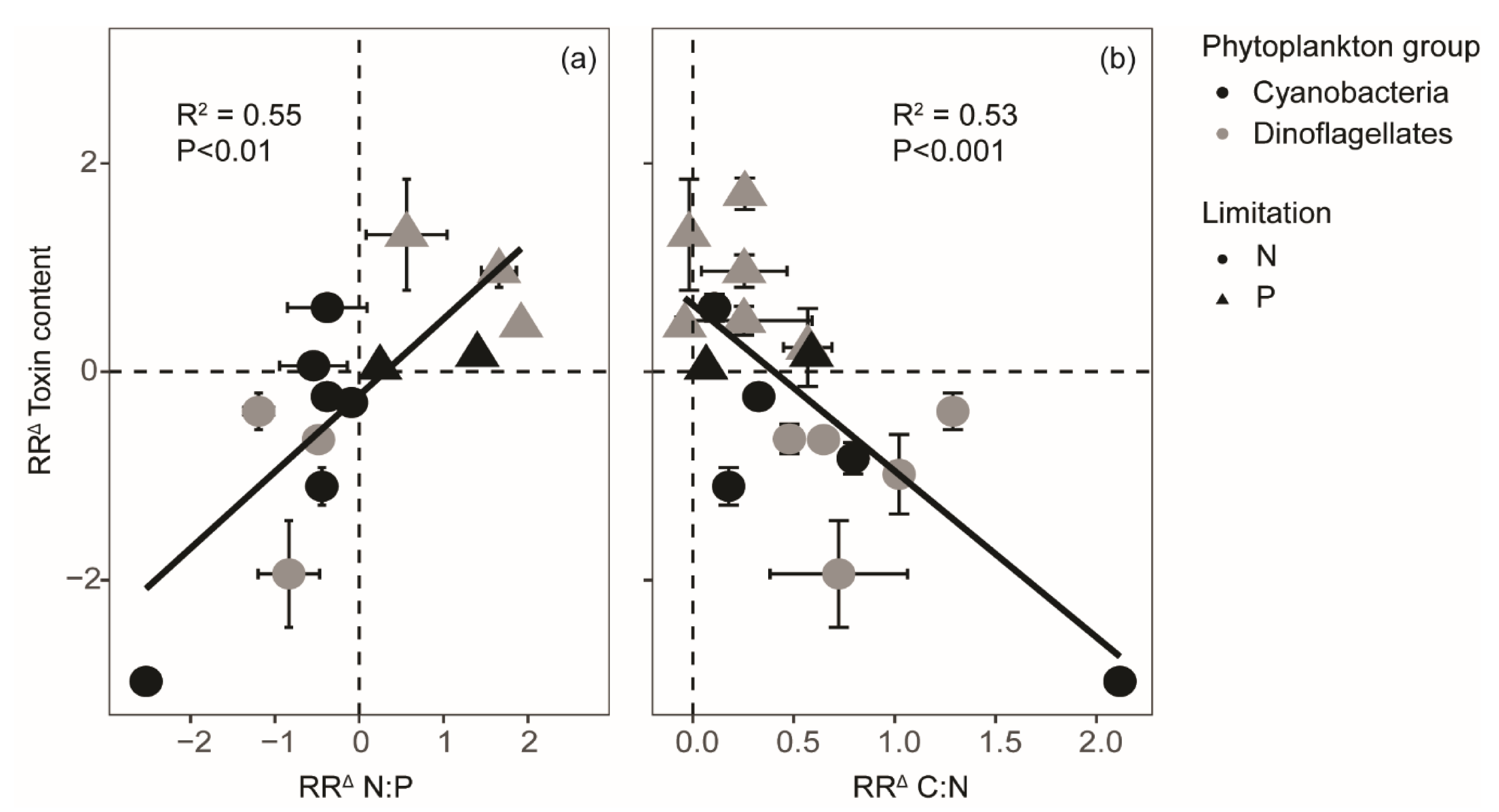
2. Results
3. Discussion
4. Materials and Methods
4.1. Data Collection
4.2. Response Ratios
4.3. Statistical Analyses
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Anderson, D.M.; Burkholder, J.M.; Cochlan, W.P.; Glibert, P.M.; Gobler, C.J.; Heil, C.A.; Kudela, R.; Parsons, M.L.; Jack, J.E.; Townsend, D.W.; et al. Harmful algal blooms and eutrophication: Examining linkages from selected coastal regions of the United States. Harmful Algae 2008, 8, 39–53. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anderson, D.M.; Cembella, A.D.; Hallegraeff, G.M. Progress in understanding harmful algal blooms: Paradigm shifts and new technologies for research, monitoring, and management. Ann. Rev. Mar. Sci. 2012, 4, 143–176. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Glibert, P.M.; Seitzinger, S.; Heil, C.; Burkholder, J.M.; Parrow, M.; Codispoti, L.; Kelly, V. The role of eutrophication in the global proliferation of harmful algal blooms. Oceanography 2005, 18, 198–209. [Google Scholar] [CrossRef] [Green Version]
- Glibert, P.M.; Burkholder, J.M. The complex relationships between increases in fertilization of the earth, coastal eutrophication and proliferation of harmful algal blooms. In Ecology of Harmful Algae; Graneli, E., Turner, J.T., Eds.; Springer: Berlin/Heidelberg, Germany, 2006; Volume 189, pp. 341–354. ISBN 978-3-540-32209-2. [Google Scholar]
- Parsons, M.L.; Dortch, Q. Sedimentological evidence of an increase in Pseudo-nitzschia (Bacillariophyceae) abundance in response to coastal eutrophication. Limnol. Oceanogr. 2002, 47, 551–558. [Google Scholar] [CrossRef]
- O’Neill, K.; Schreider, M.; McArthur, L.; Schreider, S. Changes in the water quality characteristics during a macroalgal bloom in a coastal lagoon. Ocean Coast. Manag. 2015, 118, 32–36. [Google Scholar] [CrossRef]
- Paerl, H.W.; Otten, T.G. Harmful cyanobacterial blooms: Causes, consequences, and controls. Microb. Ecol. 2013, 65, 995–1010. [Google Scholar] [CrossRef]
- Paerl, H.W.; Scott, J.T.; McCarthy, M.J.; Newell, S.E.; Gardner, W.S.; Havens, K.E.; Hoffman, D.K.; Wilhelm, S.W.; Wurtsbaugh, W.A. It takes two to tango: When and where dual nutrient (N & P) reductions are needed to protect lakes and downstream ecosystems. Environ. Sci. Technol. 2016, 50, 10805–10813. [Google Scholar]
- Elser, J.J.; Bracken, M.E.S.; Cleland, E.E.; Gruner, D.S.; Harpole, W.S.; Hillebrand, H.; Ngai, J.T.; Seabloom, E.W.; Shurin, J.B.; Smith, J.E. Global analysis of nitrogen and phosphorus limitation of primary producers in freshwater, marine and terrestrial ecosystems. Ecol. Lett. 2007, 10, 1135–1142. [Google Scholar] [CrossRef] [Green Version]
- Anderson, D.M.; Glibert, P.M.; Burkholder, J.M. Harmful algal blooms and eutrophication nutrient sources, composition, and consequences. Estuaries 2002, 25, 704–726. [Google Scholar] [CrossRef]
- Smayda, T.J. Harmful algal blooms: Their ecophysiology and general relevance to phytoplankton blooms in the sea. Limnol. Oceanogr. 1997, 42, 1137–1153. [Google Scholar] [CrossRef]
- Hallegraeff, G.M. A review of harmful algal blooms and their apparent global increase. Phycologia 1993, 32, 79–99. [Google Scholar] [CrossRef] [Green Version]
- Durbin, E.; Teegarden, G.; Campbell, R.; Cembella, A.; Baumgartner, M.F.; Mate, B.R. North Atlantic right whales, Eubalaena glacialis, exposed to paralytic shellfish poisoning (PSP) toxins via a zooplankton vector, Calanus finmarchicus. Harmful Algae 2002, 1, 243–251. [Google Scholar] [CrossRef]
- Cembella, A.D.; Quilliam, M.A.; Lewis, N.I.; Bauder, A.G.; Aversano, C.D.; Thomas, K.; Jellett, J.; Cusack, R.R. The toxigenic marine dinoflagellate Alexandrium tamarense as the probable cause of mortality of caged salmon in Nova Scotia. Harmful Algae 2002, 1, 313–325. [Google Scholar] [CrossRef]
- Doucette, G.J.; Cembella, A.D.; Martin, J.L.; Michaud, J.; Cole, T.V.N.; Rolland, R.M. Paralytic shellfish poisoning (PSP) toxins in North Atlantic right whales Eubalaena glacialis and their zooplankton prey in the Bay of Fundy, Canada. Mar. Ecol. Prog. Ser. 2006, 306, 303–313. [Google Scholar] [CrossRef]
- Sunda, W.G.; Graneli, E.; Gobler, C.J. Positive feedback and the development and persistence of ecosystem disruptive algal blooms. J. Phycol. 2006, 42, 963–974. [Google Scholar] [CrossRef]
- Bláha, L.; Babica, P.; Maršálek, B. Toxins produced in cyanobacterial water blooms—Toxicity and risks. Interdiscip. Toxicol. 2009, 2, 36–41. [Google Scholar] [CrossRef] [Green Version]
- Baselga-Cervera, B.; García Balboa, C.; Costas, E.; Lopez-Rodas, V. Why cyanobacteria produce toxins? Evolutionary game theory suggests the key. Int. J. Biol. 2014, 7, 64–77. [Google Scholar] [CrossRef] [Green Version]
- Davidson, K.; Gowen, R.J.; Harrison, P.J.; Fleming, L.E.; Hoagland, P.; Moschonas, G. Anthropogenic nutrients and harmful algae in coastal waters. J. Environ. Manag. 2014, 146, 206–216. [Google Scholar] [CrossRef] [Green Version]
- Wang, D.Z. Neurotoxins from marine dinoflagellates: A brief review. Mar. Drugs 2008, 6, 349–371. [Google Scholar] [CrossRef]
- Anderson, D.M. Toxic algae blooms and red tides: A global perspective. In Red Tides: Biology, Environmental Science and Toxicology; Okaichi, T., Anderson, D.M., Nemoto, T., Eds.; Elsevier: New York, NY, USA, 1989; pp. 11–16. [Google Scholar]
- Van Dolah, F.M. Diversity of marine and freshwater algal toxins. In Seafood Toxicology: Pharmacology, Physiology and Detection; Botana, L.M., Ed.; Luis M. Botana: New York, NY, USA, 2000; pp. 19–43. [Google Scholar]
- Zhang, D.; Xie, P.; Liu, Y.; Chen, J.; Liang, G. Bioaccumulation of the hepatotoxic microcystins in various organs of a freshwater snail from a subtropical Chinese Lake, Taihu Lake, with dense toxic Microcystis blooms. Environ. Toxicol. Chem. 2007, 26, 171–176. [Google Scholar] [CrossRef]
- Martínez-Hernández, J.; López-Rodas, V.; Costas, E. Microcystins from tap water could be a risk factor for liver and colorectal cancer: A risk intensified by global change. Med. Hypotheses 2009, 72, 539–540. [Google Scholar] [CrossRef] [PubMed]
- Chorus, I.; Bartram, J. Toxic Cyanobacteria in Water: A Guide to Their Public Health Consequences, Monitoring and Management; WHO: London, UK, 1999; ISBN 9781317436980. [Google Scholar]
- Drobac, D.; Tokodi, N.; Simeunović, J.; Baltić, V.; Stanić, D.; Svirčev, Z. Human exposure to cyanotoxins and their effects on health. Arh. Hig. Rada Toksikol. 2013, 64, 305–316. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sterner, R.W.; Elser, J.J. Ecological Stoichiometry: The Biology of Elements from Molecules to the Biosphere; Princeton Univeristy Press: Princeton, NJ, USA, 2002. [Google Scholar]
- Geider, R.J.; La Roche, J. Redfield revisited: Variability of C:N:P in marine microalgae and its biochemical basis. Eur. J. Phycol. 2002, 37, 1–17. [Google Scholar] [CrossRef] [Green Version]
- Hillebrand, H.; Steinert, G.; Boersma, M.; Malzahn, A. Goldman revisited: Faster-growing phytoplankton has lower N:P and lower stoichiometric flexibility. Limnol. Oceanogr. 2013, 58, 2076–2088. [Google Scholar] [CrossRef]
- Gao, B.; Liu, J.; Zhang, C.; Van de Waal, D.B. Biological stoichiometry of oleaginous microalgal lipid synthesis: The role of N:P supply ratios and growth rate on microalgal elemental and biochemical composition. Algal Res. 2018, 32, 353–361. [Google Scholar] [CrossRef]
- Chakraborty, S.; Pančić, M.; Andersen, K.H.; Kiørboe, T. The cost of toxin production in phytoplankton: The case of PST producing dinoflagellates. ISME J. 2019, 13, 64–75. [Google Scholar] [CrossRef]
- Van de Waal, D.B.; Smith, V.H.; Declerck, S.A.J.; Stam, E.C.M.; Elser, J.J. Stoichiometric regulation of phytoplankton toxins. Ecol. Lett. 2014, 17, 736–742. [Google Scholar] [CrossRef]
- Van de Waal, D.B.; Tillmann, U.; Zhu, M.M.; Koch, B.P.; Rost, B.; John, U. Nutrient pulse induces dynamic changes in cellular C:N:P, amino acids, and paralytic shellfish poisoning toxins in Alexandrium tamarense. Mar. Ecol. Prog. Ser. 2013, 493, 57–69. [Google Scholar] [CrossRef] [Green Version]
- Cembella, A.D. Ecophysiology and metabolism of paralytic shellfish toxins in marine microalgae. In Physiological Ecology of Harmful Algal Blooms; Anderson, D.M., Cembella, A.D., Hallegraeff, G.M., Eds.; Springer: Berlin, Germany, 1998. [Google Scholar]
- Graneli, E.; Johansson, N.; Panosso, R. Cellular toxin contents in relation to nutrient conditions for different groups of phycotoxins. In Harmful Algae; Reguera, B., Blanco, J., Fernandez, M.L., Wyatt, T., Eds.; Xunta de Galicia: Santiago de Compostela, Spain, 1998; IOC-UNESCO; pp. 321–324. [Google Scholar]
- Wagner, N.D.; Osburn, F.S.; Wang, J.; Taylor, R.B.; Boedecker, A.R.; Chambliss, C.K.; Brooks, B.W.; Scott, J.T. Biological stoichiometry regulates toxin production in Microcystis aeruginosa (UTEX 2385). Toxins 2019, 11, 601. [Google Scholar] [CrossRef] [Green Version]
- Sun, J.; Hutchins, D.A.; Feng, Y.; Seubert, E.L.; Caron, D.A.; Fu, F.X. Effects of changing pCO2 and phosphate availability on domoic acid production and physiology of the marine harmful bloom diatom Pseudo-nitzschia multiseries. Limnol. Oceanogr. 2011, 56, 829–840. [Google Scholar] [CrossRef] [Green Version]
- Van de Waal, D.B.; Verspagen, J.M.H.; Lürling, M.; Van Donk, E.; Visser, P.M.; Huisman, J. The ecological stoichiometry of toxins produced by harmful cyanobacteria: An experimental test of the carbon-nutrient balance hypothesis. Ecol. Lett. 2009, 12, 1326–1335. [Google Scholar] [CrossRef]
- Engström-Öst, J.; Hogfors, H.; El-Shehawy, R.; De Stasio, B.; Vehmaa, A.; Gorokhova, E. Toxin-producing cyanobacterium Nodularia spumigena, potential competitors and grazers: Testing mechanisms of reciprocal interactions. Aquat. Microb. Ecol. 2011, 62, 39–48. [Google Scholar] [CrossRef] [Green Version]
- Bergkvist, J.; Selander, E.; Pavia, H. Induction of toxin production in dinoflagellates: The grazer makes a difference. Oecologia 2008, 156, 147–154. [Google Scholar] [CrossRef] [PubMed]
- Selander, E.; Thor, P.; Toth, G.; Pavia, H. Copepods induce paralytic shellfish toxin production in marine dinoflagellates. Proc. R. Soc. B 2006, 273, 1673–1680. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, M.; Wang, X. Interactions between Microcystis aeruginosa and coexisting bisphenol A at different nitrogen levels. J. Hazard. Mater. 2019, 369, 132–141. [Google Scholar] [CrossRef]
- Chia, M.A.; Jankowiak, J.G.; Kramer, B.J.; Goleski, J.A.; Huang, I.S.; Zimba, P.V.; do Carmo Bittencourt-Oliveira, M.; Gobler, C.J. Succession and toxicity of Microcystis and Anabaena (Dolichospermum) blooms are controlled by nutrient-dependent allelopathic interactions. Harmful Algae 2018, 74, 67–77. [Google Scholar] [CrossRef]
- Zhou, Y.; Zhang, X.; Li, X.; Jia, P.; Dai, R. Evaluation of changes in Microcystis aeruginosa growth and microcystin production by urea via transcriptomic surveys. Sci. Total Environ. 2019, 655, 181–187. [Google Scholar] [CrossRef]
- Graneli, E.; Flynn, K. Chemical and physical factors influencing toxin content. In Ecology of Harmful Algae; Graneli, E., Turner, J.T., Eds.; Springer: Berlin/Heidelberg, Germany, 2006; Volume 189, pp. 229–242. [Google Scholar]
- Verspagen, J.M.H.; Van de Waal, D.B.; Finke, J.F.; Visser, P.M.; Huisman, J. Contrasting effects of rising CO2 on primary production and ecological stoichiometry at different nutrient levels. Ecol. Lett. 2014, 17, 951–960. [Google Scholar] [CrossRef] [Green Version]
- Eberlein, T.; Van de Waal, D.B.; Brandenburg, K.M.; John, U.; Voss, M.; Achterberg, E.P.; Rost, B. Interactive effects of ocean acidification and nitrogen limitation on two bloom-forming dinoflagellate species. Limnol. Oceanogr. 2016, 443, 127–140. [Google Scholar] [CrossRef] [Green Version]
- Huisman, J.; Arrayás, M.; Ebert, U.; Sommeijer, B. How do sinking phytoplankton species manage to persist? Am. Nat. 2002, 159, 245–254. [Google Scholar] [CrossRef]
- Pimentel, J.S.M.; Giani, A. Microcystin production and regulation under nutrient stress conditions in toxic Microcystis. Strains 2014, 80, 5836–5843. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ginn, H.P.; Pearson, L.A.; Neilan, B.A. NtcA from Microcystis aeruginosa PCC 7806 is autoregulatory and binds to the microcystin promoter. Appl. Environ. Microbiol. 2010, 76, 4362–4368. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rohrlack, T.; Dittmann, E.; Henning, M.; Börner, T.; Kohl, J.G. Role of microcystins in poisoning and food ingestion inhibition of Daphnia galeata caused by the cyanobacterium Microcystis aeruginosa. Appl. Environ. Microbiol. 1999, 65, 737–739. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Young, F.M.; Thomson, C.; Metcalf, J.S.; Lucocq, J.M.; Codd, G.A. Immunogold localisation of microcystins in cryosectioned cells of Microcystis. J. Struct. Biol. 2005, 151, 208–214. [Google Scholar] [CrossRef] [PubMed]
- Utkilen, H.; Gjolme, N. Iron-stimulated toxin production in Microcystis aeruginosa. Appl. Environ. Microbiol. 1995, 61, 797–800. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schatz, D.; Keren, Y.; Vardi, A.; Sukenik, A.; Carmeli, S.; Börner, T.; Dittmann, E.; Kaplan, A. Towards clarification of the biological role of microcystins, a family of cyanobacterial toxins. Environ. Microbiol. 2007, 9, 965–970. [Google Scholar] [CrossRef]
- Alexova, R.; Fujii, M.; Birch, D.; Cheng, J.; Waite, T.D.; Ferrari, B.C.; Neilan, B.A. Iron uptake and toxin synthesis in the bloom-forming Microcystis aeruginosa under iron limitation. Environ. Microbiol. 2011, 13, 1064–1077. [Google Scholar] [CrossRef]
- Qian, H.; Yu, S.; Sun, Z.; Xie, X.; Liu, W.; Fu, Z. Effects of copper sulfate, hydrogen peroxide and N-phenyl-2-naphthylamine on oxidative stress and the expression of genes involved photosynthesis and microcystin disposition in Microcystis aeruginosa. Aquat. Toxicol. 2010, 99, 405–412. [Google Scholar] [CrossRef]
- Latifi, A.; Ruiz, M.; Zhang, C.C. Oxidative stress in cyanobacteria. FEMS Microbiol. Rev. 2009, 33, 258–278. [Google Scholar] [CrossRef] [Green Version]
- Willis, A.; Adams, M.P.; Chuang, A.W.; Orr, P.T.; O’Brien, K.R.; Burford, M.A. Constitutive toxin production under various nitrogen and phosphorus regimes of three ecotypes of Cylindrospermopsis raciborskii ((Wołoszyńska) Seenayya et Subba Raju). Harmful Algae 2015, 47, 27–34. [Google Scholar] [CrossRef]
- Brauer, V.S.; Stomp, M.; Rosso, C.; Van Beusekom, S.A.M.; Emmerich, B.; Stal, L.J.; Huisman, J. Low temperature delays timing and enhances the cost of nitrogen fixation in the unicellular cyanobacterium Cyanothece. ISME J. 2013, 7, 2105–2115. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anderson, D.M.; Kulis, D.M.; Sullivan, J.J.; Hall, S.; Lee, C. Dynamics of physiology of saxitoxin production by the dinoflagellates Alexandrium spp. Mar. Biol. 1990, 104, 511–524. [Google Scholar] [CrossRef]
- Teegarden, G.J. Copepod grazing selection and particle discrimination on the basis of PSP toxin content. Mar. Ecol. Prog. Ser. 1999, 181, 163–176. [Google Scholar] [CrossRef]
- Colin, S.P.; Dam, H.G. Effects of the toxic dinoflagellate Alexandrium fundyense on the copepod Acartia hudsonica: A test of the mechanisms that reduce ingestion rates. Mar. Ecol. Prog. Ser. 2003, 248, 55–65. [Google Scholar] [CrossRef]
- Cembella, A.D. Chemical ecolgy of eukaryotic microalgae in marine ecosystems. Phycologia 2003, 42, 420–447. [Google Scholar] [CrossRef]
- Reguera, B.; Oshima, Y. Response of Gymnodinium catenatum to increasing levels of nitrate: Growth patterns and toxicity. In Toxic Marine Phytoplankton; Elsevier: New York, NY, USA, 1990. [Google Scholar]
- Allen, M.M.; Morris, R.; Zimmerman, W. Cyanophycin granule polypeptide protease in a unicellular cyanobacterium. Arch. Microbiol. 1984, 138, 119–123. [Google Scholar] [CrossRef]
- Van de Waal, D.B.; Verschoor, A.M.; Verspagen, J.M.H.; Van Donk, E.; Huisman, J. Climate-driven changes in the ecological stoichiometry of aquatic ecosystems. Front. Ecol. Environ. 2010, 8, 145–152. [Google Scholar] [CrossRef] [Green Version]
- Dai, R.; Liu, H.; Qu, J.; Zhao, X.; Ru, J.; Hou, Y. Relationship of energy charge and toxin content of Microcystis aeruginosa in nitrogen-limited or phosphorous-limited cultures. Toxicon 2008, 51, 649–658. [Google Scholar] [CrossRef]
- Proctor, N.H.; Chan, S.L.; Trevor, A.J. Production of saxitoxin by cultures of Gonyaulax catenella. Toxicon 1975, 13, 1–9. [Google Scholar] [CrossRef]
- Taroncher-Oldenburg, G.; Kulis, D.M.; Anderson, D.M. Coupling of saxitoxin biosynthesis to the G1 phase of the cell cycle in the dinoflagellate Alexandriun fundyense: Temperature and nutrient effects. Nat. Toxins 1999, 7, 207–219. [Google Scholar] [CrossRef]
- Selander, E.; Cervin, G.; Pavia, H. Effects of nitrate and phosphate on grazer-induced toxin production in Alexandrium minutum. Limnol. Oceanogr. 2008, 53, 523–530. [Google Scholar] [CrossRef]
- Frenken, T.; Wierenga, J.; Gsell, A.S.; van Donk, E.; Rohrlack, T.; Van de Waal, D.B. Changes in N:P supply ratios affect the ecological stoichiometry of a toxic cyanobacterium and its fungal parasite. Front. Microbiol. 2017, 8, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mowe, M.A.D.; Abbas, F.; Porojan, C.; Mitrovic, S.M.; Lim, R.P.; Furey, A.; Yeo, D.C.J. Roles of nitrogen and phosphorus in growth responses and toxin production (using LC-MS/MS) of tropical Microcystis ichthyoblabe and M. flos-aquae. J. Appl. Phycol. 2016, 28, 1543–1552. [Google Scholar] [CrossRef]
- Burkholder, J.M.; Glibert, P.M.; Skelton, H.M. Mixotrophy, a major mode of nutrition for harmful algal species in eutrophic waters. Harmful Algae 2008, 8, 77–93. [Google Scholar] [CrossRef]
- Stoecker, D.K. Mixotrophy among Dinoflagellates. J. Eukaryot. Microbiol. 1999, 46, 397–401. [Google Scholar] [CrossRef]
- Jacobson, D.M.; Anderson, D.M. Widespread phagocytosis of ciliates and other protists by marine mixotrophic and heterotrophic thecate dinoflagellates. J. Phycol. 1996, 32, 279–285. [Google Scholar] [CrossRef]
- Grzebyk, D.; Bechemin, C.; Ward, C.J.; Verite, C.; Codd, G.A.; Maestrini, S.Y. Effects of salinity and two coastal waters on the growth and toxin content of the dinoflagellate Alexandrium minutum. J. Plankton Res. 2003, 25, 1185–1199. [Google Scholar] [CrossRef]
- Navarro, J.M.; Munoz, M.G.; Contreras, A.M. Temperature as a factor regulating growth and toxin content in the dinoflagellate Alexandrium catenella. Harmful Algae 2006, 5, 762–769. [Google Scholar] [CrossRef]
- Engström-Öst, J.; Repka, S.; Mikkonen, M. Interactions between plankton and cyanobacterium Anabaena with focus on salinity, growth and toxin production. Harmful Algae 2011, 10, 530–535. [Google Scholar] [CrossRef]
- Gedaria, A.I.; Luckas, B.; Reinhardt, K.; Azanza, R.V. Growth response and toxin concentration of cultured Pyrodinium bahamense var. compressum to varying salinity and temperature conditions. Toxicon 2007, 50, 518–529. [Google Scholar] [CrossRef]
- Song, L.; Sano, T.; Li, R.; Watanabe, M.M.; Liu, Y.; Kaya, K. Microcystin production of Microcystis viridis (cyanobacteria) under different culture conditions. Phycol. Res. 1998, 46, 19–23. [Google Scholar] [CrossRef]
- Tatters, A.O.; Flewelling, L.J.; Fu, F.; Granholm, A.A.; Hutchins, D.A. High CO2 promotes the production of paralytic shellfish poisoning toxins by Alexandrium catenella from Southern California waters. Harmful Algae 2013, 30, 37–43. [Google Scholar] [CrossRef]
- Dolman, A.M.; Rücker, J.; Pick, F.R.; Fastner, J.; Rohrlack, T.; Mischke, U.; Wiedner, C. Cyanobacteria and cyanotoxins: The influence of nitrogen versus phosphorus. PLoS ONE 2012, 7, e38757. [Google Scholar] [CrossRef] [PubMed]
- Smith, V.H. Eutrophication of freshwater and coastal marine ecosystems a global problem. Environ. Sci. Pollut. Res. 2003, 10, 126–139. [Google Scholar] [CrossRef] [PubMed]
- Heisler, J.; Glibert, P.M.; Burkholder, J.M.; Anderson, D.M.; Cochlan, W.; Dennison, W.C.; Dortch, Q.; Gobler, C.J.; Heil, C.A.; Humphries, E.; et al. Eutrophication and harmful algal blooms: A scientific consensus. Harmful Algae 2008, 8, 3–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McLellan, N.L.; Manderville, R.A. Toxic mechanisms of microcystins in mammals. Toxicol. Res. 2017, 6, 391–405. [Google Scholar] [CrossRef] [Green Version]
- Vale, P. New saxitoxin analogues in the marine environment: Developments in toxin chemistry, detection and biotransformation during the 2000s. Phytochem. Rev. 2010, 9, 525–535. [Google Scholar] [CrossRef]
- Shimizu, Y. Toxigenesis and biosynthesis of saxitoxin analogues. Pure Appl. Chem. 1986, 58, 257–262. [Google Scholar] [CrossRef]
- Gobler, C.J.; Burkholder, J.A.M.; Davis, T.W.; Harke, M.J.; Johengen, T.; Stow, C.A.; Van de Waal, D.B. The dual role of nitrogen supply in controlling the growth and toxicity of cyanobacterial blooms. Harmful Algae 2016, 54, 87–97. [Google Scholar] [CrossRef]
- Rinehart, K.L.; Namikoshi, M.; Choi, B.W. Structure and biosynthesis of toxins from blue-green algae (cyanobacteria). J. Appl. Phycol. 1994, 6, 159–176. [Google Scholar] [CrossRef]
- Murata, A.; Nagashima, Y.; Taguchi, S. N:P ratios controlling the growth of the marine dinoflagellate Alexandrium tamarense: Content and composition of paralytic shellfish poison. Harmful Algae 2012, 20, 11–18. [Google Scholar] [CrossRef]
- Paerl, H.W.; Huisman, J. Climate change: A catalyst for global expansion of harmful cyanobacterial blooms. Environ. Microbiol. Rep. 2009, 1, 27–37. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, M.; Muftakhidinov, B.; Winchen, T. Engauge Digitizer, version 4.1; Github: San Francisco, CA, USA, 1991. [Google Scholar]
- Lajeunesse, M.J. Bias and correction for the log response ratio in ecological meta-analysis. Ecology 2015, 96, 2056–2063. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- R Core Team. R: A Language and Environment for Statistical Computing; R Core Team: Vienna, Austria, 2018. [Google Scholar]
- Viechtbauer, W. Conducting meta-analyses in R with the metafor package. J. Stat. Softw. 2010, 36, 1–48. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Toxin (Short Name) | (Full Name) | C:N Ratio |
|---|---|---|
| PSP | Paralytic shellfish poisoning toxins | 1.5 |
| CYN | Cylindrospermopsin | 3.0 |
| MC | Microcystin (MC-LR; MC-RR) | 4.3 |
| NOD | Nodularin | 5.1 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Brandenburg, K.; Siebers, L.; Keuskamp, J.; Jephcott, T.G.; Van de Waal, D.B. Effects of Nutrient Limitation on the Synthesis of N-Rich Phytoplankton Toxins: A Meta-Analysis. Toxins 2020, 12, 221. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins12040221
Brandenburg K, Siebers L, Keuskamp J, Jephcott TG, Van de Waal DB. Effects of Nutrient Limitation on the Synthesis of N-Rich Phytoplankton Toxins: A Meta-Analysis. Toxins. 2020; 12(4):221. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins12040221
Chicago/Turabian StyleBrandenburg, Karen, Laura Siebers, Joost Keuskamp, Thomas G. Jephcott, and Dedmer B. Van de Waal. 2020. "Effects of Nutrient Limitation on the Synthesis of N-Rich Phytoplankton Toxins: A Meta-Analysis" Toxins 12, no. 4: 221. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins12040221