Detection of Chaetomium globosum, Ch. cochliodes and Ch. rectangulare during the Diversity Tracking of Mycotoxin-Producing Chaetomium-like Isolates Obtained in Buildings in Finland

 , , and
, , and
Abstract
:1. Introduction
2. Results
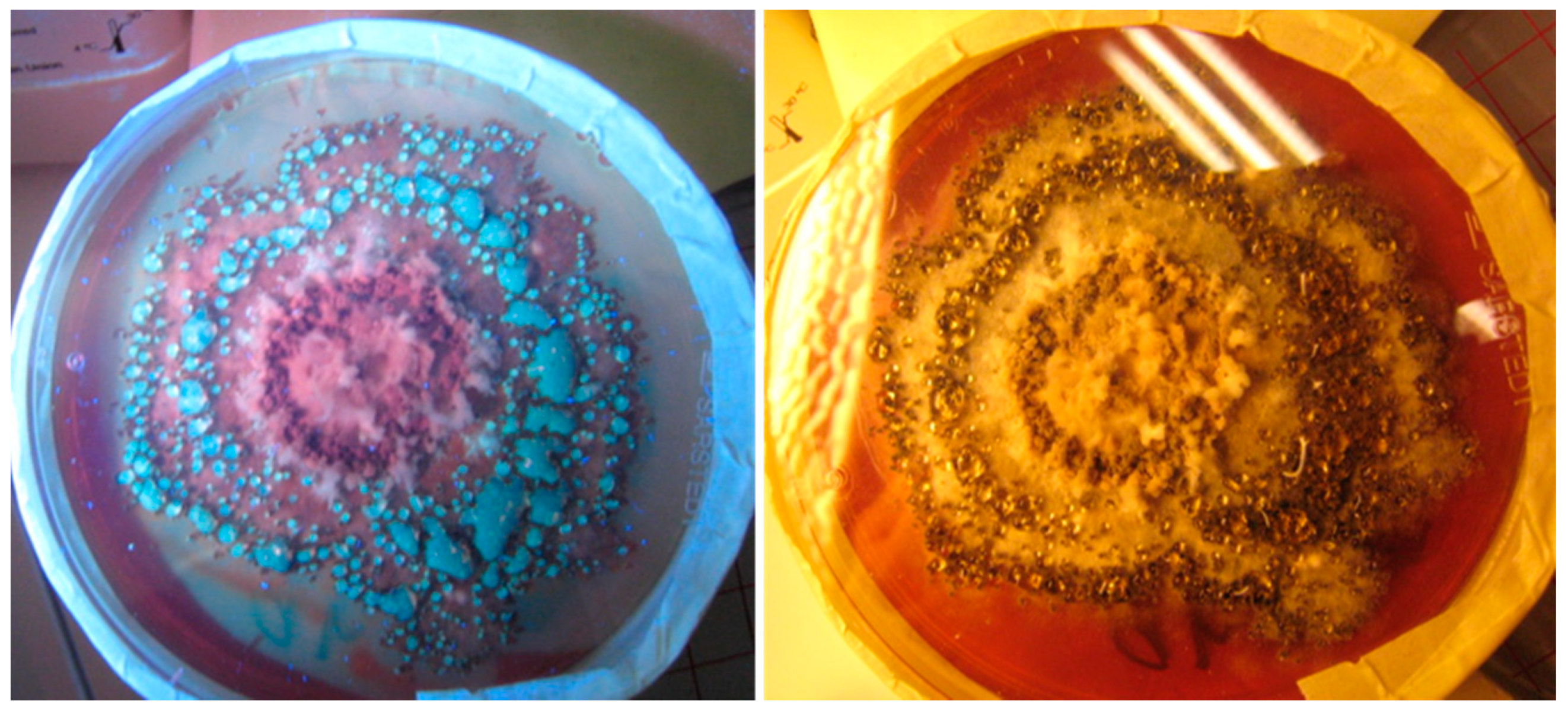
2.1. Screening for Three Toxigenic Indoor Chaetomium-Like Isolates: Ch. globosum, Ch. cochliodes and Ch. rectangulare
2.2. Mycotoxins in the Fungal Extracts
2.3. Mycotoxin and Toxicity Profiles of the Ch. globosum and Ch. cochliodes Isolates
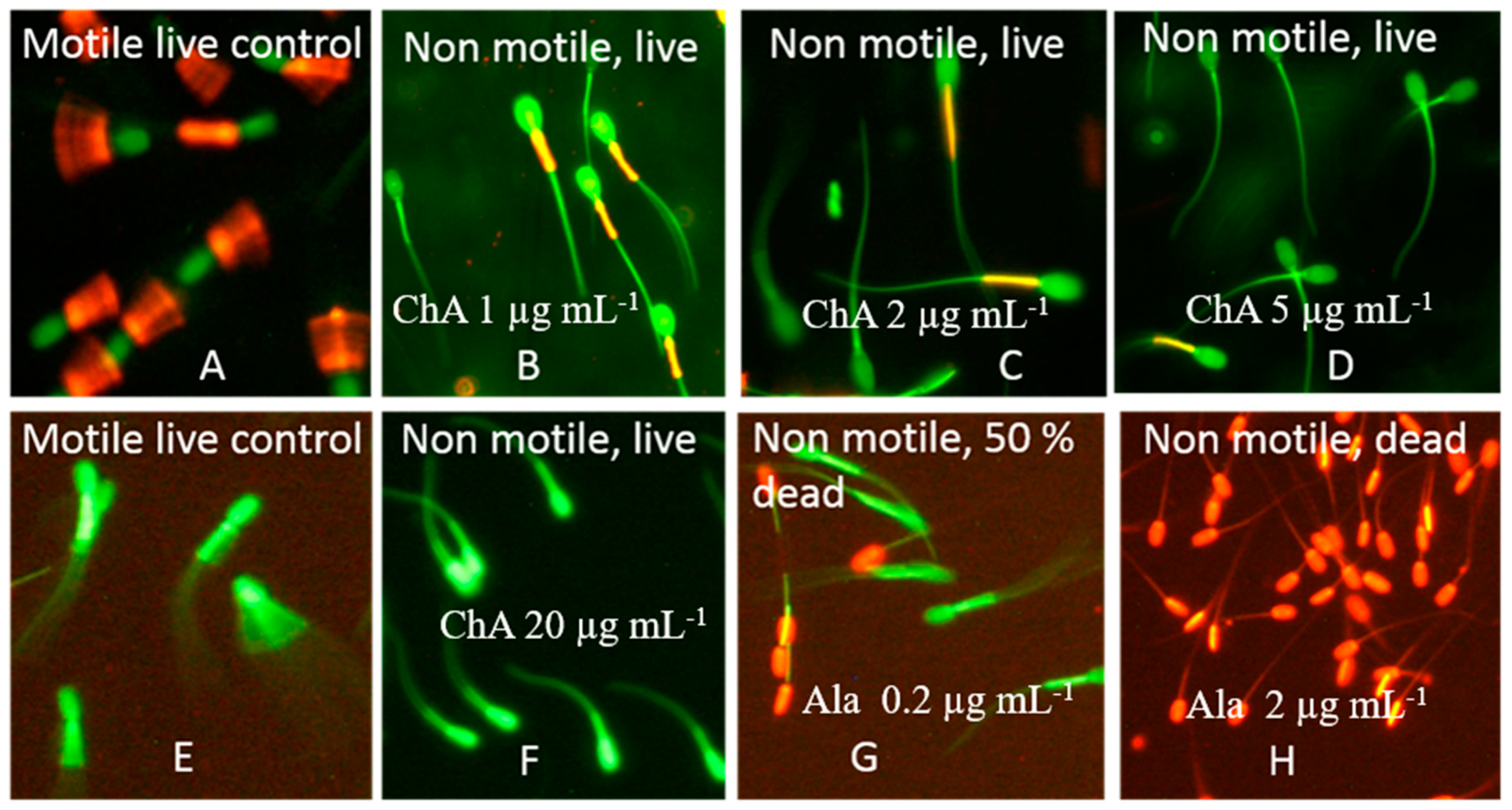
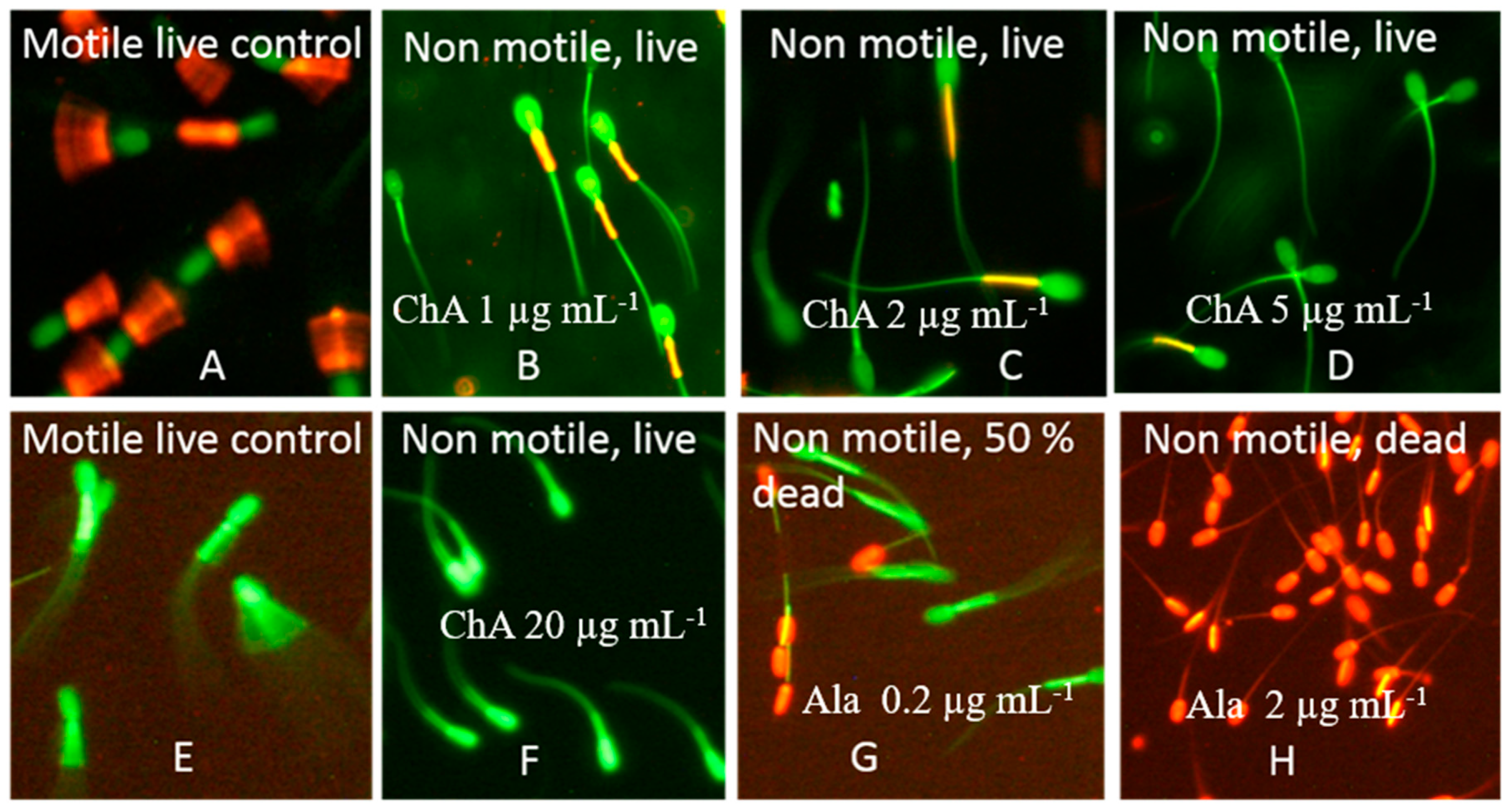
2.4. Characterization of the Toxicological Profile of Pure Chaetoglobosin A and Chaetoglobosin-Containing Extracts
2.5. Resistance of Indoor Chaetomium Strains to Biocides and the Wetting Agent Genapol-X-080
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Experimental Design
5.2. Cultivation of Mold Colonies
5.3. Rapid Screening Tests Using Ex Vivo and In Vitro Assays
5.4. Toxicity Assays for Ethanol-Soluble Dry Substances Extracted from a Plate-Grown Fungal Biomass and Pure Mycotoxins
5.5. Fluorescence Microscopy of Resting Spermatozoa Exposed in the SMIDR and the ΔΨm Assays
5.6. Identification of the Fungal Strains
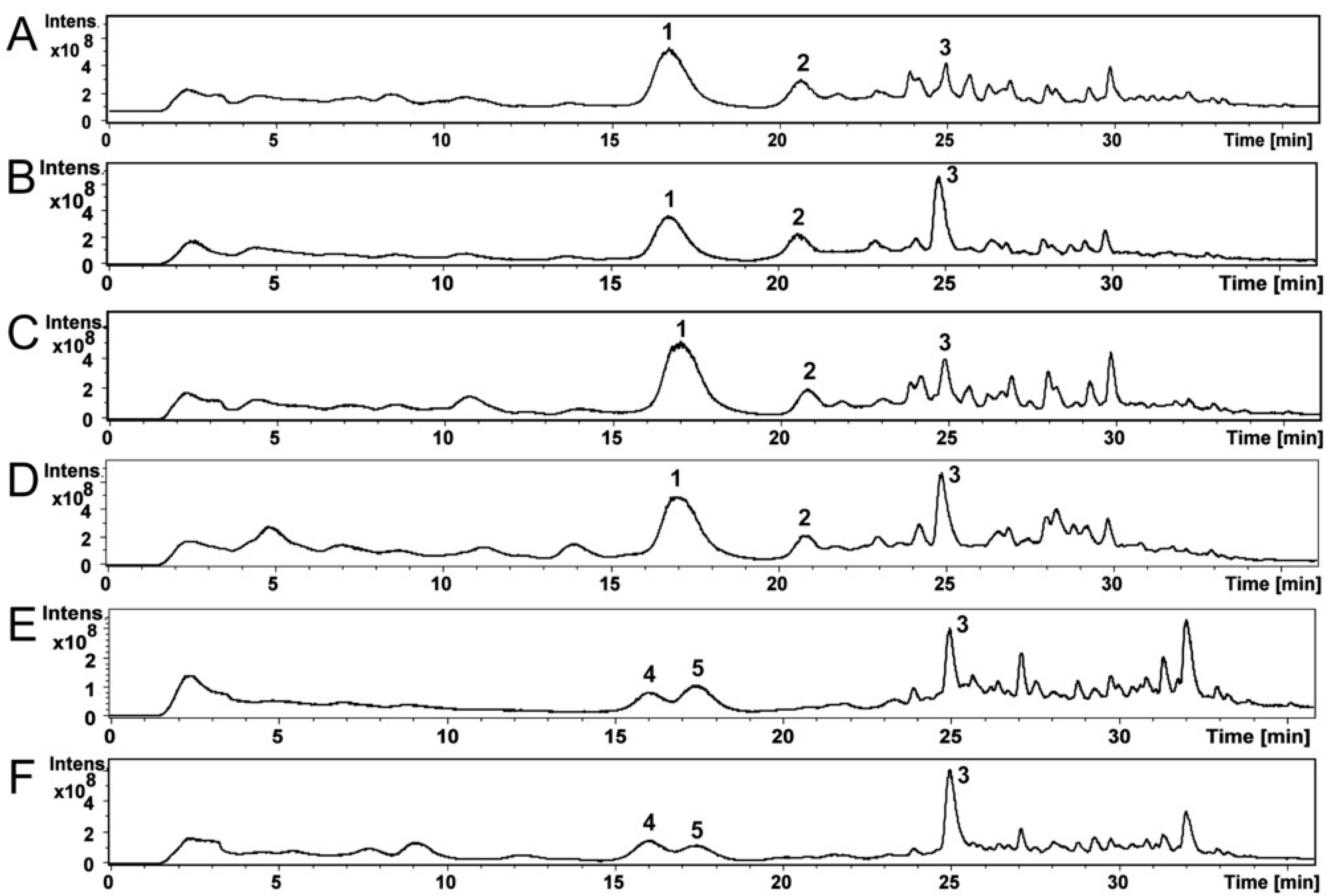
5.7. HPLC-MS Analysis and Identification of Mycotoxins in the Fungal Extracts
5.8. Methods for Testing the Toxicity of Biocides and Genapol-X080 towards the Fungi
5.9. Reagents and Supplies
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Millner, P.D. Radial growth responses to temperature by 58 Chaetomium species, and some taxonomic relationships. Mycologia 1977, 69, 492–502. [Google Scholar] [CrossRef]
- Wang, X.W.; Houbraken, J.; Groenewald, J.Z.; Meijer, M.; Andersen, B.; Nielsen, K.F.; Crous, P.W.; Samson, R.A. Diversity and taxonomy of Chaetomium and Chaetomium-like fungi from indoor environments. Stud. Mycol. 2016, 84, 145–224. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.W.; Lombard, L.; Groenewald, J.Z.; Li, J.; Videira, S.; Samson, R.; Liu, X.; Crous, P.W. Phylogenetic reassessment of the Chaetomium globosum species complex. Persoonia 2016, 36, 83–183. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Andersen, B.; Trane-Niessen, A. Evaluation of media for detection of Stachybotrys and Chaetomium species associated with water-damaged buildings. Int. Biodeterior. Biodegrad. 2000, 46, 111–116. [Google Scholar] [CrossRef]
- Andersen, B.; Frisvad, J.C.; Søndergaard, I.; Rasmussen, S.; Larsen, L. Associations between fungal species and water-damaged building materials. Appl. Environ. Microbiol. 2011, 77, 4180–4188. [Google Scholar] [CrossRef] [Green Version]
- Castagnoli, E.; Andersson, M.A.; Mikkola, R.; Kredics, L.; Marik, T.; Kurnitski, J.; Salonen, H. Indoor Chaetomium-like Isolates: Resistance to Chemicals, Fluorescence and Mycotoxin Production. In Proceedings of the Sisäilmastoseminaari 2017, Helsinki, Finland, 15 March 2017; Säteri, J., Ahola, M., Eds.; Sisäilmayhdistys Ry: Juva, Finland, 2017; Volume 35, pp. 227–232. [Google Scholar]
- Korpi, A.; Pasanen, A.-L.; Viitanen, H. Volatile metabolites of Serpula lacrymans, Coniophora puteana, Poria placenta, Stachybotrys chartarum and Chaetomium globosum. Build. Environ. 1999, 34, 205–211. [Google Scholar] [CrossRef]
- Täubel, M.; Karvonen, A.M.; Reponen, T.; Hyvärinen, A.; Vesper, S.; Pekkanena, J. Application of the environmental relative moldiness index in Finland. Appl. Environ. Microbiol. 2016, 82, 578–584. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Došen, I.; Nielsen, K.F.; Clausen, G.; Andersen, B. Potentially harmful secondary metabolites produced by indoor Chaetomium species on artificially and naturally contaminated building materials. Indoor Air 2017, 1, 34–46. [Google Scholar] [CrossRef] [Green Version]
- Bills, G.F.; Gloer, J.B. Biologically active secondary metabolites from the fungi. Microbiol. Spectr. 2016, 4. [Google Scholar] [CrossRef]
- Gathercole, P.S.; Thiel, P.G.; Hofmeyer, J.H. Inhibition of pyruvate dehydrogenase complex by moniliformin. Biochem. J. 1986, 233, 719–723. [Google Scholar] [CrossRef]
- Andersson, M.A.; Nikulin, M.; Köljalg, U.; Andersson, M.C.; Rainey, F.; Reijula, K.; Hintikka, E.L.; Salkinoja-Salonen, M. Bacteria, molds, and toxins in water-damaged building materials. Appl. Environ. Microbiol. 1997, 63, 387–393. [Google Scholar] [CrossRef] [Green Version]
- Andersson, M.A.; Mikkola, R.; Raulio, M.; Kredics, L.; Maijala, P.; Salkinoja-Salonen, M.S. Acrebol, a novel toxic peptaibol produced by an Acremonium exuviarum indoor isolate. J. Appl. Microbiol. 2009, 106, 909–923. [Google Scholar] [CrossRef] [PubMed]
- Kruglov, A.G.; Andersson, M.A.; Mikkola, R.; Roivainen, M.; Kredics, L.; Saris, N.-E.; Salkinoja-Salonen, M.S. Novel mycotoxin from Acremonium exuviarum is a powerful inhibitor of the mitochondrial respiratory chaincomplex III. Chem. Res. Toxicol. 2009, 9, 565–573. [Google Scholar] [CrossRef]
- Castagnoli, E.; Marik, T.; Mikkola, R.; Kredics, L.; Andersson, M.A.; Salonen, H.; Kurnitski, J. Indoor Trichoderma strains emitting peptaibols in guttation droplets. J. Appl. Microbiol. 2018, 125, 1408–1422. [Google Scholar] [CrossRef] [Green Version]
- Castagnoli, E.; Salo, J.; Toivonen, M.S.; Marik, T.; Mikkola, R.; Kredics, L.; Vicente-Carrillo, A.; Nagy, S.; Andersson, M.T.; Andersson, M.A.; et al. An evaluation of boar spermatozoa as a biosensor for the detection of sublethal and lethal toxicity. Toxins 2018, 10, 463. [Google Scholar] [CrossRef] [Green Version]
- Salo, M.J.; Marik, T.; Mikkola, R.; Andersson, M.A.; Kredics, L.; Salonen, H.; Kurnitski, J. Penicillium expansum strain isolated from indoor building material was able to grow on gypsum board and emitted guttation droplets containing chaetoglobosins and communesins A., B and D. J. Appl. Microbiol. 2019, 127, 1135–1147. [Google Scholar] [CrossRef] [Green Version]
- Salo, M.J.; Marik, T.; Bencsik, O.; Mikkola, R.; Kredics, L.; Szekeres, A.; Andersson, M.A.; Salonen, H.; Kurnitski, J. Screening mold colonies by using two toxicity assays revealed indoor strains of Aspergillus calidoustus producing ophiobolins G and K. Toxins 2019, 11, 683. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Corrier, D. Mycotoxicosis: Mechanisms of immunosuppression. Vet. Immunol. Immunopathol. 1991, 30, 73–87. [Google Scholar] [CrossRef]
- Garbacz, M.; Malec, A.; Duda-Saternus, S.; Suchorab, Z.; Guz, Ł.; Łagód, G. Methods for early detection of microbiological infestation of buildings based on gas sensor technologies. Chemosensors 2020, 8, 7. [Google Scholar] [CrossRef] [Green Version]
- European Agency for Safety and Health at Work. Expert Forecast on Emerging Biological Risks Related to Occupational Safety and Health. European Risk Observatory Report. 2007. Available online: https://osha.europa.eu/en/publications/report-expert-forecast-emerging-biological-risks-relatedoccupational-safety-and-health (accessed on 20 November 2019).
- Liew, W.-P.-P.; Mohd-Redzwan, S. Mycotoxin: Its impact on gut health and microbiota. Front. Cell. Infect. Microbiol. 2018, 8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Piecková, E. In vitro toxicity of indoor Chaetomium Kunze ex Fr. Ann. Agric. Environ. Med. 2003, 10, 9–14. [Google Scholar] [PubMed]
- Piecková, E.; Kunová, Z. Indoor fungi and their ciliostatic metabolites. Ann. Agric. Environ. Med. 2002, 9, 59–63. [Google Scholar]
- Piecková, E.; Wilkins, K. Airway toxicity of house dust and its fungal composition. Ann. Agric. Environ. Med. 2004, 11, 67–73. [Google Scholar]
- Nikulin, M.; Reijula, K.; Jarvis, B.B.; Hintikka, E.L. Experimental lung mycotoxicosis in mice induced by Stachybotrys atra. Int. J. Exp. Pathol. 1996, 77, 213–218. [Google Scholar] [CrossRef] [PubMed]
- Nikulin, M.; Reijula, K.; Jarvis, B.B.; Veijalainen, P.; Hintikka, E.L. Effects of intranasal exposure to spores of Stachybotrys atra in mice. Fundam. Appl. Toxicol. 1997, 35, 182–188. [Google Scholar] [CrossRef] [PubMed]
- Rand, T.G.; Chang, C.T.; McMullin, D.R.; Miller, J.D. Inflammation-associated gene expression in RAW 264.7 macrophages induced by toxins from fungi common on damp building materials. Toxicol. In Vitro 2017, 43, 16–20. [Google Scholar] [CrossRef] [PubMed]
- Fogle, M.R.; Douglas, D.R.; Jumper, C.A.; Straus, D.C. Growth and mycotoxin production by Chaetomium globosum. Mycopathologia 2007, 164, 49–56. [Google Scholar] [CrossRef] [PubMed]
- Caillaud, D.; Leynaert, B.; Keirsbulck, M.; Nadif, R. Indoor mould exposure, asthma and rhinitis: Findings from systematic reviews and recent longitudinal studies—A review. Eur. Respir. Rev. 2018, 27, 170–199. [Google Scholar] [CrossRef] [Green Version]
- Shi, D.; Lu, G.; Mei, H.; de Hoog, G.; Zheng, H.; Liang, G.; Shen, Y.; Li, T.; Liu, W. Onychomycosis due to Chaetomium globosum with yellowish black discoloration and periungual inflammation. Med. Mycol. Case Rep. 2016, 13, 12–16. [Google Scholar] [CrossRef]
- Ahmed, S.; Khan, Z.; Wang, X.; Moussa, T.; Al-Zahrani, H.; Almaghrabi, O.; Sutton, D.A.; Ahmad, S.; Groenewald, J.; Alastruey-Izquierdo, A.; et al. Chaetomium-like fungi causing opportunistic infections in humans: A possible role for extremotolerance. Fungal Divers. 2016, 76, 11–26. [Google Scholar] [CrossRef]
- Lesire, V.; Hazouard, E.; Dequin, F.-E.; Delain, M.; Thérizol-Ferly, M.; Legras, A. Possible role of Chaetomium globosum in infection after autologous bone marrow transplantation. Intensive Care Med. 1999, 25, 124–125. [Google Scholar] [CrossRef]
- Mikkola, R.; Andersson, M.A.; Grigoriev, P.; Heinonen, M.; Salkinoja-Salonen, M.S. The toxic mode of action of cyclic lipodepsipeptide fusaricidins, produced by Paenibacillus polymyxa, toward mammalian cells. J. Appl. Microbiol. 2017, 123, 436–449. [Google Scholar] [CrossRef]
- Xu, G.B.; Li, L.M.; Fang, D.M.; Li, G.Y.; Zhang, G.L.; Wu, Z.J. Electrospray ionization tandem mass spectrometry of chaetoglobosins. Rapid Commun. Mass Spectrom. 2012, 26, 2115–2122. [Google Scholar] [CrossRef] [PubMed]
- Walsh, J.P.; Renaud, J.B.; Hoogstra, S.; McMullin, D.R.; Ibrahim, A.; Visagie, C.M.; Anney, J.B.; Yeung, K.K.; Sumarah, M.W. Diagnostic fragmentation filtering for the discovery of new chaetoglobosins and cytochalasins. Rapid Commun. Mass Spectrom. 2019, 33, 133–139. [Google Scholar] [CrossRef] [PubMed]
- Nielsen, K.F.; Smedsgaard, J. Fungal metabolite screening: Database of 474 mycotoxins and fungal metabolites for dereplication by standardised liquid chromatography-UV-mass spectrometry methodology. J. Chromatogr. A 2003, 100, 111–136. [Google Scholar] [CrossRef]
- Larsen, T.O.; Smedsgaard, J.; Fog Nielsen, K.; Hansen, M.E.; Frisvad, J.C. Phenotypic taxonomy and metabolite profiling in microbial drug discovery. Nat. Prod. Rep. 2005, 22, 672–695. [Google Scholar] [CrossRef] [PubMed]
- McMullin, D.R.; Sumarah, M.W.; Miller, J.D. Chaetoglobosins and azaphilones produced by Canadian strains of Chaetomium globosum isolated from the indoor environment. Mycotoxin Res. 2013, 29, 47–54. [Google Scholar] [CrossRef]
- Wu, Z.J.; Li, G.Y.; Fang, D.M.; Qi, H.Y.; Ren, W.J.; Zhang, G.L. Analysis of epipolythiodioxopiperazines in fungus Chaetomium cochliodes using HPLC-ESI-MS/MS/MS. Anal. Chem. 2008, 80, 217–226. [Google Scholar] [CrossRef] [PubMed]
- Bencsik, O.; Papp, T.; Berta, M.; Zana, A.; Forgó, P.; Dombi, G.; Andersson, M.; Salkinoja-Salonen, M.; Vágvölgyi, C.; Szekeres, A. Ophiobolin A from Bipolaris oryzae perturbs motility and membrane integrities of porcine spermandinduces cell death on mammalian somatic cell lines. Toxins 2014, 6, 2857–2871. [Google Scholar] [CrossRef] [Green Version]
- Nielsen, K.F. Mold Growth on Building Materials. Secondary Metabolites, Mycotoxins and Biomarkers. Ph.D. Thesis, Technical University of Denmark, Lyngby, Denmark, 2002. [Google Scholar]
- Rasimus, S.; Mikkola, R.; Andersson, M.; Teplova, V.; Venediktova, N.; Ek-Kommonen, C.; Salkinoja-Salonen, M. Psychotolerant Paenibacillus tundrae from barley grains produces new cereulide-like depsipeptides, paenilide, and homopaenilide, highly toxic to mammalian cells. Appl. Environ. Microbiol. 2012, 78, 3732–3743. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tonshin, A.A.; Teplova, V.V.; Andersson, M.A.; Salkinoja-Salonen, M.S. The Fusarium mycotoxins enniatins and beauvericin cause mitochondrial dysfunction by affecting the mitochondrial volume regulation, oxidative phosphorylation and ion homeostasis. Toxicology 2010, 276, 49–57. [Google Scholar] [CrossRef] [PubMed]
- Miller, J.D.; McMullin, D.R. Fungal secondary metabolites as harmful air contaminants: 10 years on. Appl. Microbiol. Biothechnol. 2014, 98, 9953–9966. [Google Scholar] [CrossRef] [PubMed]
- Dewangan, J.; Srivastava, S.; Mishra, S.; Pandey, P.; Aman, D.; Rath, S. Chetomin induces apoptosis in human triple-negative breast cancer cells by promoting calcium overload and mitochondrial dysfunction. Biochem. Biophys. Res. Commun. 2018, 495, 1915–1921. [Google Scholar] [CrossRef]
- Viitanen, H.; Vinha, J.; Salminen, K.; Ojanen, T.; Peuhkuri, R.; Paajanen, L.; Lähdesmäki, K. Moisture and bio-deterioration risk of building materials and structures. J. Build. Phys. 2010, 33, 201–224. [Google Scholar] [CrossRef]
- Vicente-Carrillo, A.; Edebert, I.; Garside, H.; Cotgreave, I.; Rigler, R.; Loitto, V.; Magnusson, K.E.; Rodríguez-Martínez, H. Boar spermatozoa successfully predict mitochondrial modes of toxicity: Implications for drug toxicity testing and the 3R principles. Toxicol. In Vitro 2015, 29, 582–591. [Google Scholar] [CrossRef]
- Salo, J.; Andersson, M.A.; Mikkola, R.; Kredics, L.; Viljanen, M.; Salkinoja-Salonen, M. Vapor as a Carrier of Toxicity in a Health Troubled Building. In Proceedings of the Healthy Buildings 2015-Europe (ISIAQ International), Eindhoven, The Netherlands, 18–20 May 2015. [Google Scholar]
- Mikkola, R.; Andersson, M.A.; Grigoriev, P.; Teplova, V.V.; Saris, N.-E.L.; Rainey, F.A.; Salkinoja-Salonen, M.S. Bacillus amyloliquefaciens strains isolated from moisture-damaged buildings produced surfactin and a substance toxic to mammalian cells. Arch. Microbiol. 2004, 181, 314–323. [Google Scholar] [CrossRef] [PubMed]
- Ajao, C.; Andersson, M.A.; Teplova, V.V.; Nagy, S.; Gahmberg, C.G.; Andersson, L.C.; Hautaniemi, M.; Kakasi, B.; Roivainen, M.; Salkinoja-Salonen, M. Mitochondrial toxicity of triclosan on mammalian cells. Toxicol. Rep. 2015, 2, 624–637. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- An, J.; He, H.; Yao, W.; Shang, Y.; Jiang, Y.; Yu, Z. PI3K/Akt/FoxO pathway mediates glycolytic metabolism in HepG2 cells exposed to triclosan (TCS). Environ. Int. 2020, 136, 105428. [Google Scholar] [CrossRef] [PubMed]
- Hoornstra, D.; Andersson, M.A.; Mikkola, R.; Salkinoja-Salonen, M.S. A new method for in vitro detection of microbially produced mitochondrial toxins. Toxicol. In Vitro 2003, 17, 745–751. [Google Scholar] [CrossRef]
- Samson, R.A.; Hoekstra, E.S.; Frisvad, J.C.; Filtenborg, O. (Eds.) Introduction to Food and Air-Borne Fungi, 6th ed.; Centraalbureau voor Schimmelcultures: Utrecht, The Netherlands, 2002. [Google Scholar]
- Hatvani, L.; Antal, Z.; Manczinger, L.; Szekeres, A.; Druzhinina, I.S.; Kubicek, C.P.; Nagy, A.; Nagy, E.; Vágvölgyi, C.; Kredics, L. Green mold diseases of Agaricus and Pleurotus spp. are caused by related but phylogenetically different Trichoderma species. Phytopathology 2007, 4, 532–537. [Google Scholar] [CrossRef] [Green Version]
- Druzhinina, I.S.; Komoń-Zelazowska, M.; Kredics, L.; Hatvani, L.; Antal, Z.; Belayneh, T.; Kubicek, C.P. Alternative reproductive strategies of Hypocrea orientalis and genetically close but clonal Trichoderma longibrachiatum, both capable of causing invasive mycoses of humans. Microbiology 2008, 11, 3447–3459. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain Code | Origin | Sampling Site | Fluorescence/Morphotype | Species | |
|---|---|---|---|---|---|
| MTAV35 *, MTAV37 | Settled dust | University, Oulu | Blue-green | A | Chaetomium globosum |
| ABCD | Settled dust | Apartment Helsinki | Blue-green | A | Ch. globosum |
| MH5 †, M13, M14, M15, M16, L17, L18, MTA1, MTA2, MH52 †, MH10. MH10b HAS5, MH12 | Settled dust | University Espoo | Blue-green | A | Ch. globosum |
| RUK10 †, R7, R8, R9, R11, R20, R21 | Settled dust | Apartment A Vantaa | Blue-green | A | Ch. globosum |
| 2b/26 † 2c/MT | Settled dust | Apartment B Vantaa | Blue-green | A | Ch. globosum |
| 3b/AP | Exhaust air filter | University Espoo | Blue-green | A | Ch. globosum |
| C22/LM †, C23/LM, C21/LM C14/LM, C4/LM | Exhaust air filter | School B Vantaa | Blue-green | A | Ch. globosum |
| MO9 †, MO10, MO11, MO16 | Settled dust | Piggery ‡, Orimattila | Blue-green | A | Ch. globosum |
| OT7 †, OT7b † | Settled dust | Office Helsinki | None | B | Ch. cochliodes |
| Ch1/tu, Ch2/tu, Ch3/tu, Ch4/tu | Inlet air filter | University Espoo | Blue | C | Unidentified Chaetomium-like strains |
| MO15 †, MO13 †, MO12 | Settled dust | Piggery ‡, Orimattila | Yellow-green | D | Ch. rectangulare |
| Strain | Code | EC50 (µg dry wt mL−1) | Identified Metabolite | Estimated Concentration (mg mL−1) | |||
|---|---|---|---|---|---|---|---|
| BSMIM | SMIDM | ICP | |||||
| Boar Sperm | PK-15 | MNA | |||||
| 20 min | 2 h | 2 days | 2 days | ||||
| Biomasses | |||||||
| Group I blue green fluorescent | |||||||
| Ch. globosum | MTAV35 | 5 | 450 | 40 | 20 | chaetoglobosin | 3.4 |
| chaetoviridin A | 0.02 | ||||||
| chaetoviridin C | 0.2 | ||||||
| Ch. globosum | MTAV37 | 10 | 350 | 30 | 15 | No data | |
| Ch. globosum | HAS5 | 5 | 310 | 50 | 25 | chaetoglobosin | 3.9 |
| chaetoviridin A | 0.5 | ||||||
| chaetoviridin C | 0.2 | ||||||
| Ch. globosum | RUK10 | 5 | 300 | 20 | 10 | chaetoglobosin | 4.2 |
| chaetoviridin A | 0.04 | ||||||
| chaetoviridin C | 0.05 | ||||||
| Ch. globosum | ABCD | 5 | 450 | 30 | 15 | chaetoglobosin | 4.24 |
| chaetoviridin A | 0.3 | ||||||
| chaetoviridin C | 0.05 | ||||||
| Group II Non-fluorescent | |||||||
| Ch. cochliodes | OT7 | 10 | 480 | 0.5 | 0.5 | chaetomin | 1.3 |
| chaetoviridin A | 0.13 | ||||||
| chaetomugilin D | 0.02 | ||||||
| OT7b | 10 | 480 | 0.8 | 0.8 | chaetomin | 1.2 | |
| chaetoviridin A | 0.3 | ||||||
| chaetomugilin D | 0.2 | ||||||
| Exudate | 2c/MT | Chaetoglobosin 1 | |||||
| Reference mycotoxin | Biological activity | ||||||
| Alamethicin (Trichoderma arundinaceum) | 5 | 1 | 8 | 8 | Lethal toxin, K+ and Na+ ion channel former [41] | ||
| Chaetoglobosin A (Ch. globosum) | 1 | 12 | 3 | 1 | Sublethal toxin, inhibitor of glucose transport [42] | ||
| Citrinin (Penicillium citrinum) | >100 | 50 | 10 | 10 | Cytostatic toxin, nephrotoxic [41] | ||
| Sterigmatocystin (Aspergillus spp.) | >20 | >20 | 0.5 | Cytostatic toxin, inhibitor of protein synthesis [42] | |||
| Valinomycin (Streptomyces griseus) | 0.0001 | 70 | 14 | Sublethal toxin, mitochondrial toxin, potassium carrier ionophore [43] | |||
| Strain Code | EC50 µg mL−1 | |||||||
|---|---|---|---|---|---|---|---|---|
| Boar Sperm | PK-15 | |||||||
| Lethal Toxicity | Sublethal Toxicity | |||||||
| SMIDR | BSMIR | Depletion of ΔΨm | Acceleration of glycolysis | Acidosis | ||||
| 2 days | 4 days | 2 days | 4 days | 2 days | 4 days | 1 day | 2 days | |
| MTAV35 | >20 | >20 | 3 | 3 | 6 | 3 | None | None |
| MTAV37 | >20 | >20 | 4 | 4 | 4 | 4 | None | None |
| Reference toxins | ||||||||
| Lethal toxin forming ion channels in the plasma membrane | ||||||||
| Alamethicin | 0.2 | 0.2 | 0.2 | None | None | |||
| Sublethal toxin inhibiting glucose transport | ||||||||
| Chaeto-globosin A | >20 | 1 | 2 | None | None | |||
| Sublethal mitochondrial toxins | ||||||||
| Enniatin B | >50 | 5 | 5 | 5 (50) * | 5 | |||
| Moniliformin | ND | 2 | None | 4 (40) * | 4 | |||
| Valinomycin | 5 | 0.00005 | 0.00005 | 0.005 (5) * | 0.005 | |||
| Strain Code | EC50 (µg mL−1) | ||||||
|---|---|---|---|---|---|---|---|
| Borax | Boracol | PHMB | Genapol-X080 | Phenoxy-ethanol | Chloramine | Triclosan | |
| Indoor Ch. globosum strains | |||||||
| MTAV35 | 5000 | 100 | 4 | <50 | 700 | 1200 | 2 |
| MTAV37 | 5000 | 100 | 4 | <50 | 700 | 1200 | 4 |
| HAS5 | >5000 | 100 | 8 | 50 | 1500 | 2500 | 2 |
| 2b/26, | >5000 | 100 | 8 | 50 | 1500 | 2500 | 4 |
| MH 52 | 5000 | 50 | 4 | <50 | 1500 | 1200 | 2 |
| RUK10 | 5000 | 100 | 4 | <50 | 1500 | 2500 | 2 |
| ABCD | 5000 | 100 | 4 | <50 | 1500 | 1200 | 4 |
| MO9 | >5000 | 100 | 4 | <50 | 700 | 2500 | 2 |
| 2c/MT | >5000 | 100 | 8 | 50 | 1500 | 2500 | 4 |
| C22/LM | >5000 | 100 | 8 | 50 | 1500 | 2500 | 4 |
| 3b/AP | >5000 | 100 | 8 | 50 | 1500 | 2500 | 4 |
| Indoor Ch. cochliodes strains | |||||||
| OT7 | 750 | 50 | 8 | >5000 | 3000 | ND | 4 |
| OT7b | 750 | 50 | 8 | >5000 | 3000 | ND | 4 |
| Indoor Ch. rectangulare strains | |||||||
| M015 | 5000 | 100 | 4 | 50,000 | 1500 | 1200 | 2 |
| Mo13 | >5000 | 100 | 4 | 50,000 | 1500 | 1200 | 2 |
| Reference strains | |||||||
| Outdoor Chaetomium-like strains | |||||||
| CH1/tu | 1200 | 100 | 4 | <50 | 1500 | 1200 | 2 |
| Strains that could grow at 37 °C | |||||||
| Indoor strains of Aspergillus section Nigri | |||||||
| Asp21 | ≥5000 | 1600 | 30 | >50,000 | 3000 | 1200 | 30 |
| Asp33b | ≥5000 | 1600 | 30 | >50,000 | 3000 | 1200 | 16 |
| Asp 32 | ≥5000 | 1600 | 60 | >50,000 | 3000 | 1200 | 16 |
| Indoor strains of Aspergillus section Flavi | |||||||
| 7D | 2500 | 200 | 120 | >50,000 | 1500 | 600 | 8 |
| 1/37 | 5000 | 1600 | 120 | >50,000 | 1500 | 600 | 8 |
| Indoor Aspergillus calidoustus strain | |||||||
| MH4 | 5000 | 200 | 8 | >5000 | 3200 | ND | 16 |
| Indoor Paecilomyces variotii strains | |||||||
| Pac1/kop | ≥5000 | 800 | 30 | >50,000 | 3000 | 1200 | 4 |
| Pac2/kop | ≥5000 | 400 | 30 | >50,000 | 3000 | 1200 | 4 |
| Indoor Paecilomyces sp. strains | |||||||
| Pec1/skk | ≥5000 | 800 | 16 | >50,000 | 3000 | 1200 | 4 |
| Pec1/his | ≥5000 | 400 | 30 | >50,000 | 3000 | 1200 | 30 |
| Indoor Trichoderma longibrachiatum strain | |||||||
| THG | 1200 | <50 | 1 | >50,000 | 1500 | 1200 | 16 |
| Indoor Trichoderma citrinoviride strain | |||||||
| SJ40 | 1200 | <50 | 2 | >50,000 | 1500 | 1200 | 8 |
| Strains not able to grow at 37 °C | |||||||
| Aspergillus versicolor strains | |||||||
| SL/3 | 5000 | 100 | 60 | >50,000 | 800 | 600 | 16 |
| Gas/226 | 5000 | 100 | 60 | >50,000 | 400 | 1200 | 16 |
| Aspergillus westerdijkiae strains | |||||||
| PP2 | 5000 | 100 | 60 | >50,000 | 1500 | 600 | 16 |
| PP3 | 5000 | 100 | 60 | >50,000 | 1500 | 600 | 16 |
| AW/KL | 2500 | 100 | 60 | >50,000 | 1500 | 600 | 16 |
| Indoor Trichoderma atroviride strains | |||||||
| H1/226 | 1200 | 100 | 2 | >50,000 | 1500 | 1200 | 16 |
| Ke14 | 1200 | 100 | 4 | >50,000 | 1500 | 1200 | 16 |
| KIV10 | 2500 | 100 | 4 | >50,000 | 3000 | 2400 | 30 |
| Tri335 | 2500 | 100 | 4 | >50,000 | 3000 | 1200 | 30 |
| 14/AM | 1200 | 100 | 4 | >50,000 | 3000 | 1200 | 30 |
| Tri7A/SKK | 1200 | 100 | 8 | >50,000 | 1500 | 2500 | 16 |
| Outdoor T. atroviride strains | |||||||
| SZMC 12541 | 1200 | 100 | 2 | >50,000 | 1500 | 2500 | 16 |
| SZMC 12474 | 2500 | 100 | 4 | >50,000 | 3000 | 2500 | 8 |
| SZMC 207080 | 1200 | 100 | 8 | >50,000 | 3000 | 2500 | 8 |
| SZMC 1723 | 1200 | 100 | 8 | >50,000 | 1500 | 2500 | 16 |
| SZMC 12516 | 2500 | 200 | 30 | >50,000 | 1500 | 2500 | 16 |
| SZMC 12323 | 1200 | 100 | 8 | >50,000 | 1500 | 2500 | 30 |
| Indoor Penicillium sp. strains unable to grow at 37 °C | |||||||
| 35/skk | 5000 | <50 | 30 | >50,000 | 350 | 600 | ND |
| 26/skk | >5000 | <50 | 30 | >50,000 | 3000 | 600 | 30 |
| 5/skk | 2500 | 100 | 8 | >50,000 | 1500 | 600 | 2 |
| HJ2 | 2500 | 100 | 1 | >50,000 | 800 | 1200 | 8 |
| 20b/skk | >5000 | <50 | 30 | >50,000 | 1500 | 600 | 2 |
| Vaip/skk | >5000 | <50 | 8 | >50,000 | 800 | 600 | 4 |
| Mammalian cell lines (ICT assay) | |||||||
| MNA | 150 | <50 | 4 | 25 | 400 | 80 | 4–8 |
| PK-15 | 600 | <50 | 15 | 25 | 1500 | 150 | 8–15 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Salo, J.M.; Kedves, O.; Mikkola, R.; Kredics, L.; Andersson, M.A.; Kurnitski, J.; Salonen, H. Detection of Chaetomium globosum, Ch. cochliodes and Ch. rectangulare during the Diversity Tracking of Mycotoxin-Producing Chaetomium-like Isolates Obtained in Buildings in Finland. Toxins 2020, 12, 443. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins12070443
Salo JM, Kedves O, Mikkola R, Kredics L, Andersson MA, Kurnitski J, Salonen H. Detection of Chaetomium globosum, Ch. cochliodes and Ch. rectangulare during the Diversity Tracking of Mycotoxin-Producing Chaetomium-like Isolates Obtained in Buildings in Finland. Toxins. 2020; 12(7):443. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins12070443
Chicago/Turabian StyleSalo, Johanna M., Orsolya Kedves, Raimo Mikkola, László Kredics, Maria A. Andersson, Jarek Kurnitski, and Heidi Salonen. 2020. "Detection of Chaetomium globosum, Ch. cochliodes and Ch. rectangulare during the Diversity Tracking of Mycotoxin-Producing Chaetomium-like Isolates Obtained in Buildings in Finland" Toxins 12, no. 7: 443. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins12070443