Molecular Identification and Mycotoxin Production by Alternaria Species Occurring on Durum Wheat, Showing Black Point Symptoms

 , ,
, ,  ,
, 
Abstract
:1. Introduction
- (a)
- Identify the main fungal species infecting kernel wheat samples showing black point symptoms, collected in Italy;
- (b)
- Detect the presence of AOH in wheat kernels in the field;
- (c)
- Study the genetic variability of the Alternaria species isolated, by sequencing the genes coding for allergen alt a1 (alt-a1), glyceraldeyde-3-phosphate dehydrogenase (gpd) and translation elongation factor 1-α (tef), by following a multi-locus gene sequence approach;
- (d)
- Analyze the mycotoxin profile of selected strains of the Alternaria species identified.
2. Results
2.1. Black Point Disease Symptoms on Wheat Samples
2.2. Alternariol Contamination of Wheat Samples
2.3. Detection of Fungal Infection
2.4. Morphological Characterization
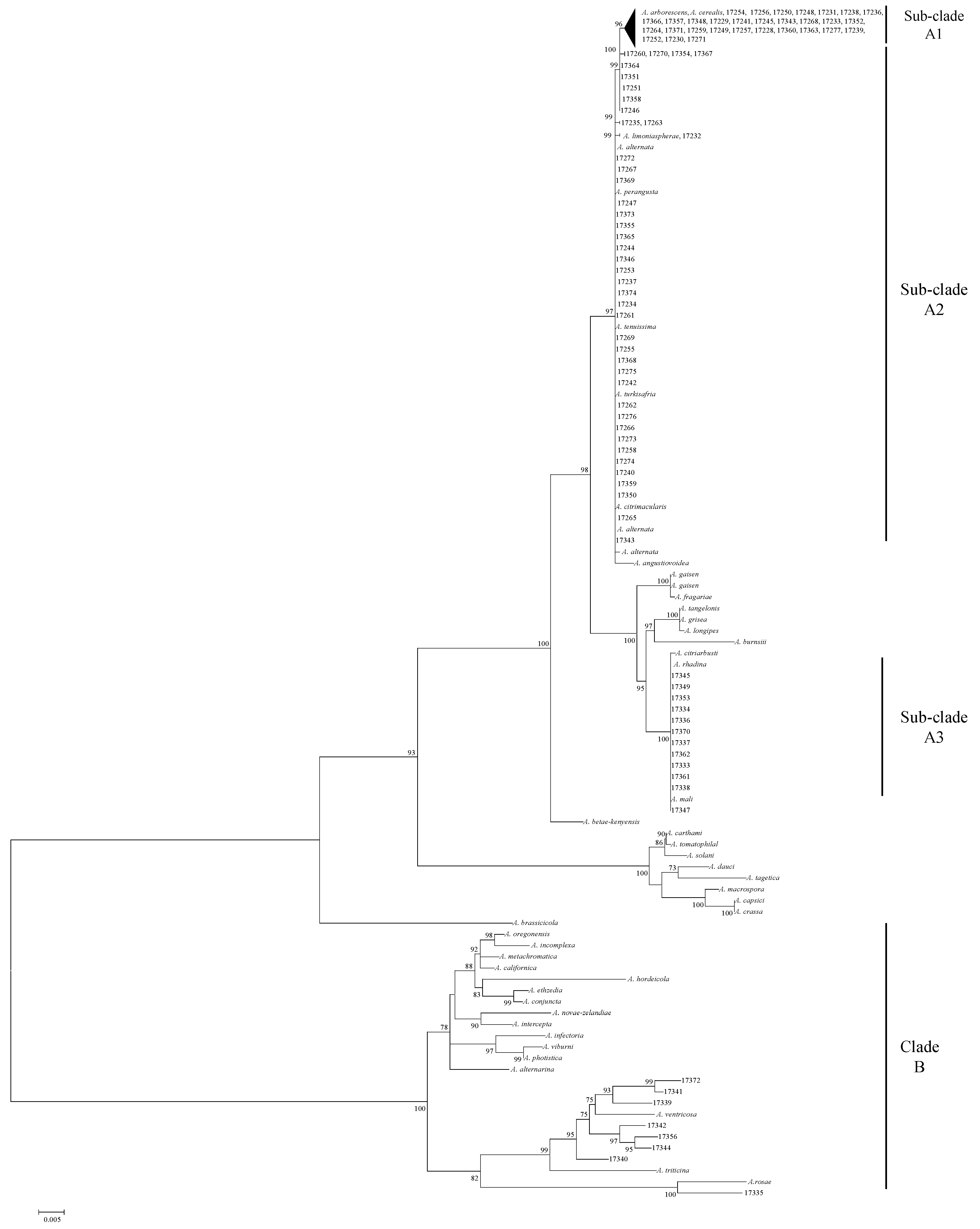
2.5. PCR Amplification and Phylogenetic Analysis
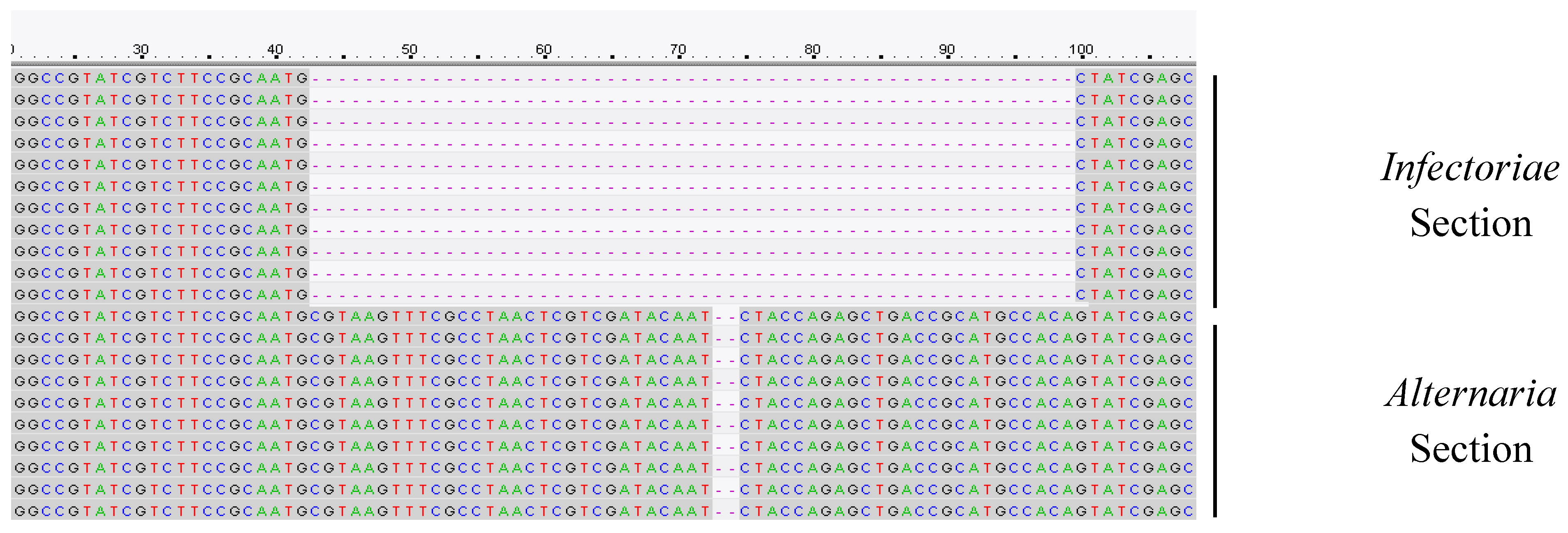
2.6. Design of Infectoriae Section Specific PCR Primers
2.7. Mycotoxins Production of Alternaria Strains
3. Discussion
4. Materials and Methods
4.1. Origin of Wheat Samples
4.2. Evaluation of Black Point Symptoms
4.3. Fungal Isolation and Growth Conditions
4.4. Molecular Characterization of Alternaria Strains
4.5. Mycotoxin Extraction
HPLC Analysis
Author Contributions
Funding
Conflicts of Interest
References
- FAOSTAT. Available online: www.fao.org (accessed on 25 January 2020).
- Conner, R.L.; Davidson, J.G.N. Resistance in wheat to black point caused by Alternaria alternata and Cochliobolus sativus. Can. J. Plant Sci. 1988, 68, 351–359. [Google Scholar] [CrossRef]
- Fernandez, M.R.; Conner, R.L. Black Point and Smudge in Wheat. Prairie Soil Crop J. 2011, 4, 158–164. [Google Scholar]
- Srivastava, J.P.; Kushwaha, G.D.; Shukla, D.N. Black point disease of wheat and its implications on seed quality. Crop Res. 2014, 47, 21–23. [Google Scholar]
- Hudec, K.; Muchova, D. Correlation between black point symptoms and fungal infestation and seedling viability of wheat kernels. Plant Prot. Sci. 2008, 44, 138–146. [Google Scholar] [CrossRef] [Green Version]
- Abdullah, S.; Atrosh, H. New Records of Fungi on Wheat Grains From Iraq. Sci. J. Univ. Zakho 2014, 2, 256–265. [Google Scholar] [CrossRef]
- Ramires, F.A.; Masiello, M.; Somma, S.; Villani, A.; Susca, A.; Logrieco, A.F.; Luz, C.; Meca, G.; Moretti, A. Phylogeny and Mycotoxin Characterization of Alternaria Species Isolated from Wheat Grown in Tuscany, Italy. Toxins 2018, 10, 472. [Google Scholar] [CrossRef] [Green Version]
- Logrieco, A.; Moretti, A.; Solfrizzo, M. Alternaria toxins and plant diseases: And overview of origin, occurrence and risks. World Mycotoxin J. 2009, 2, 129–140. [Google Scholar] [CrossRef]
- Logrieco, A.; Bottalico, A.; Mulè, G.; Moretti, A.; Perrone, G. Epidemiology of toxigenic fungi and their associated mycotoxins for some Mediterranean crops. Eur. J. Plant Pathol. 2003, 109, 645–667. [Google Scholar] [CrossRef]
- EFSA CONTAM Panel (EFSA Panel on Contaminants in the Food Chain). Scientific Opinion on the risks for animal and public health related to the presence of Alternaria toxins in feed and food. Efsa J. 2011, 9, 2407. [Google Scholar] [CrossRef]
- Arcella, D.; Eskola, M.; Gómez Ruiz, J.A. Scientific report on the dietary exposure assessment to Alternaria toxins in the European population. EFSA J. 2016, 14, 4654. [Google Scholar] [CrossRef]
- Amatulli, M.T.; Fanelli, F.; Moretti, A.; Mulè, G.; Logrieco, A.F. Alternaria species and mycotoxins associated to black point of cereals. Mycotoxins 2013, 63, 39–46. [Google Scholar] [CrossRef] [Green Version]
- Partap, M.; Solanki, V.A.; Jagdish, P.; Nathawat, B.D.S. Evaluation of wheat cultivars for resistance against Alternaria alternata and associated fungi with black point. Plant Dis. Res. 2014, 29, 170–173. [Google Scholar]
- Somma, S.; Amatulli, M.T.; Masiello, M.; Moretti, A.; Logrieco, A.F. Alternaria species associated to wheat black point identified through a multilocus sequence approach. Int. J. Food Microbiol. 2019, 293, 34–43. [Google Scholar] [CrossRef]
- Woudenberg, J.H.C.; Groenewald, J.Z.; Binder, M.; Crous, P.W. Alternaria redefined. Stud Mycol. 2013, 75, 171–212. [Google Scholar] [CrossRef] [Green Version]
- Woudenberg, J.H.C.; Seidl, M.F.; Groenewald, E.; de Vries, M.; Stielow, B.; Thomma, B.J.; Crous, P.W. Alternaria section Alternaria: Species, formaespeciales or pathotypes. Stud Mycol. 2015, 82, 1–21. [Google Scholar] [CrossRef] [Green Version]
- Lawrence, D.P.; Gannibal, P.B.; Peever, T.L.; Dugan, F.M.; Pryor, B. Characterization of Alternaria isolates from the infectoria species-group and a new taxon from Arrhenatherum, Pseudoalternaria arrhenatheria sp. nov. Mycol. Prog. 2014, 13, 257–276. [Google Scholar] [CrossRef]
- Perellò, A.; Moreno, M.; Sisterna, M. Alternaria infectoria species-group associated with Black point of wheat in Argentina. Plant Pathol. 2008, 57, 379. [Google Scholar] [CrossRef] [Green Version]
- Poursafar, A.; Ghosta, Y.; Orina, A.S.; Gannibal, P.B.; Nikkhah, M.J.; Lawrence, D.P. Taxonomic study on Alternaria sections Infectoriae and Pseudoalternaria associated with black (sooty) head mold of wheat and barley in Iran. Mycol. Prog. 2018, 17, 343–356. [Google Scholar] [CrossRef]
- Garganese, F.; Schena, L.; Siciliano, I.; Prigigallo, M.I.; Spadaro, D.; De Grassi, A.; Ippolito, A.; Sanzani, S.M. Characterization of Citrus-Associated Alternaria Species in Mediterranean Areas. PLoS ONE 2016, 11, e0163255. [Google Scholar] [CrossRef] [Green Version]
- Simmons, E.G. Alternaria: An Identification Manual, 6th ed.; CBS Fungal Biodiversity Centre: Utrecht, The Netherlands, 2007. [Google Scholar]
- Simmons, E.G.; Roberts, R. Alternaria themes and variations. Mycotaxon 1993, 48, 109–140. [Google Scholar]
- Oviedo, M.S.; Sturm, M.E.; Reynoso, M.M.; Chulze, S.N.; Ramirez, M.L. Toxigenic profile and AFLP variability of Alternaria alternata and Alternaria infectoria occurring on wheat. Braz. J. Microbiol. 2013, 44, 447–455. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Polizzotto, R.; Andersen, B.; Martini, M.; Grisan, S.; Assante, G.; Musetti, R. A polyphasic approach for the characterization of endophytic Alternaria strains isolated from grapevines. J. Microbiol. Methods 2012, 88, 162–171. [Google Scholar] [CrossRef] [PubMed]
- Da Cruz Cabrala, L.; Terminiello, L.; Pinto, V.F.; Fog, K.; Patriarca, N.A. Natural occurrence of mycotoxins and toxigenic capacity of Alternaria strains from mouldy peppers. Int. J. Food Microbiol. 2016, 7, 155–160. [Google Scholar] [CrossRef] [PubMed]
- Yekeler, H.; Bitmis, K.; Ozcelik, N.; Doymaz, M.Z.; Calta, M. Analysis of toxic effects of Alternaria toxins on oesophagus of mice by light and electron microscopy. Toxicol. Pathol. 2001, 29, 492–497. [Google Scholar] [CrossRef] [Green Version]
- Siegel, D.; Feist, M.; Proske, M.; Koch, M.; Nehls, I. Degradation of the Alternaria mycotoxins alternariol, alternariol monomethyl ether, and altenuene upon bread baking. J. Agric. Food Chem. 2010, 58, 9622–9630. [Google Scholar] [CrossRef]
- Romero Bernal, A.R.; Reynoso, C.M.; García Londoño, V.A.; Broggi, L.E.; Resnik, S.L. Alternaria toxins in Argentinean wheat, bran, and flour. Food Addit Contam Part B Surveill 2019, 12, 24–30. [Google Scholar] [CrossRef]
- Azcarate, M.P.; Patriarca, A.; Terminiello, L.; Fernandez Pinto, V. Alternaria Toxins in Wheat during the 2004 to 2005 Argentinean Harvest. J. Food Prot. 2008, 71, 1262–1265. [Google Scholar] [CrossRef]
- Zhao, K.; Shao, B.; Yang, D.; Li, F.; Zhu, J. Natural Occurrence of Alternaria Toxins in Wheat-Based Products and Their Dietary Exposure in China. PLoS ONE 2015, 10, e0132019. [Google Scholar] [CrossRef] [Green Version]
- Xu, W.; Han, X.; Li, F.; Zhang, L. Natural Occurrence of Alternaria Toxins in the 2015 Wheat from Anhui Province, China. Toxins 2016, 8, 308. [Google Scholar] [CrossRef] [Green Version]
- Topi, D.; Tavcar-Kalcher, G.; Pavšič-Vrtač, K.; Babič, J.; Jakovac-Strajn, B. Alternaria mycotoxins in grains from Albania: Alternariol, alternariol monomethyl ether, tenuazonic acid and tentoxin. World Mycotoxin J. 2019, 12, 89–99. [Google Scholar] [CrossRef]
- Wenderoth, M.; Garganese, F.; Schmidt-Heydt, M.; Soukup, S.T.; Ippolito, A.; Sanzani, S.M.; Fischer, R. Alternariol as virulence and colonization factor of Alternaria alternata during plant infection. Mol. Microbiol. 2019, 112, 131–146. [Google Scholar] [CrossRef]
- Rozen, S.; Skaletsky, H. Primer3 on the WWW for general users and for biologist programmers. Methods Mol. Biol. 2000, 132, 365–386. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Avenot, H.F.; Michailides, T.J. Resistance to boscalid fungicide in Alternaria alternata isolates from pistachio in California. Plant Dis. 2007, 91, 1345–1350. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Greco, M.; Patriarca, A.; Terminiello, L.; Fernández Pinto, V.; Pose, G. Toxigenic Alternaria species from Argentinean blueberries. Int. J. Food Microbiol. 2012, 154, 187–191. [Google Scholar] [CrossRef]
- Andersen, B.; Hansen, M.E.; Smedsgaard, J. Automated and unbiased image analyses as tool in phenotypic classification of small-spored Alternaria spp. Phytopathology 2005, 95, 1021–1029. [Google Scholar] [CrossRef] [Green Version]
- Andersen, B.; Dongo, A.; Pryor, B.M. Secondary metabolite profiling of Alternaria dauci, A. porri, A. solani and A. tomatophila. Mycol. Res. 2008, 112, 241–250. [Google Scholar] [CrossRef]
- Brun, S.; Madrid, H.; Van Den Ende, B.G.; Andersen, B.; Marinach-Patrice, C.; Mazier, D. Multilocus phylogeny and MALDI-TOF analysis of the plant pathogenic species Alternaria dauci and relatives. Fungal Biol. 2013, 117, 32–40. [Google Scholar] [CrossRef]
- Chou, H.H.; Wu, W.S. Phylogenetic analysis of internal transcribed spacer regions of the genus Alternaria, and the significance of filament-beaked conidia. Mycol. Res. 2002, 106, 164–169. [Google Scholar] [CrossRef]
- Hong, S.G.; Cramer, R.A.; Lawrence, C.B.; Pryor, B.M. Alt a1 allergen homologs from Alternaria and related taxa: Analysis of phylogenetic content and secondary structure. Fungal Genet. Biol. 2005, 42, 119–129. [Google Scholar] [CrossRef]
- Inderbitzin, P.; Shoemaker, R.A.; O’Neill, N.R.; Turgeon, B.G.; Berbee, M.L. Systematics and mating systems of two fungal pathogens of opium poppy: The heterothallic Crivellia papaveracea with a Brachycladium penicillatum asexual state and a homotallic species with a Brachycladium papaveris asexual state. Can. J. Bot. 2006, 84, 1304–1326. [Google Scholar] [CrossRef]
- Pryor, B.M.; Creamer, R.; Shoemaker, R.A.; McLain-Romero, J.; Hambleton, S. Undifilum, a new genus for endophytic Embellisia oxytropis and parasitic Helminthosporium bornmuelleri on legumes. Botany 2009, 87, 178–194. [Google Scholar] [CrossRef] [Green Version]
- Runa, M.; Park, M.; Pryor, B. Ulocladium systematics revisited: Phylogeny and taxonomic status. Mycol. Prog. 2009, 8, 35–47. [Google Scholar] [CrossRef]
- Somma, S.; Pose, G.; Pardo, A.; Mulè, G.; Pinto, V.F.; Moretti, A.; Logrieco, A.F. AFLP variability, toxin production and pathogenicity of Alternaria species from Argentinian tomato fruits and puree. Int. J. Food Microbiol. 2011, 145, 414–419. [Google Scholar] [CrossRef] [PubMed]
- Pryor, B.M.; Michailides, T. Morphological, Pathogenic, and Molecular Characterization of Alternaria Isolates Associated with Alternaria Late Blight of Pistachio. Phytopathology 2002, 92, 406–416. [Google Scholar] [CrossRef] [Green Version]
- Patriarca, A.; Azcarate, M.P.; Terminiello, L.; Fernández Pinto, V. Mycotoxin production by Alternaria strains isolated from Argentinean wheat. Int. J. Food Microbiol. 2007, 119, 219–222. [Google Scholar] [CrossRef]
- Gargouri-Kammoun, L.; Bensassi, F.; Mnari-Hattab, M.; Rhouma, A.; Bacha, H.; Hajlaoui, M.R. Identification of Alternaria Species Recovered from Stored Durum Wheat Kernels in Tunisia. Tunis. J. Plant Prot. 2014, 9, 119–129. [Google Scholar]
- Kahl, S.M.; Ulrich, A.; Kirichenko, A.A.; Muller, M.E.H. Phenotypic and phylogenetic segregation of Alternaria infectoria from small-spored Alternaria species isolated from wheat in Germany and Russia. J. Appl. Microbiol. 2015, 119, 1637–1650. [Google Scholar] [CrossRef]
- Kosiak, B.; Torp, M.; Skjerve, E.; Andersen, B. Alternaria and Fusarium in Norwegian grains of reduced quality—A matched pair sample study. Int. J. Food Microbiol. 2004, 93, 51–62. [Google Scholar] [CrossRef]
- Müller, M.E.H.; Korn, U. Alternaria mycotoxins in wheat—A 10 years survey in the Northeast of Germany. Food Control 2013, 34, 191–197. [Google Scholar] [CrossRef]
- Liu, G.T.; Qian, Y.Z.; Zhang, P.; Dong, Z.M.; Shi, Z.Y.; Zhen, Y.Z.; Miao, J.; Xu, Y.M. Relationships between Alternaria alternata and oesophageal cancer. IARC Sci. Publ. 1991, 105, 258–262. [Google Scholar]
- Berbee, M.L.; Pirseyedi, M.; Hubbard, S. Cochliobolus phylogenetics and the origin of known, highly virulent pathogens, inferred from ITS and glyceraldehyde-3-phosphate dehydrogenase gene sequences. Mycologia 1999, 91, 964–977. [Google Scholar] [CrossRef]
- Tamura, K.; Peterson, D.; Peterson, N.; Stecher, G.; Nei, M.; Kumar, S. MEGA5: Molecular Evolutionary Genetics Analysis Using Maximum Likelihood, Evolutionary Distance, and Maximum Parsimony Methods. Mo. Biol. Evol. 2011, 28, 2731–2739. [Google Scholar] [CrossRef] [Green Version]
- Li, F.Q.; Toyazaki, N.; Yoshizawa, T. Production of Alternaria mycotoxins by Alternaria alternata isolated from weather-damaged wheat. J. Food Prot. 2001, 64, 567–571. [Google Scholar] [CrossRef]
- Rubert, J.; Dzuman, Z.; Vaclavikova, M.; Zachariasova, M.; Soler, C.; Hajslova, J. Analysis of mycotoxins in barley using ultra high liquid chromatography high resolution mass spectrometry: Comparison of efficiency and efficacy of different extraction procedures. Talanta 2012, 99, 712–719. [Google Scholar] [CrossRef]
- Myresiotis, C.K.; Testempasis, S.; Vryzas, Z.; Karaoglanidis, G.S.; Papadopoulou-Mourkidou, E. Determination of mycotoxins in pomegranate fruits and juices using a QuEChERS-based method. Food Chem. 2015, 182, 81–88. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Sample Code | Genotype | Treated with Fungicides * | Untreated | ||||
|---|---|---|---|---|---|---|---|
| Blackpoint Symptoms (0–9 Scale) | Alternaria Incidence of Infected Kernels (%) | Alternariol (µg Kg−1) | Blackpoint Symptoms (0–9 Scale) | Alternaria Incidence of Infected Kernels (%) | Alternariol (µg Kg−1) | ||
| G1 | D04802 | 4.0 | 5 | 34 | 5.0 | 0 | 87 |
| G2 | D05422 | 2.5 | 7 | 72 | 4.5 | 12 | 148 |
| G3 | 508Gd05/101 | 1.0 | 6 | 24 | 4.0 | 12 | 101 |
| G4 | 508gd05/209 | 0.5 | 8 | 31 | 1.5 | 11 | Nd |
| G5 | 508gd07/10T | 0.5 | 10 | Nd | 0.5 | 8 | 109 |
| G6 | D07422 | 1.0 | 2 | 47 | 3.0 | 4 | 88 |
| G7 | 909gd08/20B | 0.5 | 6 | 44 | 2.0 | 11 | 109 |
| G8 | 909gd08/77 | 2.5 | 5 | 79 | 6.0 | 13 | Nd |
| G9 | 909gd08/94A | 6.0 | 5 | Nd | 8.0 | 16 | 124 |
| G10 | 905gd09/37 | 2.0 | 15 | 55 | 4.0 | 9 | 115 |
| G11 | 505gd09/131A | 1.5 | 12 | 51 | 3.5 | 8 | - |
| G12 | 505gd09/131B | 3.0 | 26 | 31 | 4.5 | 5 | 84 |
| G13 | 906gd10/4T | 3.0 | 16 | 37 | 4.0 | 7 | 85 |
| G14 | 906gd104P | 3.5 | 8 | 65 | 4.0 | 14 | 112 |
| G15 | 906gd10/21A | 2.0 | 4 | 44 | 4.0 | 13 | 77 |
| G16 | 906gd10/21B | 4.0 | 10 | 36 | 6.0 | 8 | 97 |
| G17 | 906gd10/40T | 0.0 | 5 | 27 | 1.5 | 8 | 88 |
| G18 | 906gd10/40P | 0.5 | 11 | 85 | 1.0 | 6 | 98 |
| G19 | 906gd10/72T | 1.5 | 0 | 39 | 3.5 | 9 | 106 |
| G20 | 906gd10/72P | 1.5 | 8 | 40 | 3.0 | 3 | 89 |
| G21 | 906gd010/88A | 1.0 | 0 | 43 | 1.5 | 5 | 77 |
| G22 | 906gd10/88B | 0.5 | 5 | 26 | 3.0 | 8 | 40 |
| G23 | 906gd10/118T | 0.5 | 3 | Nd | 1.5 | 12 | 93 |
| G24 | 906gd10/118P | 1.5 | 1 | 28 | 4.0 | 16 | 93 |
| Min value | 0 | 0 | 0 | 0.5 | 0 | 0 | |
| Max value | 6 | 26 | 85 | 8 | 16 | 148 | |
| Average | 1.9 | 7.4 | 39.1 | 3.5 | 9.1 | 84.2 | |
| Standard Deviation | 1.5 | 5.8 | 22.3 | 1.8 | 4.1 | 34.3 | |
| G25 | CLAUDIO | 1.0 | 6 | 42 | 6.0 | 9 | 97 |
| G26 | IRIDE | 3.0 | 9 | 91 | 6.0 | 6 | Nd |
| G27 | LIBERDUR | 1.0 | 20 | 44 | 2.0 | 11 | 77 |
| G28 | MARCO AURELIO | 3.0 | 2 | 66 | 5.0 | 21 | 110 |
| G29 | MIRADOUX | 1.0 | 7 | 87 | 2.0 | 10 | 87 |
| G30 | SARAGOLLA | 2.5 | 9 | 92 | 4.0 | 17 | 110 |
| Sample Code | Genotype | Treated with Fungicides * | Untreated | ||||
|---|---|---|---|---|---|---|---|
| Blackpoint Symptoms (0–9 Scale) | Alternaria Incidence of Infected Kernels (%) | Alternariol (µg Kg−1) | Blackpoint Symptoms (0–9 Scale) | Alternaria Incidence of Infected Kernels (%) | Alternariol (µg Kg−1) | ||
| G1 | D04802 | 3.0 | 8 | 118 | 5.0 | 14 | 118 |
| G2 | D05422 | 6.0 | 11 | 141 | 6.0 | 15 | Nd |
| G3 | 508Gd05/101 | 1.5 | 13 | 124 | 3.0 | 13 | 133 |
| G4 | 508gd05/209 | 0.5 | 6 | 97 | 1.0 | 11 | Nd |
| G5 | 508gd07/10T | 0.5 | 4 | 262 | 1.0 | 12 | 105 |
| G6 | D07422 | 0.5 | 7 | 161 | 1.5 | 9 | 230 |
| G7 | 909gd08/20B | 1.0 | 6 | 76 | 2.0 | 12 | Nd |
| G8 | 909gd08/77 | 5.0 | 10 | 147 | 6.0 | 14 | 84 |
| G9 | 909gd08/94A | 2.0 | 6 | 64 | 3.5 | 13 | Nd |
| G10 | 905gd09/37 | 0.5 | 2 | Nd | 1.0 | 10 | 97 |
| G11 | 505gd09/131A | 1.0 | 6 | 100 | 3.0 | 12 | Nd |
| G12 | 505gd09/131B | 1.0 | 4 | 100 | 2.0 | 12 | Nd |
| G13 | 906gd10/4T | 2.0 | 11 | 111 | 3.5 | 5 | 120 |
| G14 | 906gd104P | 1.0 | 12 | 127 | 4.0 | 13 | Nd |
| G15 | 906gd10/21A | 1.5 | 17 | 111 | 4.0 | 15 | Nd |
| G16 | 906gd10/21B | 5.0 | 6 | 107 | 3.0 | 14 | 95 |
| G17 | 906gd10/40T | 1.0 | 8 | 180 | 2.0 | 12 | Nd |
| G18 | 906gd10/40P | 0.5 | 5 | Nd | 2.0 | 12 | Nd |
| G19 | 906gd10/72T | 1.0 | 9 | 104 | 5.0 | 14 | 125 |
| G20 | 906gd10/72P | 0.5 | 7 | Nd | 1.5 | 12 | 68 |
| G21 | 906gd010/88A | 2.0 | 4 | 91 | 4.0 | 15 | 94 |
| G22 | 906gd10/88B | 2.0 | 10 | 111 | 3.0 | 3 | 171 |
| G23 | 906gd10/118T | 0.5 | 3 | 139 | 1.0 | 5 | 78 |
| G24 | 906gd10/118P | 0.5 | 7 | 141 | 2.0 | 11 | 71 |
| Min value | 0.5 | 2 | 0 | 1 | 3 | 0 | |
| Max value | 6 | 17 | 262 | 6 | 15 | 230 | |
| Average | 1.7 | 7.6 | 108.8 | 2.9 | 11.6 | 66.2 | |
| Standard Deviation | 1.6 | 3.5 | 57.3 | 1.5 | 3.2 | 65.8 | |
| G25 | CLAUDIO | 0.5 | 9 | 76 | 3.0 | 3 | Nd |
| G26 | IRIDE | 0.5 | 5 | 71 | 2.0 | 8 | 155 |
| G27 | LIBERDUR | 1.0 | 9 | 157 | 1.5 | 17 | 83 |
| G28 | MARCO AURELIO | 2.0 | 7 | 109 | 4.0 | 5 | 97 |
| G29 | MIRADOUX | 0.5 | 16 | Nd | 1.0 | 20 | 160 |
| G30 | SARAGOLLA | 1.0 | 10 | Nd | 2.0 | 13 | 169 |
| Strain (ITEM) | mg kg−1 | Strain (ITEM) | mg kg−1 | Strain (ITEM) | mg kg−1 | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| AOH | AME | ALT | TA | AOH | AME | ALT | TA | AOH | AME | ALT | TA | |||
| 17235 A2 | 67 | 182 | - | - | 17272 A2 | 38 | 310 | Nd | 1030 | 17348 A1 | 127 | 501 | Nd | 1775 |
| 17237 A2 | 815 | 2618 | Nd | 603 | 17274 A2 | 46 | 185 | Nd | 965 | 17271 A1 | 448 | 2703 | - | - |
| 17242 A2 | 1070 | 2557 | Nd | 1022 | 17276 A2 | 26 | 353 | Nd | 977 | 17277 A1 | 1085 | 2113 | Nd | 311 |
| 17243 A2 | 251 | 2167 | Nd | 306 | 17350 A2 | 65 | 371 | Nd | 476 | 17352 A1 | 1002 | 1306 | Nd | 157 |
| 17244 A2 | 100 | 1000 | 25 | Nd | 17351 A2 | 69 | 798 | Nd | 934 | 17357 A1 | 752 | 3849 | Nd | 2725 |
| 17247 A2 | 1682 | 2349 | - | - | 17354 A2 | 85 | 9 | Nd | 1596 | 17360 A1 | 1 | 5 | - | - |
| 17258 A2 | 46 | 65 | - | 575 | 17355 A2 | 372 | 2239 | - | - | 17363 A1 | 145 | 14 | Nd | Nd |
| 17260 A2 | 1169 | 7965 | Nd | 755 | 17358 A2 | 138 | 369 | Nd | 572 | 17366 A1 | 22 | 30 | Nd | Nd |
| 17269 A2 | 378 | 129 | Nd | Nd | 17364 A2 | 872 | 2396 | - | - | 17371 A1 | 1512 | 2035 | Nd | Nd |
| 17270 A2 | 872 | 50 | Nd | Nd | 17374 A2 | 992 | 3357 | Nd | 642 | Average A1 | 884 | 1449 | - | 730 |
| 17273 A2 | 1822 | 5603 | Nd | 1654 | Average A2 | 774 | 2099 | - | 723 | 17333 A3 | 1633 | 1934 | - | - |
| 17275 A2 | 7372 | 13,156 | Nd | 1009 | 17228 A1 | 651 | 548 | Nd | Nd | 17334 A3 | 60 | 91 | - | - |
| 17346 A2 | 3152 | 7143 | Nd | 938 | 17229 A1 | 23 | 33 | - | - | 17336 A3 | 62 | 620 | Nd | 2143 |
| 17359 A2 | 11 | 5 | - | - | 17230 A1 | 4176 | 4213 | 2.5 | 822 | 17337 A3 | 415 | 2866 | - | - |
| 17365 A2 | 3581 | 14,341 | Nd | 1656 | 17231 A1 | 1283 | 2567 | Nd | Nd | 17338 A3 | 1781 | 2763 | Nd | 1084 |
| 17367 A2 | 62 | 39 | Nd | Nd | 17233 A1 | 22 | 163 | Nd | 3043 | 17345 A3 | 28 | 385 | Nd | 2485 |
| 17368 A2 | 40 | 374 | Nd | 377 | 17236 A1 | 1283 | 1612 | - | - | 17347 A3 | 213 | 288 | Nd | 1767 |
| 17369 A2 | 57 | 230 | - | - | 17238 A1 | 925 | 1833 | - | - | 17349 A3 | 73 | 1039 | Nd | 3684 |
| 17373 A2 | 245 | 823 | Nd | 985 | 17239 A1 | 89 | 10 | Nd | Nd | 17353 A3 | 451 | 287 | Nd | 773 |
| 17232 A2 | 1962 | 4485 | Nd | Nd | 17241 A1 | 154 | 29 | Nd | Nd | 17361 A3 | 23 | 16 | Nd | 736 |
| 17234 A2 | 1296 | 2775 | Nd | Nd | 17245 A1 | 8064 | 8597 | - | - | 17362 A3 | 636 | 2261 | Nd | 2111 |
| 17246 A2 | 842 | 1257 | Nd | 914 | 17248 A1 | 1309 | 1067 | Nd | 1417 | 17370 A3 | 2434 | 8730 | Nd | 816 |
| 17251 A2 | 34 | 30 | Nd | 710 | 17249 A1 | 948 | 2606 | Nd | 920 | Average A3 | 651 | 1773 | 1733 | |
| 17253 A2 | 32 | 127 | Nd | 2059 | 17250 A1 | 531 | 2437 | Nd | 750 | 17335 B | 68 | 4 | Nd | Nd |
| 17255 A2 | 162 | 605 | Nd | 621 | 17252 A1 | 611 | 787 | - | - | 17339 B | 25 | 64 | Nd | Nd |
| 17261 A2 | 221 | 2174 | - | - | 17254 A1 | 50 | 53 | Nd | Nd | 17340 B | 12 | 19 | Nd | Nd |
| 17262 A2 | 25 | 6 | Nd | 901 | 17256 A1 | 505 | 1045 | Nd | 953 | 17341 B | 8 | 11 | - | - |
| 17263 A2 | Nd | 49 | Nd | 183 | 17257 A1 | Nd | Nd | Nd | 605 | 17342 B | 5 | 4 | Nd | Nd |
| 17240 A2 | 316 | 166 | Nd | Nd | 17259 A1 | 240 | 87 | Nd | 680 | 17344 B | Nd | Nd | Nd | Nd |
| 17265 A2 | 44 | 357 | Nd | 2388 | 17264 A1 | 112 | 152 | Nd | 1911 | 17356 B | 223 | 151 | Nd | Nd |
| 17266 A2 | 420 | 2178 | Nd | 463 | 17268 A1 | 300 | 635 | Nd | Nd | 17372 B | 0 | 1 | Nd | 23 |
| 17267 A2 | 880 | 2765 | Nd | Nd | 17343 A1 | 143 | 2442 | - | - | Average B | 43 | 32 | - | - |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Masiello, M.; Somma, S.; Susca, A.; Ghionna, V.; Logrieco, A.F.; Franzoni, M.; Ravaglia, S.; Meca, G.; Moretti, A. Molecular Identification and Mycotoxin Production by Alternaria Species Occurring on Durum Wheat, Showing Black Point Symptoms. Toxins 2020, 12, 275. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins12040275
Masiello M, Somma S, Susca A, Ghionna V, Logrieco AF, Franzoni M, Ravaglia S, Meca G, Moretti A. Molecular Identification and Mycotoxin Production by Alternaria Species Occurring on Durum Wheat, Showing Black Point Symptoms. Toxins. 2020; 12(4):275. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins12040275
Chicago/Turabian StyleMasiello, Mario, Stefania Somma, Antonia Susca, Veronica Ghionna, Antonio Francesco Logrieco, Matteo Franzoni, Stefano Ravaglia, Giuseppe Meca, and Antonio Moretti. 2020. "Molecular Identification and Mycotoxin Production by Alternaria Species Occurring on Durum Wheat, Showing Black Point Symptoms" Toxins 12, no. 4: 275. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins12040275