The Comparative Toxicity of 10 Microcystin Congeners Administered Orally to Mice: Clinical Effects and Organ Toxicity

 ,
,
Abstract
:1. Introduction
2. Results
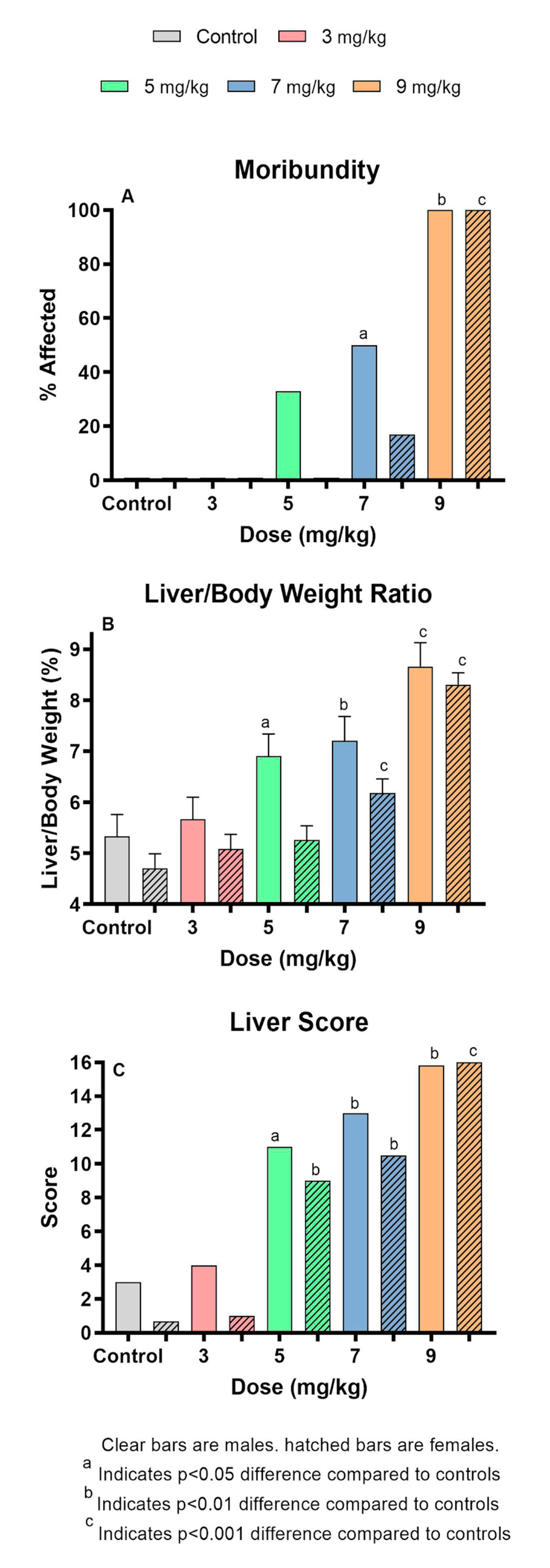
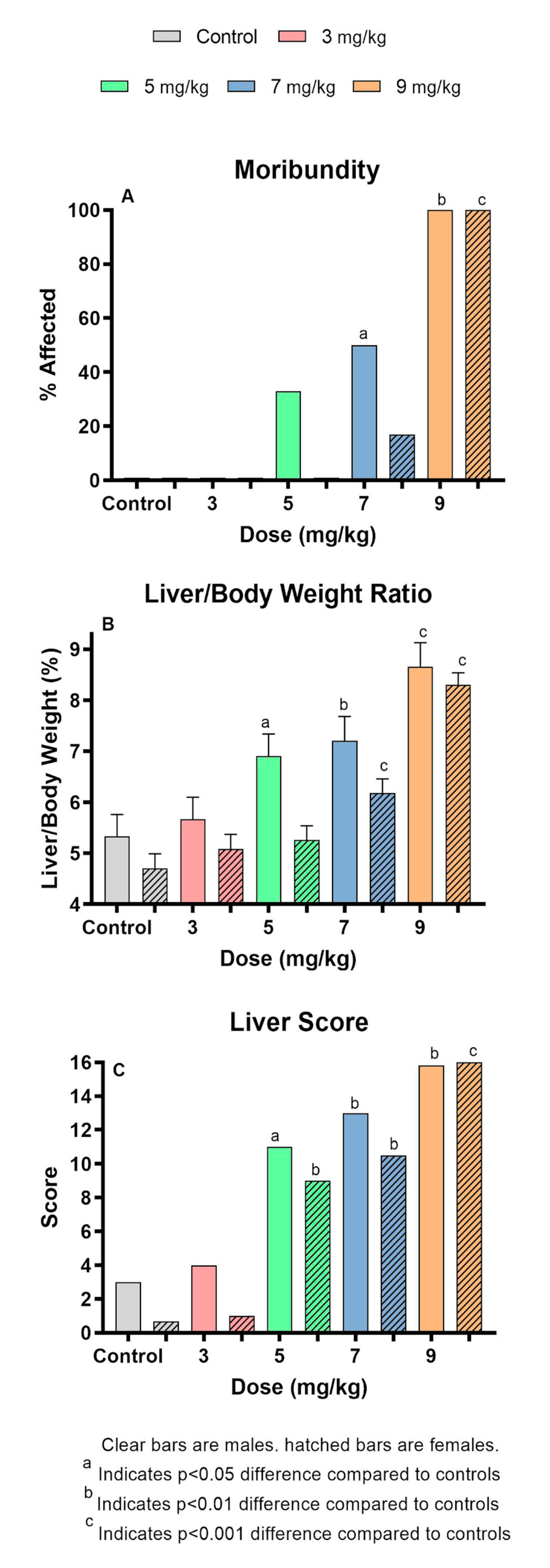
2.1. MCLR Dose Response After Oral Administration
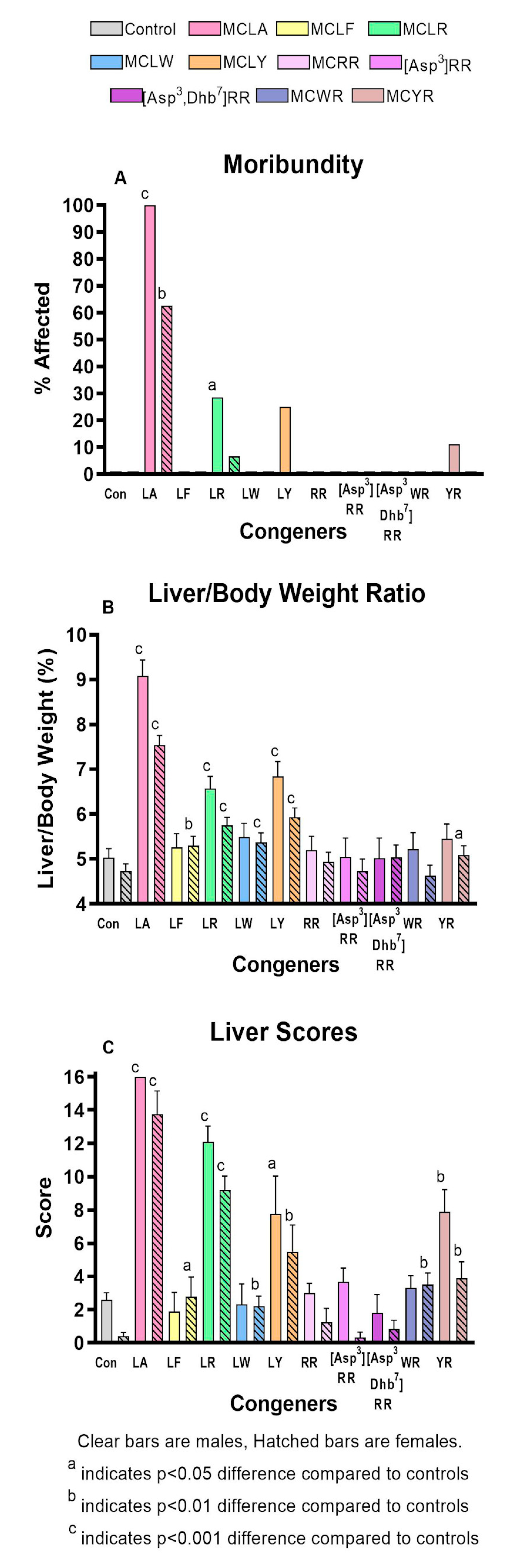
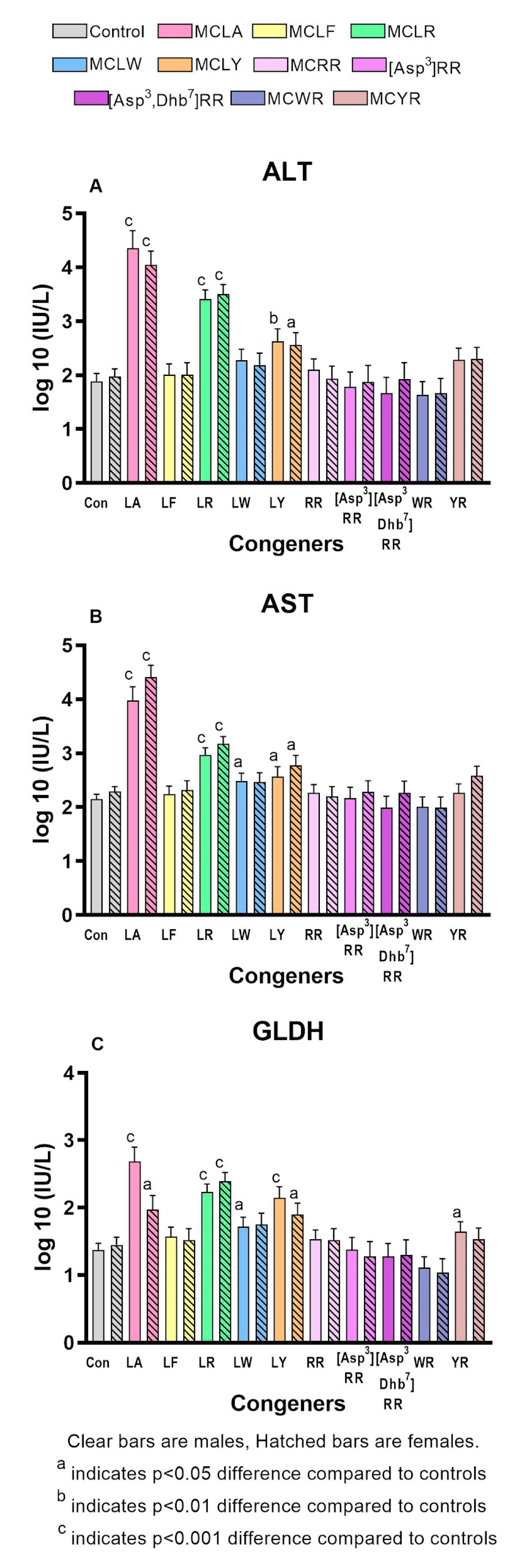
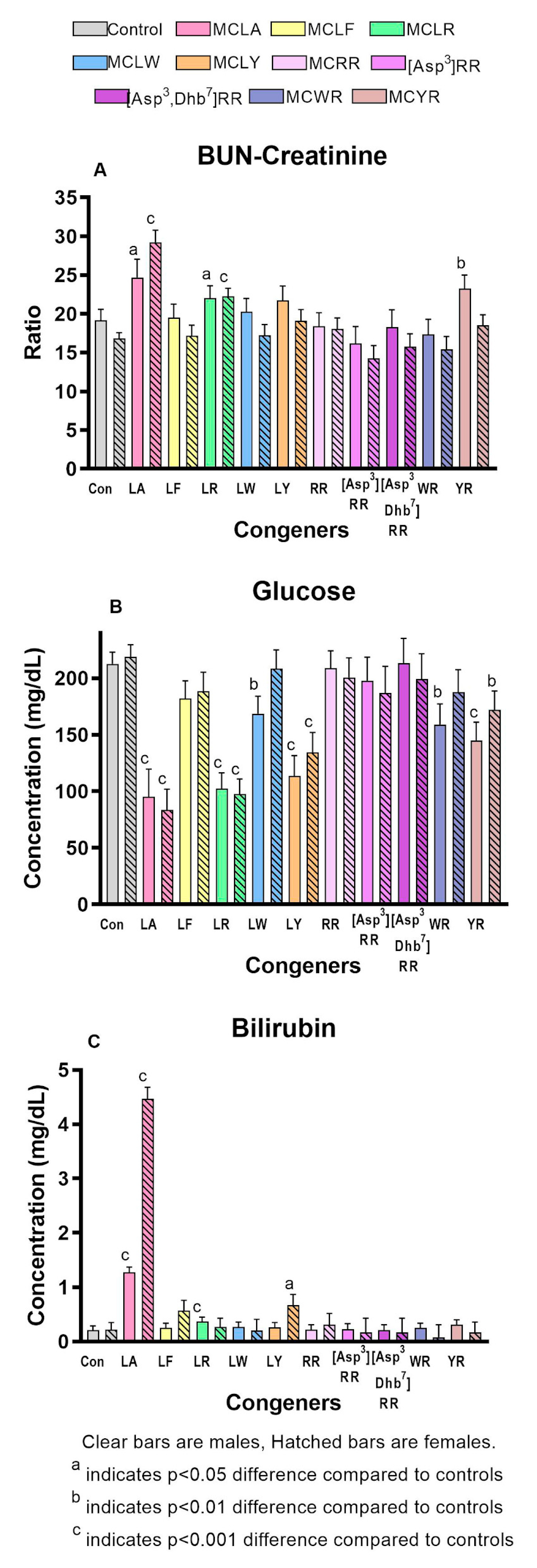
2.2. Comparative Toxicity of Microcystin Congeners MCLA, MCLF, MCLR, MCLW, MCLY, MCRR, [Asp3]MCRR, [Asp3,Dhb7]MCRR, MCWR, and MCYR Administered a Single Oral Dose of 7mg/kg
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Compounds
4.3. Dosing, Animal Observation and Necropsy
4.4. Clinical Chemistry
4.5. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Buratti, F.M.; Manganelli, M.; Vichi, S.; Stefanelli, M.; Scardala, S.; Testai, E.; Funari, E. Cyanotoxins: Producing organisms, occurrence, toxicity, mechanism of action and human health toxicological risk evaluation. Arch. Toxicol. 2017, 91, 1049–1130. [Google Scholar] [CrossRef]
- Merel, S.; Walker, D.; Chicana, R.; Snyder, S.; Baures, E.; Thomas, O. State of knowledge and concerns on cyanobacterial blooms and cyanotoxins. Environ. Int. 2013, 59, 303–327. [Google Scholar] [CrossRef] [PubMed]
- Paerl, H.W.; Otten, T.G. Harmful cyanobacterial blooms: Causes, consequences, and controls. Microb. Ecol. 2013, 65, 995–1010. [Google Scholar] [CrossRef] [PubMed]
- D’Anglada, L.V.; Donohue, J.M.; Strong, J.; Hawkins, B. Health Effects Support Document for the Cyanobacterial Toxin Microcystins; U.S. Environmental Protection Agency, Office of Water: Washington, DC, USA, 2015.
- Dolman, A.M.; Rucker, J.; Pick, F.R.; Fastner, J.; Rohrlack, T.; Mischke, U.; Wiedner, C. Cyanobacteria and cyanotoxins: The influence of nitrogen versus phosphorus. PLoS ONE 2012, 7, e38757. [Google Scholar] [CrossRef]
- Gobler, C.J.; Burkholder, J.M.; Davis, T.W.; Harke, M.J.; Johengen, T.; Stow, C.A.; Van de Waal, D.B. The dual role of nitrogen supply in controlling the growth and toxicity of cyanobacterial blooms. Harmful Algae 2016, 54, 87–97. [Google Scholar] [CrossRef]
- Paerl, H.W.; Xu, H.; Hall, N.S.; Zhu, G.; Qin, B.; Wu, Y.; Rossignol, K.L.; Dong, L.; McCarthy, M.J.; Joyner, A.R. Controlling cyanobacterial blooms in hypertrophic Lake Taihu, China: Will nitrogen reductions cause replacement of non-N2 fixing by N2 fixing taxa? PLoS ONE 2014, 9, e113123. [Google Scholar] [CrossRef] [Green Version]
- Yang, Y.; Colom, W.; Pierson, D.; Pettersson, K. Water column stability and summer phytoplankton dynamics in a temperate lake (Lake Erken, Sweden). Inland Waters 2016, 6, 499–508. [Google Scholar] [CrossRef] [Green Version]
- Johnk, K.D.; Huisman, J.; Sharples, J.; Sommeijer, B.; Visser, P.M.; Stroom, J.M. Summer heatwaves promote blooms of harmful cyanobacteria. Glob. Chang. Biol. 2008, 14, 495–512. [Google Scholar] [CrossRef] [Green Version]
- Paerl, H.W.; Huisman, J. Climate. Blooms like it hot. Science 2008, 320, 57–58. [Google Scholar] [CrossRef] [Green Version]
- Vasconcelos, V. Eutrophication, toxic cyanobacteria and cyanotoxins: When ecosystems cry for help. Limnetica 2006, 25, 425–432. [Google Scholar]
- Blaha, L.; Babica, P.; Marsalek, B. Toxins produced in cyanobacterial water blooms-toxicity and risks. Interdiscip. Toxicol. 2009, 2, 36–41. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mantzouki, E.; Lurling, M.; Fastner, J.; de Senerpont Domis, L.; Wilk-Wozniak, E.; Koreiviene, J.; Seelen, L.; Teurlincx, S.; Verstijnen, Y.; Krzton, W.; et al. Temperature Effects Explain Continental Scale Distribution of Cyanobacterial Toxins. Toxins 2018, 10, 156. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Araoz, R.; Molgo, J.; Tandeau de Marsac, N. Neurotoxic cyanobacterial toxins. Toxicon Off. J. Int. Soc. Toxinol. 2010, 56, 813–828. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, H.; Machii, K. Comparison of toxicity between saxitoxin and decarbamoyl saxitoxin in the mouse bioassay for paralytic shellfish poisoning toxins. J. Vet. Med. Sci. 2014, 76, 1523–1525. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.; Shen, D.; Fang, D. Nodularins in poisoning. Clin. Chim. Acta Int. J. Clin. Chem. 2013, 425, 18–29. [Google Scholar] [CrossRef]
- Dawson, R.M. The toxicology of microcystins. Toxicon Off. J. Int. Soc. Toxinol. 1998, 36, 953–962. [Google Scholar] [CrossRef]
- de la Cruz, A.A.; Hiskia, A.; Kaloudis, T.; Chernoff, N.; Hill, D.; Antoniou, M.G.; He, X.; Loftin, K.; O’Shea, K.; Zhao, C.; et al. A review on cylindrospermopsin: The global occurrence, detection, toxicity and degradation of a potent cyanotoxin. Environ. Sci. Process. Impacts 2013, 15, 1979–2003. [Google Scholar] [CrossRef]
- Rastogi, R.P.; Sinha, R.P.; Incharoensakdi, A. The cyanotoxin-microcystins: Current overview. Rev. Environ. Sci. Biol. 2014, 13, 215–249. [Google Scholar] [CrossRef]
- Loftin, K.A.; Graham, J.L.; Hilborn, E.D.; Lehmann, S.C.; Meyer, M.T.; Dietze, J.E.; Griffith, C.B. Cyanotoxins in inland lakes of the United States: Occurrence and potential recreational health risks in the EPA National Lakes Assessment 2007. Harmful Algae 2016, 56, 77–90. [Google Scholar] [CrossRef] [Green Version]
- Backer, L.C.; Manassaram-Baptiste, D.; LePrell, R.; Bolton, B. Cyanobacteria and algae blooms: Review of health and environmental data from the Harmful Algal Bloom-Related Illness Surveillance System (HABISS) 2007–2011. Toxins 2015, 7, 1048–1064. [Google Scholar] [CrossRef] [Green Version]
- Bengis, R.; Govender, D.; Lane, E.; Myburgh, J.; Oberholster, P.; Buss, P.; Prozesky, L.; Keet, D. Eco-epidemiological and pathological features of wildlife mortality events related to cyanobacterial bio-intoxication in the Kruger National Park, South Africa. J. South. Afr. Vet. Assoc. 2016, 87, e1–e9. [Google Scholar] [CrossRef] [Green Version]
- Foss, A.J.; Miles, C.O.; Samdal, I.A.; Lovberg, K.E.; Wilkins, A.L.; Rise, F.; Jaabaek, J.A.H.; McGowan, P.C.; Aubel, M.T. Analysis of free and metabolized microcystins in samples following a bird mortality event. Harmful Algae 2018, 80, 117–129. [Google Scholar] [CrossRef] [PubMed]
- Fitzgerald, S.D.; Poppenga, R.H. Toxicosis due to microcystin hepatotoxins in three Holstein heifers. J. Vet. Diagn. Investig. Off. Publ. Am. Assoc. Vet. Lab. Diagn. Inc 1993, 5, 651–653. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Puschner, B.; Galey, F.D.; Johnson, B.; Dickie, C.W.; Vondy, M.; Francis, T.; Holstege, D.M. Blue-green algae toxicosis in cattle. J. Am. Vet. Med. Assoc. 1998, 213, 1571, 1605–1607. [Google Scholar] [PubMed]
- Backer, L.C.; Landsberg, J.H.; Miller, M.; Keel, K.; Taylor, T.K. Canine cyanotoxin poisonings in the United States (1920s-2012): Review of suspected and confirmed cases from three data sources. Toxins 2013, 5, 1597–1628. [Google Scholar] [CrossRef]
- Foss, A.J.; Aubel, M.T.; Gallagher, B.; Mettee, N.; Miller, A.; Fogelson, S.B. Diagnosing Microcystin Intoxication of Canines: Clinicopathological Indications, Pathological Characteristics, and Analytical Detection in Postmortem and Antemortem Samples. Toxins 2019, 11, 456. [Google Scholar] [CrossRef] [Green Version]
- Peacock, M.B.; Gibble, C.M.; Senn, D.B.; Cloern, J.E.; Kudela, R.M. Blurred lines: Multiple freshwater and marine algal toxins at the land-sea interface of San Francisco Bay, California. Harmful Algae 2018, 73, 138–147. [Google Scholar] [CrossRef] [PubMed]
- Miller, M.A.; Kudela, R.M.; Mekebri, A.; Crane, D.; Oates, S.C.; Tinker, M.T.; Staedler, M.; Miller, W.A.; Toy-Choutka, S.; Dominik, C.; et al. Evidence for a novel marine harmful algal bloom: Cyanotoxin (microcystin) transfer from land to sea otters. PLoS ONE 2010, 5. [Google Scholar] [CrossRef] [PubMed]
- Jochimsen, E.M.; Carmichael, W.W.; An, J.S.; Cardo, D.M.; Cookson, S.T.; Holmes, C.E.; Antunes, M.B.; de Melo Filho, D.A.; Lyra, T.M.; Barreto, V.S.; et al. Liver failure and death after exposure to microcystins at a hemodialysis center in Brazil. N. Engl. J. Med. 1998, 338, 873–878. [Google Scholar] [CrossRef] [PubMed]
- Hilborn, E.D.; Soares, R.M.; Servaites, J.C.; Delgado, A.G.; Magalhaes, V.F.; Carmichael, W.W.; Azevedo, S.M. Sublethal microcystin exposure and biochemical outcomes among hemodialysis patients. PLoS ONE 2013, 8, e69518. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Svircev, Z.; Drobac, D.; Tokodi, N.; Mijovic, B.; Codd, G.A.; Meriluoto, J. Toxicology of microcystins with reference to cases of human intoxications and epidemiological investigations of exposures to cyanobacteria and cyanotoxins. Arch. Toxicol. 2017, 91, 621–650. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Chen, J.A.; Zhao, Q.; Pu, C.; Qiu, Z.; Zhang, R.; Shu, W. A cross-sectional investigation of chronic exposure to microcystin in relationship to childhood liver damage in the Three Gorges Reservoir Region, China. Environ. Health Perspect. 2011, 119, 1483–1488. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McCarty, C.L.; Nelson, L.; Eitniear, S.; Zgodzinski, E.; Zabala, A.; Billing, L.; DiOrio, M. Community Needs Assessment After Microcystin Toxin Contamination of a Municipal Water Supply-Lucas County, Ohio, September 2014. Mmwr. Morb. Mortal. Wkly. Rep. 2016, 65, 925–929. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wilson, A.E.; Gossiaux, D.C.; Hook, T.O.; Berry, J.P.; Landrum, P.F.; Dyble, J.; Guildford, S.J. Evaluation of the human health threat associated with the hepatotoxin microcystin in the muscle and liver tissues of yellow perch (Perca flavescens). Can. J. Fish. Aquat. Sci. 2008, 65, 1487–1497. [Google Scholar] [CrossRef] [Green Version]
- Rinehart, K.L.; Harada, K.; Namikoshi, M.; Chen, C.; Harvis, C.A.; Munro, M.H.G.; Blunt, J.W.; Mulligan, P.E.; Beasley, V.R.; Dahlem, A.M.; et al. Nodularin, Microcystin, and the Configuration of Adda. J. Am. Chem. Soc. 1988, 110, 8557–8558. [Google Scholar] [CrossRef]
- Sivonen, K.; Jones, G. Cyanobacterial Toxins. In Toxic Cyanobacteria in Water: A Guide to Their Public Health Consequences, Monitoring and Management; Chorus, I., Bartram, J., Eds.; E and FN Spon: London, UK, 1999; pp. 41–111. [Google Scholar]
- Puddick, J.; Prinsep, M.R.; Wood, S.A.; Kaufononga, S.A.; Cary, S.C.; Hamilton, D.P. High levels of structural diversity observed in microcystins from Microcystis CAWBG11 and characterization of six new microcystin congeners. Mar. Drugs 2014, 12, 5372–5395. [Google Scholar] [CrossRef]
- Spoof, L.; Catherine, L. Appendix 3: Tables of microcystins and nodularins. In Handbook of Cyanobacterial Monitoring and Toxin Analysis; Meriluoto, J., Spoof, L., Codd, G.A., Eds.; John Wiley & Sons: Hoboken, NJ, USA, 2017; pp. 526–537. [Google Scholar]
- Diez-Quijada, L.; Prieto, A.I.; Guzman-Guillen, R.; Jos, A.; Camean, A.M. Occurrence and toxicity of microcystin congeners other than MC-LR and MC-RR: A review. Food Chem. Toxicol. Int. J. Publ. Br. Ind. Biol. Res. Assoc. 2019, 125, 106–132. [Google Scholar] [CrossRef]
- Kaebernick, M.; Neilan, B.A. Ecological and molecular investigations of cyanotoxin production. Fems Microbiol. Ecol. 2001, 35, 1–9. [Google Scholar] [CrossRef]
- Keil, C.; Forchert, A.; Fastner, J.; Szewzyk, U.; Rotard, W.; Chorus, I.; Kratke, R. Toxicity and microcystin content of extracts from a Planktothrix bloom and two laboratory strains. Water Res. 2002, 36, 2133–2139. [Google Scholar] [CrossRef]
- Catherine, A. Microcystins and nodularins. In Handbook of Cyanobacterial Monitoring and Cyantoxin Analysis; Meriluoto, J., Spoof, L., Codd, G.A., Eds.; John Wiley & Sons: Hoboken, NJ, USA, 2017; pp. 109–126. [Google Scholar]
- Graham, J.L.; Loftin, K.A.; Meyer, M.T.; Ziegler, A.C. Cyanotoxin mixtures and taste-and-odor compounds in cyanobacterial blooms from the Midwestern United States. Environ. Sci. Technol. 2010, 44, 7361–7368. [Google Scholar] [CrossRef]
- Zurawell, R.W.; Chen, H.; Burke, J.M.; Prepas, E.E. Hepatotoxic cyanobacteria: A review of the biological importance of microcystins in freshwater environments. J. Toxicol. Environ. Health. Part. B Crit. Rev. 2005, 8, 1–37. [Google Scholar] [CrossRef] [PubMed]
- Stoner, R.D.; Adams, W.H.; Slatkin, D.N.; Siegelman, H.W. The effects of single L-amino acid substitutions on the lethal potencies of the microcystins. Toxicon Off. J. Int. Soc. Toxinology 1989, 27, 825–828. [Google Scholar] [CrossRef]
- Eriksson, J.E.; Gronberg, L.; Nygard, S.; Slotte, J.P.; Meriluoto, J.A. Hepatocellular uptake of 3H-dihydromicrocystin-LR, a cyclic peptide toxin. Biochim. Et. Biophys. Acta 1990, 1025, 60–66. [Google Scholar] [CrossRef]
- Hagenbuch, B.; Meier, P.J. Organic anion transporting polypeptides of the OATP/ SLC21 family: Phylogenetic classification as OATP/ SLCO superfamily, new nomenclature and molecular/functional properties. Pflug. Arch. Eur. J. Physiol. 2004, 447, 653–665. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, H.; Choudhuri, S.; Ogura, K.; Csanaky, I.L.; Lei, X.; Cheng, X.; Song, P.Z.; Klaassen, C.D. Characterization of organic anion transporting polypeptide 1b2-null mice: Essential role in hepatic uptake/toxicity of phalloidin and microcystin-LR. Toxicol. Sci. Off. J. Soc. Toxicol. 2008, 103, 35–45. [Google Scholar] [CrossRef] [Green Version]
- Fischer, A.; Hoeger, S.J.; Stemmer, K.; Feurstein, D.J.; Knobeloch, D.; Nussler, A.; Dietrich, D.R. The role of organic anion transporting polypeptides (OATPs/SLCOs) in the toxicity of different microcystin congeners in vitro: A comparison of primary human hepatocytes and OATP-transfected HEK293 cells. Toxicol. Appl. Pharmacol. 2010, 245, 9–20. [Google Scholar] [CrossRef] [Green Version]
- Weng, D.; Lu, Y.; Wei, Y.; Liu, Y.; Shen, P. The role of ROS in microcystin-LR-induced hepatocyte apoptosis and liver injury in mice. Toxicology 2007, 232, 15–23. [Google Scholar] [CrossRef]
- Chen, T.; Wang, Q.; Cui, J.; Yang, W.; Shi, Q.; Hua, Z.; Ji, J.; Shen, P. Induction of apoptosis in mouse liver by microcystin-LR: A combined transcriptomic, proteomic, and simulation strategy. Mol. Cell. Proteom. Mcp 2005, 4, 958–974. [Google Scholar] [CrossRef] [Green Version]
- Runnegar, M.; Berndt, N.; Kaplowitz, N. Microcystin uptake and inhibition of protein phosphatases: Effects of chemoprotectants and self-inhibition in relation to known hepatic transporters. Toxicol. Appl. Pharmacol. 1995, 134, 264–272. [Google Scholar] [CrossRef] [PubMed]
- Mattos, L.J.; Valenca, S.S.; Azevedo, S.M.; Soares, R.M. Dualistic evolution of liver damage in mice triggered by a single sublethal exposure to Microcystin-LR. Toxicon Off. J. Int. Soc. Toxinol. 2014, 83, 43–51. [Google Scholar] [CrossRef] [PubMed]
- Zhou, M.; Tu, W.W.; Xu, J. Mechanisms of microcystin-LR-induced cytoskeletal disruption in animal cells. Toxicon Off. J. Int. Soc. Toxinol. 2015, 101, 92–100. [Google Scholar] [CrossRef]
- Lovell, R.A.; Schaeffer, D.J.; Hooser, S.B.; Haschek, W.M.; Dahlem, A.M.; Carmichael, W.W.; Beasley, V.R. Toxicity of intraperitoneal doses of microcystin-LR in two strains of male mice. J. Environ. Pathol. Toxicol. Oncol. Off. Organ. Int. Soc. Environ. Toxicol. Cancer 1989, 9, 221–237. [Google Scholar]
- Chernoff, N.; Hunter, E.S., 3rd; Hall, L.L.; Rosen, M.B.; Brownie, C.F.; Malarkey, D.; Marr, M.; Herkovits, J. Lack of teratogenicity of microcystin-LR in the mouse and toad. J. Appl. Toxicol. Jat 2002, 22, 13–17. [Google Scholar] [CrossRef] [PubMed]
- Yoshida, T.; Makita, Y.; Nagata, S.; Tsutsumi, T.; Yoshida, F.; Sekijima, M.; Tamura, S.; Ueno, Y. Acute oral toxicity of microcystin-LR, a cyanobacterial hepatotoxin, in mice. Nat. Toxins 1997, 5, 91–95. [Google Scholar] [CrossRef] [PubMed]
- Fawell, J.K.; Mitchell, R.E.; Everett, D.J.; Hill, R.E. The toxicity of cyanobacterial toxins in the mouse: I microcystin-LR. Hum. Exp. Toxicol. 1999, 18, 162–167. [Google Scholar] [CrossRef]
- Heinze, R. Toxicity of the cyanobacterial toxin microcystin-LR to rats after 28 days intake with the drinking water. Environ. Toxicol. 1999, 14, 57–60. [Google Scholar] [CrossRef]
- Sedan, D.; Laguens, M.; Copparoni, G.; Aranda, J.O.; Giannuzzi, L.; Marra, C.A.; Andrinolo, D. Hepatic and intestine alterations in mice after prolonged exposure to low oral doses of Microcystin-LR. Toxicon Off. J. Int. Soc. Toxinol. 2015, 104, 26–33. [Google Scholar] [CrossRef] [PubMed]
- Huang, P.; Zheng, Q.; Xu, L.H. The apoptotic effect of oral administration of microcystin-RR on mice liver. Environ. Toxicol. 2011, 26, 443–452. [Google Scholar] [CrossRef]
- Liu, X.; Zhang, H.; Liang, J. Blood urea nitrogen is elevated in patients with non-alcoholic fatty liver disease. Hepato-Gastroenterology 2013, 60, 343–345. [Google Scholar]
- He, J.; Li, G.; Chen, J.; Lin, J.; Zeng, C.; Chen, J.; Deng, J.; Xie, P. Prolonged exposure to low-dose microcystin induces nonalcoholic steatohepatitis in mice: A systems toxicology study. Arch. Toxicol. 2017, 91, 465–480. [Google Scholar] [CrossRef] [PubMed]
- Gaudin, J.; Huet, S.; Jarry, G.; Fessard, V. In vivo DNA damage induced by the cyanotoxin microcystin-LR: Comparison of intra-peritoneal and oral administrations by use of the comet assay. Mutat. Res. 2008, 652, 65–71. [Google Scholar] [CrossRef] [PubMed]
- Nishiwaki, R.; Ohta, T.; Sueoka, E.; Suganuma, M.; Harada, K.; Watanabe, M.F.; Fujiki, H. Two significant aspects of microcystin-LR: Specific binding and liver specificity. Cancer Lett. 1994, 83, 283–289. [Google Scholar] [CrossRef]
- Ito, E.; Kondo, F.; Harada, K. Hepatic necrosis in aged mice by oral administration of microcystin-LR. Toxicon Off. J. Int. Soc. Toxinol. 1997, 35, 231–239. [Google Scholar] [CrossRef]
- Ito, E.; Kondo, F.; Harada, K. First report on the distribution of orally administered microcystin-LR in mouse tissue using an immunostaining method. Toxicon Off. J. Int. Soc. Toxinol. 2000, 38, 37–48. [Google Scholar] [CrossRef]
- He, J.; Chen, J.; Wu, L.; Li, G.; Xie, P. Metabolic response to oral microcystin-LR exposure in the rat by NMR-based metabonomic study. J. Proteome Res. 2012, 11, 5934–5946. [Google Scholar] [CrossRef] [PubMed]
- Mrdjen, I.; Morse, M.A.; Ruch, R.J.; Knobloch, T.J.; Choudhary, S.; Weghorst, C.M.; Lee, J. Impact of Microcystin-LR on Liver Function Varies by Dose and Sex in Mice. Toxins 2018, 10, 435. [Google Scholar] [CrossRef] [Green Version]
- Feurstein, D.; Holst, K.; Fischer, A.; Dietrich, D.R. Oatp-associated uptake and toxicity of microcystins in primary murine whole brain cells. Toxicol. Appl. Pharmacol. 2009, 234, 247–255. [Google Scholar] [CrossRef] [Green Version]
- Vesterkvist, P.S.; Misiorek, J.O.; Spoof, L.E.; Toivola, D.M.; Meriluoto, J.A. Comparative cellular toxicity of hydrophilic and hydrophobic microcystins on Caco-2 cells. Toxins 2012, 4, 1008–1023. [Google Scholar] [CrossRef] [Green Version]
- Feurstein, D.; Kleinteich, J.; Heussner, A.H.; Stemmer, K.; Dietrich, D.R. Investigation of microcystin congener-dependent uptake into primary murine neurons. Environ. Health Perspect. 2010, 118, 1370–1375. [Google Scholar] [CrossRef]
- Loftin, K.A.; Clark, J.M.; Journey, C.A.; Kolpin, D.W.; Van Metre, P.C.; Carlisle, D.; Bradley, P.M. Spatial and temporal variation in microcystin occurrence in wadeable streams in the southeastern United States. Environ. Toxicol. Chem. 2016, 35, 2281–2287. [Google Scholar] [CrossRef]
- Service, P.H. Policy on the Humane Care and Use of Laboratory Animals; American Association for Laboratory Animal Science: Memphis, TN, USA, 2015. [Google Scholar]
- Wharton, R.E.; Ojeda-Torres, G.; Cunningham, B.; Feyereisen, M.C.; Hill, K.L.; Abbott, N.L.; Seymour, C.; Hill, D.; Lang, J.; Hamelin, E.I.; et al. Quantification of Microcystin-LR in Human Urine by Immunocapture Liquid Chromatography Tandem Mass Spectrometry. Chem. Res. Toxicol. 2018, 31, 898–903. [Google Scholar] [CrossRef] [PubMed]
- SAS. SAS/STAT®, 13.1; Cary, N.C., Ed.; SAS: Cary, NC, USA, 2013. [Google Scholar]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Number of Animals | Moribundity (%) | Weight Change (g.) | Liver wt (g.) | |||||
| Dose | Male | Female | Male | Female | Male | Female | Male | Female |
| 0 | 6 | 6 | 0 | 0 | −1.47 ± 0.28 | −1.72 ± 0.17 | 1.03 ± 0.09 | 0.79 ± 0.06 |
| 3 | 6 | 6 | 0 | 0 | −1.82 ± 0.28 | −1.73 ± 0.17 | 1.05 ± 0.09 | 0.82 ± 0.06 |
| 5 | 6 | 6 | 33 | 0 | −2.62 ± 0.28** | −2.62 ± 0.17*** | 1.30 ± 0.09 | 0.82 ± 0.06 |
| 7 | 6 | 6 | 50 | 17 | −2.18 ± 0.31 | −2.52 ± 0.17** | 1.31 ± 0.10 | 1.01 ± 0.06** |
| 9 | 6 | 9 | 100** | 100*** | −1.34 ± 0.31 | −0.50 ± 0.15*** | 1.69 ± 0.10*** | 1.45 ± 0.05*** |
| Liver/Body wt | ALT (log10(IU/L)) | AST (log10(IU/L)) | GLDH (log10(IU/L)) | |||||
| Dose | Male | Female | Male | Female | Male | Female | Male | Female |
| 0 | 5.33 ± 0.43 | 4.71 ± 0.28 | 1.89 ± 0.28 | 1.76±0.26 | 2.08 ± 0.23 | 2.11 ± 0.24 | 1.55 ± 0.20 | 1.30 ± 0.18 |
| 3 | 5.67 ± 0.43 | 5.09 ± 0.28 | 2.37 ± 0.28 | 2.13±0.29 | 2.39 ± 0.23 | 2.22 ± 0.26 | 1.76 ± 0.20 | 1.51 ± 0.20 |
| 5 | 6.91 ± 0.43* | 5.26 ± 0.28 | 3.19 ± 0.28** | 3.00±0.26** | 2.85 ± 0.23* | 2.82 ± 0.24* | 2.03 ± 0.20 | 2.10 ± 0.18** |
| 7 | 7.21 ± 0.47** | 6.18 ± 0.28*** | 3.76 ± 0.31*** | 3.84±0.26*** | 3.19 ± 0.26** | 3.55 ± 0.24*** | 2.37 ± 0.22 | 2.66 ± 0.18*** |
| 9 | 8.66 ± 0.47*** | 8.31 ± 0.23*** | 3.91 ± 0.35*** | 3.39±0.27*** | 3.34 ± 0.29** | 3.20 ± 0.24** | 2.41 ± 0.25 | 1.82 ± 0.18 |
| Liver Score | BUN (mg/dl) | Creatinine (mg/dl) | BUN/Creatinine | |||||
| Dose | Male | Female | Male | Female | Male | Female | Male | Female |
| 0 | 3.00 | 0.67 | 8.59 ± 0.76 | 6.92 ± 0.87 | 0.36 ± 0.03 | 0.42 ± 0.03 | 23.78 ± 4.17 | 16.61 ± 2.13 |
| 3 | 4.00 | 1.00 | 9.96 ± 0.76 | 7.45 ± 0.96 | 0.43 ± 0.03 | 0.39 ± 0.03 | 23.48 ± 4.17 | 18.96 ± 2.34 |
| 5 | 11.00* | 9.00** | 9.53 ± 0.76 | 8.98 ± 0.87* | 0.37 ± 0.03 | 0.43 ± 0.03 | 26.05 ± 4.17 | 21.12 ± 2.13 |
| 7 | 13.00** | 10.50** | 10.30 ± 0.84 | 11.43 ± 0.87** | 0.36 ± 0.04 | 0.43 ± 0.03 | 28.73 ± 4.59 | 26.41 ± 2.13** |
| 9 | 15.83** | 16.0*** | 9.26 ± 0.95 | 9.43 ± 0.88* | 0.32 ± 0.04 | 0.44 ± 0.03 | 38.17 ± 5.19 | 22.41 ± 2.15* |
| Albumin (g/dl) | Globulin (g/dl) | Total Protein (g/L) | Glucose (mg/dl) | |||||
| Dose | Male | Female | Male | Female | Male | Female | Male | Female |
| 0 | 2.96 ± 0.19 | 3.30 ± 0.17 | 1.61 ± 0.14 | 1.65 ± 0.09 | 4.72 ± 0.30 | 4.96 ± 0.26 | 176.3 ± 25.8 | 204.4 ± 45.5 |
| 3 | 3.43 ± 0.19 | 3.30 ± 0.19 | 2.07 ± 0.14* | 1.69 ± 0.10 | 5.51 ± 0.30 | 5.00 ± 0.28 | 163.3 ± 25.8 | 115.6 ± 50.0 |
| 5 | 3.09 ± 0.19 | 3.52 ± 0.17 | 1.71 ± 0.14 | 1.78 ± 0.09 | 4.79 ± 0.30 | 5.30 ± 0.26 | 129.8 ± 25.8 | 97.1 ± 45.5 |
| 7 | 2.87 ± 0.20 | 3.28 ± 0.17 | 1.69 ± 0.16 | 1.61 ± 0.09 | 4.57 ± 0.33 | 4.89 ± 0.26 | 75.4 ± 28.4 | 77.1 ± 45.5 |
| 9 | 2.16 ± 0.23* | 2.56 ± 0.17** | 1.25 ± 0.18 | 1.29 ± 0.09* | 3.42 ± 0.37* | 3.85 ± 0.26** | 132.6 ± 32.1 | 215.1 ± 46.0 |
| Total Bilirubin (mg/dl) | ||||||||
| Dose | Male | Female | ||||||
| 0 | 0.17 ± 0.07 | 0.21 ± 0.08 | ||||||
| 3 | 0.20 ± 0.06 | 0.18 ± 0.09 | ||||||
| 5 | 0.37 ± 0.08 | 0.22 ± 0.08 | ||||||
| 7 | 0.38 ± 0.07 | 0.33 ± 0.08 | ||||||
| 9 | 0.25 ± 0.08 | 0.37 ± 0.08 | ||||||
| Variable | Control Male | Female | MCLA Male | MCLR Male | MCLA Female | MCLR Female |
|---|---|---|---|---|---|---|
| Number of animals | 24 | 24 | 8 | 14 | 8 | 15 |
| Moribundity (%) | 0 | 0 | 100 ** | 29 | 63 ** | 7 |
| Weight Change (g.) | −1.68 ± 0.18 | −1.43 ±0.09 | −1.30 ± 0.30 *** | −2.66 ± 0.22 | −2.02 ± 0.16 ** | −2.66 ± 0.11 |
| Liver Wt (g.) | 1.06 ± 0.05 | 0.82 ± 0.03 | 1.89 ± 0.10*** | 1.26 ± 0.07 | 1.26 ± 0.05*** | 0.94 ± 0.04 |
| Liver/Body wt ratio | 5.06 ± 0.23 | 4.75 ± 0.18 | 9.09 ± 0.39*** | 6.58 ± 0.29 | 7.56 ± 0.23 *** | 5.77 ± 0.20 |
| Liver score (avg.) | 2.21 ± 0.44 | 0.17 ± 0.16 | 16 ± 0 * | 12.10 ± 0.97 | 13.75 ± 1.40 ** | 9.20 ± 0.82 |
| ALT (log10 (IU/L)) | 1.94 ± 0.16 | 1.99 ± 0.16 | 4.37 ± 0.35 * | 3.43 ± 0.19 | 4.05 ± 0.27 | 3.50 ± 0.19 |
| AST (log10 (IU/L)) | 2.17 ± 0.11 | 2.27 ± 0.11 | 3.99 ± 0.27*** | 2.97 ± 0.14 | 4.38 ± 0.24*** | 3.17 ± 0.14 |
| GLDH (log10 (IU/L)) | 1.40 ± 0.12 | 1.43 ± 0.13 | 2.71 ± 0.23* | 2.25 ± 0.14 | 1.95 ± 0.23 | 2.38 ± 0.15 |
| BUN (mg/dL) | 8.58 ± 0.66 | 8.37 ± 0.50 | 12.90 ± 1.37 * | 9.90 ± 0.77 | 11.93 ± 0.91 | 10.56 ± 0.62 |
| Creatinine (mg/dL) | 0.45 ± 0.03 | 0.50 ± 0.02 | 0.51 ± 0.04 | 0.45 ± 0.03 | 0.37 ± 0.03** | 0.48 ± 0.03 |
| BUN/Creatinine ratio | 19.39 ± 1.66 | 16.67 ± 0.91 | 24.88 ± 2.53 | 22.24 ± 1.78 | 29.24 ± 1.64** | 22.23 ± 1.12 |
| Albumin (g/dL) | 3.43 ± 0.15 | 3.74 ± 0.19 | 2.72 ± 0.23*** | 3.45 ± 0.16 | 4.39 ± 0.28 | 3.69 ± 0.21 |
| Globulin (g/dL) | 1.90 ± 0.15 | 1.80 ± 0.17 | 1.33 ± 0.20*** | 1.92 ± 0.16 | 1.67 ± 0.19 | 1.65 ± 0.17 |
| Total Protein (g/dL) | 5.37 ± 0.21 | 5.49 ± 0.21 | 4.09 ± 0.33*** | 5.36 ± 0.23 | 6.11 ± 0.33 | 5.31 ± 0.24 |
| Glucose (mg/dL) | 215.3 ± 11.6 | 217.3 ± 11.6 | 95.8 ± 25.2 | 103.6 ± 14.7 | 82.2 ± 17.9 | 97.0 ± 13.4 |
| Total bilirubin (mg/dL) | 0.23 ± 0.09 | 0.24 ± 0.15 | 1.29 ± 0.11*** | 0.38 ± 0.09 | 4.47 ± 0.23*** | 0.27 ± 0.17 |
| [M-H]- | Dosing Solution Vendor | External Calibration Quantification (g/L) | Standard Addition Quantification (g/L) | Reference Material Vendor | |
|---|---|---|---|---|---|
| MCLF | 984.5083 | Enzo Lot #30325 | 0.94 ± 0.12 | 1.3 | Sigma Aldrich Lot #BCBS3614V |
| MCLY | 1000.5032 | Enzo Lot #30326 | 0.91 ± 0.05 | 1.1 | Abraxis Lot#L3032060917 |
| MCRR | 1036.558 | Beagle Lot #MCRR-2004 | 0.74 ± 0.06 | 0.75 | NRC Lot #200702071111 |
| MCLR | 993.541 | Enzo Lot #30362 | 1.0 ± 0.11 | 0.88 | NRC Lot #200701311388 |
| MCWR | 1066.5355 | Enzo Lot #L30471 | 1.1 ± 0.03 | 1.2 | Abraxis Lot # 300652 |
| MCLA | 908.4772 | Beagle Lot #MCLA-2009 | 1.1 ± 0.11 | 1 | Abraxis Lot #B1071017 |
| MCYR | 1043.5052 | Enzo Lot #L30470 | 1.3 ± 0.21 | 1 | Sigma Aldrich Lot #BCBR8280V |
| MCLW | 1023.5192 | Enzo Lot #30392 | 1.1 ± 0.29 | 1 | Sigma Aldrich Lot #BCBS5567V |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chernoff, N.; Hill, D.; Lang, J.; Schmid, J.; Le, T.; Farthing, A.; Huang, H. The Comparative Toxicity of 10 Microcystin Congeners Administered Orally to Mice: Clinical Effects and Organ Toxicity. Toxins 2020, 12, 403. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins12060403
Chernoff N, Hill D, Lang J, Schmid J, Le T, Farthing A, Huang H. The Comparative Toxicity of 10 Microcystin Congeners Administered Orally to Mice: Clinical Effects and Organ Toxicity. Toxins. 2020; 12(6):403. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins12060403
Chicago/Turabian StyleChernoff, Neil, Donna Hill, Johnsie Lang, Judy Schmid, Thao Le, Amy Farthing, and Hwa Huang. 2020. "The Comparative Toxicity of 10 Microcystin Congeners Administered Orally to Mice: Clinical Effects and Organ Toxicity" Toxins 12, no. 6: 403. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins12060403