In Vitro Assessment of Biocontrol Effects on Fusarium Head Blight and Deoxynivalenol (DON) Accumulation by DON-Degrading Bacteria

 ,
,
Abstract
:1. Introduction
2. Results
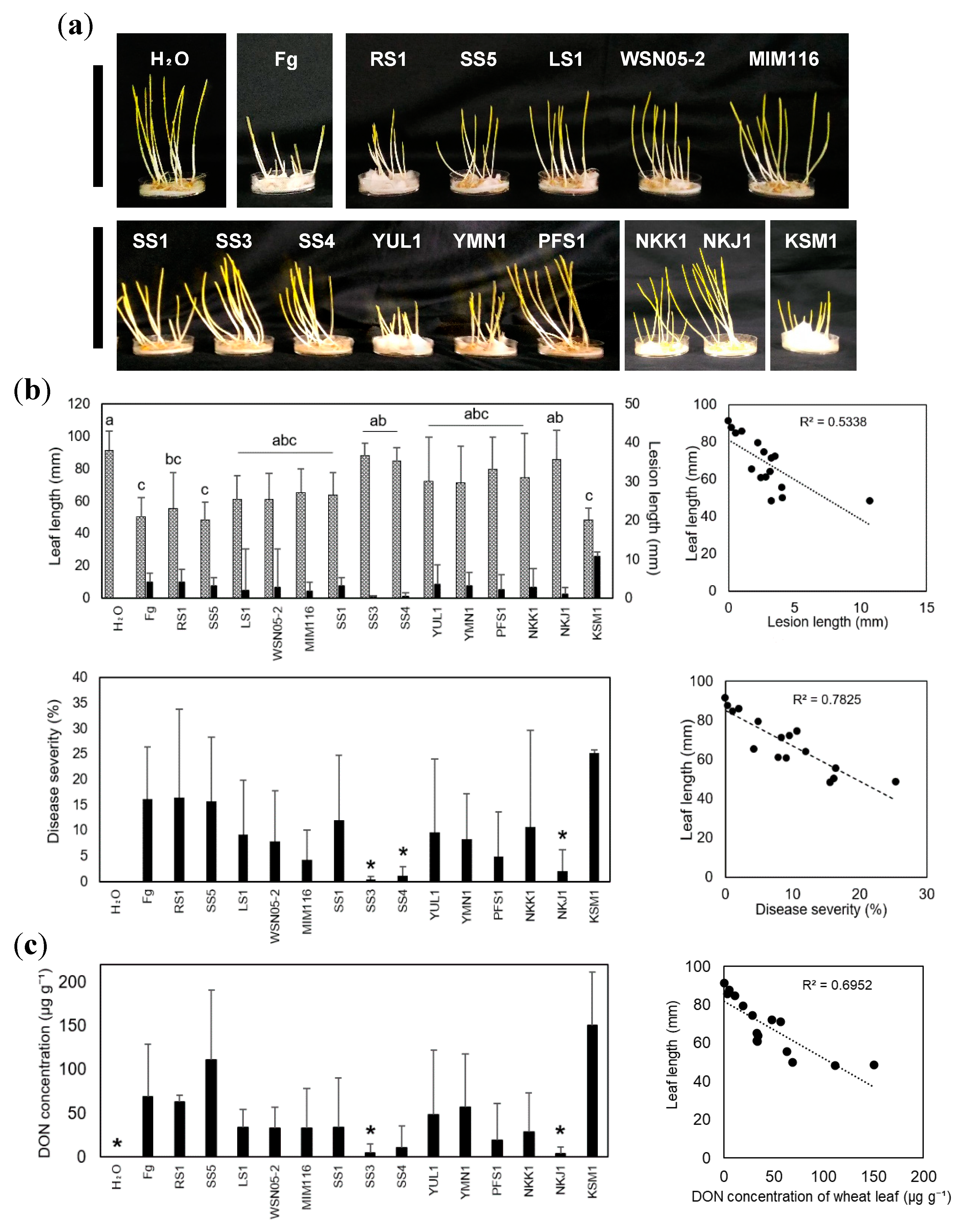
2.1. Devosia sp. NKJ1 and Nocardioides spp. SS3 and SS4 Suppress the Disease Progression of Fg
2.2. Devosia sp. NKJ1 and Nocardioides spp. SS3 and SS4 Prevent the DON Accumulation in Wheat Leaves
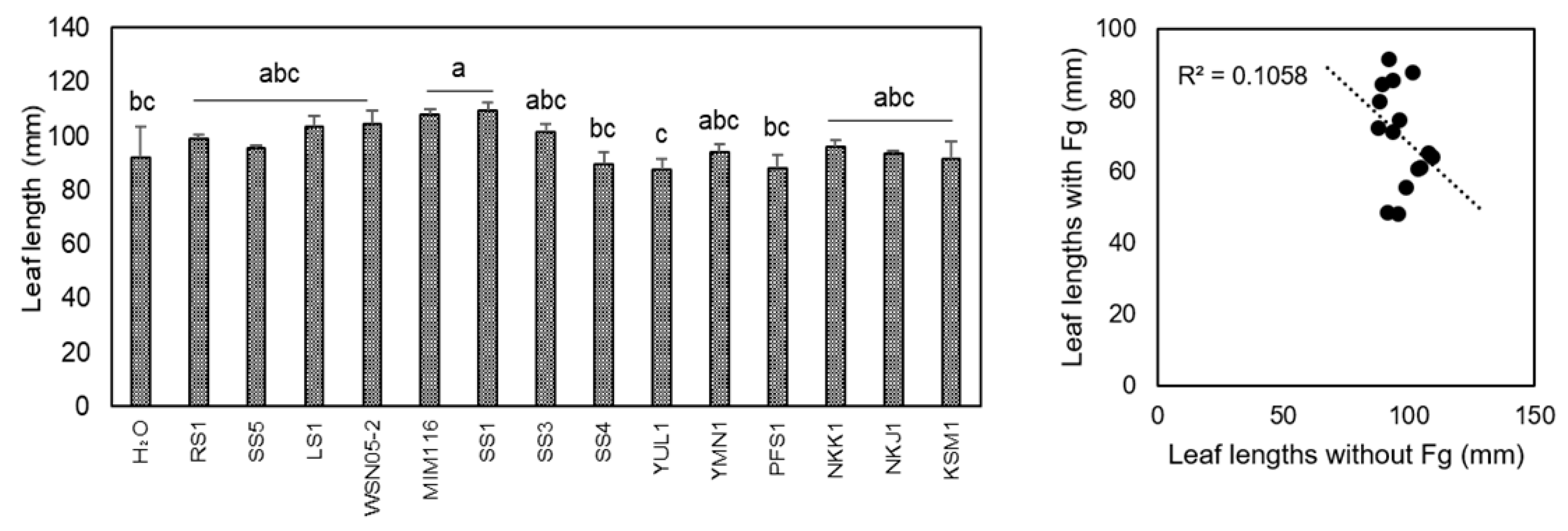
2.3. Devosia sp. NKJ1 and Nocardioides spp. SS3 or SS4 Did Not Promote Wheat Growth
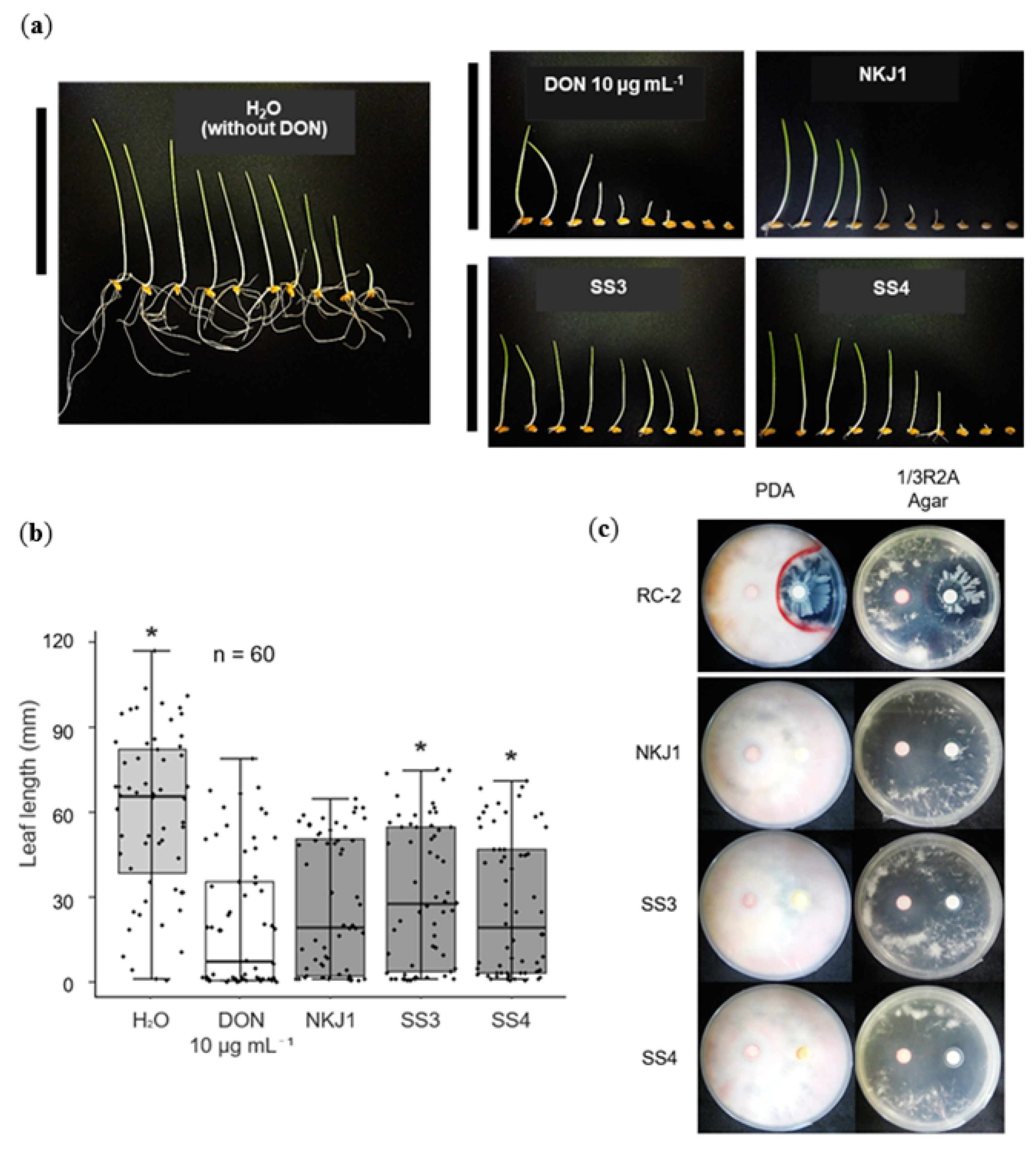
2.4. Nocardioides spp. SS3 and SS4 Reduce the Effects of DON
2.5. None of the DDBs Showed Antagonistic Activity
3. Discussion
4. Materials and Methods
4.1. Chemicals, Strains, and Media
4.2. Fg Microconidia Preparation
4.3. DON-Degrading Bacteria Suspension
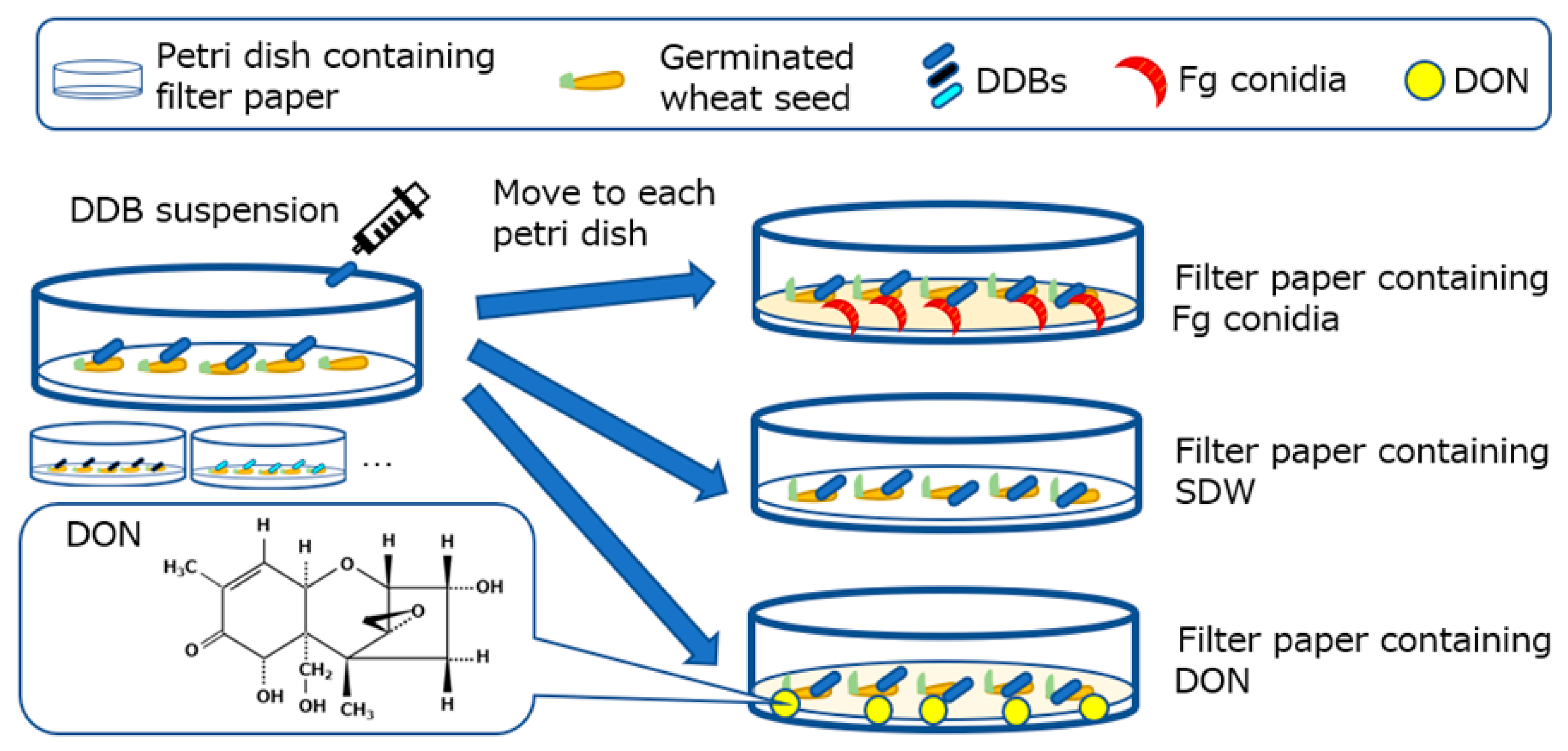
4.4. Wheat Seeds and Petri Dish Test
4.5. DON Extraction and HPLC Analysis
4.6. Antagonistic Activities Assay against F. graminearum on Solid Media
4.7. Statistical Analysis
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Windels, C.E. Economic and social impacts of Fusarium head blight: Changing farms and rural communities in the northern great plains. Phytopathology 2000, 90, 17–21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goswami, R.S.; Kistler, H.C. Heading for disaster: Fusarium graminearum on cereal crops. Mol. Plant Pathol. 2004, 5, 515–525. [Google Scholar] [CrossRef] [PubMed]
- Goswami, R.S.; Xu, J.R.; Trail, F.; Hilburn, K.; Kistler, H.C. Genomic analysis of host-pathogen interaction between Fusarium graminearum and wheat during early stages of disease development. Microbiology 2006, 152, 1877–1890. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parry, D.W.; Jenkinson, P.; McLeod, L. Fusarium ear blight (Scab) in small-grain cereals—A review. Plant Pathol. 1995, 44, 207–238. [Google Scholar] [CrossRef]
- Yoshida, M.; Nakajima, T. Deoxynivalenol and nivalenol accumulation in wheat infected with Fusarium graminearum during grain development. Phytopathology 2010, 100, 763–773. [Google Scholar] [CrossRef] [Green Version]
- Yoshizawa, T.; Jin, Y.Z. Natural occurrence of acetylated derivatives of deoxynivalenol and nivalenol in wheat and barley in japan. Food Addit. Contam. 1995, 12, 689–694. [Google Scholar] [CrossRef]
- Rotter, B.A.; Prelusky, D.B.; Pestka, J.J. Toxicology of deoxynivalenol (vomitoxin). J. Toxicol. Environ. Health 1996, 48, 1–34. [Google Scholar] [CrossRef]
- Pestka, J.J. Deoxynivalenol: Mechanisms of action, human exposure, and toxicological relevance. Arch. Environ. Contam. Toxicol. 2010, 84, 663–679. [Google Scholar] [CrossRef]
- Mesterházy, Á.; Bartók, T.; Lamper, C. Influence of wheat cultivar, species of Fusarium, and isolate aggressiveness on the efficacy of fungicides for control of Fusarium head blight. Plant Dis. 2003, 87, 1107–1115. [Google Scholar] [CrossRef] [Green Version]
- Chen, C.J.; Wang, J.X.; Luo, Q.Q.; Yuan, S.K.; Zhou, M.G. Characterization and fitness of carbendazim-resistant strains of Fusarium graminearum (wheat scab). Pest Manage. Sci. 2007, 63, 1201–1207. [Google Scholar] [CrossRef]
- Kogan, M. Integrated pest management: Historical perspectives and contemporary developments. Annu. Rev. Entomol. 1998, 43, 243–270. [Google Scholar] [CrossRef] [PubMed]
- Arrebola, E.; Jacobs, R.; Korsten, L. Iturin A is the principal inhibitor in the biocontrol activity of Bacillus amyloliquefaciens PPCB004 against postharvest fungal pathogens. J. Appl. Microbiol. 2010, 108, 386–395. [Google Scholar] [CrossRef] [PubMed]
- Crane, J.M.; Gibson, D.M.; Vaughan, R.H.; Bergstrom, G.C. Iturin levels on wheat spikes linked to biological control of fusarium head blight by Bacillus amyloliquefaciens. Phytopathology 2013, 103, 146–155. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gu, Q.; Yang, Y.; Yuan, Q.M.; Shi, G.M.; Wu, L.M.; Lou, Z.Y.; Huo, R.; Wu, H.J.; Borriss, R.; Gao, X.W. Bacillomycin D Produced by Bacillus amyloliquefaciens is involved in the antagonistic interaction with the plant-pathogenic fungus Fusarium graminearum. Appl. Environ. Microbiol. 2017, 83, 17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shi, C.J.; Yan, P.S.; Li, J.F.; Wu, H.Q.; Li, Q.W.; Guan, S.S. Biocontrol of Fusarium graminearum growth and deoxynivalenol production in wheat kernels with bacterial antagonists. Int. J. Environ. Res. Public Health 2014, 11, 1094–1105. [Google Scholar] [CrossRef] [Green Version]
- Scheepmaker, J.W.A.; Busschers, M.; Sundh, I.; Eilenberg, J.; Butt, T.M. Sense and nonsense of the secondary metabolites data requirements in the EU for beneficial microbial control agents. Biol. Control 2019, 136, 10. [Google Scholar] [CrossRef]
- Audenaert, K.; Vanheule, A.; Höfte, M.; Haesaert, G. Deoxynivalenol: A major player in the multifaceted response of Fusarium to its environment. Toxins 2014, 6, 1–19. [Google Scholar] [CrossRef] [Green Version]
- Desmond, O.J.; Manners, J.M.; Stephens, A.E.; MaClean, D.J.; Schenk, P.M.; Gardiner, D.M.; Munn, A.L.; Kazan, K. The Fusarium mycotoxin deoxynivalenol elicits hydrogen peroxide production, programmed cell death and defence responses in wheat. Mol. Plant Pathol. 2008, 9, 435–445. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kazan, K.; Gardiner, D.M.; Manners, J.M. On the trail of a cereal killer: Recent advances in Fusarium graminearum pathogenomics and host resistance. Mol. Plant Pathol. 2012, 13, 399–413. [Google Scholar] [CrossRef]
- Jansen, C.; von Wettstein, D.; Schäfer, W.; Kogel, K.H.; Felk, A.; Maier, F.J. Infection patterns in barley and wheat spikes inoculated with wild-type and trichodiene synthase gene disrupted Fusarium graminearum. Proc. Natl. Acad. Sci. USA 2005, 102, 16892–16897. [Google Scholar] [CrossRef] [Green Version]
- Miura, I.; Kamakura, T.; Maeno, S.; Hayashi, S.; Yamaguchi, I. Inhibition of Enzyme-secretion in plant-pathogens by mepanipyrim, a novel fungicide. Pestic. Biochem. Physiol. 1994, 48, 222–228. [Google Scholar] [CrossRef]
- Howell, C.R. Mechanisms employed by Trichoderma species in the biological control of plant diseases: The history and evolution of current concepts. Plant Dis. 2003, 87, 4–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Köhl, J.; Kolnaar, R.; Ravensberg, W.J. Mode of action of microbial biological control agents against plant diseases: Relevance beyond efficacy. Front. Plant. Sci. 2019, 10, 19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ouchi, S.; Toyoda, H.; Utsumi, R.; Hashimoto, H.; Hadama, T. A promising strategy for the control of fungal diseases by the use of toxin-degrading microbes. In Phytotoxins and Plant Pathogenesis; Graniti, A., Durbin, R.D., Ballio, A., Eds.; Springer: Berlin/Heidelberg, Germany, 1989; pp. 301–317. [Google Scholar]
- Achari, G.A.; Ramesh, R. Characterization of bacteria degrading 3-hydroxy palmitic acid methyl ester (3OH-PAME), a quorum sensing molecule of Ralstonia solanacearum. Lett. Appl. Microbiol. 2015, 60, 447–455. [Google Scholar] [CrossRef] [PubMed]
- Ji, C.; Fan, Y.; Zhao, L.H. Review on biological degradation of mycotoxins. Anim Nutr. 2016, 2, 127–133. [Google Scholar] [CrossRef]
- Hassan, Y.I.; Zhou, T. Addressing the mycotoxin deoxynivalenol contamination with soil-derived bacterial and enzymatic transformations targeting the C3 carbon. World Mycotoxin J. 2018, 11, 101–111. [Google Scholar] [CrossRef]
- Toyoda, H.; Hashimoto, H.; Utsumi, R.; Kobayashi, H.; Ouchi, S. Detoxification of fusaric acid by a fusaric acid-resistant mutant of Pseudomonas solanacearum and its application to biological-control of fusarium-wilt of tomato. Phytopathology 1988, 78, 1307–1311. [Google Scholar] [CrossRef]
- Kosawang, C.; Karlsson, M.; Vélëz, H.; Rasmussen, P.H.; Collinge, D.B.; Jensen, B.; Jensen, D.F. Zearalenone detoxification by zearalenone hydrolase is important for the antagonistic ability of Clonostachys rosea against mycotoxigenic Fusarium graminearum. Fungal Biol. 2014, 118, 364–373. [Google Scholar] [CrossRef]
- Parry, D.W.; Bayles, D.W.; Priestley, R.H. Resistance of winter wheat varieties to ear blight (F. culmorum). J. Natl. Inst. Agric. Bot. 1984, 16, 465–468. [Google Scholar]
- Wu, A.B.; Li, H.P.; Zhao, C.S.; Liao, Y.C. Comparative pathogenicity of Fusarium graminearum isolates from China revealed by wheat coleoptile and floret inoculations. Mycopathologia 2005, 160, 75–83. [Google Scholar] [CrossRef]
- Mesterházy, Á. Comparative Analysis of artificial inoculation methods with Fusarium spp. on winter-wheat varieties. J. Phytopathol. 1978, 93, 12–25. [Google Scholar] [CrossRef]
- Purahong, W.; Alkadri, D.; Nipoti, P.; Pisi, A.; Lemmens, M.; Prodi, A. Validation of a modified Petri-dish test to quantify aggressiveness of Fusarium graminearum in durum wheat. Eur. J. Plant Pathol. 2012, 132, 381–391. [Google Scholar] [CrossRef]
- Ikunaga, Y.; Sato, I.; Grond, S.; Numaziri, N.; Yoshida, S.; Yamaya, H.; Hiradate, S.; Hasegawa, M.; Toshima, H.; Koitabashi, M.; et al. Nocardioides sp. strain WSN05−2, isolated from a wheat field, degrades deoxynivalenol, producing the novel intermediate 3-epi-deoxynivalenol. Appl. Microbiol. Biotechnol. 2011, 89, 419–427. [Google Scholar] [CrossRef] [Green Version]
- Sato, I.; Ito, M.; Ishizaka, M.; Ikunaga, Y.; Sato, Y.; Yoshida, S.; Koitabashi, M.; Tsushima, S. Thirteen novel deoxynivalenol-degrading bacteria are classified within two genera with distinct degradation mechanisms. FEMS Microbiol. Lett. 2012, 327, 110–117. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ito, M.; Sato, I.; Koitabashi, M.; Yoshida, S.; Imai, M.; Tsushima, S. A novel actinomycete derived from wheat heads degrades deoxynivalenol in the grain of wheat and barley affected by Fusarium head blight. Appl. Microbiol. Biotechnol. 2012, 96, 1059–1070. [Google Scholar] [CrossRef]
- Ito, M.; Sato, I.; Ishizaka, M.; Yoshida, S.; Koitabashi, M.; Tsushima, S. Bacterial cytochrome P450 system catabolizing the Fusarium toxin deoxynivalenol. Appl. Environ. Microbiol. 2013, 79, 1619–1628. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suga, H.; Kageyama, K.; Shimizu, M.; Hyakumachi, M. A natural mutation involving both pathogenicity and perithecium formation in the Fusarium graminearum species complex. G3 Genes Genomes Genet. 2016, 6, 3883–3892. [Google Scholar] [CrossRef] [Green Version]
- Yoshida, S.; Hiradate, S.; Tsukamoto, T.; Hatakeda, K.; Shirata, A. Antimicrobial activity of culture filtrate of Bacillus amyloliquefaciens RC-2 isolated from mulberry leaves. Phytopathology 2001, 91, 181–187. [Google Scholar] [CrossRef] [Green Version]
- Brennan, J.M.; Fagan, B.; van Maanen, A.; Cooke, B.M.; Doohan, F.M. Studies on in vitro growth and pathogenicity of european Fusarium fungi. Eur. J. Plant Pathol. 2003, 109, 577–587. [Google Scholar] [CrossRef]
- Weller, D.M. Biological—Control of Soilborne plant-pathogens. Annu. Rev. Phytopathol 1988, 26, 379–407. [Google Scholar] [CrossRef]
- Trivedi, P.; Delgado-Baquerizo, M.; Trivedi, C.; Hamonts, K.; Anderson, I.C.; Singh, B.K. Keystone microbial taxa regulate the invasion of a fungal pathogen in agro-ecosystems. Soil Biol. Biochem. 2017, 111, 10–14. [Google Scholar] [CrossRef]
- Vurukonda, S.; Giovanardi, D.; Stefani, E. Plant growth promoting and biocontrol activity of Streptomyces spp. as endophytes. Int. J. Mol. Sci. 2018, 19, 952. [Google Scholar] [CrossRef] [Green Version]
- Aldesuquy, H.S.; Mansour, F.A.; Abo-Hamed, S.A. Effect of the culture filtrates of Streptomyces on growth and productivity of wheat plants. Folia Microbiol. 1998, 43, 465–470. [Google Scholar] [CrossRef]
- Jog, R.; Nareshkumar, G.; Rajkumar, S. Plant growth promoting potential and soil enzyme production of the most abundant Streptomyces spp. from wheat rhizosphere. J. Appl. Microbiol. 2012, 113, 1154–1164. [Google Scholar] [CrossRef] [PubMed]
- Clifford, L.J.; Jia, Q.S.; Pestka, J.J. An improved method for the purification of the trichothecene deoxynivalenol (Vomitoxin) from Fusarium graminearum culture. J. Agric. Food Chem. 2003, 51, 521–523. [Google Scholar] [CrossRef] [PubMed]
- Gale, L.R.; Bryant, J.D.; Calvo, S.; Giese, H.; Katan, T.; O’Donnell, K.; Suga, H.; Taga, M.; Usgaard, T.R.; Ward, T.J.; et al. Chromosome complement of the fungal plant pathogen Fusarium graminearum based on genetic and physical mapping and cytological observations. Genetics 2005, 171, 985–1001. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sulyok, M.; Berthiller, F.; Krska, R.; Schuhmacher, R. Development and validation of a liquid chromatography/tandem mass spectrometric method for the determination of 39 mycotoxins in wheat and maize. Rapid Commun. Mass Spectrom. 2006, 20, 2649–2659. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2018; Available online: https://www.R-project.org/ (accessed on 16 June 2020).





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Class | Strain [Ref. No.] | Source of Isolation | Closely Related Species (% 16S rRNA Gene Sequence Similarity) |
|---|---|---|---|
| Actinobacteria | WSN05-2 [34] | Wheat field soil | Nocardioides panacihumi Gsoil 616 (98.37) |
| SS3 [35] | Wheat field soil | Nocardioides panacihumi Gsoil 616 (98.47) | |
| YMN1 [35] | Uncultivated soil | Nocardioides panacihumi Gsoil 616 (98.33) | |
| SS1 [35] | Wheat field soil | Nocardioides panacihumi Gsoil 616 (98.98) | |
| YUL1 [35] | Wheat field soil | Nocardioides panacihumi Gsoil 616 (98.73) | |
| PFS1 [35] | Paddy field soil | Nocardioides panacihumi Gsoil 616 (98.72) | |
| SS4 [35] | Wheat field soil | Nocardioides ginsengisegetis Gsoil 485 (100) | |
| LS1 [35] | Wheat leaf | Nocardioides ginsengisegetis Gsoil 485 (100) | |
| MIM116 [36] | Wheat head | Marmoricola aequoreus SST-45 (98.80) | |
| Alphaproteobacteria | SS5 [35] † | Wheat field soil | Youhaiella tibetensis fig4 (99.84) |
| RS1 [35] † | Wheat field soil | Youhaiella tibetensis fig4 (99.10) | |
| NKJ1 [35] | Paddy field soil | Devosia insulae DS-56 (99.34) | |
| NKK1 [35] | Wheat field soil | Devosia insulae DS-56 (99.10) | |
| KSM1 [37] | Lake water | Sphingomonas naphthae DKC-5-1 (97.40) |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Morimura, H.; Ito, M.; Yoshida, S.; Koitabashi, M.; Tsushima, S.; Camagna, M.; Chiba, S.; Takemoto, D.; Kawakita, K.; Sato, I. In Vitro Assessment of Biocontrol Effects on Fusarium Head Blight and Deoxynivalenol (DON) Accumulation by DON-Degrading Bacteria. Toxins 2020, 12, 399. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins12060399
Morimura H, Ito M, Yoshida S, Koitabashi M, Tsushima S, Camagna M, Chiba S, Takemoto D, Kawakita K, Sato I. In Vitro Assessment of Biocontrol Effects on Fusarium Head Blight and Deoxynivalenol (DON) Accumulation by DON-Degrading Bacteria. Toxins. 2020; 12(6):399. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins12060399
Chicago/Turabian StyleMorimura, Hiroyuki, Michihiro Ito, Shigenobu Yoshida, Motoo Koitabashi, Seiya Tsushima, Maurizio Camagna, Sotaro Chiba, Daigo Takemoto, Kazuhito Kawakita, and Ikuo Sato. 2020. "In Vitro Assessment of Biocontrol Effects on Fusarium Head Blight and Deoxynivalenol (DON) Accumulation by DON-Degrading Bacteria" Toxins 12, no. 6: 399. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins12060399