Neuroprotective Fragment C of Tetanus Toxin Modulates IL-6 in an ALS Mouse Model

 , , ,
, , ,
Abstract
:1. Introduction
2. Results
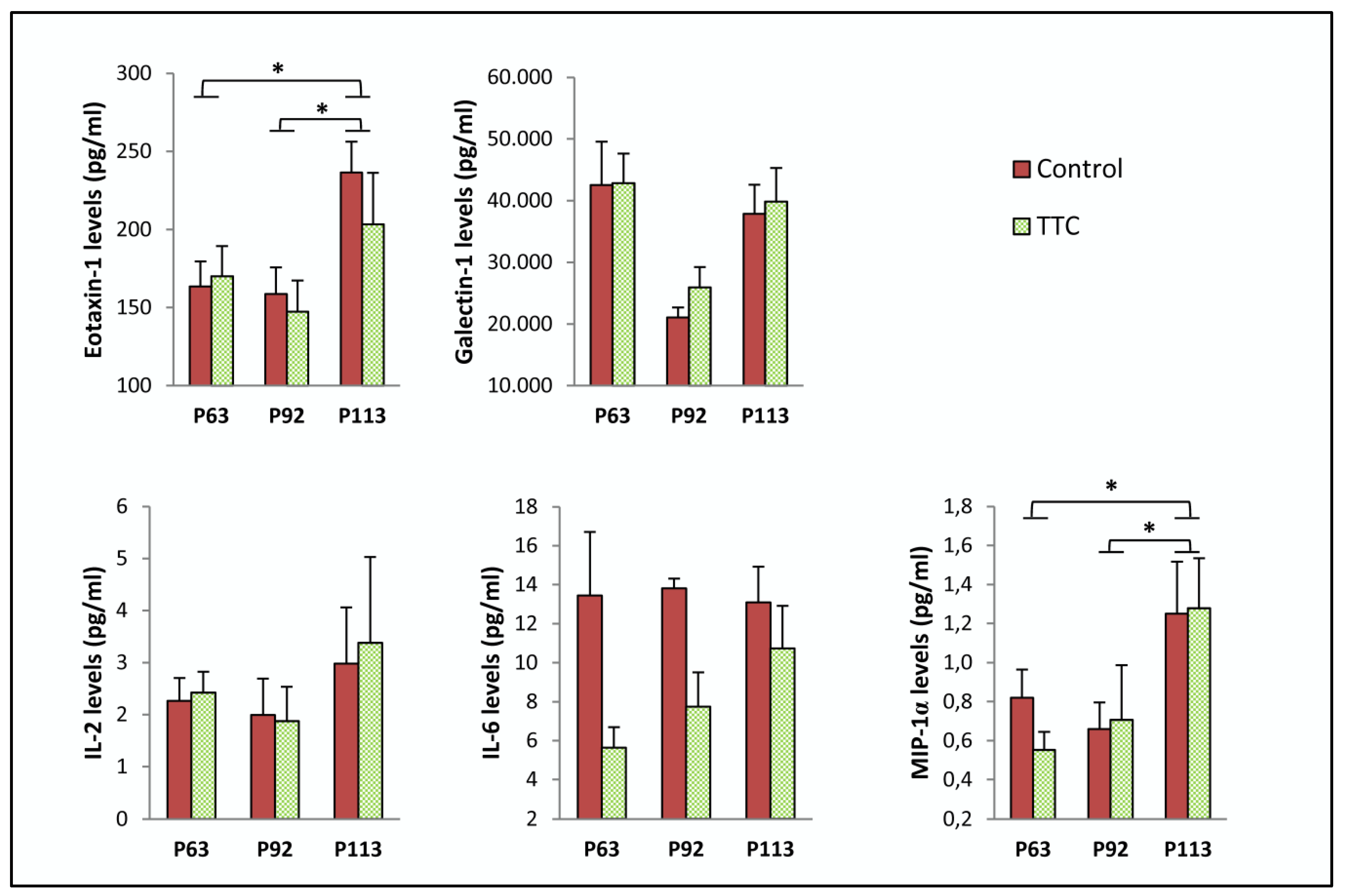
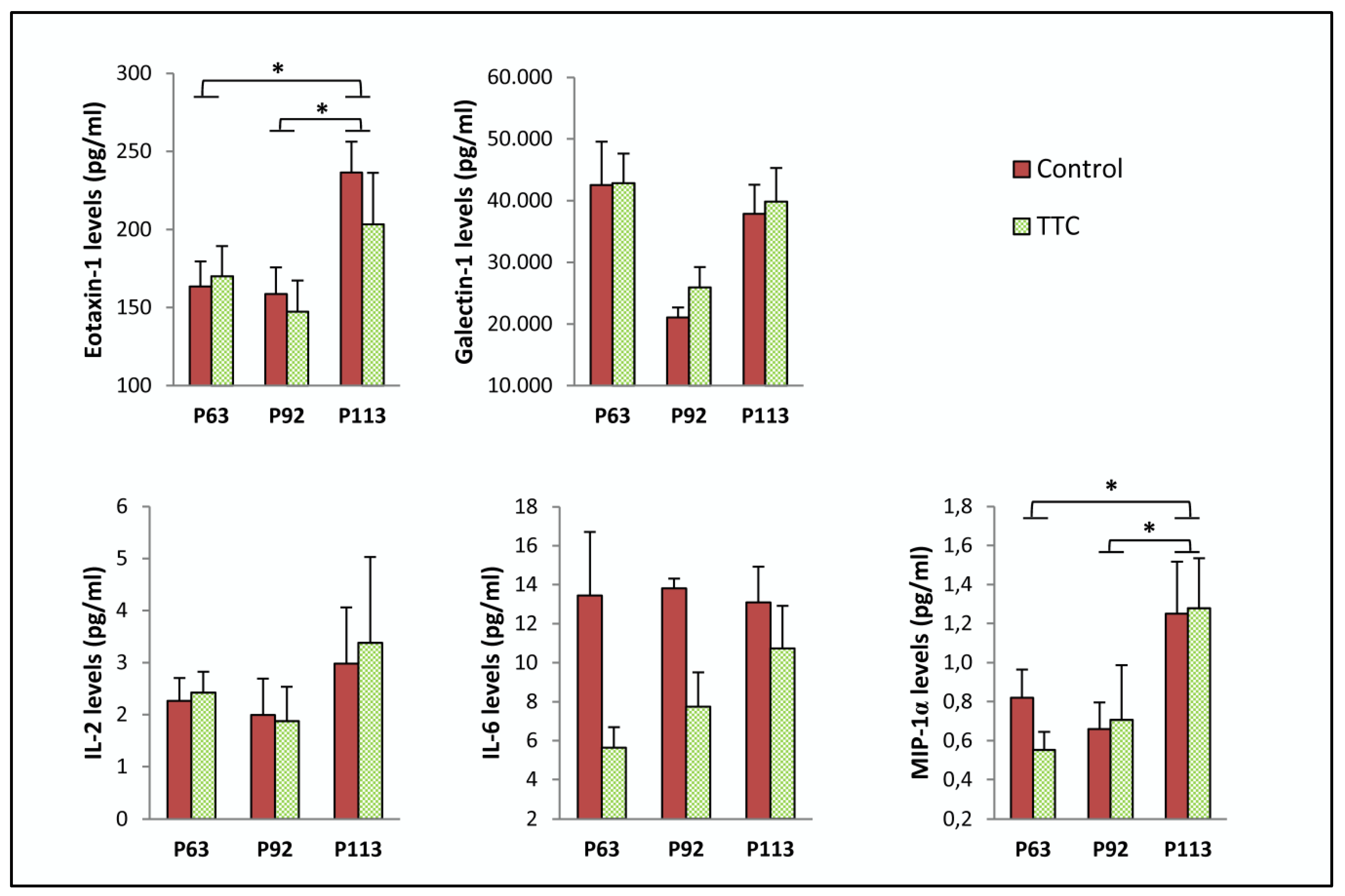
2.1. TTC Did Not Reduce Any Cytokine in Plasma from SOD1G93A Mice
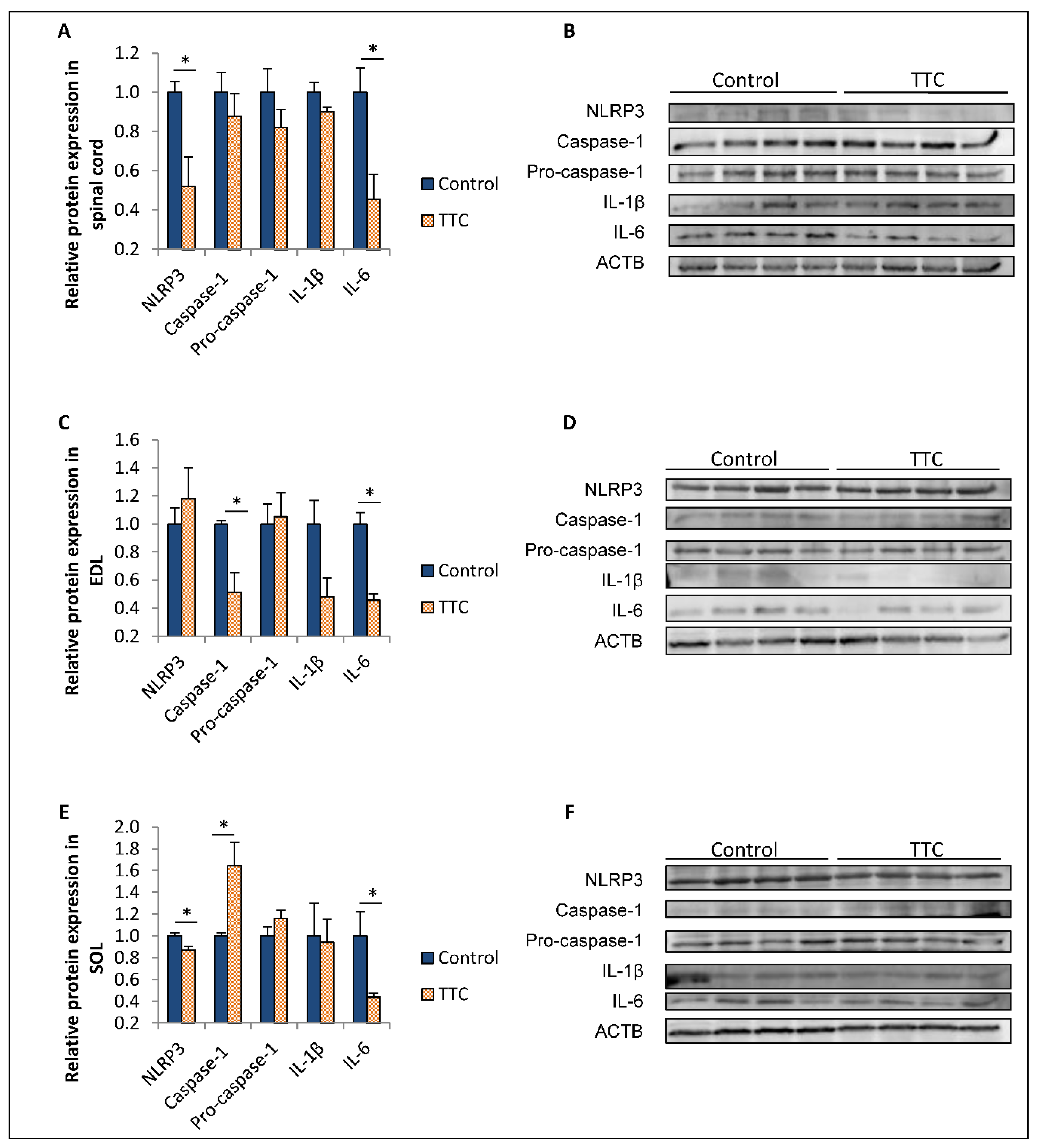
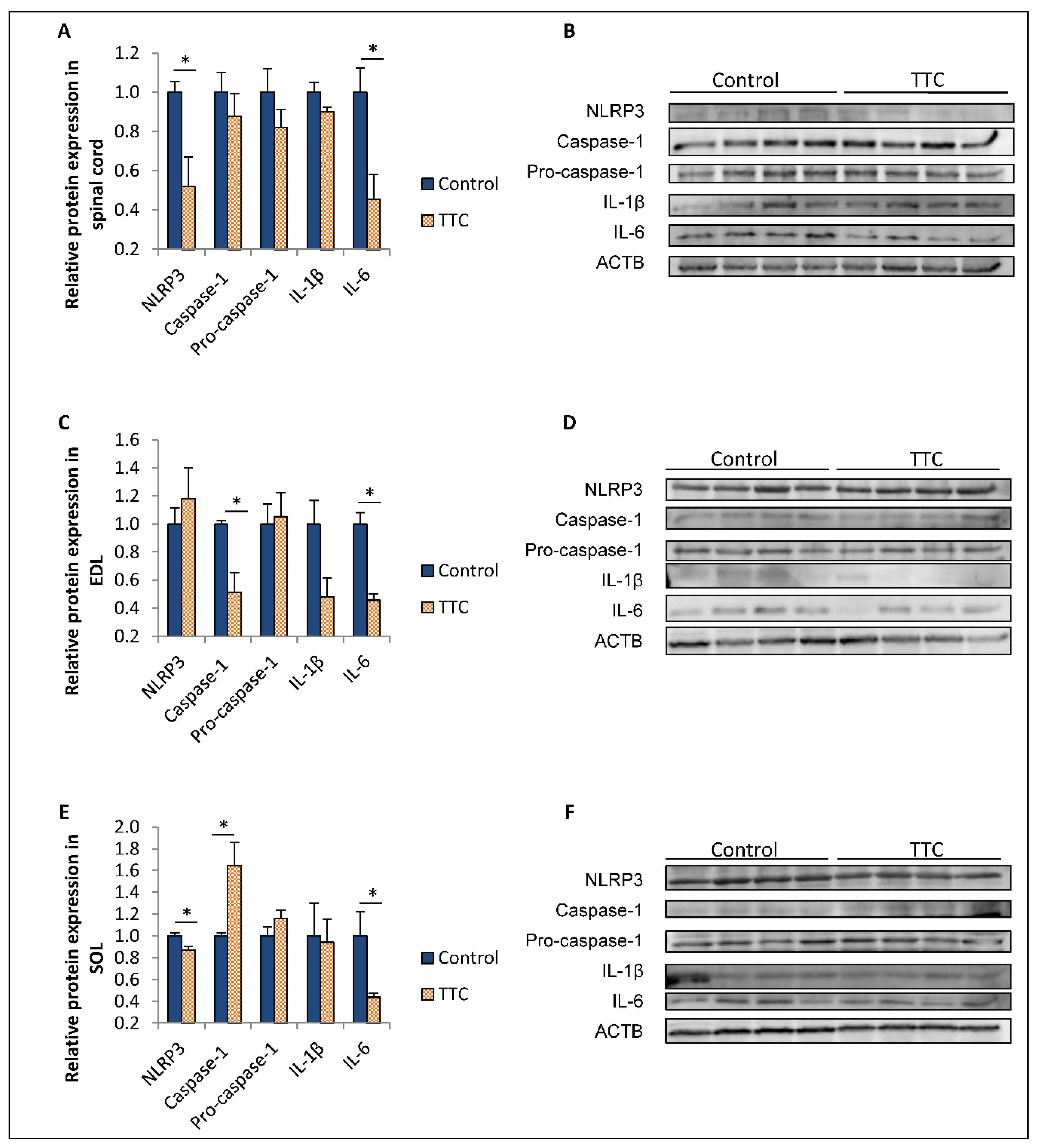
2.2. TTC Differently Modulates the Expression of NLRP3, Caspase-1 and IL-6 in Spinal Cord and Two Types of Skeletal Muscle Fibers from SOD1G93A Mice
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Animals
5.2. Treatment with Recombinant TTC Protein
5.3. Sample Collection
5.4. ELISA and Multiplex Immunoassays
5.5. Western Blotting
5.6. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Scott, A. Drug Therapy: On the Treatment Trail for ALS. Nature 2017, 550, S120–S121. [Google Scholar] [CrossRef] [PubMed]
- Ransohoff, R.M. How Neuroinflammation Contributes to Neurodegeneration. Science 2016, 353, 777–783. [Google Scholar] [CrossRef] [PubMed]
- Thonhoff, J.R.; Simpson, E.P.; Appel, S.H. Neuroinflammatory Mechanisms in Amyotrophic Lateral Sclerosis Pathogenesis. Curr. Opin. Neurol. 2018, 31, 635–639. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Wang, F. Role of Neuroinflammation in Amyotrophic Lateral Sclerosis: Cellular Mechanisms and Therapeutic Implications. Front. Immunol. 2017, 8, 1005. [Google Scholar] [CrossRef] [Green Version]
- Moreno-Martinez, L.; Calvo, A.C.; Munoz, M.J.; Osta, R. Are Circulating Cytokines Reliable Biomarkers for Amyotrophic Lateral Sclerosis? Int. J. Mol. Sci. 2019, 20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bellezza, I.; Grottelli, S.; Costanzi, E.; Scarpelli, P.; Pigna, E.; Morozzi, G.; Mezzasoma, L.; Peirce, M.J.; Moresi, V.; Adamo, S.; et al. Peroxynitrite Activates the NLRP3 Inflammasome Cascade in SOD1(G93A) Mouse Model of Amyotrophic Lateral Sclerosis. Mol. Neurobiol. 2018, 55, 2350–2361. [Google Scholar] [CrossRef] [Green Version]
- Debye, B.; Schmulling, L.; Zhou, L.; Rune, G.; Beyer, C.; Johann, S. Neurodegeneration and NLRP3 Inflammasome Expression in the Anterior Thalamus of SOD1(G93A) ALS Mice. Brain Pathol. 2018, 28, 14–27. [Google Scholar] [CrossRef]
- Gugliandolo, A.; Giacoppo, S.; Bramanti, P.; Mazzon, E. NLRP3 Inflammasome Activation in a Transgenic Amyotrophic Lateral Sclerosis Model. Inflammation 2018, 41, 93–103. [Google Scholar] [CrossRef]
- He, Y.; Hara, H.; Nunez, G. Mechanism and Regulation of NLRP3 Inflammasome Activation. Trends Biochem. Sci. 2016, 41, 1012–1021. [Google Scholar] [CrossRef] [Green Version]
- Michaelson, N.; Facciponte, D.; Bradley, W.; Stommel, E. Cytokine Expression Levels in ALS: A Potential Link between Inflammation and BMAA-Triggered Protein Misfolding. Cytokine Growth Factor Rev. 2017, 37, 81–88. [Google Scholar] [CrossRef]
- Kim, S.H.; Oh, K.W.; Jin, H.K.; Bae, J.S. Immune Inflammatory Modulation as a Potential Therapeutic Strategy of Stem Cell Therapy for ALS and Neurodegenerative Diseases. BMB Rep. 2018, 51, 545–546. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thonhoff, J.R.; Beers, D.R.; Zhao, W.; Pleitez, M.; Simpson, E.P.; Berry, J.D.; Cudkowicz, M.E.; Appel, S.H. Expanded Autologous Regulatory T-Lymphocyte Infusions in ALS: A Phase I, First-in-Human Study. Neurol. Neuroimmunol. Neuroinflamm. 2018, 5, e465. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beers, D.R.; Appel, S.H. Immune Dysregulation in Amyotrophic Lateral Sclerosis: Mechanisms and Emerging Therapies. Lancet Neurol. 2019, 18, 211–220. [Google Scholar] [CrossRef]
- Toivonen, J.M.; Olivan, S.; Osta, R. Tetanus Toxin C-Fragment: The Courier and the Cure? Toxins 2010, 2, 2622–2644. [Google Scholar] [CrossRef] [Green Version]
- Calvo, A.C.; Moreno-Igoa, M.; Mancuso, R.; Manzano, R.; Olivan, S.; Munoz, M.J.; Penas, C.; Zaragoza, P.; Navarro, X.; Osta, R. Lack of a Synergistic Effect of a Non-Viral ALS Gene Therapy Based on BDNF and a TTC Fusion Molecule. Orphanet J. Rare Dis. 2011, 6, 10. [Google Scholar] [CrossRef] [Green Version]
- Moreno-Igoa, M.; Calvo, A.C.; Penas, C.; Manzano, R.; Olivan, S.; Munoz, M.J.; Mancuso, R.; Zaragoza, P.; Aguilera, J.; Navarro, X.; et al. Fragment C of Tetanus Toxin, More than a Carrier. Novel Perspectives in Non-Viral ALS Gene Therapy. J. Mol. Med. 2010, 88, 297–308. [Google Scholar] [CrossRef]
- Ciriza, J.; Moreno-Igoa, M.; Calvo, A.C.; Yague, G.; Palacio, J.; Miana-Mena, F.J.; Munoz, M.J.; Zaragoza, P.; Brulet, P.; Osta, R. A Genetic Fusion GDNF-C Fragment of Tetanus Toxin Prolongs Survival in a Symptomatic Mouse ALS Model. Restor. Neurol. Neurosci. 2008, 26, 459–465. [Google Scholar]
- Ehrhart, J.; Smith, A.J.; Kuzmin-Nichols, N.; Zesiewicz, T.A.; Jahan, I.; Shytle, R.D.; Kim, S.H.; Sanberg, C.D.; Vu, T.H.; Gooch, C.L.; et al. Humoral Factors in ALS Patients during Disease Progression. J. Neuroinflamm. 2015, 12, 127. [Google Scholar] [CrossRef] [Green Version]
- Ngo, S.T.; Steyn, F.J.; Huang, L.; Mantovani, S.; Pfluger, C.M.; Woodruff, T.M.; O’Sullivan, J.D.; Henderson, R.D.; McCombe, P.A. Altered Expression of Metabolic Proteins and Adipokines in Patients with Amyotrophic Lateral Sclerosis. J. Neurol. Sci. 2015, 357, 22–27. [Google Scholar] [CrossRef] [Green Version]
- Lu, C.H.; Allen, K.; Oei, F.; Leoni, E.; Kuhle, J.; Tree, T.; Fratta, P.; Sharma, N.; Sidle, K.; Howard, R.; et al. Systemic Inflammatory Response and Neuromuscular Involvement in Amyotrophic Lateral Sclerosis. Neurol. Neuroimmunol. Neuroinflamm. 2016, 3, e244. [Google Scholar] [CrossRef] [Green Version]
- Blasco, H.; Garcon, G.; Patin, F.; Veyrat-Durebex, C.; Boyer, J.; Devos, D.; Vourc’h, P.; Andres, C.R.; Corcia, P. Panel of Oxidative Stress and Inflammatory Biomarkers in ALS: A Pilot Study. Can. J. Neurol. Sci. 2017, 44, 90–95. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, Y.; Cao, C.; Qin, X.Y.; Yu, Y.; Yuan, J.; Zhao, Y.; Cheng, Y. Increased Peripheral Blood Inflammatory Cytokine Levels in Amyotrophic Lateral Sclerosis: A Meta-Analysis Study. Sci. Rep. 2017, 7, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Prado, L.G.R.; Rocha, N.P.; de Souza, L.C.; Bicalho, I.C.S.; Gomez, R.S.; Vidigal-Lopes, M.; Braz, N.F.T.; Vieira, E.L.M.; Teixeira, A.L. Longitudinal Assessment of Clinical and Inflammatory Markers in Patients with Amyotrophic Lateral Sclerosis. J. Neurol. Sci. 2018, 394, 69–74. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Xia, K.; Chen, L.; Fan, D. Increased Interleukin-6 Levels in the Astrocyte-Derived Exosomes of Sporadic Amyotrophic Lateral Sclerosis Patients. Front. Neurosci. 2019, 13, 574. [Google Scholar] [CrossRef] [Green Version]
- Moreno-Martinez, L.; de la Torre, M.; Toivonen, J.M.; Zaragoza, P.; Garcia-Redondo, A.; Calvo, A.C.; Osta, R. Circulating Cytokines could Not be Good Prognostic Biomarkers in a Mouse Model of Amyotrophic Lateral Sclerosis. Front. Immunol. 2019, 10, 801. [Google Scholar] [CrossRef]
- Moreau, C.; Devos, D.; Brunaud-Danel, V.; Defebvre, L.; Perez, T.; Destee, A.; Tonnel, A.B.; Lassalle, P.; Just, N. Elevated IL-6 and TNF-Alpha Levels in Patients with ALS: Inflammation Or Hypoxia? Neurology 2005, 65, 1958–1960. [Google Scholar] [CrossRef]
- Pronto-Laborinho, A.; Pinto, S.; Gromicho, M.; Pereira, M.; Swash, M.; de Carvalho, M. Interleukin-6 and Amyotrophic Lateral Sclerosis. J. Neurol. Sci. 2019, 398, 50–53. [Google Scholar] [CrossRef]
- Qin, X.Y.; Zhang, S.P.; Cao, C.; Loh, Y.P.; Cheng, Y. Aberrations in Peripheral Inflammatory Cytokine Levels in Parkinson Disease: A Systematic Review and Meta-Analysis. JAMA Neurol. 2016, 73, 1316–1324. [Google Scholar] [CrossRef]
- Lai, K.S.P.; Liu, C.S.; Rau, A.; Lanctot, K.L.; Kohler, C.A.; Pakosh, M.; Carvalho, A.F.; Herrmann, N. Peripheral Inflammatory Markers in Alzheimer’s Disease: A Systematic Review and Meta-Analysis of 175 Studies. J. Neurol. Neurosurg. Psychiatry. 2017, 88, 876–882. [Google Scholar] [CrossRef]
- Madaro, L.; Passafaro, M.; Sala, D.; Etxaniz, U.; Lugarini, F.; Proietti, D.; Alfonsi, M.V.; Nicoletti, C.; Gatto, S.; De Bardi, M.; et al. Denervation-Activated STAT3-IL-6 Signalling in Fibro-Adipogenic Progenitors Promotes Myofibres Atrophy and Fibrosis. Nat. Cell Biol. 2018, 20, 917–927. [Google Scholar] [CrossRef]
- Sironi, F.; Vallarola, A.; Violatto, M.B.; Talamini, L.; Freschi, M.; De Gioia, R.; Capelli, C.; Agostini, A.; Moscatelli, D.; Tortarolo, M.; et al. Multiple Intracerebroventricular Injections of Human Umbilical Cord Mesenchymal Stem Cells Delay Motor Neurons Loss but Not Disease Progression of SOD1G93A Mice. Stem Cell Res. 2017, 25, 166–178. [Google Scholar] [CrossRef] [PubMed]
- Korhonen, P.; Pollari, E.; Kanninen, K.M.; Savchenko, E.; Lehtonen, S.; Wojciechowski, S.; Pomeshchik, Y.; Van Den Bosch, L.; Goldsteins, G.; Koistinaho, J.; et al. Long-Term Interleukin-33 Treatment Delays Disease Onset and Alleviates Astrocytic Activation in a Transgenic Mouse Model of Amyotrophic Lateral Sclerosis. IBRO Rep. 2019, 6, 74–86. [Google Scholar] [CrossRef] [PubMed]
- Lu, J.; Zheng, Y.; Yang, J.; Zhang, J.; Cao, W.; Chen, X.; Fang, S. Resveratrol Alleviates Inflammatory Injury and Enhances the Apoptosis of Fibroblastlike Synoviocytes Via Mitochondrial Dysfunction and ER Stress in Rats with Adjuvant Arthritis. Mol. Med. Rep. 2019, 20, 463–472. [Google Scholar] [PubMed] [Green Version]
- Kuhle, J.; Lindberg, R.L.; Regeniter, A.; Mehling, M.; Steck, A.J.; Kappos, L.; Czaplinski, A. Increased Levels of Inflammatory Chemokines in Amyotrophic Lateral Sclerosis. Eur. J. Neurol. 2009, 16, 771–774. [Google Scholar] [CrossRef]
- Yang, X.; Gao, L.; Wu, X.; Zhang, Y.; Zang, D. Increased Levels of MIP-1alpha in CSF and Serum of ALS. Acta Neurol. Scand. 2016, 134, 94–100. [Google Scholar] [CrossRef]
- Guo, J.; Yang, X.; Gao, L.; Zang, D. Evaluating the Levels of CSF and Serum Factors in ALS. Brain Behav. 2017, 7, e00637. [Google Scholar] [CrossRef]
- Meissner, F.; Molawi, K.; Zychlinsky, A. Mutant Superoxide Dismutase 1-Induced IL-1beta Accelerates ALS Pathogenesis. Proc. Natl. Acad. Sci. USA 2010, 107, 13046–13050. [Google Scholar] [CrossRef] [Green Version]
- Johann, S.; Heitzer, M.; Kanagaratnam, M.; Goswami, A.; Rizo, T.; Weis, J.; Troost, D.; Beyer, C. NLRP3 Inflammasome is Expressed by Astrocytes in the SOD1 Mouse Model of ALS and in Human Sporadic ALS Patients. Glia 2015, 63, 2260–2273. [Google Scholar] [CrossRef]
- Lehmann, S.; Esch, E.; Hartmann, P.; Goswami, A.; Nikolin, S.; Weis, J.; Beyer, C.; Johann, S. Expression Profile of Pattern Recognition Receptors in Skeletal Muscle of SOD1((G93A)) Amyotrophic Lateral Sclerosis (ALS) Mice and Sporadic ALS Patients. Neuropathol. Appl. Neurobiol. 2018, 44, 606–627. [Google Scholar] [CrossRef]
- Okumura, N.; Hashida-Okumura, A.; Kita, K.; Matsubae, M.; Matsubara, T.; Takao, T.; Nagai, K. Proteomic Analysis of Slow- and Fast-Twitch Skeletal Muscles. Proteomics 2005, 5, 2896–2906. [Google Scholar] [CrossRef]
- Jia, W.H.; Wang, N.Q.; Yin, L.; Chen, X.; Hou, B.Y.; Qiang, G.F.; Chan, C.B.; Yang, X.Y.; Du, G.H. Effect of Skeletal Muscle Phenotype and Gender on Fasting-Induced Myokine Expression in Mice. Biochem. Biophys. Res. Commun. 2019, 514, 407–414. [Google Scholar] [CrossRef] [PubMed]
- Grottelli, S.; Mezzasoma, L.; Scarpelli, P.; Cacciatore, I.; Cellini, B.; Bellezza, I. Cyclo(His-Pro) Inhibits NLRP3 Inflammasome Cascade in ALS Microglial Cells. Mol. Cell. Neurosci. 2019, 94, 23–31. [Google Scholar] [CrossRef] [PubMed]
- Heitzer, M.; Kaiser, S.; Kanagaratnam, M.; Zendedel, A.; Hartmann, P.; Beyer, C.; Johann, S. Administration of 17beta-Estradiol Improves Motoneuron Survival and Down-Regulates Inflammasome Activation in Male SOD1(G93A) ALS Mice. Mol. Neurobiol. 2017, 54, 8429–8443. [Google Scholar] [CrossRef] [PubMed]
- Manzano, R.; Toivonen, J.M.; Olivan, S.; Calvo, A.C.; Moreno-Igoa, M.; Munoz, M.J.; Zaragoza, P.; Garcia-Redondo, A.; Osta, R. Altered Expression of Myogenic Regulatory Factors in the Mouse Model of Amyotrophic Lateral Sclerosis. Neurodegener Dis. 2011, 8, 386–396. [Google Scholar] [CrossRef]
- Manzano, R.; Toivonen, J.M.; Calvo, A.C.; Munoz, M.J.; Zaragoza, P.; Osta, R. Housekeeping Gene Expression in Myogenic Cell Cultures from Neurodegeneration and Denervation Animal Models. Biochem. Biophys. Res. Commun. 2011, 407, 758–763. [Google Scholar] [CrossRef]
- Manzano, R.; Toivonen, J.M.; Calvo, A.C.; Olivan, S.; Zaragoza, P.; Munoz, M.J.; Montarras, D.; Osta, R. Quantity and Activation of Myofiber-Associated Satellite Cells in a Mouse Model of Amyotrophic Lateral Sclerosis. Stem Cell. Rev. Rep. 2012, 8, 279–287. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
| Protein | Age | Control | TTC | p-Value |
|---|---|---|---|---|
| Eotaxin-1 (pg/mL) | P63 | 163.407 ± 15.992 | 170.119 ± 19.104 | 0.799 |
| P92 | 158.566 ± 17.151 | 147.400 ± 19.803 | 0.687 | |
| P113 | 236.529 ± 19.625 | 203.274 ± 33.074 | 0.412 | |
| Galectin-1 (pg/mL) | P63 | 42,562.638 ± 6977.809 | 42,872.392 ± 4757.342 | 0.971 |
| P92 | 21,041.705 ± 1657.878 | 25,930.396 ± 3328.63 | 0.192 | |
| P113 | 37,897.955 ± 4724.214 | 39,839.61 ± 5479.457 | 0.798 | |
| IL-2 (pg/mL) | P63 | 2.262 ± 0.442 | 2.420 ± 0.403 | 0.816 |
| P92 | 1.994 ± 0.697 | 1.870 ± 0.667 | 0.902 | |
| P113 | 2.980 ± 1.077 | 3.376 ± 1.654 | 0.853 | |
| IL-6 (pg/mL) | P63 | 13.451 ± 3.255 | 5.626 ± 1.064 | 0.062 |
| P92 | 13.810 ± 0.502 | 7.735 ± 1.764 | * 0.021 | |
| P113 | 13.097 ± 1.828 | 10.727 ± 2.188 | 0.429 | |
| MIP-1α (pg/mL) | P63 | 0.820 ± 0.144 | 0.552 ± 0.093 | 0.157 |
| P92 | 0.659 ± 0.137 | 0.706 ± 0.281 | 0.877 | |
| P113 | 1.250 ± 0.267 | 1.278 ± 0.256 | 0.941 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Moreno-Martinez, L.; de la Torre, M.; Muñoz, M.J.; Zaragoza, P.; Aguilera, J.; Calvo, A.C.; Osta, R. Neuroprotective Fragment C of Tetanus Toxin Modulates IL-6 in an ALS Mouse Model. Toxins 2020, 12, 330. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins12050330
Moreno-Martinez L, de la Torre M, Muñoz MJ, Zaragoza P, Aguilera J, Calvo AC, Osta R. Neuroprotective Fragment C of Tetanus Toxin Modulates IL-6 in an ALS Mouse Model. Toxins. 2020; 12(5):330. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins12050330
Chicago/Turabian StyleMoreno-Martinez, Laura, Miriam de la Torre, María J. Muñoz, Pilar Zaragoza, José Aguilera, Ana C. Calvo, and Rosario Osta. 2020. "Neuroprotective Fragment C of Tetanus Toxin Modulates IL-6 in an ALS Mouse Model" Toxins 12, no. 5: 330. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins12050330