The Occurrence of Potential Harmful Cyanobacteria and Cyanotoxins in the Obrzyca River (Poland), a Source of Drinking Water


Abstract
:1. Introduction
2. Results
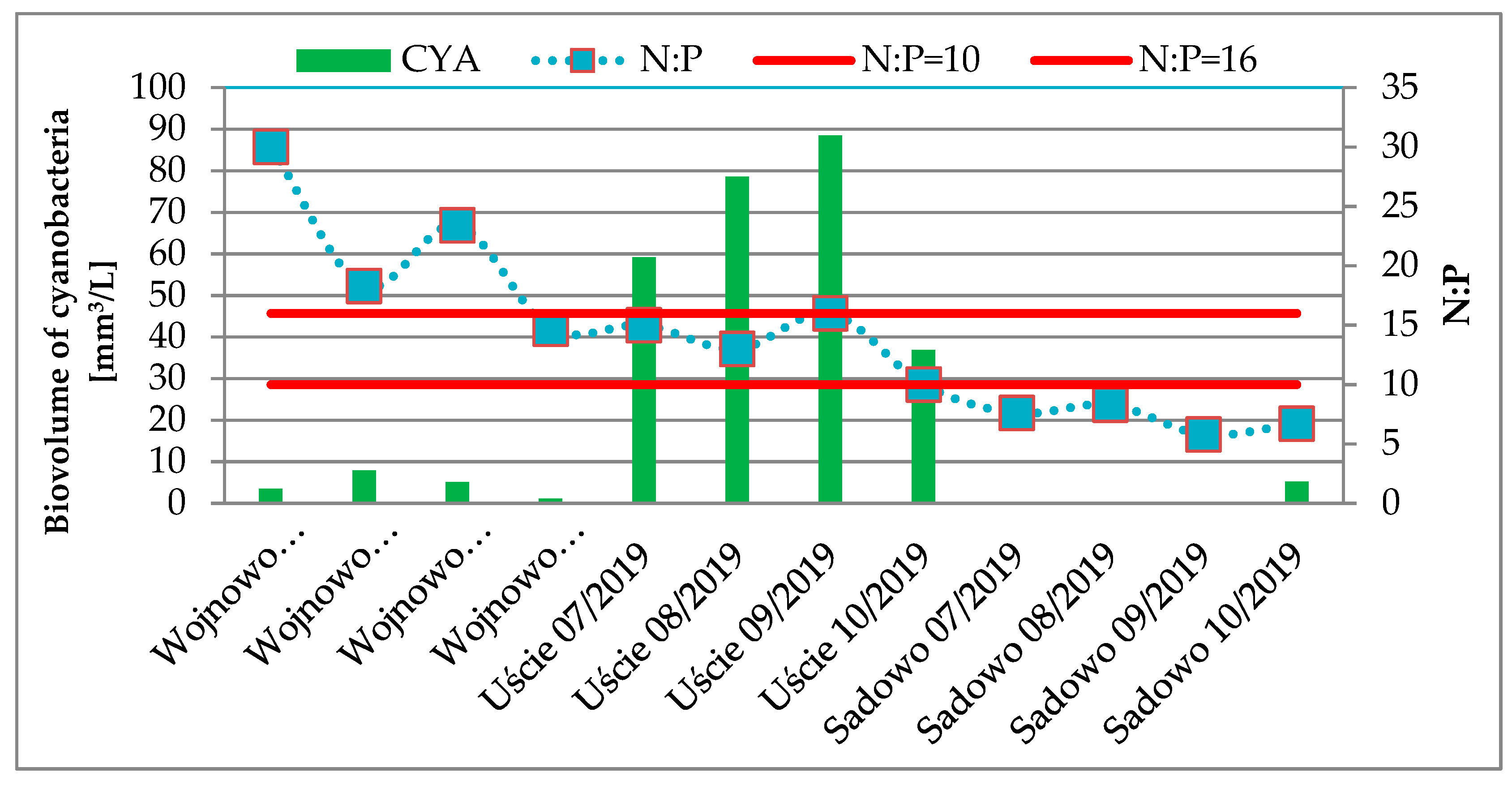
2.1. Cyanobacterial Blooms
2.2. Cyanotoxins
2.3. Bioassay Test
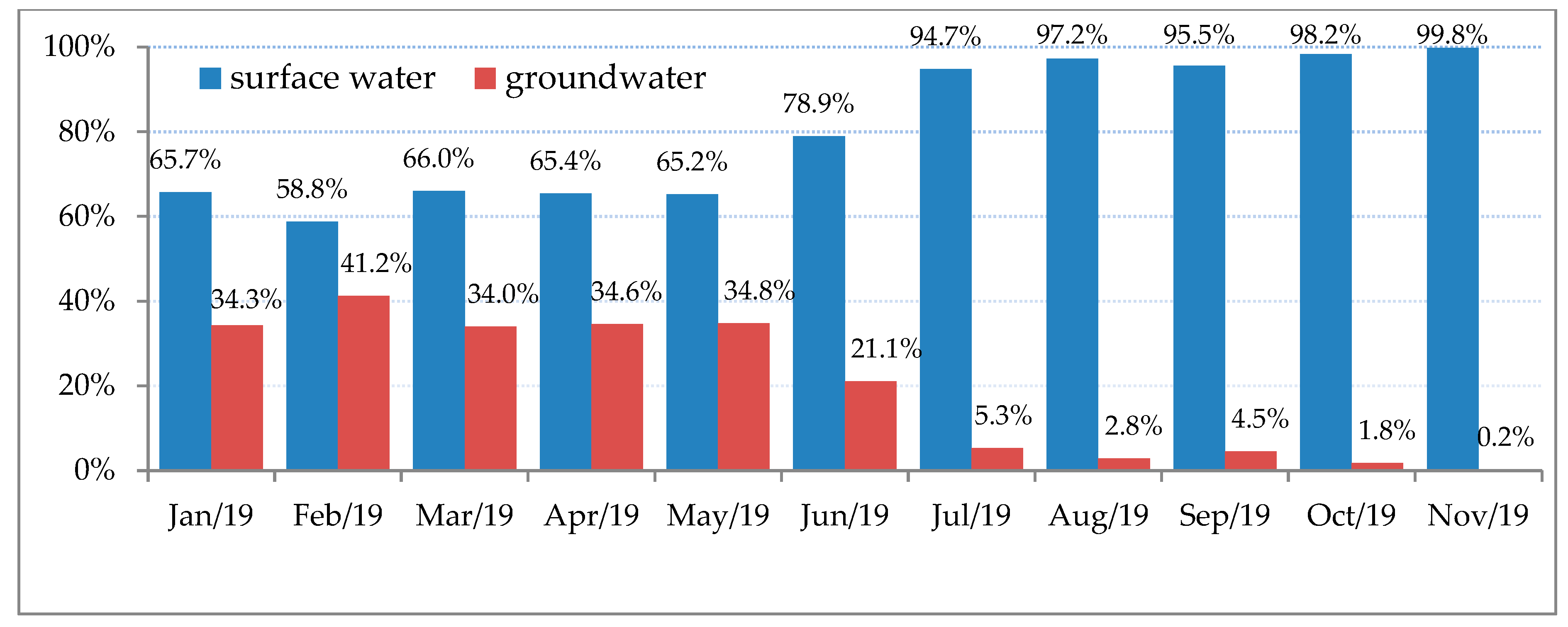
2.4. Physical and Chemical Water Quality Indicators
3. Discussion
4. Conclusions
- The occurrence of cyanobacterial blooms in the Obrzyca River is punctual.
- The highest intracellular microcystin concentration (3.97 μg/L) was determined in the Obrzyca River at the Uście point, where cyanobacterial bloom was noticed.
- Cyanobacterial extracts collected from August to October at the Uście point were toxic for D. tigrina where intracellular microcystins were present. The most toxic sample for planarians was taken in September (LC 50 was 21% extract concentration). Bioassays with D. tigrina are sensitive and applicable for the assessment of the toxicity of cyanobacterial blooms.
- A correlation between cyanobacterial abundance and pH, turbidity, total nitrogen, and total suspension was found. Water quality indicators, i.e., pH, total phosphorus, total nitrogen, turbidity, and total suspension were strongly correlated with intracellular microcystins MC-RR and MC-LR. The analysis showed that cyanobacterial blooms took place when the N:P ratio was in the range of 10–16.
- The conducted studies (biological, chemical, and physical) complement each other well and thus are a great tool to analyze the risks of harmful cyanobacteria in drinking water, especially in DWTPs.
5. Materials and Methods
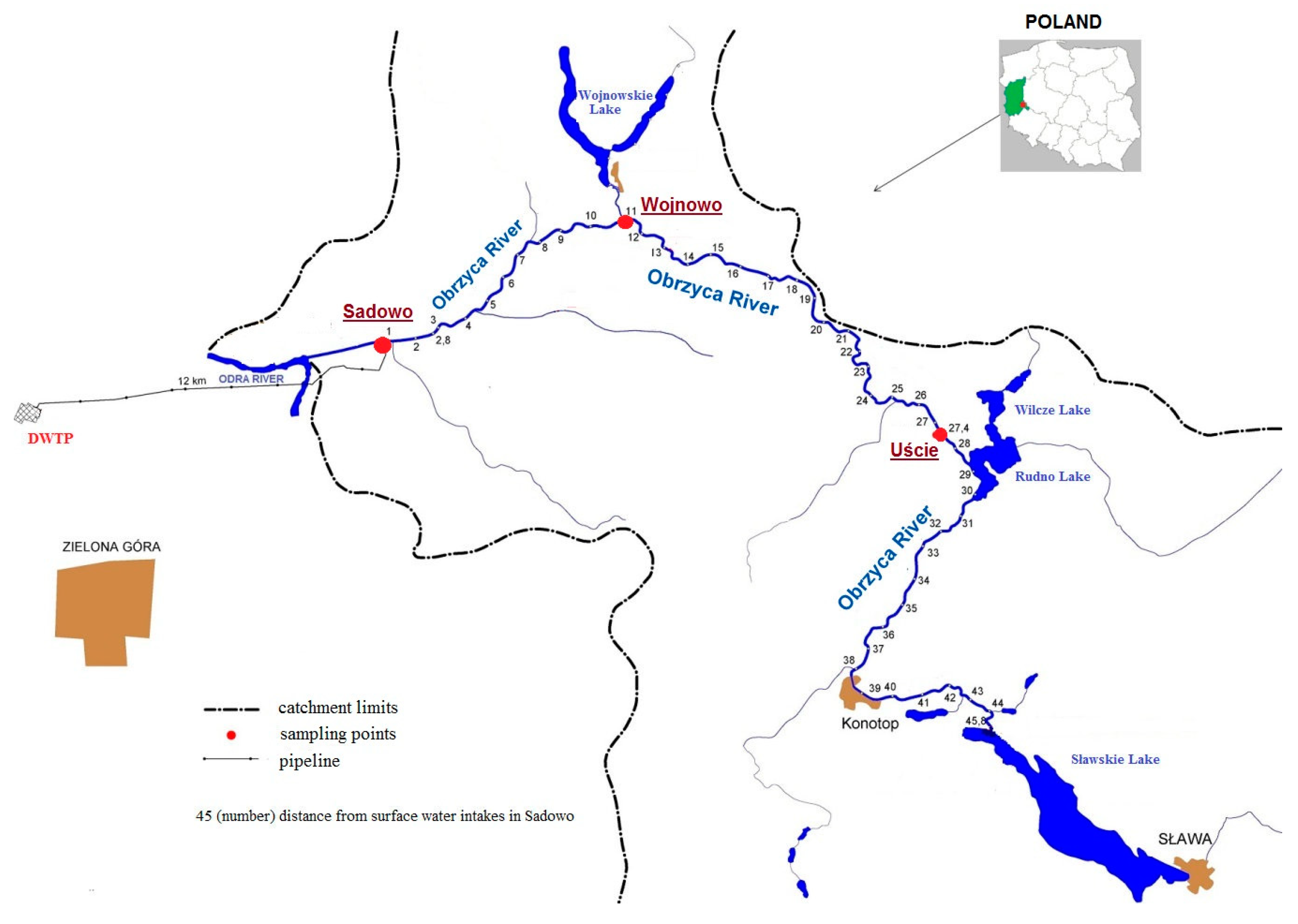
5.1. Study Area
5.2. Sampling
5.3. Physical and Chemical Water Quality Indicator Analysis
5.4. Microscopic Analysis
5.5. Chlorophyll a
5.6. Cyanotoxin Analysis
5.7. Bioassays Tests
5.8. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Mantzouki, E.; Lürling, M.; Fastner, J.; de Senerpont Domis, L.; Wilk-Wożniak, E.; Korelviené, J.; Seelen, L.; Teurlincx, S.; Verstijnen, Y.; Krztoń, W.; et al. Temperature effects explain continental scale distribution of cyanobacterial toxins. Toxins 2018, 10, 156. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Szlag, D.C.; Sinclair, J.L.; Southwell, B.; Westrick, J.A. Cyanobacteria and cyaonotoxins occurrence and removal from five high risk conventional drinking water plants. Toxins 2015, 7, 2198–2220. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carmichael, W.W.; Azevedo, S.M.; An, J.S.; Molica, R.J.; Jochimsen, E.M.; Lau, S.; Rinehart, K.L.; Shaw, G.R.; Eaglesham, G.K. Human fatalities from cyanobacteria: Chemical and biological evidence for cyanotoxins. Environ. Health Perspect 2001, 109, 663–668. [Google Scholar] [CrossRef] [PubMed]
- Ibrahim, W.M.; Salim, E.H.; Azab, Y.A.; Ismail, A.-H.M. Monitoring removal of cyanobacterial toxins from drinking water by algal-activated carbon. Toxicol. Ind. Health 2015, 32, 1752–1762. [Google Scholar] [CrossRef] [PubMed]
- Trung, B.; Dao, T.-S.; Faassen, E.; Lürling, M. Cyanobacterial blooms and microcystins in Southern Vietnam. Toxins 2018, 10, 471. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoeger, S.J.; Shaw, G.; Hitzfeld, B.C.; Dietrich, D.R. Occurrence and elimination of cyanobacterial toxins in two Australian drinking water treatment plants. Toxicon 2004, 43, 639–649. [Google Scholar] [CrossRef] [Green Version]
- Teneva, I.; Mladenov, R.; Belkinova, D.; Dimitrova-Dyulgerova, I.; Dzhambazov, B. Phytoplankton community of the drinking water supply reservoir Borovitsa (South Bulgaria) with emphasis on cyatoxins and water quality. Cent. Eur. J. Biol. 2010, 5, 231–239. [Google Scholar]
- Pitois, F.; Fastner, J.; Pagotto, C.; Dechesne, M. Multi-toxin occurences in ten French water resource reservoirs. Toxins 2018, 10, 283. [Google Scholar] [CrossRef] [Green Version]
- Piontek, M.; Czyżewska, W.; Mankiewicz-Boczek, J. The occurrence of cyanobacteria blooms in the Obrzyca River catchment area (Poland), a source of drinking water. Pol. J. Environ. Stud. 2017, 26, 1191–1201. [Google Scholar] [CrossRef]
- Xe, X.; Liu, Y.-L.; Conklin, A.; Westrick, J.; Weavers, L.K.; Dionysiou, D.D.; Lenhart, J.J.; Mouser, J.; Szlag, D.; Walker, H.W. Toxic cyanobacteria and drinking water: Impacts, detection and treatment. Harmful Algae 2016, 4, 174–193. [Google Scholar]
- Huang, I.-S.; Zimba, P.V. Cyanobacterial active metabolites. A review of their chemistry and biology. Harmful Algae 2019, 86, 139–209. [Google Scholar] [PubMed]
- Jochimsen, E.M.; Carmichael, W.W.; An, J.S.; Cardo, D.M.; Cookson, S.T.; Holmes, C.E.; Antunes, M.B.; de Melo Filho, D.A.; Lyra, T.M.; Barreto, V.S.; et al. Liver failure and death after exposure to microcystins at a hemodlialysis center in Brazil. N. Engl. J. Med 1998, 338, 873–878. [Google Scholar] [CrossRef] [PubMed]
- Carmichael, W.W.; Boyer, G. Health impacts from cyanobacterial harmful algae blooms: Implications, for the North American Great Lakes. Harmful Algae 2016, 54, 194–212. [Google Scholar] [CrossRef] [PubMed]
- Hilborn, E.D.; Beasley, V.R. One health and cyanobacteria in freshwater systems: Animal illness and death are sentinel events for human health risks. Toxins 2015, 7, 1374–1395. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lévesque, B.; Gervais, M.-C.; Chevalier, P.; Gauvin, D.; Anassour-Laouan-Sidi, E.; Gingras, S.; Fortin, N.; Brissson, G.; Greer, C.; Bird, D. Prospective study of acute health effects in relation to exposure to cyanobacteria. Sci. Total Environ. 2014, 466, 397–403. [Google Scholar] [CrossRef]
- Svirčev, Z.; Lalič, D.; Savič, G.B.; Tokodi, N.; Backovič, D.D.; Chen, L.; Meriluoto, J.; Codd, G.A. Global geographical and historical overview of cyanotoxin distribution and cyanobacterial poisonings. Arch. Toxicol 2019, 93, 2429–2481. [Google Scholar] [CrossRef]
- Piontek, M.; Czyżewska, W. Influence of cyanobacterial bloom on freshwater biocoenosis. Use of bioassays for cyanobacterial microcystins toxicity assessment. Civ. Environ. Eng. Rep. 2017, 24, 47–67. [Google Scholar] [CrossRef]
- Rzymski, P.; Poniedziałek, B.; Karczewski, J. Gastroenteritis and liver carcinogenesis induced by cyanobacterial toxins. Gastroenterol. Pol. 2011, 18, 159–162. [Google Scholar]
- Žegura, B.; Štraser, A.; Filipič, M. Genotoxicity and potential carcinogenicity of cyanobacterial toxins—A review. Mutat. Res. 2011, 727, 16–41. [Google Scholar] [CrossRef]
- Hu, C.; Rzymski, P. Programmed cell death—Like and accompanying release of microcystin in freshwater bloom-forming cyanobacterium Microcystis: From identification to ecological relevance. Toxins 2019, 11, 706. [Google Scholar] [CrossRef] [Green Version]
- Bartram, J.; Burch, M.; Falconer, I.R.; Jones, G.; Kuiper-Goodman, T. Situation assessment, planning and management. In Toxic Cyanobacteria in Water. A Guide to the Their Public Health Consequences, Monitoring and Management; Chorus, I., Bartram, J., Eds.; Taylor & Francis: London, UK; New York, NY, USA, 1999; pp. 179–209. [Google Scholar]
- Burch, M.D. Effective doses, guidelines & regulations. In Cyanobacterial Harmful Algal Blooms: State of the Science and Research Needs; Hudnell, K.H., Ed.; Springer Sience & Business Media: Berlin/Heidelberg, Germany, 2008; Volume 619, pp. 831–853. [Google Scholar]
- Santé, O.M.D.L. Guidelines for Drinking Water Quality, 4th ed.; World Health Organization: Geneva, Switzerland, 2011. [Google Scholar]
- ISAP – Internetowy System Aktów Prawnych. Available online: http://prawo.sejm.gov.pl/isap.nsf/DocDetails.xsp?id=WDU20070610417 (accessed on 29 March 2007).
- Fristachi, A.; Sinclair, J.L. Occurrence of cyanobacterial harmful algal blooms workgroup report. In Cyanobacterial Harmful Algal Blooms State of the Science and Research Needs, 1st ed.; Hudnell, H.K., Ed.; Springer: New York, NY, USA, 2008; pp. 45–103. [Google Scholar]
- Mohamed, Z.A. Breakthrough of Oscillatoria limnetica and microcystin toxins into drinking water treatment plants—Examples from the Nile River. Water SA 2016, 42, 161–165. [Google Scholar] [CrossRef] [Green Version]
- Grabowska, M.; Mazur-Marzec, H. The effect of cyanobacterial bloom in the Siemianówka Dam Reservoir on the phytoplankton structure in the Narew River. Oceanol. Hydrobiol. Stud. 2011, 40, 19–26. [Google Scholar] [CrossRef]
- Piontek, M. Use of a planarian Dugesia tigrina Girard in the studies of acute toxicity of organic substances. Pol. Arch. Hydrobiol. 1999, 46, 331–338. [Google Scholar]
- Czyżewska, W.; Piontek, M. The efficiency of microstrainers filtration in the process of removing phytoplankton with special consideration of cyanobacteria. Toxins 2019, 11, 285. [Google Scholar] [CrossRef] [Green Version]
- Polish Provincial Inspectorate of Environmental Protection. The state of purity of Rudno Lake based on studies conducted in 1991-2015 (Stan czystości jeziora Rudno na podstawie badań WIOŚ w Zielonej Górze zrealizowanych w latach 1991–2015 (in Polish)). Available online: http://www.zgora.pios.gov.pl/wp-content/uploads/2017/09/2017-09-15-RUDNO-opracowanie-PS.pdf (accessed on 23 September 2017).
- Kobos, J.; Błaszczyk, A.; Hohlfeld, N.; Toruńska-Sitarz, A.; Krakowiak, A.; Hebel, A.; Sutryk, K.; Grabowska, M.; Toporowska, M.; Kokociński, M.; et al. Cyanobacteria and cyanotoxins in Polish freshwater bodies. Oceanol. Hydrobiol. Stud. 2013, 42, 358–378. [Google Scholar] [CrossRef] [Green Version]
- Kurmayer, R.; Gumpenberger, M. Diversity of microcystin genotypes among populations of the filamentous cyanobacteria Planktothrix rubescens and Planktothrix agardhii. Mol. Ecol. 2006, 15, 3849–3861. [Google Scholar] [CrossRef] [Green Version]
- Briand, E.; Gugger, M.; Francois, J.-C.; Bernard, C.; Humbert, J.-C.; Quiblier, C. Temporal Variations in the Dynamics of Potentially Microcystin-Producing Strains in a Bloom-Forming Planktothrix agardhii (Cyanobacterium) Population. Appl. Environ. Microbiol. 2008, 74, 3839–3848. [Google Scholar] [CrossRef] [Green Version]
- Kufel, L.; Prejs, A.; Rybak, J.I. Factors controlling the dominance of Planktothrix agardhii and Limnothrix redekei in eutrophic shallow lakes. Hydrobiologia 1997, 342/343, 107–115. [Google Scholar] [CrossRef]
- Churro, C.; Azevedo, J.; Vasconcelos, V.; Silva, A. Detection of a Planktothrix agardhii Bloom in Portuguese Marine Coastal Waters. Toxins 2017, 9, 391. [Google Scholar] [CrossRef] [Green Version]
- Komárek, J.; Zapomélowa, E. Planktic morphospecies of the cyanobacterial genus Anabaena = subg. Dolichospermum—1. part: Coiled types. Fottea 2007, 7, 1–31. [Google Scholar]
- Du, X.; Liu, H.; Yuan, L.; Wang, Y.; Ma, Y.; Wang, R.; Chen, X.; Losiewicz, M.D.; Guo, H.; Zhang, H. The diversity of cyanobacterial toxins on structural characterization, distribution and identification: A systematic review. Toxins 2019, 11, 530. [Google Scholar] [CrossRef] [Green Version]
- Mankiewicz-Boczek, J.; Gągała, I.; Kokociński, M.; Jurczak, T.; Stefaniak, K. Perennial toxigenic Planktothrix agardhii bloom in selected lakes of Western Poland. Environ. Toxicol 2011, 26, 10–20. [Google Scholar] [CrossRef]
- Grabowska, M.; Mazur-Marzec, H. Vertical distribution of cyanobacteria biomass and cyanotoxin production in the polymictic Siemianówka Dam Reservoir (Easter Poland). Arch. Pol. Fish 2014, 22, 41–51. [Google Scholar] [CrossRef]
- Krüger, T.; Wiegand, C.; Kun, L.; Luckas, B.; Pflugmacher, S. More and more toxins around – analysis of cyanobacterial strains isolated from Lake Chao (Anhui Province, China). Toxicon 2010, 56, 1520–1524. [Google Scholar] [CrossRef] [PubMed]
- Pawlik-Skowrońska, B.; Toporowska, M. Blooms of toxin-producing cyanobacteria—A real threat in small dam reservoirs at the beginning of their operation. Oceanol. Hydrobiol. Stud. 2011, 40, 30–37. [Google Scholar] [CrossRef]
- Rzymski, P.; Brygider, A.; Kokociński, M. On the occurence and toxicity of Cylindrospermopsis raciborskii. Limnol. Rev. 2017, 17, 23–29. [Google Scholar] [CrossRef] [Green Version]
- Mur, L.R.; Skulberg, O.M.; Utkilen, H. Cyanobacteria in the environment. In Toxic Cyanobacteria in Water. A Guide to the Their Public Health Consequences, Monitoring and Management; Chorus, I., Bartram, J., Eds.; Taylor & Francis: London, UK; New York, NY, USA, 1999; pp. 15–40. [Google Scholar]
- Rigosi, A.; Hanson, P.; Hamilton, D.P.; Hipsey, M.; Rusak, J.A.; Bois, J.; Sparber, K.; Chorus, I.; Watkinson, A.J.; Qin, B.; et al. Determination the probability of cyanobacterial blooms: The application of Bayesian networks in multiple lake systems. Ecol. Appl. 2015, 186–199. [Google Scholar] [CrossRef]
- Nõges, T.; Laugaste, R.; Nõges, P.; Tõnno, I. Critical N:P ratio for cyanobacteria and N2-fixing species in the large shallow temperate lakes Peipsi and Võrtsjãrv, North East Europe. Hydrobiologia 2008, 599, 77–86. [Google Scholar] [CrossRef]
- Filstrup, C.T.; Heathcote, A.J.; Kendall, D.L.; Downing, J.A. Phytoplankton taxonomic compositional shifts across nutrient and light gradients in temperate lakes. Inland Waters 2016, 6, 234–249. [Google Scholar] [CrossRef] [Green Version]
- Beversdorf, L.J.; Ryde, K.; Weirich, C.A.; Bartlett, S.L.; Seaman, M.; Kozik, C.; Biese, P.; Gosz, T.; Suha, M.; Stempa, C.; et al. Analysis of cyanobacterial metabolites in surface water and raw drinking waters reveals more than microcystins. Water Res. 2018, 140, 280–290. [Google Scholar] [CrossRef]
- Peretyatko, A.; Teissier, S.; Backer, S.D.; Triest, L. Assessment of the risk of cyanobacterial bloom occurrence in urban ponds: Probabilistic approach. Ann. Lilmnol. Int. J. Limnol. 2010, 46, 121–133. [Google Scholar] [CrossRef]
- Kokociński, M.; Stefaniak, K.; Izydorczyk, K.; Jurczak, T.; Mankiewicz-Boczek, J.; Soininen, J. Temporal variation in microcystin production by Planktothrix agardhii (Gomont) Anagnostidis and Komárek (Cyanobacteria, Oscillatoriales) in a temperate lake. Ann. Limnol. Int. J. Limnol. 2011, 47, 363–371. [Google Scholar] [CrossRef] [Green Version]
- Lu, J.; Zhu, B.; Struewing, I.; Xu, N.; Duan, S. Nitrogen–phosphorus-associated metabolic activities during the development of a cyanobacterial bloom revealed by metatranscriptomics. Sci. Rep. 2019, 9, 2480. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- DeMott, W.R.; Zhang, Q.X.; Carmichael, W.W. Effects of toxic cyanobacteria and purified toxins on the survival and feeding of a copepod and three species of Daphnia. Limnol. Oceanogr. 1991, 36, 1346–1357. [Google Scholar] [CrossRef] [Green Version]
- Pawlik-Skowrónska, B.; Toporowska, M.; Mazur-Marzec, H. Effects of secondary metabolites produced by different cyanobacterial populations on the freshwater zooplankters Brachionus calyciflorus and Daphnia pulex. Environ. Sci. Pollut. Res. 2019, 26, 11793–11804. [Google Scholar] [CrossRef] [Green Version]
- Piontek, M.; Czyżewska, W. Efficiency of drinking water treatment processes. Removal of phytoplankton with special consideration for cyanobacteria and improving physical and chemical parameters. Pol. J. Environ. Stud. 2012, 21, 1797–1805. [Google Scholar]
- Method 8038, Ammonia Measurement Interferences; Water Analysis Hach, Handbook; Hach Company: Loveland, Colorado, 2003.
- ISO 7887. Water Quality. Examination and Determination of Colour; International Standards Organization: Geneva, Switzerland, 2011. [Google Scholar]
- ISO 5814. Water Quality. Determination of Dissolved Oxygen Electrochemical Probe Method; Standards Organization: Geneva, Switzerland, 2012. [Google Scholar]
- ISO 10304-1. Water Quality. Determination of Dissolved Anions by Liquid Chromatography of Ions—Part 1: Determination of Bromide, Chloride, Fluoride, Nitrate, Nitrite, Phosphate and Sulfate; International Standards Organization: Geneva, Switzerland, 2007. [Google Scholar]
- Method 8048, Phosphorus Reactive; Water Analysis Hach, Handbook; Hach Company: Loveland, Colorado, 2003.
- ISO 10523. Water Quality. Determination of PH; International Standards Organization: Geneva, Switzerland, 2008. [Google Scholar]
- Hach Lange Test Kit No. 138. Available online: https://uk.hach.com/laton-total-nitrogen-cuvette-test-1-16-mg-l-tn-25-tests/product-downloads?id=26370268941 (accessed on 1 June 2017).
- Hach Lange Test Kit No. 349. Available online: https://uk.hach.com/phosphate-ortho-total-cuvette-test-0-05-1-5-mg-l-po-sub-4-sub-p-25-tests/product-downloads?id=26370291448 (accessed on 1 December 2017).
- ISO 872. Water Quality. Determination of Suspended Solids. Method by Filtration through Glass Fibre Filters; International Standards Organization: Geneva, Switzerland, 2005. [Google Scholar]
- PN-C-05560-02. Water and Wastewater. Determination of Chlorophyll in Surface Water. Determination of Chlorophyll Alpha in Plankton Algae by Monochromatic Spectrophotometric Method with Correction for Alpha Feopigments; The Polish Committee for Standardization: Warsaw, Poland, 1986. [Google Scholar]
- Almuhtaram, H.; Cui, Y.; Zamyadi, A.; Hofmann, R. Cyanotoxins and cyanobacteria cell accumulations in drinking water treatment plants with a low risk of bloom formation at the source. Toxins 2018, 10, 430. [Google Scholar] [CrossRef] [Green Version]
- Piontek, M. The regenerative ability of the planarian Dugesia tigrina (Girard) and the possibility of its use in reproduction of this species. Acta Hydrobiol. 1984, 25, 81–88. [Google Scholar]
- Piontek, M. Application of Dugesia tigrina Girard in toxicological studies of aquatic environments. Pol. Arch. Hydrobiol. 1998, 45, 565–572. [Google Scholar]
- Piontek, M. Use of the planarian Dugesia tigrina Girard in studies of acute intoxication. Pol. Arch. Hydrobiol. 1999, 46, 41–48. [Google Scholar]
- Weber, E. Grundriss der biologischen Statistik für Naturwissenschaftler, Landwirte und Mediziner; G. Fischer Verlag: Jena, Germany, 1972; pp. 1–674. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sampling Months [month/year] | Chlorophyll a [µg/L] | Cyanobacteria Biovolume mm3/L] | Dominant * Species | Percentage of Cyanobacteria Biovolume |
|---|---|---|---|---|
| SADOWO | ||||
| 07/2019 | 2.14 | n.d.** | - | |
| 08/2019 | 1.07 | 0.05 | Pseudanabaena limnetica | 100% |
| 09/2019 | 3.20 | 0.12 | Pseudanabaena limnetica | 96% |
| 10/2019 | 3.47 | 5.21 | Microcystis aeruginosa | 85% |
| UŚCIE | ||||
| 07/2019 | 32.8 | 59.2 | Dolichospermum flos-aquae | 96% |
| 08/2019 | 88.1 | 78.6 | Microcystis aeruginosa Dolichospermum flos-aquae | 49% 36% |
| 09/2019 | 79.0 | 88.5 | Microcystis aeruginosa | 89% |
| 10/2019 | 20.8 | 36.9 | Planktothrix agardhii | 69% |
| WOJNOWO | ||||
| 07/2019 | 6.14 | 3.52 | Dolichospermum flos-aquae | 65% |
| 08/2019 | 28.8 | 7.93 | Dolichospermum flos-aquae | 63% |
| 09/2019 | 28.6 | 5.51 | Oscillatoria angusta | 76% |
| 10/2019 | 7.21 | 1.06 | Oscillatoria angusta Microcystis wesenbergii | 47%31% |
| Sampling Months [month/year] | CYANOTOXINS [µg/L] | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| ANA | CYN | dmMC- RR | MC- RR | dmMC- LR | MC-LF | MC- LR | MC- LY | MC- LW | MC- YR | ∑MCs | |
| SADOWO | |||||||||||
| 07/2019 | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. |
| 08/2019 | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. |
| 09/2019 | n.d. | n.d. | n.d. | <0.01 | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. |
| 10/2019 | <0.01 | n.d. | n.d. | 0.01 | 0.73 | 0.03 | 0.06 | <0.01 | <0.01 | n.d. | 0.83 |
| UŚCIE | |||||||||||
| 07/2019 | n.d. | n.d. | n.d. | <0.01 | n.d. | 0.03 | <0.01 | n.d. | n.d. | n.d. | 0.03 |
| 08/2019 | n.d. | n.d. | n.d. | 0.14 | n.d. | 0.06 | 0.32 | <0.01 | <0.01 | 0.09 | 0.61 |
| 09/2019 | 0.05 | n.d. | 1.56 | 0.58 | n.d. | 0.21 | 1.33 | 0.03 | 0.03 | 0.23 | 3.97 |
| 10/2019 | 0.02 | n.d. | 1.66 | 0.17 | n.d. | 0.06 | 0.23 | 0.01 | 0.01 | 0.07 | 2.21 |
| WOJNOWO | |||||||||||
| 07/2019 | 0.01 | n.d. | n.d. | <0.01 | n.d. | 0.07 | <0.01 | n.d. | n.d. | n.d. | 0.07 |
| 08/2019 | 0.47 | 0.02 | n.d. | <0.01 | n.d. | 0.01 | n.d. | n.d. | <0.01 | n.d. | 0.01 |
| 09/2019 | 0.56 | 0.01 | n.d. | 0.3 | 0.41 | 0.02 | n.d. | <0.01 | <0.01 | n.d. | 0.46 |
| 10/2019 | 0.15 | 0.01 | n.d. | 0.01 | 0.73 | n.d. | n.d. | <0.01 | <0.01 | 0.13 | 0.86 |
| Sampling Months/Sites | SADOWO | UŚCIE | WOJNOWO |
|---|---|---|---|
| 07/2019 | n.t. | n.t. | n.t. |
| 08/2019 | n.t. | 41.7% | n.t. |
| 09/2019 | n.t. | 20.9% | n.t. |
| 10/2019 | n.t. | 35.5% | n.t. |
| Water Quality Indicators | Cya | ANA | dmMC-RR | MC-RR | dmMC-LR | MC-LR | ∑MCs |
|---|---|---|---|---|---|---|---|
| pH | 0.69 | 0.00 | −0.27 | 0.71 | −0.16 | 0.70 | 0.55 |
| NH4 | 0.36 | 0.17 | 0.31 | −0.06 | −0.37 | −0.11 | −0.05 |
| Dissolved oxygen | 0.24 | 0.38 | −0.46 | −0.24 | 0.20 | −0.26 | −0.31 |
| Ptot | 0.43 | −0.50 | 0.46 | 0.70 | −0.40 | 0.70 | 0.68 |
| Ntot | 0.60 | −0.22 | 0.00 | 0.91 | −0.43 | 0.91 | 0.77 |
| PO4 | 0.07 | −0.51 | 0.52 | 0.49 | −0.33 | 0.49 | 0.51 |
| Color | 0.03 | −0.31 | 0.41 | 0.33 | −0.29 | 0.32 | 0.32 |
| Turbidity | 0.84 | 0.01 | −0.23 | 0.79 | −0.27 | 0.81 | 0.61 |
| Total suspension | 0.84 | 0.22 | −0.09 | 0.77 | −0.18 | 0.74 | 0.69 |
| Water temperature | 0.15 | 0.22 | −0.36 | −0.06 | −0.46 | −0.02 | −0.39 |
| N:P | 0.02 | 0.47 | −0.40 | 0.04 | −0.05 | 0.03 | −0.08 |
| Physical–Chemical Indicators | Measurement | Equipment | Standards |
|---|---|---|---|
| Ammonium nitrogen | Spectrophotometric | spectrometer DR 5000, Hach Germany | WAH HACH 2003 met. 8038 [54] |
| Color | Visual | - | EN ISO 7887:2011 [55] |
| Dissolved oxygen | Electrochemical | oxygen meter HQ30d, Hach Germany | EN ISO 5814:2012 [56] |
| Nitrate nitrogen | Ion chromatography | ion chromatograph 881 IC Compact Pro, Metrohm Switzerland | EN ISO 10304-1:2009 [57] |
| Orthophosphate | Spectrophotometric | spectrometer DR 5000, Hach Germany | WAH HACH 1997 met. 8048 [58] |
| pH | Electrochemical | pH meter 540 GLP, WTW Germany | EN ISO 10523:2012 [59] |
| Temperature | Electrochemical | oxygen meter HQ30d, Hach Germany | Manufacturer‘s instructions |
| Total nitrogen | Spectrophotometric | spectrometer DR 5000, Hach Germany | Cuvette Test LCK 138, Hach [60] |
| Total phosphorus | Spectrophotometric | spectrometer DR 5000, Hach Germany | Cuvette Test LCK 349, Hach [61] |
| Total suspended solids | Gravimetric | weight CP224S-OCE, Sartorius, Germany | EN 872:2005 [62] |
| Turbidity | Nephelometric | turbidimeter 2100 IS AN, Hach Germany | Manufacturer‘s instructions |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Czyżewska, W.; Piontek, M.; Łuszczyńska, K. The Occurrence of Potential Harmful Cyanobacteria and Cyanotoxins in the Obrzyca River (Poland), a Source of Drinking Water. Toxins 2020, 12, 284. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins12050284
Czyżewska W, Piontek M, Łuszczyńska K. The Occurrence of Potential Harmful Cyanobacteria and Cyanotoxins in the Obrzyca River (Poland), a Source of Drinking Water. Toxins. 2020; 12(5):284. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins12050284
Chicago/Turabian StyleCzyżewska, Wanda, Marlena Piontek, and Katarzyna Łuszczyńska. 2020. "The Occurrence of Potential Harmful Cyanobacteria and Cyanotoxins in the Obrzyca River (Poland), a Source of Drinking Water" Toxins 12, no. 5: 284. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins12050284