Blooms of Toxic Cyanobacterium Nodularia spumigena in Norwegian Fjords During Holocene Warm Periods

 , ,
, ,  ,
,
Abstract
:1. Introduction
2. Results
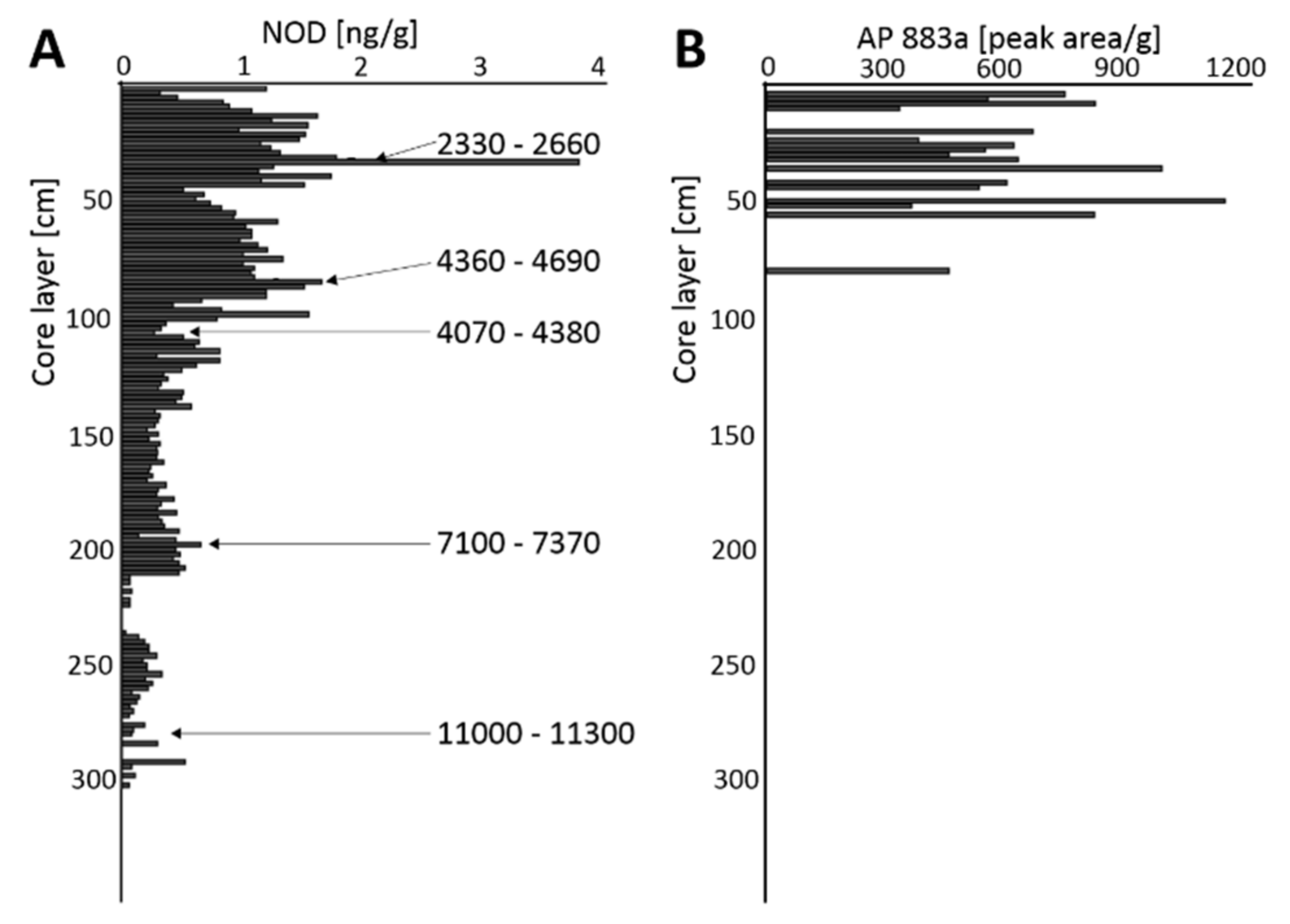
2.1. N. spumigena Chemical Markers in Sediment Samples
2.2. Genetic Markers in Sediment Samples
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Sediment Sampling
5.2. Extraction and LC-MS/MS Analysis
5.3. Genetic Analysis
5.4. Sediment 14C-Dating
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Johnsen, G.; Sakshaug, E. Monitoring of harmful algal blooms along the Norwegian Coast using bio-optical methods. S. Afr. J. Mar. Sci. 2000, 22, 309–321. [Google Scholar] [CrossRef]
- Bratbak, G.; Jacquet, S.; Larsen, A.; Pettersson, L.H.; Sazhin, A.F.; Thyrhaug, R. The plankton community in Norwegian coastal waters-abundance, composition, spatial distribution and diel variation. Cont. Shelf Res. 2011, 31, 1500–1514. [Google Scholar] [CrossRef]
- Dittami, S.M.; Hostyeva, V.; Egge, E.S.; Kegel, J.U.; Eikrem, W.; Edvardsen, B. Seasonal dynamics of harmful algae in outer Oslofjorden monitored by microarray, qPCR, and microscopy. Environ. Sci. Pollut. Res. Int. 2013, 20, 6719–6732. [Google Scholar] [CrossRef] [PubMed]
- Lännergren, C. Nitrogen fixation by blue-green algae in a Norwegian land-locked fjord, determined by the acetylene reduction technique. Sarsia 1980, 65, 73–77. [Google Scholar] [CrossRef]
- Kataržytė, M.; Vaičiūtė, D.; Nasvytis, P. Excellent bathing waters in coastal areas: Is microbial pollution the only important parameter? Ocean Coast. Manage. 2019, 182, 104922. [Google Scholar] [CrossRef]
- Stal, L.J.; Albertano, P.; Bergman, B.; von Bröckel, K.; Gallon, J.R.; Hayes, P.K.; Sivonen, K.; Walsby, A.E. BASIC: Baltic Sea cyanobacteria. An investigation of the structure and dynamics of water blooms of cyanobacteria in the Baltic Sea-responses to a changing environment. Cont. Shelf Res. 2003, 23, 1695–1714. [Google Scholar] [CrossRef]
- McGregor, G.B.; Stewart, I.; Sendall, B.C.; Sadler, R.; Reardon, K.; Carter, S.; Wruck, D.; Wickramasinghe, W. First Report of a Toxic Nodularia spumigena (Nostocales/Cyanobacteria) Bloom in Sub-Tropical Australia. I. Phycological and Public Health Investigations. Int. J. Environ. Res. Public Health 2012, 9, 2396–2411. [Google Scholar] [CrossRef] [Green Version]
- Woodward, C.A.; Shulmeister, J. A Holocene record of human induced and natural environmental change from Lake Forsyth (Te Wairewa), New Zealand. J. Paleolimnol. 2005, 34, 481–501. [Google Scholar] [CrossRef]
- Nehring, S. Mortality of dogs associated with a mass development of Nodularia spumigena (Cyanophyceae) in a brackish lake at the German North Sea Coast. J. Plankton Res. 1993, 15, 867–872. [Google Scholar] [CrossRef]
- Descy, J.P.; Stoyneva-Gärtner, M.P.; Uzunov, B.A.; Dimitrova, P.H.; Pavlova, V.T.; Gärtner, G. Studies on cyanoprokaryotes of the water bodies along the Bulgarian Black Sea Coast (1890–2017): A review, with special reference to new, rare and harmful taxa. Acta Zool. Bulg. 2018, 11, 43–52. [Google Scholar]
- Fewer, D.P.; Jokela, J.; Rouhlainen, L.; Wahlsten, M.; Koskenniemi, K.; Stal, L.J.; Sivonen, K. The non-ribosomal assembly and frequent occurrence of the protease inhibitors spumigins in the bloom-forming cyanobacteria Nodularia spumigena. Mol. Microbiol. 2009, 73, 924–937. [Google Scholar] [CrossRef]
- Mazur-Marzec, H.; Bertos-Fortis, M.; Toruńska-Sitarz, A.; Fidor, A.; Legrand, C. Chemical and Genetic Diversity of Nodularia spumigena from the Baltic Sea. Mar. Drugs. 2016, 14, E209. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cegłowska, M.; Toruńska-Sitarz, A.; Kowalewska, G.; Mazur-Marzec, H. Specific Chemical and Genetic Markers Revealed a Thousand-Years Presence of Toxic Nodularia spumigena in the Baltic Sea. Mar. Drugs. 2018, 16, 116. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mazur, H.; Pliński, M. Nodularia spumigena blooms and the occurrence of hepatotoxin in the Gulf of Gdańsk. Oceanologia 2003, 45, 305–316. [Google Scholar]
- Mazur-Marzec, H.; Krężel, A.; Kobos, J.; Pliński, M. Toxic Nodularia spumigena blooms in the coastal waters of the Gulf of Gdańsk: A ten-year survey. Oceanologia 2006, 48, 255–273. [Google Scholar]
- Jokela, J.; Helnillä, L.M.P.; Shishido, T.K.; Wahlsten, M.; Fewer, D.P.; Fiore, M.F.; Wang, H.; Haapaniemi, E.; Permi, P.; Sivonen, K. Production of High Amounts of Hepatotoxin Nodularin and New Protease Inhibitors Pseudospumigins by the Brazilian Benthic Nostoc sp. CENA543. Front. Microbiol. 2017, 8, 1963. [Google Scholar] [CrossRef] [PubMed]
- Kaasalainen, U.; Fewer, D.P.; Jokela, J.; Wahlsten, M.; Sivonen, K.; Rikkinen, J. Cyanobacteria produce a high variety of hepatotoxic peptides in lichen symbiosis. Proc. Natl. Acad. Sci. USA 2012, 109, 5886–5891. [Google Scholar] [CrossRef] [Green Version]
- Gehringer, M.M.; Adler, L.; Roberts, A.A.; Moffitt, M.C.; Mihali, T.K.; Mills, T.J.T.; Fieker, C.; Neilan, B.A. Nodularin, a cyanobacterial toxin, is synthesized in planta by symbiotic Nostoc sp. ISME J. 2012, 6, 1834–1847. [Google Scholar] [CrossRef] [Green Version]
- McGregor, G.B.; Sendall, B.C. Iningainema pulvinus gen nov., sp nov. (Cyanobacteria, Scytonemataceae) a new nodularin producer from Edgbaston Reserve, north-eastern Australia. Harmful Algae 2017, 62, 10–19. [Google Scholar] [CrossRef]
- Teikari, J.E.; Fewer, D.P.; Shrestha, R.; Hou, S.; Leikoski, N.; Mäkelä, M.; Simojoki, A.; Hess, W.R.; Sivonen, K. Strains of the toxic and bloom-forming Nodularia spumigena (cyanobacteria) can degrade methylphosphonate and release methane. ISME J. 2018, 12, 1619–1630. [Google Scholar] [CrossRef] [Green Version]
- Kankaanpää, H.T.; Sjövall, O.; Huttunen, M.; Olin, M.; Karlsson, K.; Hyvärinen, K.; Sneitz, L.; Härkönen, J.; Sipiä, V.O.; Meriluoto, J.A. Production and sedimentation of peptide toxins nodularin-R and microcystin-LR in the northern Baltic Sea. Environ. Pollut. 2009, 157, 1301–1309. [Google Scholar] [CrossRef] [PubMed]
- Seidenkrantz, M.S.; Aagaard-Sørensen, S.; Sulsbrück, H.; Kuijpers, A.; Jensen, K.G.; Kunzendorf, H. Hydrography and climate of the last 4400 years in a SW Greenland fjord: Implication for Labrador Sea palaeoceanography. Holocene 2007, 17, 387–401. [Google Scholar] [CrossRef]
- Wang, T.; Surge, D.; Mithen, S. Seasonal temperature variability of the Neoglacial (3300–2500 BP) and Roman Warm Period (2500–1600 BP) reconstructed from oxygen isotope ratios of limpet shells (Patella vulgata), Northwest Scotland. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2012, 317-318, 104–113. [Google Scholar] [CrossRef]
- Kuylenstierna, M.; Karlson, B. Seasonality and Composition of Pico- and Nanoplanktonic Cyanobacteria and Protists in the Skagerrak. Bot. Mar. 1994, 37, 17–33. [Google Scholar] [CrossRef]
- Larsen, A.; Flaten, G.A.F.; Sandaa, R.A.; Castberg, T.; Thyrhaug, R.; Erga, S.R.; Jacquet, S.; Bratbak, G. Spring phytoplankton bloom dynamics in Norwegian coastal waters: Microbial community succession and diversity. Limnol. Oceanogr. 2004, 49, 180–190. [Google Scholar] [CrossRef] [Green Version]
- Davis, B.A.S.; Brewer, S.; Stevenson, A.C.; Guiot, J. The temperature of Europe during the Holocene reconstructed from pollen data. Quat. Sci. Rev. 2003, 22, 15–17. [Google Scholar] [CrossRef]
- Seppä, H.; Hammarlund, D.; Antonsson, K. Low-frequency and high-frequency changes in temperature and effective humidity during the Holocene in south-central Sweden: Implications for atmospheric and oceanic forcings of climate. Clim. Dyn. 2005, 25, 285–297. [Google Scholar] [CrossRef]
- Mauri, A.; Davis, B.A.S.; Collins, P.M.; Kaplan, J.O. The climate of Europe during the Holocene: A gridded pollen-based reconstruction and its multi-proxy evaluation. Quat. Sci. Rev. 2015, 112, 109–127. [Google Scholar] [CrossRef]
- Szymczak-Żyła, M.; Krajewska, M.; Witak, M.; Ciesielski, T.M.; Ardelan, M.V.; Jenssen, B.M.; Goslar, T.; Winogradow, A.; Filipkowska, A.; Lubecki, L.; et al. Present and Past-Millennial Eutrophication in the Gulf of Gdańsk (southern Baltic Sea). Paleoceanogr. Paleocl. 2019, 34, 136–152. [Google Scholar] [CrossRef] [Green Version]
- Wang, T.; Surge, D.; Walker, K.J. Seasonal climate change across the Roman Warm Period/Vandal Minimum transition using isotope sclerochronology in archaeological shells and otoliths, southwest Florida, USA. Quatern. Int. 2013, 308-309, 230–241. [Google Scholar] [CrossRef]
- Hong, J.W.; Choi, H.G.; Kang, S.H.; Yoon, H.S. Axenic purification and cultivation of an Arctic cyanobacterium, Nodularia spumigena KNUA005, with cold tolerance potential for sustainable production of algae-based biofuel. Algae 2010, 25, 99–104. [Google Scholar] [CrossRef]
- Taton, A.; Grubisic, S.; Brambilla, E.; De Wit, R.; Wilmotte, A. Cyanobacterial Diversity in Natural and Artificial Microbial Mats of Lake Fryxell (McMurdo Dry Valleys, Antarctica): A Morphological and Molecular Approach. Appl. Environ. Microbiol. 2003, 69, 5157–5169. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hassan, N.; Anesio, A.M.; Rafiq, M.; Holtvoeth, J.; Bull, I.; Williamson, C.J.; Hasan, F. Cell membrane fatty acid and pigment composition of the psychrotolerant cyanobacterium Nodularia spumigena CHS1 isolated from Hopar glacier, Pakistan. Extremophiles 2020, 24, 135–145. [Google Scholar] [CrossRef] [PubMed]
- Chrismas, N.A.; Anesio, A.M.; Sánchez-Baracaldo, P. Multiple adaptations to polar and alpine environments within cyanobacteria: A phylogenomic and Bayesian approach. Front. Microbiol. 2015, 6, 1070. [Google Scholar] [CrossRef] [Green Version]
- Lyra, K.; Sinkko, H.; Rantanen, M.; Paulin, L.; Kotilainen, A. Sediment bacterial communities reflect the history of a Sea Basin. PLoS ONE 2013, 8, e54326. [Google Scholar] [CrossRef] [Green Version]
- Monchamp, M.E.; Walser, J.C.; Pomati, F.; Spaak, P. Sedimentary DNA reveals cyanobacterial community diversity over 200 years in two perialpine lakes. Appl. Environ. Microbiol. 2016, 82, 6472–6482. [Google Scholar] [CrossRef] [Green Version]
- Legrand, B.; Miras, Y.; Beauger, A.; Dussauze, M.; Latour, D. Akinetes and ancient DNA reveal toxic cyanobacterial recurrences and their potential for resurrection in a 6700-year-old core from a eutrophic lake. Sci. Total. Environ. 2019, 687, 1369–1380. [Google Scholar] [CrossRef]
- Rinta-Kanto, J.M.; Saxton, M.; DeBruym, J.M.; Smith, J.L.; Marvin, C.H.; Krieger, K.A.; Sayler, G.S.; Boyer, G.L.; Wilhelm, S.W. The diversity and distribution of toxygenic Microcystis spp. in the present day and archived pelagic and sediment samples from Lake Erie. Harmful Algae 2009, 8, 385–394. [Google Scholar] [CrossRef]
- Lindahl, T. Instability and decay of the primary structure of DNA. Nature 1993, 362, 709–715. [Google Scholar] [CrossRef]
- Moffitt, M.C.; Neilan, B.A. Characterization of the Nodularin Synthetase Gene Cluster and Proposed Theory of the Evolution of Cyanobacterial Hepatotoxins. Appl. Environ. Microbiol. 2004, 70, 6353–6362. [Google Scholar] [CrossRef] [Green Version]
- Jungblut, A.D.; Hawes, I.; Mountfort, D.; Hitzfeld, B.; Dietrich, D.R.; Burns, B.P.; Neilan, B.A. Diversity within cyanobacterial mat communities in variable salinity meltwater ponds of McMurdo Ice Shelf, Antarctica. Environ. Microbiol. 2005, 7, 519–529. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nübel, U.; Garcia-Pichel, F.; Muyzer, G. PCR primers to amplify 16S rRNA genes from cyanobacteria. Appl. Environ. Microbiol. 1997, 63, 3327–3332. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Neilan, B.A.; Jacobs, D.; Goodman, A.E. Genetic diversity and phylogeny of toxic cyanobacteria determined by DNA polymorphisms within the phycocyanin locus. Appl. Environ. Microbiol. 1995, 61, 3875–3883. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jungblut, A.; Neilan, B.A. Molecular identification and evolution of the cyclic peptide hepatotoxins, microcystin and nodularin, synthetase gene in three orders of cyanobacteria. Arch. Microbiol. 2006, 185, 107–114. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis Version 7.0 for Bigger Datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reimer, P.J.; Bard, E.; Bayliss, A.; Beck, J.W.; Blackwell, P.G.; Bronk Ramsey, C.; Buck, C.E.; Cheng, H.; Edwards, R.L.; Friedrich, M.; et al. IntCal13 and MARINE13 radiocarbon age calibration curves 0–50,000 years calBP. Radiocarbon 2013, 55, 1869–1887. [Google Scholar] [CrossRef] [Green Version]
- Stuiver, M.; Reimer, P.J.; Reimer, R.W. CALIB. 2020. Available online: http://calib.org (accessed on 31 March 2020).




{kind=link}
{kind=link}
{kind=link}
| Sample | Sediment Layer [cm] | DNA [ng/µL] | A260/280 | 16S rDNA | cpcBA-IGS | mcyE/ndaF |
|---|---|---|---|---|---|---|
| N. spumigena CCNP1401 | + | + | + | |||
| N. spumigena CCY9414 | + | + | + | |||
| Oslofjorden SC | 0–1 | 242 | 1.2 | + | - | - |
| 1–5 | 139 | 1.3 | + | - | - | |
| 5–10 | 140 | 1.3 | + | - | - | |
| 10–15 | 210 | 1.3 | - | - | - | |
| 15–20 | 100 | 1.3 | + | - | - | |
| Oslofjorden LC | 32–34 | 72 | 1.2 | + | + | - |
| 34–36 | 118 | 1.4 | + | + | + | |
| 42–44 | 75 | 1.2 | + | - | + | |
| 68–70 | 44 | 1.3 | - | + | - | |
| 84–86 | 70 | 1.2 | + | + | + | |
| 88–90 | 71 | 1.2 | + | + | - | |
| 114–116 | 85 | 1.1 | + | + | + | |
| 126–128 | 91 | 1.2 | + | + | + | |
| 128–130 | 96 | 1.2 | - | + | + | |
| 146–148 | 140 | 1.1 | - | - | + | |
| 148–150 | 132 | 1.2 | + | + | + | |
| 178–180 | 62 | 1.3 | + | + | + | |
| 180–182 | 105 | 1.2 | + | + | - | |
| 224–226 | 86 | 1.2 | + | - | - | |
| 232–234 | 40 | 1.1 | + | + | + | |
| 260–262 | 19 | 1.4 | + | + | - | |
| 262–264 | 50 | 1.2 | + | - | + | |
| 314–316 | 47 | 1.2 | + | + | - | |
| 334–336 | 46 | 1.2 | + | + | - | |
| 336–338 | 31 | 1.2 | + | + | - | |
| 338–340 | 40 | 1.3 | + | + | + | |
| 362–364 | 31 | 1.3 | + | - | + | |
| 364–366 | 38 | 1.8 | + | - | - | |
| Balsfjorden LC * | 152–154 | 64 | 1.2 | + | - | - |
| 220–222 | 45 | 1.2 | + | - | - | |
| 230–232 | 177 | 1.2 | + | - | - | |
| Trondheimsfjorden LC * | 92–94 | 65 | 1.0 | + | + | - |
| 134–136 | - | - | - | - | - |
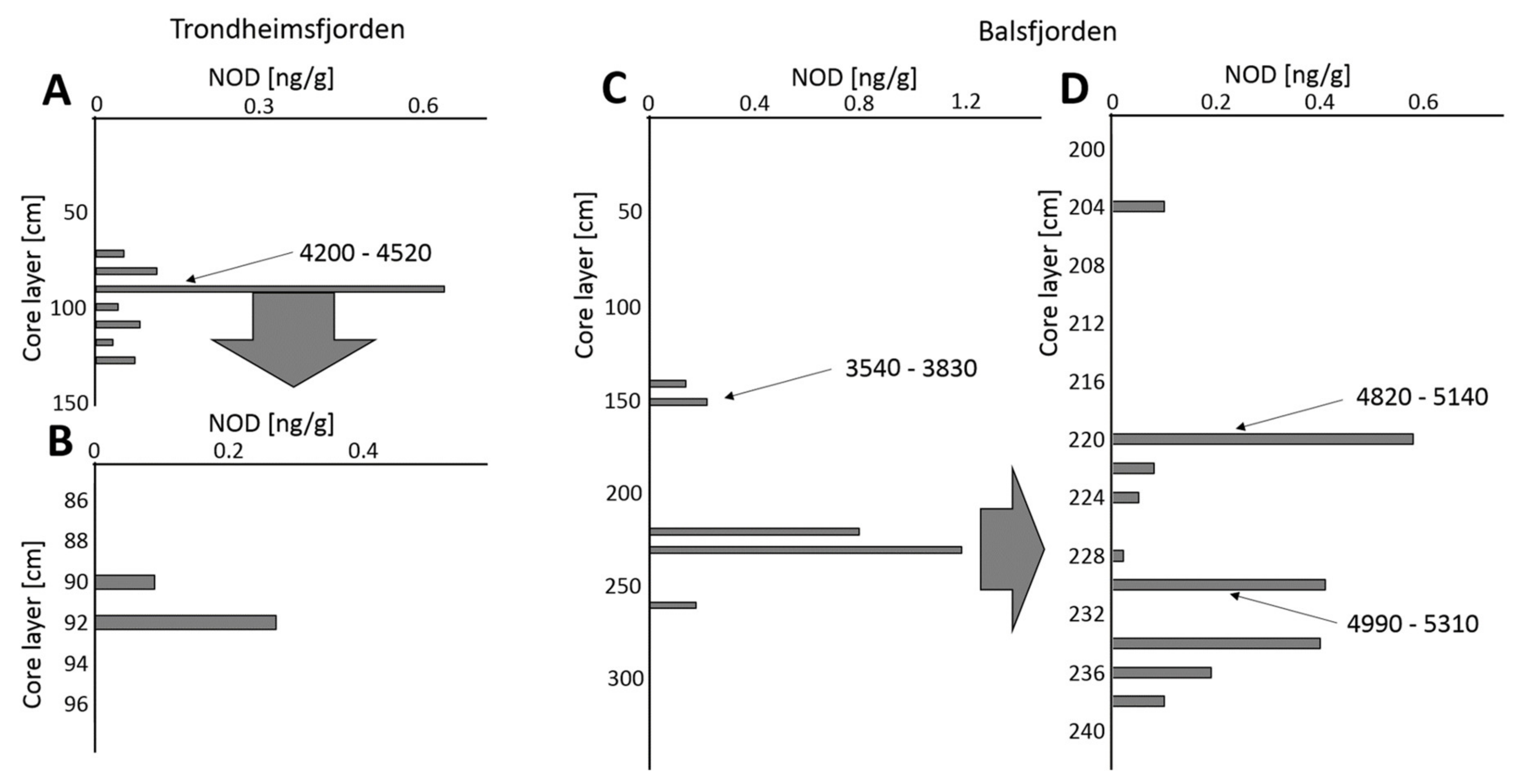
| Presenceof NOD in the Sediment Core | Norwegian Fjords | Baltic Sea | ||
|---|---|---|---|---|
| Balsfjorden | Trondheimsfjorden | Oslofjorden | Gdańsk Depth | |
| NOD Peaks [cal yr BP] | 2330–2660 | ~2500 * | ||
| 3540–3830 | 4200–4520 | 4360–4690 | ~4200 * | |
| 7100–7370 | ||||
| 4820–5140 | 1100–1130 | |||
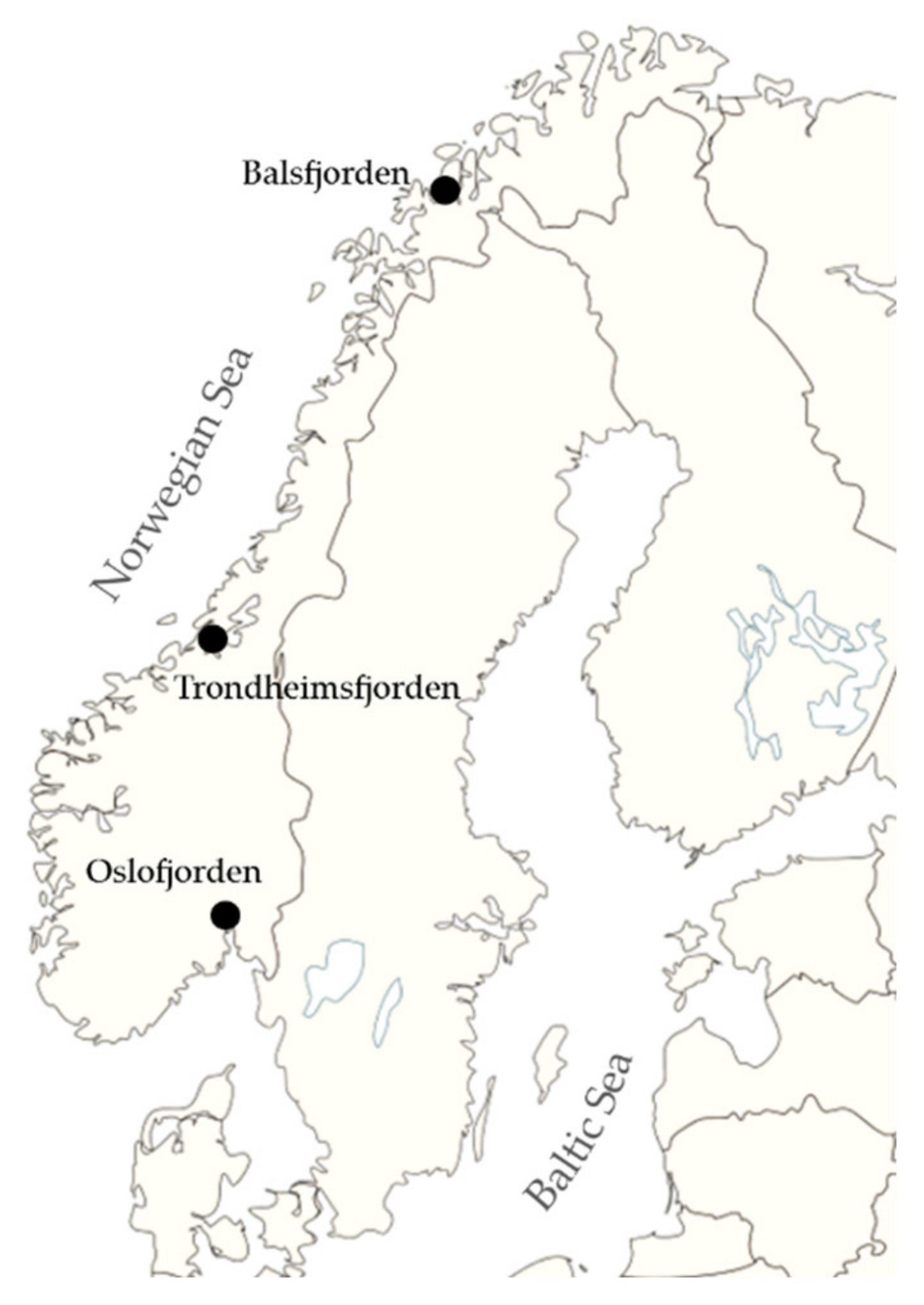
| Sampling Station | Depth [m] | Core Length [cm] |
|---|---|---|
| Oslofjorden 59° 50.648′ N; 10° 43.560′ E | 77 | 336 |
| Trondheimsfjorden 63° 28.370′ N; 10° 11.650′ E | 502 | 136 |
| Balsfjorden 69° 17.361′ N; 19° 22.586′ E | 112 | 440 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Konkel, R.; Toruńska-Sitarz, A.; Cegłowska, M.; Ežerinskis, Ž.; Šapolaitė, J.; Mažeika, J.; Mazur-Marzec, H. Blooms of Toxic Cyanobacterium Nodularia spumigena in Norwegian Fjords During Holocene Warm Periods. Toxins 2020, 12, 257. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins12040257
Konkel R, Toruńska-Sitarz A, Cegłowska M, Ežerinskis Ž, Šapolaitė J, Mažeika J, Mazur-Marzec H. Blooms of Toxic Cyanobacterium Nodularia spumigena in Norwegian Fjords During Holocene Warm Periods. Toxins. 2020; 12(4):257. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins12040257
Chicago/Turabian StyleKonkel, Robert, Anna Toruńska-Sitarz, Marta Cegłowska, Žilvinas Ežerinskis, Justina Šapolaitė, Jonas Mažeika, and Hanna Mazur-Marzec. 2020. "Blooms of Toxic Cyanobacterium Nodularia spumigena in Norwegian Fjords During Holocene Warm Periods" Toxins 12, no. 4: 257. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins12040257