Vaccination with VLPs Presenting a Linear Neutralizing Domain of S. aureus Hla Elicits Protective Immunity

 , ,
, , 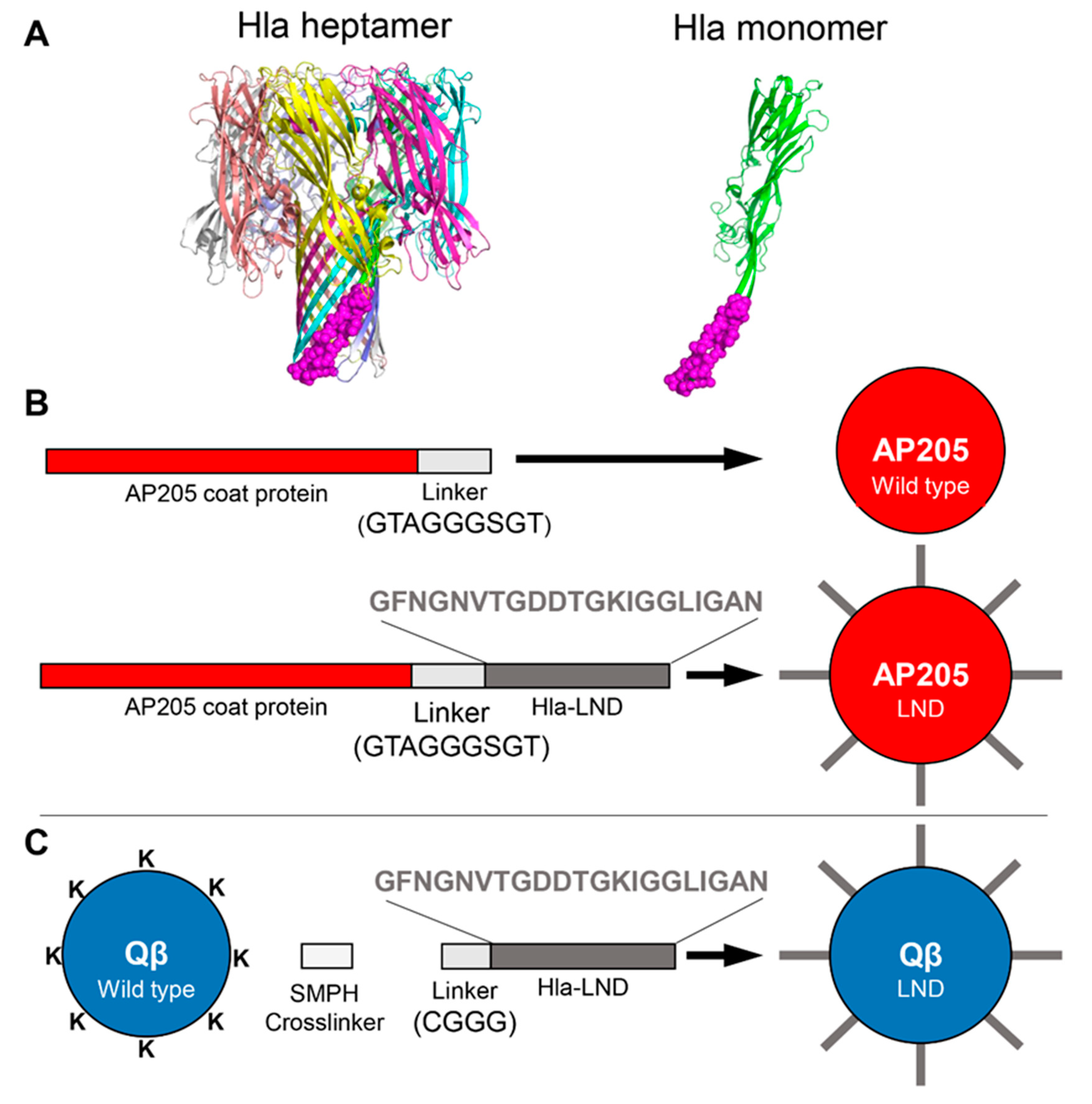{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
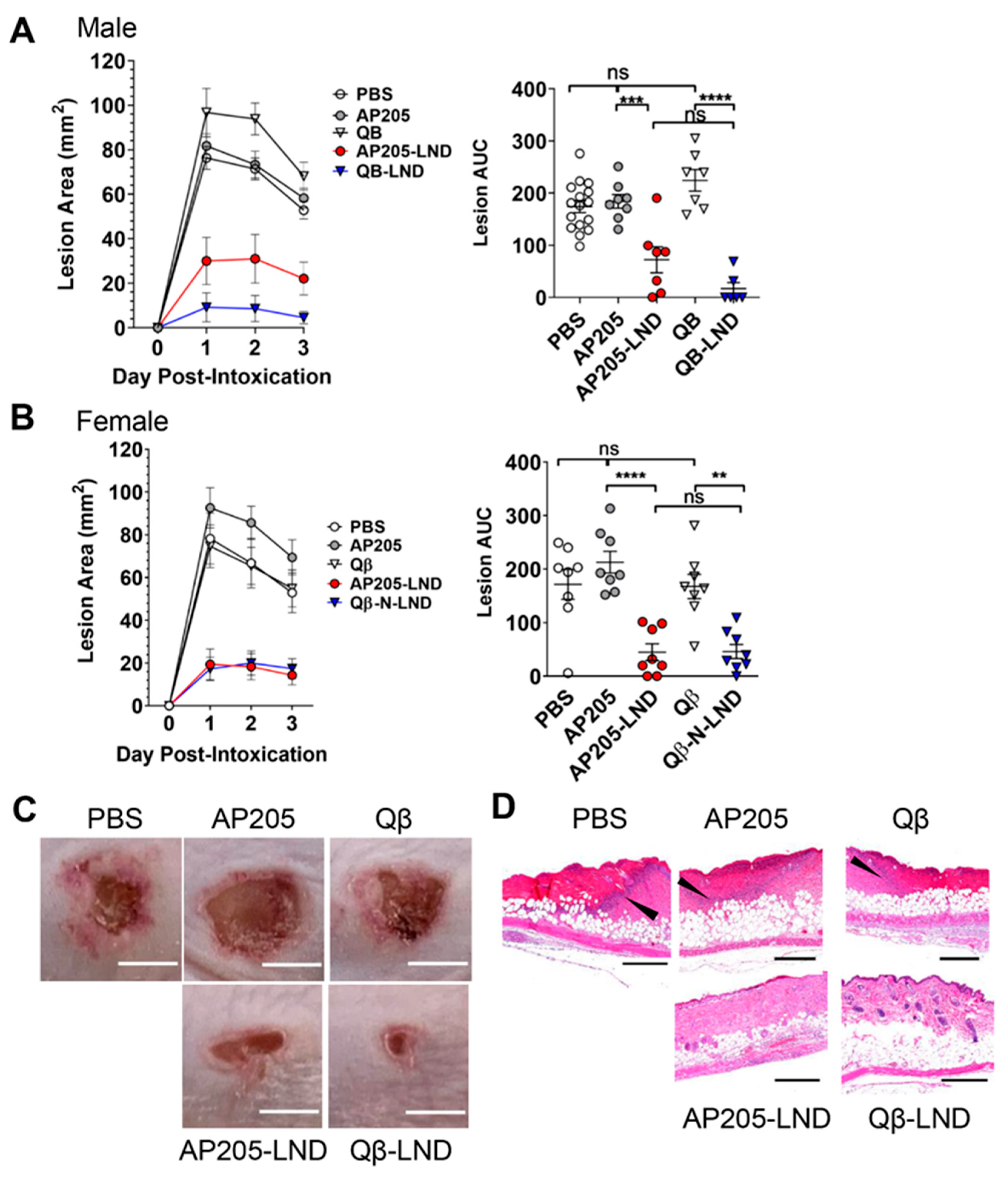
2.1. Vaccination with VLPs Displaying LND Protect against Hla Challenge
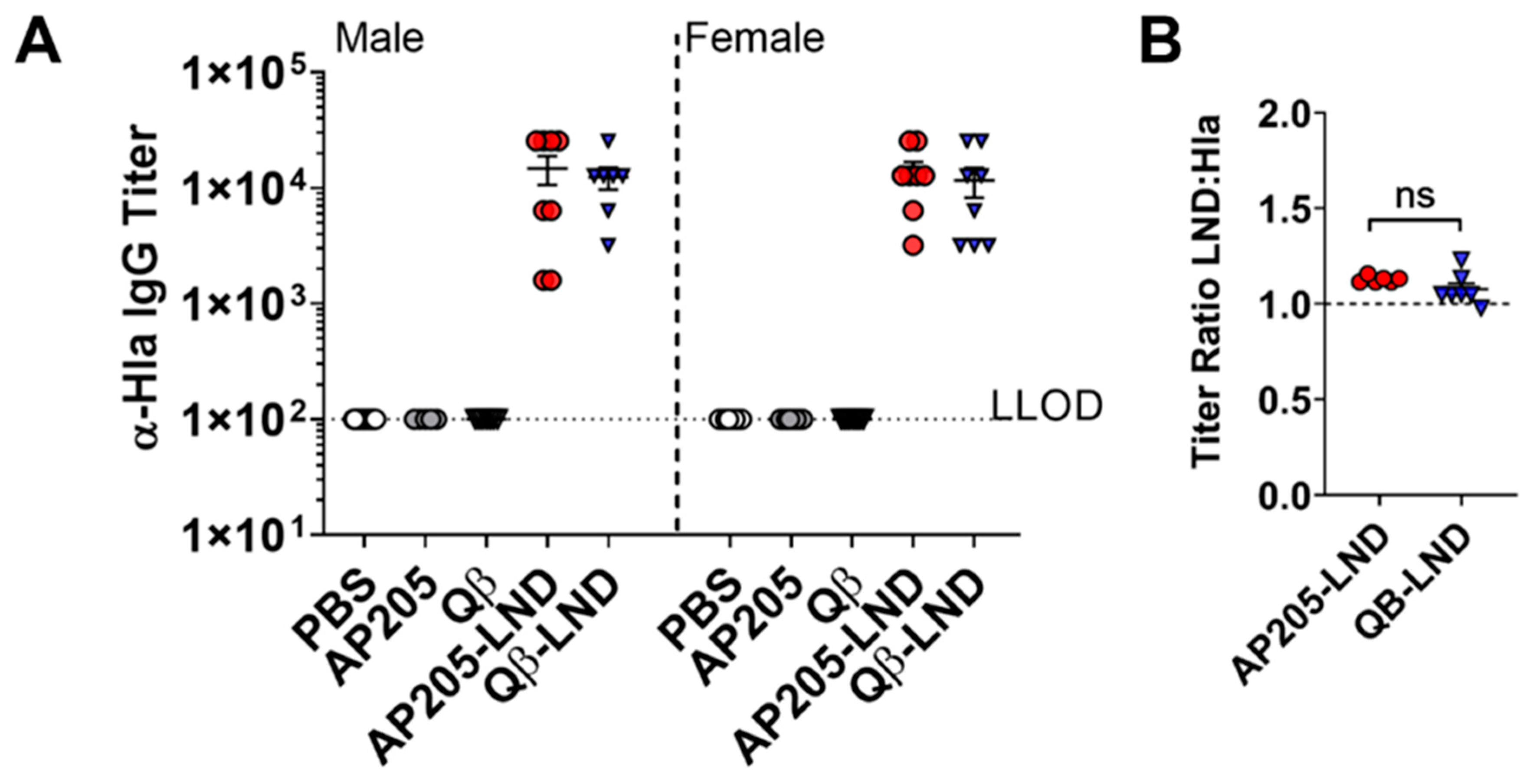
2.2. Antibodies Induced by VLP-LND Vaccination Bind to Intact Hla
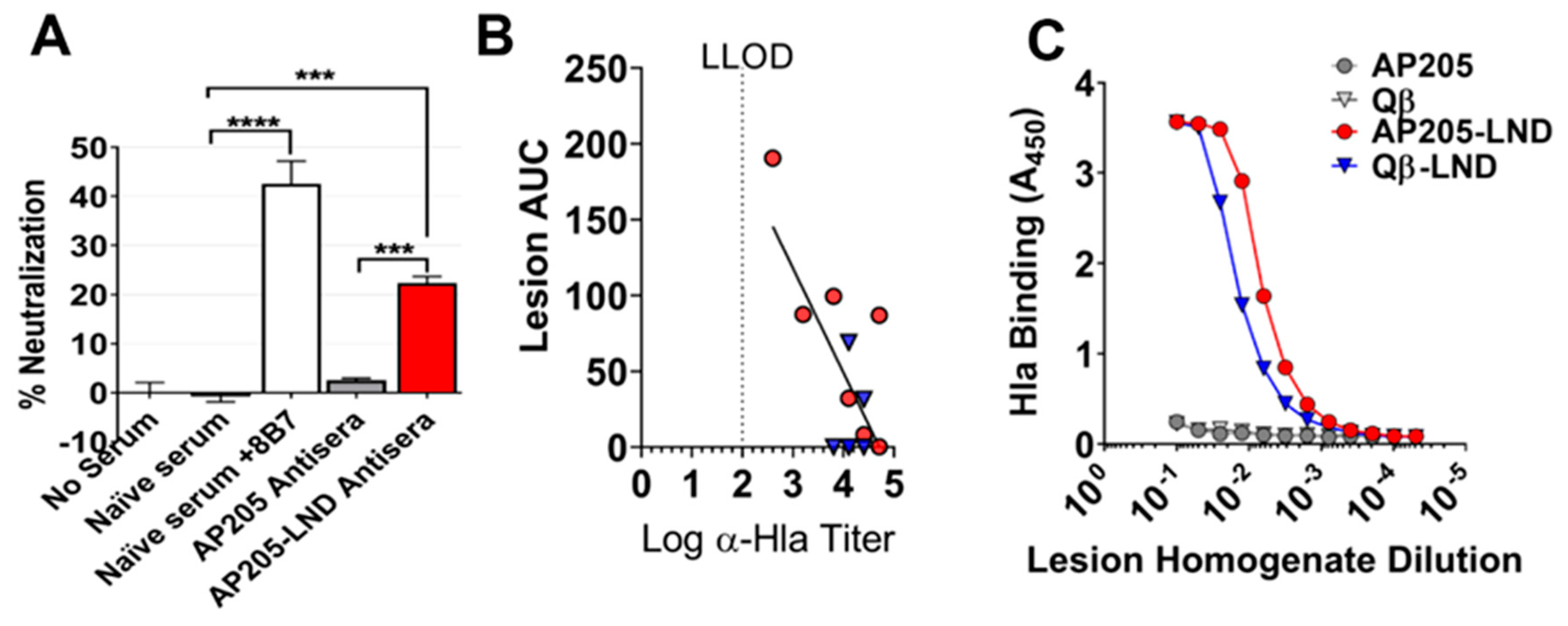
2.3. AP205-LND Vaccination Induces Hla-Neutralizing Antibodies
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Ethics Statement
5.2. VLP Cloning, Expression, and Purification
5.3. Mouse Immunizations
5.4. Hla and LND Peptide ELISA
5.5. Toxin Neutralization Assay
5.6. Mouse Skin Intoxication Model
5.7. Histology
5.8. Statistical analyses
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Rauch, S.; DeDent, A.C.; Kim, H.K.; Bubeck Wardenburg, J.; Missiakas, D.M.; Schneewind, O. Abscess Formation and Alpha-Hemolysin Induced Toxicity in a Mouse Model of Staphylococcus aureus Peritoneal Infection. Infect. Immun. 2012, 80, 3721–3732. [Google Scholar] [CrossRef] [Green Version]
- Bubeck Wardenburg, J.; Bae, T.; Otto, M.; Deleo, F.R.; Schneewind, O. Poring over Pores: Alpha-Hemolysin and Panton-Valentine Leukocidin in Staphylococcus aureus Pneumonia. Nat. Med. 2007, 13, 1405–1406. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, S.D.; Malachowa, N.; Whitney, A.R.; Braughton, K.R.; Gardner, D.J.; Long, D.; Bubeck Wardenburg, J.; Schneewind, O.; Otto, M.; DeLeo, F.R. Comparative Analysis of USA300 Virulence Determinants in a Rabbit Model of Skin and Soft Tissue Infection. J. Infect. Dis. 2011, 204, 937–941. [Google Scholar] [CrossRef]
- Powers, M.E.; Kim, H.K.; Wang, Y.; Bubeck Wardenburg, J. Adam10 Mediates Vascular Injury Induced by Staphylococcus aureus Alpha-Hemolysin. J. Infect. Dis. 2012, 206, 352–356. [Google Scholar] [CrossRef] [PubMed]
- Ragle, B.E.; Bubeck Wardenburg, J. Anti-Alpha-Hemolysin Monoclonal Antibodies Mediate Protection against Staphylococcus aureus Pneumonia. Infect. Immun. 2009, 77, 2712–2718. [Google Scholar] [CrossRef] [Green Version]
- Inoshima, N.; Wang, Y.; Bubeck Wardenburg, J. Genetic Requirement for ADAM10 in Severe Staphylococcus aureus Skin Infection. J. Investig. Dermatol. 2012, 132, 1513–1516. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berube, B.J.; Bubeck Wardenburg, J. Staphylococcus aureus Alpha-Toxin: Nearly a Century of Intrigue. Toxins 2013, 5, 1140–1166. [Google Scholar] [CrossRef] [Green Version]
- Sampedro, G.R.; DeDent, A.C.; Becker, R.E.; Berube, B.J.; Gebhardt, M.J.; Cao, H.; Bubeck Wardenburg, J. Targeting Staphylococcus aureus Alpha-Toxin as a Novel Approach to Reduce Severity of Recurrent Skin and Soft-Tissue Infections. J. Infect. Dis. 2014, 210, 1012–1018. [Google Scholar] [CrossRef] [Green Version]
- Inoshima, I.; Inoshima, N.; Wilke, G.A.; Powers, M.E.; Frank, K.M.; Wang, Y.; Bubeck Wardenburg, J. A Staphylococcus aureus pore-forming toxin subverts the activity of ADAM10 to cause lethal infection in mice. Nat. Med. 2011, 17, 1310–1314. [Google Scholar] [CrossRef] [Green Version]
- Kennedy, A.D.; Bubeck Wardenburg, J.; Gardner, N.J.; Long, D.; Whitney, A.R.; Braughton, K.R.; Schneewind, O.; DeLeo, F.R. Targeting of alpha-hemolysin by active or passive immunization decreases severity of USA300 skin infection in a mouse model. J. Infect. Dis. 2010, 202, 1050–1058. [Google Scholar] [CrossRef] [Green Version]
- Wilke, G.A.; Bubeck Wardenburg, J. Role of a Disintegrin and Metalloprotease 10 in Staphylococcus aureus Alpha-Hemolysin-Mediated Cellular Injury. Proc. Natl. Acad. Sci. USA 2010, 107, 13473–13478. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bubeck Wardenburg, J.; Schneewind, O. Vaccine protection against Staphylococcus aureus pneumonia. J. Exp. Med. 2008, 205, 287–294. [Google Scholar] [CrossRef]
- Adhikari, R.P.; Karauzum, H.; Sarwar, J.; Abaandou, L.; Mahmoudieh, M.; Boroun, A.R.; Vu, H.; Nguyen, T.; Devi, V.S.; Shulenin, S.; et al. Novel Structurally Designed Vaccine for S. aureus α-Hemolysin: Protection against Bacteremia and Pneumonia. PLoS ONE 2012, 7, e38567. [Google Scholar] [CrossRef] [PubMed]
- Fiaschi, L.; Di Palo, B.; Scarselli, M.; Pozzi, C.; Tomaszewski, K.; Galletti, B.; Nardi-Dei, V.; Arcidiacono, L.; Mishra, R.P.N.; Mori, E.; et al. Auto-Assembling Detoxified Staphylococcus aureus Alpha-Hemolysin Mimicking the Wild-Type Cytolytic Toxin. Clin. Vaccine Immunol. 2016, 23, 442–450. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chackerian, B. Virus-like particles: Flexible platforms for vaccine development. Expert Rev. Vaccines 2007, 6, 381–390. [Google Scholar] [CrossRef]
- Mohsen, M.O.; Zha, L.; Cabral-Miranda, G.; Bachmann, M.F. Major findings and recent advances in virus–like particle (VLP)-based vaccines. Semin. Immunol. 2017, 34, 123–132. [Google Scholar] [CrossRef]
- Oscherwitz, J.; Cease, K.B. Identification and Validation of a Linear Protective Neutralizing Epitope in the Beta-Pore Domain of Alpha Toxin. PLoS ONE 2015, 10, e0116882. [Google Scholar] [CrossRef] [PubMed]
- Tissot, A.C.; Renhofa, R.; Schmitz, N.; Cielens, I.; Meijerink, E.; Ose, V.; Jennings, G.T.; Saudan, P.; Pumpens, P.; Bachmann, M.F. Versatile Virus-Like Particle Carrier for Epitope Based Vaccines. PLoS ONE 2010, 5, e9809. [Google Scholar] [CrossRef] [Green Version]
- Malachowa, N.; Kobayashi, S.D.; Braughton, K.R.; DeLeo, F.R. Mouse Model of Staphylococcus aureus Skin Infection. Methods Mol. Biol. 2013, 1031, 109–116. [Google Scholar] [PubMed]
- Ridder, M.J.; Daly, S.M.; Triplett, K.D.; Seawell, N.A.; Hall, P.R.; Bose, J.L. Staphylococcus aureus fatty acid kinase FakA modulates pathogenesis during skin infection via proteases. Infect. Immun. 2020. [Google Scholar] [CrossRef]
- Wright, J.S.; Jin, R.; Novick, R.P. Transient interference with staphylococcal quorum sensing blocks abscess formation. Proc. Natl. Acad. Sci. USA 2005, 102, 1691–1696. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Castleman, M.J.; Febbraio, M.; Hall, P.R. Cd36 Is Essential for Regulation of the Host Innate Response to Staphylococcus aureus Alpha-Toxin-Mediated Dermonecrosis. J. Immunol. 2015, 195, 2294–2302. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Castleman, M.J.; Pokhrel, S.; Triplett, K.D.; Kusewitt, D.F.; Elmore, B.O.; Joyner, J.A.; Femling, J.K.; Sharma, G.; Hathaway, H.J.; Prossnitz, E.R.; et al. Innate Sex Bias of Staphylococcus aureus Skin Infection Is Driven by Alpha-Hemolysin. J. Immunol. 2018, 200, 657–668. [Google Scholar] [CrossRef] [Green Version]
- Hall, P.R. Omega-Hydroxyemodin Limits Staphylococcus aureus Quorum Sensing-Mediated Pathogenesis and Inflammation. J. Immunol. 2015, 59, 2223–2235. [Google Scholar]
- Daly, S.; Joyner, J.A.; Triplett, K.D.; Elmore, B.O.; Pokhrel, S.; Frietze, K.M.; Peabody, D.S.; Chackerian, B.; Hall, P.R. VLP-based vaccine induces immune control of Staphylococcus aureus virulence regulation. Sci. Rep. 2017, 7, 637. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Triplett, K.D.; Pokhrel, S.; Castleman, M.J.; Daly, S.M.; Elmore, B.O.; Joyner, J.A.; Sharma, G.; Herbert, G.; Campen, M.J.; Hathaway, H.J.; et al. GPER activation protects against epithelial barrier disruption by Staphylococcus aureus α-toxin. Sci. Rep. 2019, 9, 1343. [Google Scholar] [CrossRef] [Green Version]
- Kim, H.K.; Kim, H.-Y.; Schneewind, O.; Missiakas, D. Identifying protective antigens of Staphylococcus aureus, a pathogen that suppresses host immune responses. FASEB J. 2011, 25, 3605–3612. [Google Scholar] [CrossRef] [Green Version]
- Atkins, K.L.; Burman, J.D.; Chamberlain, E.S.; Cooper, J.E.; Poutrel, B.; Bagby, S.; Jenkins, A.T.A.; Feil, E.; Elsen, J.M.V.D.; Jenkins, A.T.A.S. aureus IgG-binding proteins SpA and Sbi: Host specificity and mechanisms of immune complex formation. Mol. Immunol. 2008, 45, 1600–1611. [Google Scholar] [CrossRef]
- Sun, Y.; Emolo, C.; Holtfreter, S.; Wiles, S.; Kreiswirth, B.; Missiakas, D.; Schneewind, O. Staphylococcal Protein A Contributes to Persistent Colonization of Mice with Staphylococcus aureus. J. Bacteriol. 2018, 200, e00735-17. [Google Scholar] [CrossRef] [Green Version]
- Miller, L.S.; Fowler, V.G.; Shukla, S.K.; Rose, W.E.; Proctor, R.A. Development of a vaccine against Staphylococcus aureus invasive infections: Evidence based on human immunity, genetics and bacterial evasion mechanisms. FEMS Microbiol. Rev. 2020, 44, 123–153. [Google Scholar] [CrossRef] [Green Version]
- Bouma, G.; Ancliff, P.J.; Thrasher, A.J.; Burns, S. Recent advances in the understanding of genetic defects of neutrophil number and function. Br. J. Haematol. 2010, 151, 312–326. [Google Scholar] [CrossRef]
- Miller, L.S.; Cho, J.S. Immunity against Staphylococcus aureus cutaneous infections. Nat. Rev. Immunol. 2011, 11, 505–518. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Andrews, T.; Sullivan, K.E. Infections in Patients with Inherited Defects in Phagocytic Function. Clin. Microbiol. Rev. 2003, 16, 597–621. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adhikari, R.P.; Ajao, A.O.; Karauzum, H.; Sarwar, J.; Lydecker, A.D.; Aman, M.J.; Johnson, J.K.; Nguyen, C.; Chen, W.H.; Roghmann, M.-C. Lower Antibody Levels to Staphylococcus aureus Exotoxins Are Associated With Sepsis in Hospitalized Adults With Invasive S. aureus Infections. J. Infect. Dis. 2012, 206, 915–923. [Google Scholar] [CrossRef]
- Fritz, S.A.; Tiemann, K.M.; Hogan, P.G.; Epplin, E.K.; Rodriguez, M.; Al-Zubeidi, D.N.; Bubeck Wardenburg, J.; Hunstad, D.A. A Serologic Correlate of Protective Immunity against Community-Onset Staphylococcus aureus Infection. Clin. Infect. Dis. 2013, 56, 1554–1561. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tran, V.G.; Venkatasubramaniam, A.; Adhikari, R.P.; Krishnan, S.; Wang, X.; Le, V.T.M.; Le, H.N.; Vu, T.T.T.; Schneider-Smith, E.; Aman, M.J.; et al. Efficacy of Active Immunization With Attenuated α-Hemolysin and Panton-Valentine Leukocidin in a Rabbit Model of Staphylococcus aureus Necrotizing Pneumonia. J. Infect. Dis. 2019, 221, 267–275. [Google Scholar] [CrossRef] [PubMed]
- Oscherwitz, J.; Muñoz-Planillo, R.; Yu, F.; Nunez, G.; Cease, K.B. In vivo mapping of a protective linear neutralizing epitope at the N-terminus of alpha hemolysin from Staphylococcus aureus. Mol. Immunol. 2014, 60, 62–71. [Google Scholar] [CrossRef]
- Adhikari, R.P.; Thompson, C.D.; Aman, M.J.; Lee, J.C. Protective efficacy of a novel alpha hemolysin subunit vaccine (AT62) against Staphylococcus aureus skin and soft tissue infections. Vaccine 2016, 34, 6402–6407. [Google Scholar] [CrossRef] [Green Version]
- Proctor, R.A. Immunity to Staphylococcus aureus: Implications for Vaccine Development. Microbiol. Spectr. 2019, 7, 766–775. [Google Scholar] [CrossRef]
- National Research Council Committee for the Update of the Guide for the Care and Use of Laboratory Animals. Guide for the Care and Use of Laboratory Animals, 8th ed.; National Academies Press: Washington, DC, USA, 2011. [Google Scholar]
- Crossey, E.; Amar, M.J.A.; Sampson, M.; Peabody, J.; Schiller, J.T.; Chackerian, B.; Remaley, A.T. A cholesterol-lowering VLP vaccine that targets PCSK9. Vaccine 2015, 33, 5747–5755. [Google Scholar] [CrossRef] [Green Version]
- Akache, B.; Weeratna, R.D.; Deora, A.; Thorn, J.M.; Champion, B.; Merson, J.R.; Davis, H.L.; McCluskie, M.J. Anti-IgE Qb-VLP Conjugate Vaccine Self-Adjuvants through Activation of TLR7. Vaccines 2016, 4, 3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chackerian, B.; Rangel, M.; Hunter, Z.; Peabody, D.S. Virus and virus-like particle-based immunogens for Alzheimer’s disease induce antibody responses against amyloid-beta without concomitant T cell responses. Vaccine 2006, 24, 6321–6331. [Google Scholar] [CrossRef] [PubMed]




© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Joyner, J.A.; Daly, S.M.; Peabody, J.; Triplett, K.D.; Pokhrel, S.; Elmore, B.O.; Adebanjo, D.; Peabody, D.S.; Chackerian, B.; Hall, P.R. Vaccination with VLPs Presenting a Linear Neutralizing Domain of S. aureus Hla Elicits Protective Immunity. Toxins 2020, 12, 450. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins12070450
Joyner JA, Daly SM, Peabody J, Triplett KD, Pokhrel S, Elmore BO, Adebanjo D, Peabody DS, Chackerian B, Hall PR. Vaccination with VLPs Presenting a Linear Neutralizing Domain of S. aureus Hla Elicits Protective Immunity. Toxins. 2020; 12(7):450. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins12070450
Chicago/Turabian StyleJoyner, Jason A., Seth M. Daly, Julianne Peabody, Kathleen D. Triplett, Srijana Pokhrel, Bradley O. Elmore, Diane Adebanjo, David S. Peabody, Bryce Chackerian, and Pamela R. Hall. 2020. "Vaccination with VLPs Presenting a Linear Neutralizing Domain of S. aureus Hla Elicits Protective Immunity" Toxins 12, no. 7: 450. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins12070450