Characterization of Bacillus cereus in Dairy Products in China


1
College of Veterinary Medicine, China Agricultural University, Beijing 100193, China
2
Department of Mechanics and Engineering Science, College of Engineering, Academy for Advanced Interdisciplinary Studies, and Beijing Advanced Innovation Center for Engineering Science and Emerging Technology, College of Engineering, Peking University, Beijing 100871, China
3
National Center for Veterinary Drug Safety Evaluation, College of Veterinary Medicine, China Agricultural University, Beijing 100193, China
4
National Feed Drug Reference Laboratory, Feed Research Institute, Chinese Academy of Agricultural Sciences, Beijing 100081, China
*
Author to whom correspondence should be addressed.
†
These authors contributed equally to this review.
Toxins 2020, 12(7), 454; https://0-doi-org.brum.beds.ac.uk/10.3390/toxins12070454
Submission received: 28 May 2020
/
Revised: 30 June 2020
/
Accepted: 9 July 2020
/
Published: 14 July 2020
(This article belongs to the Special Issue Bacillus cereus Toxins)
Abstract
:Bacillus cereus is a common and ubiquitous foodborne pathogen with an increasing prevalence rate in dairy products in China. High and unmet demands for such products, particularly milk, raise the risk of B. cereus associated contamination. The presence of B. cereus and its virulence factors in dairy products may cause food poisoning and other illnesses. Thus, this review first summarizes the epidemiological characteristics and analytical assays of B. cereus from dairy products in China, providing insights into the implementation of intervention strategies. In addition, the recent achievements on the cytotoxicity and mechanisms of B. cereus are also presented to shed light on the therapeutic options for B. cereus associated infections.
Key Contribution: This review summarizes the prevalence of B. cereus from dairy products in China. It also focuses on the analytical methods of B. cereus and gives an overview of the current knowledge about the virulence factors and the underlying mechanisms of infections.
1. Introduction
Bacillus cereus is a Gram-positive, endospore-forming, foodborne pathogenic bacterium that is widely distributed in the natural environments, frequently found in foods especially dairy products and even persisting in host epithelial cells [1,2,3,4,5,6,7]. As an opportunistic pathogen, B. cereus has long-term emerged as a health threat to humans and animals, involving both domestic and wild animals [8,9,10,11]. Foodborne outbreaks involving B. cereus in China usually occurred through dairy products [12,13,14,15]. Major symptoms of food-poisoning caused by B. cereus are divided into either diarrhea or emesis [5,16]. Diarrhea is mainly induced by three enterotoxins that belong to the family of pore-forming toxins (PFTs) [17], including non-hemolytic enterotoxin (Nhe) [18,19], hemolysin BL (Hbl) [20] and cytolysin K (CytK) [21], while the emetic syndrome is tightly connected to a lethal toxin known as “cereulide”, which is synthesized by a non-ribosomal peptide synthetase (NRPS) encoded by a ces gene [22,23].
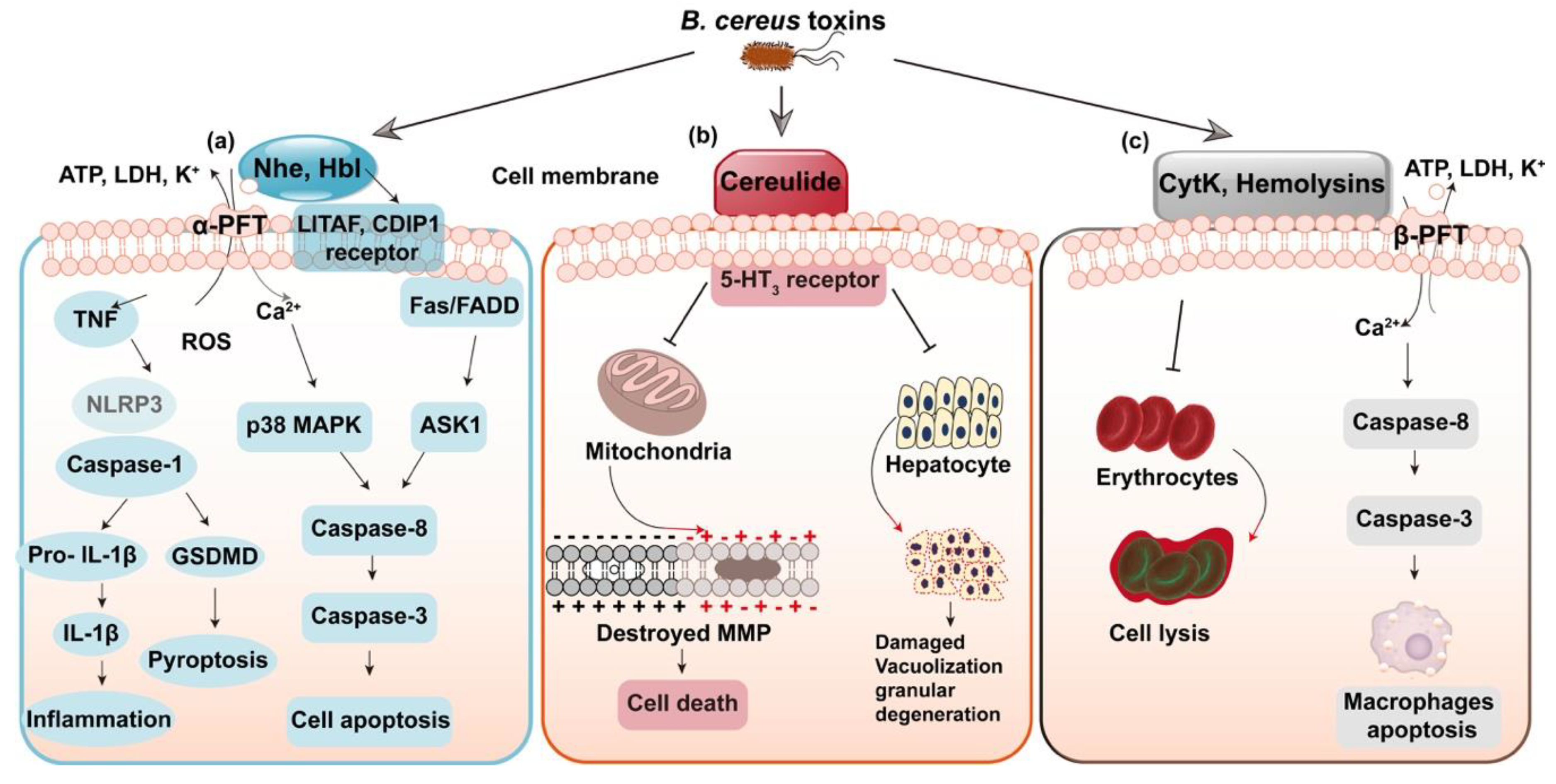
Most toxins of B. cereus belong to the family of pore forming toxins (PFTs); among them, Nhe and Hbl are similar to the well-known cytolysin A (ClyA) of α-PFTs family, while CytK and hemolysins are members of β-PFTs [17,24,25]. PFTs have the capacities of altering the plasma membrane permeability of target cells, potentially leading to cell death and triggering the signaling pathways [26,27,28]. Nhe and Hbl have a similar mechanism of action; the three components of Nhe complex display the highest cytotoxicity at a ratio close to 10:10:1 for NheA, B and C [29,30]. In terms of Hbl, the ratio at L2: L1: B = 1:1:10/ 10:1:10 prompts the most rapid pore formation [20]. Recent studies also found that LITAF and CDIP1 work as the Hbl receptor [31]. The signaling pathways triggered by Nhe, as shown in Figure 1a, include the induced cell apoptosis though ASK1 and Fas-p38 MAPK mediated caspase-8 dependent pathways [32]. In addition, the most recent research suggested that Nhe and Hbl operate synergistically to activate the NLRP3 inflammasome and induce inflammation [33]. Moreover, Nhe have the concerted action with sphingomyelinase in pathogenic B. cereus to cause full virulence and formation of disease [34]. The two caspase-1 dependent inflammatory pathways triggered by Nhe include the form of inflammation initiated by IL-1β release and pyroptosis induced by the activation of GSDMD (Figure 1a). Moreover, the emetic toxin, cereulide is a K+ ionophore toxin that damages the cellular membrane potential through inhibiting the synthesis of RNA [35], affecting mitochondrial function, resulting in expansion of mitochondria and formation of vacuoles in the protoplasm of sensitive cells, inducing cell apoptosis and even fulminant liver failure [36,37] (Figure 1b). Moreover, cereulide is terribly unwholesome and could accumulate in multiple organs [38]. CytK and hemolysins otherwise do harm to the target membrane, bringing cell lysis and apoptosis in macrophages [39] (Figure 1c).
Besides, B. cereus can survive in the gastrointestinal tract with versatile virulence factors [40,41]. Therefore, many other infections associated with B. cereus have been reported including meningitis, brain abscess [42], cellulitis [43], endophthalmitis [44,45], pneumonia [46], endocarditis [47] and osteomyelitis [48]. More seriously, persistent B. cereus strains, which are highly detrimental pathogenic bacteria against antibiotic therapies, were also found in patients in the USA [49]. Altogether, tracing the source of B. cereus, such as in dairy products, is of particular concern.
At present, to the best of our knowledge, there is no detailed information focusing on the potential risk of B. cereus from dairy products in China, not to mention the summaries about the detection and toxicity mechanisms of Bacillus virulence factors. In the current review, we summarized the demand for dairy products in China, the prevalence and detection of B. cereus and the virulence factors. We aim to give an overview of B. cereus in dairy products, which may contribute to the implementation of effective strategies to prevent and control foodborne pathogenic B. cereus in dairy products.
2. Dairy Products with Bacillus cereus
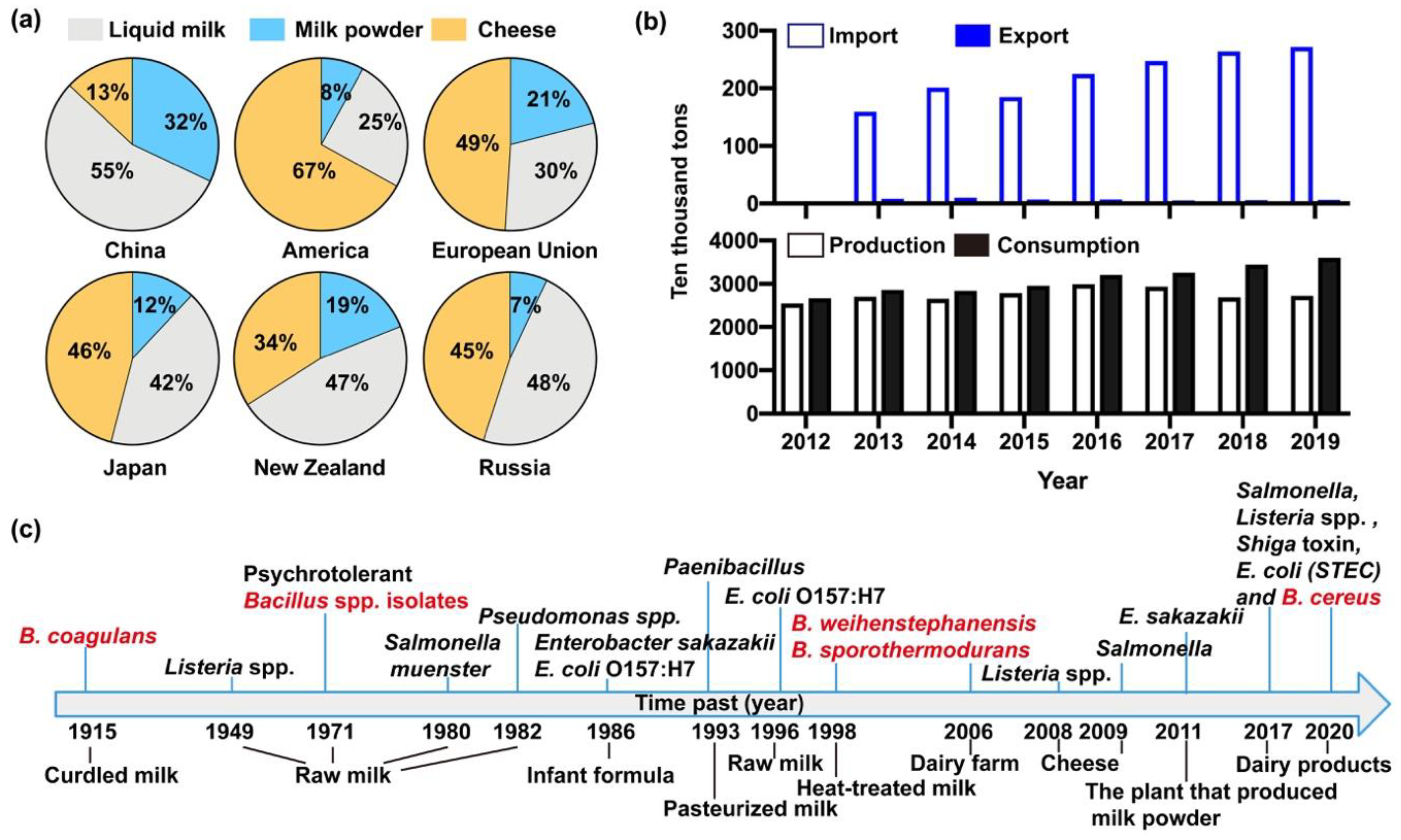
Dairy products are excellent nutrition for both young animals and human beings [50,51,52]. Notably, milk contains all eight kinds of essential amino acids, minerals, vitamins, and fatty acids with optimal proportions of nutrients [8,9]. To satisfy the increasing needs of human, milk associated dairy products are derived from diverse sources, which are known to fall into various categories including liquid milk, milk powder, cheese, condensed milk, milk fat and ice cream [51,52,53]. Demand for dairy products varies sharply from one region to another [54]. For instance, liquid milk had a top priority for Chinese customers in contrast to American and those from other countries, the proportions being 55% in China and 25% in America, respectively, while cheese had the smallest demand (just 13%, yellow part) in China compared with other countries (Figure 2a). In addition, dairy products in the Chinese market have been in short supply, and the export volume of these was far less than the import amount in China from the year of 2013 to 2018 (Figure 2b). According to the phased consumption targets of the National Food and Nutrition Advisory Committee, per capita consumption of milk will reach 28kg by 2020 and 41kg by 2030 [53]. Likewise, the sales of milk in China will increase from 119.5 to 128.3 billion yuan from 2018 to 2022, with an average annual compound growth rate of 1.7%, and the size of the milk market will sustain a steady growth, as put forward by the China Business Research Institute. Consequently, the sale volume of milk in China will gradually augment, and the market prospect is considerable [55,56]. In view of the high demand for dairy products and the outbreaks of bacterial contamination in liquid milk and milk powder in China, it is urgent to carry out effective analytical tests, especially the detection of microbes during manufacturing, selling and importing of milk [12,57,58,59]. In addition, the uncertain microbial growth rate and toxicity of microbial metabolites all threaten food safety [60,61,62,63]. As shown in Figure 2c, although preventive measures have been made to manage the contamination of dairy products in the last century [64], the outbreaks of B. cereus spp. contamination constantly arose in various milk products all over the world [61,62,65,66,67,68]. The prevalence of B. cereus in dairy products is difficult to estimate, and food poisoning incidents caused by B. cereus still remain a thorny problem worldwide due to the high tolerance of B. cereus to various environments and strong propagation capacity of B. cereus spores [2,5,13,47,69]. The B. cereus isolates correspondingly are suspected of threatening the safety of raw milk and dairy products in China [5,13,70]. Therefore, we will next focus on the pattern of B. cereus contamination in dairy products in China.
3. Prevalence of Bacillus cereus from Dairy Products in China
3.1. Contamination of Bacillus cereus Isolates
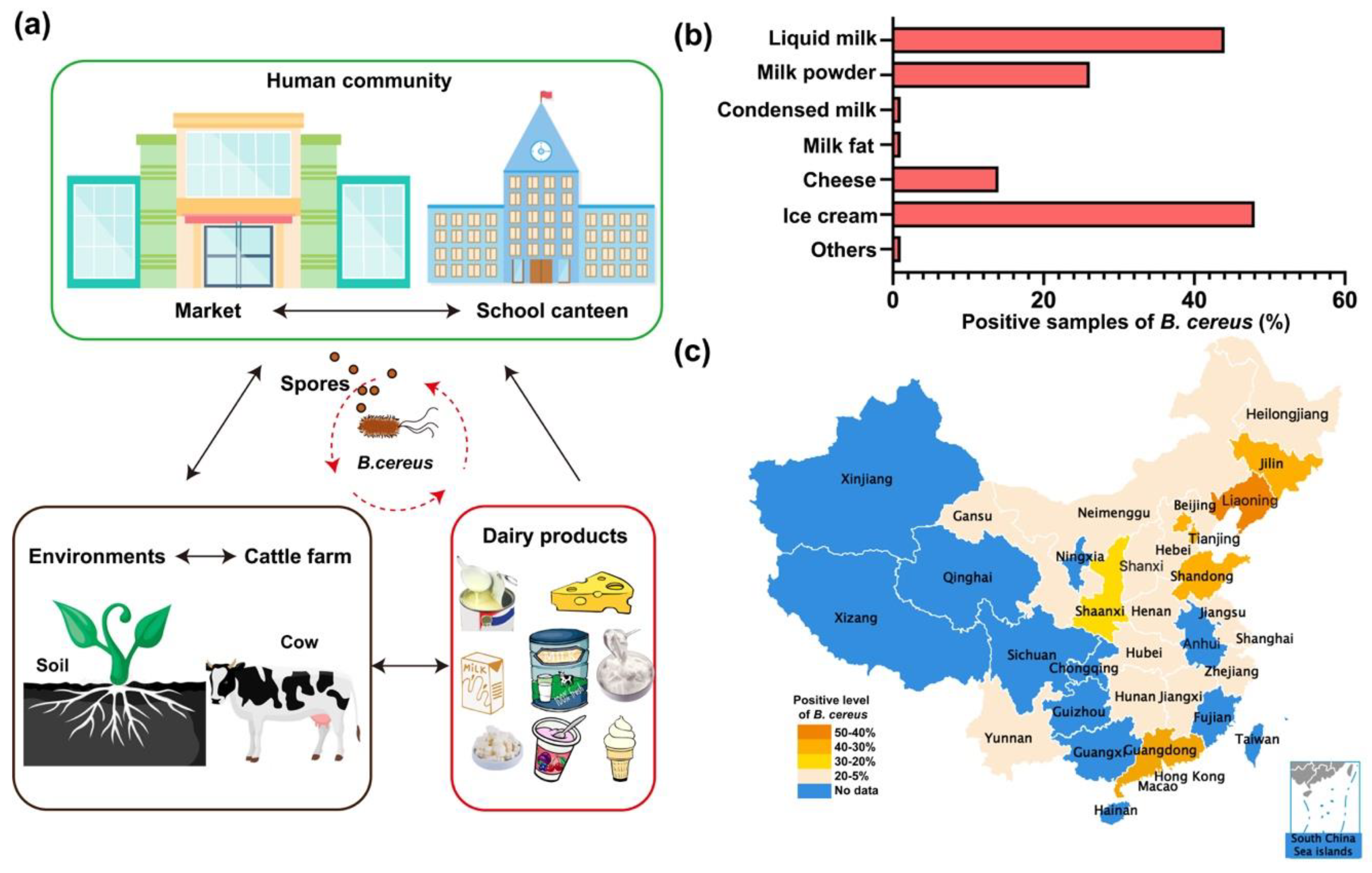
B. cereus prevails in soil and dairy farms and often pollutes foods like raw milk and all dairy products [3,71]. Spores of B. cereus can primarily spread through soil and air [69]. Researches showed that 1g of soil contains 50–380,000 CFUs (colony-forming units) B. cereus spores, and 1 m3 of air has at least 100 CFUs of B. cereus spores. Thus, the abundant B. cereus group spores in the environment are a major cause of the high prevalence rate of B. cereus [5,72]. B. cereus and its spores subsequently have a great opportunity to circularly contaminate dairy farms, human market, food supply places, and dairy products and colonize the intestinal tract of invertebrates and cause illness in humans afterwards (Figure 3a). The Centers for Disease Control (CDC) website claimed that there were 619 confirmed outbreaks of Bacillus-related poisoning from 1998 to 2015 [2,73]. Specifically, the diarrheal illness caused by B. cereus is often related to meats, milk, vegetables and fish, while the emetic type is most possibly associated with rice products [15]. Previous studies have shown that B. cereus isolated from raw milk have the ability to remain active after pasteurization or ultra-high temperature (UHT) sterilization, which ensues in bacterial pollution in the final products [12,66,73,74]. Thus, the health hazards originated from B. cereus in milk industry in China require rapid and proper handling.
3.2. Distribution of Bacillus cereus in Milk and Milk Products in China
The prevalence of B. cereus in China has distinct traits, owing to the specific market demands and physical differences. Liquid milk and milk powder are consumed by the largest part of the population in China (Figure 2a). It is quite a coincidence that a high prevalence rate of B. cereus appears in liquid milk (44%) and milk powder (26.1%) (Figure 3b). A recent report claimed that B. cereus were widely present in pasteurized milk in China, showing that 100 of B. cereus isolates are distributed in most Chinese cities including Hong Kong, Guangzhou, Shenzhen, Harbin, Ningxia, Beihai, Hai kou, etc. [14]. In general, there is a relatively lower prevalence rate of B. cereus in dairy products in southern China than in the northern region with exceptions (Table 1 and Figure 3c). For instance, infant formula in the Liaoning province (42%) was contaminated with B. cereus more seriously than that in Yunnan province (12%) (Table 1). The regular rates of B. cereus in other cities or provinces, such as Beijing, Liaoning, Gansu, Yunnan, northeast China, were found to be 30%, 27%, 19%, 10% and 16%, respectively [13,75,76,77]. Among these data, Yunnan province and the Northeast China indeed had a relatively low prevalence of B. cereus in powdered infant formula (PIF). Dairy products in Gansu province was polluted by B. cereus to a moderate extent (19%). The most striking data in Table 1 were observed in Beijing and Liaoning province, and the investigation conducted in major cities of China during 2011–2016 suggested that approximately 27% of the pasteurized milk contained B. cereus, and the contamination of B. cereus was 31% (11/36) in northern China and 25% (33/132) in southern China. Concerning infant formula in the Chinese market, two reports were produced in 2012–2013 and 2015, according to which 14% and 42% of formula contained B. cereus, respectively. It was also revealed that 8.2% of PIF samples in China were contaminated with B. cereus strains [78]. Overall, the regional characteristics of the prevalence of B. cereus in dairy products in China cannot be clearly defined by latitudes or longitudes. More scientific research into the epidemiological nature of B. cereus in milk is worth pursuing. From an international perspective, dairy products from African nations are more likely to be polluted by B. cereus, which is aided by the poor sanitary conditions [74]. A nationwide survey conducted in America manifested that a total of 18 (8.9%) of 202 samples were positive for presumptive B. cereus using the MPN technique (<10 to 50 CFU/mL), which cannot be directly compared with those of most other studies [79]. Remarkably, ice cream tested in Bavaria, Germany, was found to carry B. cereus with a high rate of 62.7%, and artisan cheese sold in Mexico had a rather low rate of 28.4% [80]. Nevertheless, no certain epidemiological profile can be obtained from the scanty information on dairy products, domestically and globally. Therefore, it is urgent for countries to address the prevalence of Bacillus cereus in their territory.
4. Virulence Factors of Bacillus cereus and Detection Techniques
Many diseases caused by Bacillus groups, such as bovine mastitis, are associated with the virulence factors [40,89,90]. Therefore, it is necessary to further detect the virulence factors secreted from B. cereus isolates in a clinical setting. In particular, the analytical assays of virulence factors in a clinical setting would be enormously beneficial to understand the pathogenic mechanism of virulence factors of B. cereus and to develop effective treatment strategies.
4.1. Virulence Factors of Bacillus cereus
The PlcR regulator in B. cereus is a transcriptional regulator that controls some of the most known virulence factors. It activates gene expression by binding to a nucleotidic sequence called the “PlcR box” [16,90,91]. PlcR regulator is mainly responsible for the transcription of the genes of metalloproteases (InhA2 and Enhancin), hemolysins (CLO and CytK), enterotoxins (Haemolysin BL, Hbl and Nonhemolytic enterotoxin, Nhe) and phospholipases (PI-PLC, PC-PLC and SM-PLC) [24,92]. Another emetic toxin of B. cereus, cereulide, whose synthesis is independent of PlcR, belongs to the Spo0A-AbrB regulon [93]. Cereulide is encoded by the 24-kb cereulide synthetase gene (ces) cluster that located on a megaplasmid of pXO1 [94,95].
4.2. Detection of Bacillus cereus Isolates
B. cereus is a ubiquitous Gram-positive, aerobic or facultative anaerobic, endospore-forming, rod-shaped bacterium [13,71]. The detection and isolation of B. cereus strains are mainly based on the colony count technique ISO 7932 [96]. B. cereus or presumptive colonies of Bacillus are counted on the varieties of Bacillus agar by spiral-plating or spread-plating techniques, most probable number (MPN) method and so on [97]. Both B. cereus cells and spores in the examined products can be counted according to the colony-forming units (CFUs) [5,98,99]. In addition, B. cereus isolates have the capacity for casein, starch and tributyrin hydrolysis as well as lactose fermentation, which inspired the invention of chromogenic medium for B. cereus [100,101]. Instead, the identification or analysis of B. cereus isolates also can use the PCR, the quantitative real-time PCR by targeting the 16S rRNA gene [102,103], groEL/gyrB genes [104] and panC gene [105,106,107], or cross-priming amplification [108] and so on.
4.3. Detection of Toxins Secreted from Bacillus cereus
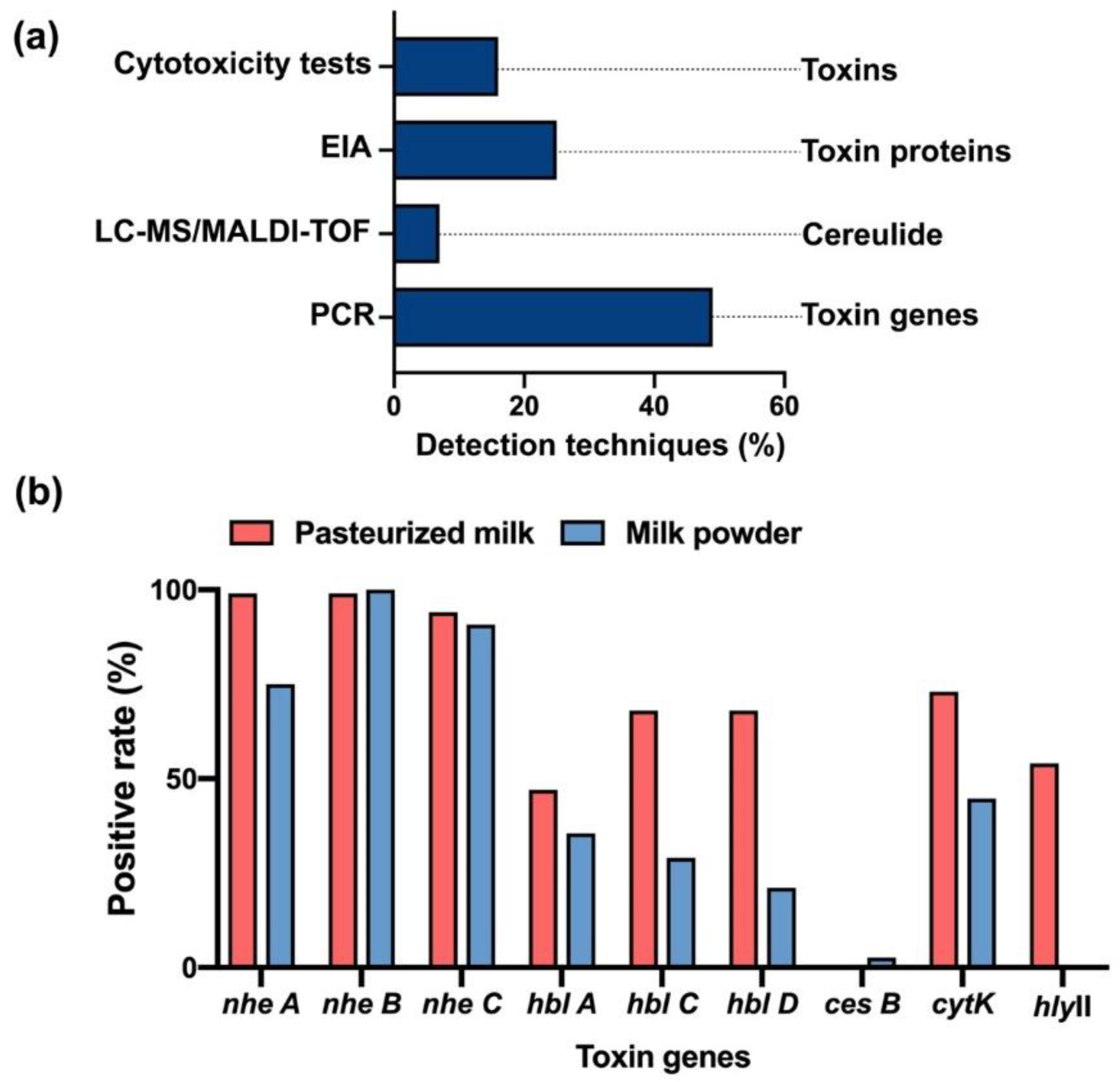
PCR, RT-PCR and multiplex PCR are the major analytical techniques that are reported in research articles to identify the virulence factors by detecting toxin genes in B. cereus (Figure 4a). As we know, the tripartite enterotoxins–Nhe complex consists of NheA, NheB and NheC that were encoded by nheA, nheB and nheC genes [19], as well as the components of Hbl-L2, L1, B were encoded by hblA, hblC and hblD genes separately [21]. Thus, Nhe and Hbl were usually recognized though PCR by targeting their toxic genes [109]. Clinically, a higher rate of nheA, B, C genes than that of hblA, hblC and hblD genes was disclosed in Bacillus samples (Table 1). Other toxin genes such as cytK and cytK-2 genes of CytK, hly gene of hemolysins or cesB gene of cereulide are also served as the main approaches to determine the positive strains of B. cereus by PCR [23,110,111,112,113,114,115,116]. Furthermore, another detection tool, Enzyme Immunoassay (EIA), is employed for the direct inspection on the protein level of the toxic components of B. cereus by targeting specific mAbs [117,118], which are able to purify toxins such as NheB and the NheB-C complex and neutralize the cytotoxicity [21,119].
Liquid chromatography–mass spectrometry (LC-MS) and matrix assisted laser desorption/ionization-time of flight (MALDI-TOF) analysis are often used for the rapid detection of emetic Bacillus isolates in food products, by analyzing ribosomal subunit proteins [120] or targeting the distinct molecular of cereulide [70,121,122,123,124]. Cereulide is a cyclic molecule composed of a 36-membered ring and a small heat and acid stable cyclic dodecadepsipeptide of 1165 Da, which consist of alternating ester and amide bonds and the structure [-D-O-Leu-D-Ala-L-O-Val-L-Val-]3 [23]. Besides, cereulide belongs to the surfactin-like peptides and is biosynthesized via nonribosomal peptide synthesis (NRPS) [94]. Recent reports revealed that the identification of cereulide from bacterial extracts peak at m/z 1191 with a limit of detection (LOD) of 30 ng/mL [121].
Normally, cytotoxicity tests or cell culture methods are used for evaluating the virulence of B. cereus isolates. The cytotoxicity of the complexes of diarrheal Bacillus enterotoxins accounts for over 90% of the total toxicity [109]. A study showed that Vero and primary endothelial cells (HUVEC) were most sensitive to Nhe, whereas Hep-G2, Vero and A549 cell lines were highly susceptible to Nhe and Hbl. CytK exhibited the highest toxicity on CaCo-2 cells [40], and the emetic toxin cereulide prevented cell proliferation in HepG2 cells by 2 nM [35], also causing vacuolation in HEp-2 cells [125,126]. In general, B. cereus toxins exhibited a wide cytotoxicity to those epithelial cells [127] and some toxins also acted on immune cells [39,128].
4.4. Analysis of Bacillus Toxin Detections in Dairy Products in China
B. cereus and its virulence factors are frequently present in dairy products in China [12,13,14,15,57,58]. This not only impacts the quality of dairy products but also potentially impairs human health. Since there is a huge demand for dairy products among Chinese people of all ages [50,53], once Bacillus contamination has occurred, the scope of the damage is extensive and inestimable.
Currently, there is a large number of potential toxin genes related to diarrhea in these B. cereus strains, including the genes of hcbl, nhe, cytK and enterotoxin FM (entFM), as well as potential enterotoxins hlyII and enterotoxin T (BceT) [14]. As shown in Table 2, a study conducted in 10 local dairy farms in Beijing suggested that the nhe, hbl, and ces genes were detected at the rate of 100%, 79.3%, and 1.1%, respectively [13]. Meanwhile, a research involving 12 provinces in China showed that the average virulence gene number in powdered milk was 5.71, and no comparison of the distribution of those genes between different provinces was made [129]. It is logical to conclude that the virulence factors of B. cereus can exert influence on the quality of dairy products during their processing, transporting and selling. Still, like B. cereus, no striking feature of the distribution of their virulence factors in diverse provinces could be obtained. Similarly, high numbers of isolates carried nheA (84.1%), nheB (89.9%), nheC (84.1%), hblA (59.4%), hblC (44.9%), hblD (53.6%) and cytK (53.6%) genes in the production chain of milk in Brazil [62], which is notably bigger than the rate found in Ghana [73]. It is really challenging to sum up the rule of distribution of Bacillus toxins in dairy products in China, and this is limited by the deficient detection methods and shortage of useful data, as the notion of Bacillus contamination in dairy products only came into view in recent years.
To produce a more forthright analysis of the Bacillus toxins, we collected the prevalence of the toxin genes in Bacillus samples from pasteurized milk and milk powder as they are the most popular dairy products in China [12,13,14,15,53,57,82,112] (Figure 3a). The data showed that the genes of nhe A B C were found in almost all the Bacillus isolates from pasteurized milk, while at least 80% of nhe A and approximate 100% nhe B and C in milk powder (Table 1 and Table 2 and Figure 4b). More genes of hbl A C D were discovered in pasteurized milk (45%) than in milk powder (36.8%). Similarly, the positive rate of cytK genes in pasteurized milk and milk powder was 73% and 44.7%, respectively. The cesB gene was mainly found in milk powder (2.6%), and hlyII gene only grew in pasteurized milk (54%) (Figure 4b).
However, the detection techniques of B. cereus and its virulence factors are strongly limited [5,110,114], and B. cereus diagnostic method is still a field to be developed [135]. The widely used nucleic acid-based detection technology cannot accurately determine the bacterial activity and toxin expression [110,114]. Thus, the data analysis was incomplete since it is only based on the occurrence of toxin genes, while the level of protein expression and its toxic effects stay implicit. Therefore, the diversity and perfection of detection techniques of B. cereus and its virulence factors are necessary to improve the safety of dairy products in the future. Beyond that, the toxicity mechanisms of Bacillus virulence factors deserve to be illustrated, as host cells have frequently interacted with B. cereus infection [1,31,33,136,137]. Thus, comprehending the cytotoxicity of B. cereus is greatly beneficial in providing the therapeutic strategy for related illnesses caused by intaking Bacillus-contaminated dairy products. We believe that the summary of detection techniques of B. cereus will certainly be helpful for a more accurate examination and evaluation in the future.
5. Conclusions and Future Perspectives
The awareness of food safety in China has risen significantly with the mounting needs for high-quality foods. However, there still exist food poisoning incidents caused by bacteria like B. cereus in dairy products [5,12,13,70]. B. cereus group is an opportunistic spore-producing pathogen that causes food poisoning with symptoms of vomiting and diarrhea, exhalation of toxins that are the main culprit of damaging liver tissue and inflammatory diseases such as gastroenteritis and meningitis [16,17]. The infectious bacteria were the main focus of some widespread epidemics in history, and therefore, the safety of dairy products should not ever be ignored. In the past decades, although we have made arduous efforts to ensure food safety, the contamination of dairy products with B. cereus is still an issue in China. With regard to the average level in China, the investigation carried out from 2011 to 2016 unveiled that about 27.1% of the pasteurized milk on shelf were infested with B. cereus and also that the environments of milk production, handling and processing could introduce B. cereus into milk products. Together, these assessments implied the high prevalence of B. cereus and existence of potential hazards in contaminated pasteurized milk (Figure 3 and Table 1).
On the other hand, Bacillus strains can also be used as human probiotics [138], and this field is gaining greater attention [90,139], as B. cereus strains or spores serve as probiotics for human use [140,141]. Some countries even utilize their connection with the dairy chain as a source to culture novel probiotic products [142]; however, the consequences could be unfavorable. In contrast to chemical drugs that quantitatively decrease or remain unchanged after metabolic process, the variation trend of the amounts of microbes like B. cereus in probiotics is undefined and could even rise exponentially.
In this review, we summarized the risk of B. cereus in dairy products in China and provided the analytical assays of B. cereus and its toxins. PCR is the most commonly used analysis method, accounting for 49% of articles on B. cereus (Figure 4a). However, the expression of genes does not completely represent the toxicity of the virulence factors. We need more comprehensive and rapid testing methods such as cytotoxicity tests or LC-MS analysis. Thus, the role in cell toxicity of the virulence factors of B. cereus is unquestionably important, and understanding the actions of B. cereus and its toxins at the cellular level would benefit the prevention of Bacillus infections. In addition, the cellular mechanism and the interaction between different virulence factors should be further studied. Recent reports showed that the persistent B. cereus interaction with host cells is even hard to control [1,49,143], and compared to other persistent bacteria, B. cereus are more dangerous due to the high transmission and viability of their spores [69,99,144]. In sum, strong emphasis should be placed on the B. cereus in dairy products to guarantee the safety of human life in China.
Author Contributions
K.Z. and S.-Y.D. conceived the project. X.-Y.L. and Q.H. prepared the figures and tables. X.-Y.L., Q.H. and F.X. performed data collection. X.-Y.L., Q.H. and K.Z. wrote the manuscript. All authors read and approved the manuscript.
Funding
This work was supported by the National Key Research and Development Program of China (2017YFC1600305), Fund of Beijing Innovation Team of Dairy Industry and Start-up Fund from China Agricultural University (2020RC002).
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Liu, X.; Liu, F.; Ding, S.; Shen, J.; Zhu, K. Sublethal levels of antibiotics promote bacterial persistence in epithelial cells. Adv. Sci. 2020. [Google Scholar] [CrossRef]
- Stenfors Arnesen, L.P.; Fagerlund, A.; Granum, P.E. From soil to gut: Bacillus cereus and its food poisoning toxins. FEMS Microbiol. Rev. 2008, 32, 579–606. [Google Scholar] [CrossRef] [Green Version]
- Rigourd, V.; Barnier, J.P.; Ferroni, A.; Nicloux, M.; Hachem, T.; Magny, J.F.; Lapillonne, A.; Frange, P.; Nassif, X.; Bille, E. Recent actuality about Bacillus cereus and human milk bank: A new sensitive method for microbiological analysis of pasteurized milk. Eur. J. Clin. Microbiol. Infect. Dis. 2018, 37, 1297–1303. [Google Scholar] [CrossRef] [PubMed]
- Abdeen, E.E.; Hussien, H.; Hadad, G.A.E.; Mousa, W.S. Prevalence of virulence determinants among Bacillus cereus isolated from milk products with potential public health concern. Pak. J. Biol. Sci. 2020, 23, 206–212. [Google Scholar] [PubMed] [Green Version]
- Vidic, J.; Chaix, C.; Manzano, M.; Heyndrickx, M. Food sensing: detection of Bacillus cereus spores in dairy products. Biosensors 2020, 10, 15. [Google Scholar] [CrossRef] [Green Version]
- Fiedler, G.; Schneider, C.; Igbinosa, E.O.; Kabisch, J.; Brinks, E.; Becker, B.; Stoll, D.A.; Cho, G.S.; Huch, M.; Franz, C. Antibiotics resistance and toxin profiles of Bacillus cereus-group isolates from fresh vegetables from German retail markets. BMC Microbiol. 2019, 19, 250. [Google Scholar] [CrossRef]
- Zhang, Y.; Chen, J.; Feng, C.; Zhan, L.; Zhang, J.; Li, Y.; Yang, Y.; Chen, H.; Zhang, Z.; Zhang, Y.; et al. Quantitative prevalence, phenotypic and genotypic characteristics of Bacillus cereus isolated from retail infant foods in China. Foodborne Pathog. Dis. 2017, 14, 564–572. [Google Scholar] [CrossRef]
- Tunick, M.H.; Van Hekken, D.L. Dairy products and health: recent insights. J. Agric. Food Chem. 2015, 63, 9381–9388. [Google Scholar] [CrossRef]
- Godos, J.; Tieri, M.; Ghelfi, F.; Titta, L.; Marventano, S.; Lafranconi, A.; Gambera, A.; Alonzo, E.; Sciacca, S.; Buscemi, S.; et al. Dairy foods and health: An umbrella review of observational studies. Int. J. Food Sci. Nutr. 2020, 71, 138–151. [Google Scholar] [CrossRef]
- Romero-Alvarez, D.; Peterson, A.T.; Salzer, J.S.; Pittiglio, C.; Shadomy, S.; Traxler, R.; Vieira, A.R.; Bower, W.A.; Walke, H.; Campbell, L.P. Potential distributions of Bacillus anthracis and Bacillus cereus biovar anthracis causing anthrax in Africa. PLoS Negl. Trop. Dis. 2020, 14, e0008131. [Google Scholar] [CrossRef]
- Guo, R.; Tian, Y.; Zhang, H.; Guo, D.; Pei, X.; Wen, H.; Li, P.; Mehmood, K.; Yang, K.; Chang, Y.F.; et al. Biological characteristics and genetic evolutionary analysis of emerging pathogenic Bacillus cereus isolated from Pere David’s Deer (Elaphurus davidianus). Microb. Pathog. 2020. [Google Scholar] [CrossRef] [PubMed]
- Zhao, S.; Chen, J.; Fei, P.; Feng, H.; Wang, Y.; Ali, M.A.; Li, S.; Jing, H.; Yang, W. Prevalence, molecular characterization, and antibiotic susceptibility of Bacillus cereus isolated from dairy products in China. J. Dairy Sci. 2020, 103, 3994–4001. [Google Scholar] [CrossRef] [PubMed]
- Cui, Y.; Liu, X.; Dietrich, R.; Martlbauer, E.; Cao, J.; Ding, S.; Zhu, K. Characterization of Bacillus cereus isolates from local dairy farms in China. FEMS Microbiol. Lett. 2016, 363. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, T.; Ding, Y.; Wu, Q.; Wang, J.; Zhang, J.; Yu, S.; Yu, P.; Liu, C.; Kong, L.; Feng, Z.; et al. Prevalence, virulence Genes, antimicrobial susceptibility, and genetic diversity of Bacillus cereus isolated from pasteurized milk in China. Front Microbiol 2018, 9, 533. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, S.; Yu, P.; Wang, J.; Li, C.; Guo, H.; Liu, C.; Kong, L.; Yu, L.; Wu, S.; Lei, T.; et al. A study on prevalence and characterization of Bacillus cereus in ready-to-eat foods in China. Front. Microbiol. 2019, 10, 3043. [Google Scholar] [CrossRef] [Green Version]
- Granum, P.E. Spotlight on Bacillus cereus and its food poisoning toxins. FEMS Microbiol. Lett. 2017, 364. [Google Scholar] [CrossRef] [Green Version]
- Dal Peraro, M.; van der Goot, F.G. Pore-forming toxins: Ancient, but never really out of fashion. Nat Rev. Microbiol. 2016, 14, 77–92. [Google Scholar] [CrossRef]
- Zhu, K.; Didier, A.; Dietrich, R.; Heilkenbrinker, U.; Waltenberger, E.; Jessberger, N.; Martlbauer, E.; Benz, R. Formation of small transmembrane pores: An intermediate stage on the way to Bacillus cereus non-hemolytic enterotoxin (Nhe) full pores in the absence of NheA. Biochem. Biophys. Res. Commun. 2016, 469, 613–618. [Google Scholar] [CrossRef]
- Fagerlund, A.; Lindback, T.; Storset, A.K.; Granum, P.E.; Hardy, S.P. Bacillus cereus Nhe is a pore-forming toxin with structural and functional properties similar to the ClyA (HlyE, SheA) family of haemolysins, able to induce osmotic lysis in epithelia. Microbiology 2008, 154, 693–704. [Google Scholar] [CrossRef] [Green Version]
- Jessberger, N.; Dietrich, R.; Schwemmer, S.; Tausch, F.; Schwenk, V.; Didier, A.; Martlbauer, E. Binding to the target cell surface is the crucial step in pore formation of hemolysin BL from Bacillus cereus. Toxins 2019, 11, 281. [Google Scholar] [CrossRef] [Green Version]
- Fagerlund, A.; Lindback, T.; Granum, P.E. Bacillus cereus cytotoxins Hbl, Nhe and CytK are secreted via the Sec translocation pathway. BMC Microbiol. 2010, 10, 304. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ehling-Schulz, M.; Svensson, B.; Guinebretiere, M.H.; Lindback, T.; Andersson, M.; Schulz, A.; Fricker, M.; Christiansson, A.; Granum, P.E.; Martlbauer, E.; et al. Emetic toxin formation of Bacillus cereus is restricted to a single evolutionary lineage of closely related strains. Microbiology 2005, 151, 183–197. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lucking, G.; Frenzel, E.; Rutschle, A.; Marxen, S.; Stark, T.D.; Hofmann, T.; Scherer, S.; Ehling-Schulz, M. Ces locus embedded proteins control the non-ribosomal synthesis of the cereulide toxin in emetic Bacillus cereus on multiple levels. Front. Microbiol. 2015, 6, 1101. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramarao, N.; Sanchis, V. The pore-forming haemolysins of Bacillus cereus: A review. Toxins 2013, 5, 1119–1139. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Madegowda, M.; Eswaramoorthy, S.; Burley, S.K.; Swaminathan, S. X-ray crystal structure of the B component of hemolysin BL from Bacillus cereus. Proteins 2008, 71, 534–540. [Google Scholar] [CrossRef] [Green Version]
- Los, F.C.; Randis, T.M.; Aroian, R.V.; Ratner, A.J. Role of pore-forming toxins in bacterial infectious diseases. Microbiol. Mol. Biol. Rev. 2013, 77, 173–207. [Google Scholar] [CrossRef] [Green Version]
- Bischofberger, M.; Iacovache, I.; van der Goot, F.G. Pathogenic pore-forming proteins: function and host response. Cell Host Microbe 2012, 12, 266–275. [Google Scholar] [CrossRef] [Green Version]
- Alves, G.G.; Machado de Avila, R.A.; Chavez-Olortegui, C.D.; Lobato, F.C. Clostridium perfringens epsilon toxin: the third most potent bacterial toxin known. Anaerobe 2014, 30, 102–107. [Google Scholar] [CrossRef]
- Lindback, T.; Fagerlund, A.; Rodland, M.S.; Granum, P.E. Characterization of the Bacillus cereus Nhe enterotoxin. Microbiology 2004, 150, 3959–3967. [Google Scholar] [CrossRef] [Green Version]
- Heilkenbrinker, U.; Dietrich, R.; Didier, A.; Zhu, K.; Lindback, T.; Granum, P.E.; Martlbauer, E. Complex formation between NheB and NheC is necessary to induce cytotoxic activity by the three-component Bacillus cereus Nhe enterotoxin. PLoS ONE 2013, 8, e63104. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.; Zuo, Z.; Sastalla, I.; Liu, C.; Jang, J.Y.; Sekine, Y.; Li, Y.; Pirooznia, M.; Leppla, S.H.; Finkel, T.; et al. Sequential CRISPR-Based Screens Identify LITAF and CDIP1 as the Bacillus cereus Hemolysin BL Toxin Host Receptors. Cell Host Microbe 2020. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Ding, S.; Shi, P.; Dietrich, R.; Martlbauer, E.; Zhu, K. Non-hemolytic enterotoxin of Bacillus cereus induces apoptosis in Vero cells. Cell. Microbiol. 2017, 19. [Google Scholar] [CrossRef] [PubMed]
- Fox, D.; Mathur, A.; Xue, Y.; Liu, Y.; Tan, W.H.; Feng, S.; Pandey, A.; Ngo, C.; Hayward, J.A.; Atmosukarto, I.I.; et al. Bacillus cereus non-haemolytic enterotoxin activates the NLRP3 inflammasome. Nat. Commun. 2020, 11, 760. [Google Scholar] [PubMed]
- Doll, V.M.; Ehling-Schulz, M.; Vogelmann, R. Concerted action of sphingomyelinase and non-hemolytic enterotoxin in pathogenic Bacillus cereus. PLoS ONE 2013, 8, e61404. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Andersson, M.A.; Hakulinen, P.; Honkalampi-Hamalainen, U.; Hoornstra, D.; Lhuguenot, J.C.; Maki-Paakkanen, J.; Savolainen, M.; Severin, I.; Stammati, A.L.; Turco, L.; et al. Toxicological profile of cereulide, the Bacillus cereus emetic toxin, in functional assays with human, animal and bacterial cells. Toxicon 2007, 49, 351–367. [Google Scholar] [CrossRef]
- Paananen, A.; Jarvinen, K.; Sareneva, T.; Salkinoja-Salonen, M.S.; Timonen, T.; Holtta, E. Valinomycin-induced apoptosis of human NK cells is predominantly caspase independent. Toxicology 2005, 212, 37–45. [Google Scholar] [CrossRef]
- Vangoitsenhoven, R.; Rondas, D.; Crevecoeur, I.; D’Hertog, W.; Baatsen, P.; Masini, M.; Andjelkovic, M.; Van Loco, J.; Matthys, C.; Mathieu, C.; et al. Foodborne cereulide causes beta-cell dysfunction and apoptosis. PLoS ONE 2014, 9, e104866. [Google Scholar] [CrossRef]
- Bauer, T.; Sipos, W.; Stark, T.D.; Kaser, T.; Knecht, C.; Brunthaler, R.; Saalmuller, A.; Hofmann, T.; Ehling-Schulz, M. First insights into within host translocation of the Bacillus cereus toxin cereulide using a porcine model. Front. Microbiol. 2018, 9, 2652. [Google Scholar] [CrossRef] [Green Version]
- Tran, S.-L.; Guillemet, E.; Ngo-Camus, M.; Clybouw, C.; Puhar, A.; Moris, A.; Gohar, M.; Lereclus, D.; Ramarao, N. Haemolysin II is a Bacillus cereus virulence factor that induces apoptosis of macrophages. Cell. Microbiol. 2011, 13, 92–108. [Google Scholar] [CrossRef]
- Jessberger, N.; Kranzler, M.; Da Riol, C.; Schwenk, V.; Buchacher, T.; Dietrich, R.; Ehling-Schulz, M.; Martlbauer, E. Assessing the toxic potential of enteropathogenic Bacillus cereus. Food Microbiol. 2019, 84, 103276. [Google Scholar] [CrossRef]
- Berthold-Pluta, A.; Pluta, A.; Garbowska, M. The effect of selected factors on the survival of Bacillus cereus in the human gastrointestinal tract. Microb. Pathog. 2015, 82, 7–14. [Google Scholar] [CrossRef] [PubMed]
- Koizumi, Y.; Okuno, T.; Minamiguchi, H.; Hodohara, K.; Mikamo, H.; Andoh, A. Survival of a case of Bacillus cereus meningitis with brain abscess presenting as immune reconstitution syndrome after febrile neutropenia—A case report and literature review. BMC Infect. Dis. 2020, 20, 15. [Google Scholar] [CrossRef] [PubMed]
- Meredith, F.T.; Fowler, V.G.; Gautier, M.; Corey, G.R.; Reller, L.B. Bacillus cereus necrotizing cellulitis mimicking clostridial myonecrosis: case report and review of the literature. Scand. J. Infect. Dis. 1997, 29, 528–529. [Google Scholar] [CrossRef] [PubMed]
- Pan, Q.; Liu, Y.; Wang, R.; Chen, T.; Yang, Z.; Deng, Y.; Zhao, Z.; Hu, X.; Chen, X.; Wei, W.; et al. Treatment of Bacillus cereus endophthalmitis with endoscopy-assisted vitrectomy. Medicine 2017, 96, e8701. [Google Scholar] [CrossRef] [PubMed]
- David, D.B.; Kirkby, G.R.; Noble, B.A. Bacillus cereus endophthalmitis. Br. J. Ophthalmol. 1994, 78, 577–580. [Google Scholar] [CrossRef] [Green Version]
- Ishida, R.; Ueda, K.; Kitano, T.; Yamamoto, T.; Mizutani, Y.; Tsutsumi, Y.; Imoto, K.; Yamamori, Y. Fatal community-acquired Bacillus cereus pneumonia in an immunocompetent adult man: A case report. BMC Infect. Dis. 2019, 19, 197. [Google Scholar] [CrossRef]
- Soudet, S.; Becquart, C.; Dezoteux, F.; Faure, K.; Staumont-Salle, D.; Delaporte, E. Bacillus cereus endocarditis and a probable cutaneous gateway. Ann. Dermatol. Venereol. 2017, 144, 45–48. [Google Scholar] [CrossRef]
- Avcu, G.; Sahbudak Bal, Z.; Cavusoglu, C.; Vardar, F. Osteomyelitis caused by Bacillus calmette-guerin vaccination in a healthy toddler. J. Trop. Pediatr. 2020, 66, 103–105. [Google Scholar] [CrossRef]
- Schaefer, G.; Campbell, W.; Jenks, J.; Beesley, C.; Katsivas, T.; Hoffmaster, A.; Mehta, S.R.; Reed, S. Persistent Bacillus cereus bacteremia in 3 persons who inject drugs, San Diego, California, USA. Emerg. Infect. Dis. 2016, 22, 1621–1623. [Google Scholar] [CrossRef] [Green Version]
- Ballard, O.; Morrow, A.L. Human milk composition: nutrients and bioactive factors. Pediatr. Clin. North Am. 2013, 60, 49–74. [Google Scholar] [CrossRef] [Green Version]
- Schroeter, H.; Holt, R.R.; Orozco, T.J.; Schmitz, H.H.; Keen, C.L. Nutrition: Milk and absorption of dietary flavanols. Nature 2003, 426, 787–788. [Google Scholar] [CrossRef] [PubMed]
- Rosa, D.D.; Dias, M.M.S.; Grzeskowiak, L.M.; Reis, S.A.; Conceicao, L.L.; Peluzio, M. Milk kefir: nutritional, microbiological and health benefits. Nutr. Res. Rev. 2017, 30, 82–96. [Google Scholar] [CrossRef] [PubMed]
- He, Y.; Yang, X.; Xia, J.; Zhao, L.; Yang, Y. Consumption of meat and dairy products in China: A review. Proc. Nutr. Soc. 2016, 75, 385–391. [Google Scholar] [CrossRef] [Green Version]
- Lagrange, V.; Whitsett, D.; Burris, C. Global market for dairy proteins. J. Food. Sci. 2015, 80, A16–A22. [Google Scholar] [CrossRef] [PubMed]
- Bian, S.; Hu, J.; Zhang, K.; Wang, Y.; Yu, M.; Ma, J. Dairy product consumption and risk of hip fracture: A systematic review and meta-analysis. BMC Public Health 2018, 18, 165. [Google Scholar] [CrossRef] [Green Version]
- Gao, Y.; Sun, Y.; Chen, J.; Tian, J. The quality of overviews on milk and dairy product consumption and cardiovascular diseases and milk and dairy product consumption and risk of mortality can be improved. Adv. Nutr. 2020, 11, 464. [Google Scholar] [CrossRef] [PubMed]
- Chen, D.; Li, Y.; Lv, J.; Liu, X.; Gao, P.; Zhen, G.; Zhang, W.; Wu, D.; Jing, H.; Li, Y.; et al. A foodborne outbreak of gastroenteritis caused by Norovirus and Bacillus cereus at a university in the Shunyi District of Beijing, China 2018: A retrospective cohort study. BMC Infect. Dis. 2019, 19, 910. [Google Scholar] [CrossRef] [Green Version]
- Thirkell, C.E.; Sloan-Gardner, T.S.; Kacmarek, M.C.; Polkinghorne, B. An outbreak of Bacillus cereus toxin-mediated emetic and diarrhoeal syndromes at a restaurant in Canberra, Australia 2018. Commun. Dis. Intell. (2018) 2019, 43. [Google Scholar] [CrossRef]
- Chai, S.J.; Gu, W.; O’Connor, K.A.; Richardson, L.C.; Tauxe, R.V. Incubation periods of enteric illnesses in foodborne outbreaks, United States, 1998–2013. Epidemiol. Infect. 2019, 147, e285. [Google Scholar] [CrossRef] [Green Version]
- Ceuppens, S.; Rajkovic, A.; Heyndrickx, M.; Tsilia, V.; Van De Wiele, T.; Boon, N.; Uyttendaele, M. Regulation of toxin production by Bacillus cereus and its food safety implications. Crit. Rev. Microbiol. 2011, 37, 188–213. [Google Scholar] [CrossRef]
- Ombui, J.N.; Nduhiu, J.G. Prevalence of enterotoxigenic Bacillus cereus and its enterotoxins in milk and milk products in and around Nairobi. East Afr. Med. J. 2005, 82, 280–284. [Google Scholar] [CrossRef] [PubMed]
- Rossi, G.A.M.; Silva, H.O.; Aguilar, C.E.G.; Rochetti, A.L.; Pascoe, B.; Méric, G.; Mourkas, E.; Hitchings, M.D.; Mathias, L.A.; de Azevedo Ruiz, V.L.; et al. Comparative genomic survey of Bacillus cereus sensu stricto isolates from the dairy production chain in Brazil. FEMS Microbiol. Lett. 2018, 365. [Google Scholar] [CrossRef] [PubMed]
- Fasolato, L.; Cardazzo, B.; Carraro, L.; Fontana, F.; Novelli, E.; Balzan, S. Edible processed insects from e-commerce: Food safety with a focus on the Bacillus cereus group. Food Microbiol. 2018, 76, 296–303. [Google Scholar] [CrossRef] [PubMed]
- Boor, K.J.; Wiedmann, M.; Murphy, S.; Alcaine, S. A 100-year review: microbiology and safety of milk handling. J. Dairy Sci. 2017, 100, 9933–9951. [Google Scholar] [CrossRef] [Green Version]
- Becker, H.; Schaller, G.; von Wiese, W.; Terplan, G. Bacillus cereus in infant foods and dried milk products. Int. J. Food Microbiol. 1994, 23, 1–15. [Google Scholar] [CrossRef]
- Zhou, G.; Liu, H.; He, J.; Yuan, Y.; Yuan, Z. The occurrence of Bacillus cereus, B. thuringiensis and B. mycoides in Chinese pasteurized full fat milk. Int. J. Food Microbiol. 2008, 121, 195–200. [Google Scholar] [CrossRef]
- Yusuf, U.; Kotwal, S.K.; Gupta, S.; Ahmed, T. Identification and antibiogram pattern of Bacillus cereus from the milk and milk products in and around Jammu region. Vet. World 2018, 11, 186–191. [Google Scholar] [CrossRef]
- Pedonese, F.; Verani, G.; Torracca, B.; Turchi, B.; Felicioli, A.; Nuvoloni, R. Effect of an Italian propolis on the growth of Listeria monocytogenes, Staphylococcus aureus and Bacillus cereus in milk and whey cheese. Ital. J. Food Saf. 2019, 8, 8036. [Google Scholar] [CrossRef] [Green Version]
- Huang, Y.; Flint, S.H.; Palmer, J.S. Bacillus cereus spores and toxins—The potential role of biofilms. Food Microbiol. 2020, 90, 103493. [Google Scholar] [CrossRef]
- Cui, Y.; Liu, Y.; Liu, X.; Xia, X.; Ding, S.; Zhu, K. Evaluation of the toxicity and toxicokinetics of cereulide from an emetic Bacillus cereus strain of milk origin. Toxins 2016, 8, 156. [Google Scholar] [CrossRef] [Green Version]
- Sankararaman, S.; Velayuthan, S. Bacillus cereus. Pediatr. Rev. 2013, 34, 196–197. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Christiansson, A.; Bertilsson, J.; Svensson, B. Bacillus cereus spores in raw milk: factors affecting the contamination of milk during the grazing period. J. Dairy Sci. 1999, 82, 305–314. [Google Scholar] [CrossRef]
- Owusu-Kwarteng, J.; Wuni, A.; Akabanda, F.; Tano-Debrah, K.; Jespersen, L. Prevalence, virulence factor genes and antibiotic resistance of Bacillus cereus sensu lato isolated from dairy farms and traditional dairy products. BMC Microbiol. 2017, 17, 65. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hanson, M.L.; Wendorff, W.L.; Houck, K.B. Effect of heat treatment of milk on activation of Bacillus spores. J. Food Prot. 2005, 68, 1484–1486. [Google Scholar] [CrossRef] [PubMed]
- Ma, H.; Li, J.; Xi, X.; Xu, H.; Wuri, L.; Bian, Y.; Yu, Z.; Ren, M.; Duo, L.; Sun, Y.; et al. Evaluation of Bacterial contamination in goat milk powder using PacBio single molecule real-time sequencing and droplet digital PCR. J. Food Prot. 2018, 81, 1791–1799. [Google Scholar] [CrossRef] [PubMed]
- Sun, T.; Wang, W.; Han, J.; Zhang, M. Investigation of Bacillus cereus contamination in infant formula food and cereals in Liaoning Province during 2016–2017. Chin. J. Health Lab. Tech. 2019, 29, 1525–1527. [Google Scholar]
- Fei, P.; Yuan, X.; Zhao, S.; Yang, T.; Xiang, J.; Chen, X.; Zhou, L.; Ji, M. Prevalence and genetic diversity of Bacillus cereus isolated from raw milk and cattle farm environments. Curr. Microbiol. 2019, 76, 1355–1360. [Google Scholar] [CrossRef]
- Zhuang, K.; Li, H.; Zhang, Z.; Wu, S.; Zhang, Y.; Fox, E.M.; Man, C.; Jiang, Y. Typing and evaluating heat resistance of Bacillus cereus sensu stricto isolated from the processing environment of powdered infant formula. J. Dairy Sci. 2019, 102, 7781–7793. [Google Scholar] [CrossRef]
- Jackson, E.E.; Erten, E.S.; Maddi, N.; Graham, T.E.; Larkin, J.W.; Blodgett, R.J.; Schlesser, J.E.; Reddy, R.M. Detection and enumeration of four foodborne pathogens in raw commingled silo milk in the United States. J. Food Prot. 2012, 75, 1382–1393. [Google Scholar] [CrossRef]
- Zhou, G.; Zheng, D.; Dou, L.; Cai, Q.; Yuan, Z. Occurrence of psychrotolerant Bacillus cereus group strains in ice creams. Int. J. Food Microbiol. 2010, 137, 143–146. [Google Scholar] [CrossRef]
- Xie, A.; Wu, K.; Li, Y.; Hong, C.; Hu, Y. Contamination survey and toxin genes distribution analysis of Bacillus cereus in milk powder in Wenzhou. Chin. J. Health Lab. Tech. 2018, 28, 2930–2932. [Google Scholar]
- Yang, Y.; Yu, X.; Zhan, L.; Chen, J.; Zhang, Y.; Zhang, J.; Chen, H.; Zhang, Z.; Zhang, Y.; Lu, Y. Multilocus sequence type profiles of Bacillus cereus isolates from infant formula in China. Food Microbiol. 2017, 62, 46–50. [Google Scholar] [CrossRef] [PubMed]
- Yan, S.; Yan, X.; Gan, X.; Li, Z.; Yu, D.; Zhao, X.; Li, F. Survey on contamination of Bacillus cereus and its virulence gene profiles isolated from retail infant formula in China. Chin. J. Food Hyg. 2015, 27, 286–291. [Google Scholar]
- Sun, T.; Wang, W.; Han, J.; Li, X.; Zhang, M.; Liu, H.; Wang, Y.; Zhang, M. Detection of virulence genes of Bacillus cereus in infant formula foods and cereal-based complementary foods in Liaoning in 2016. Chin. J. Microecol. 2018, 30, 791–794. [Google Scholar]
- Tang, X.; Yang, Z.; Fan, L.; Guo, Z. Investigation and assessment of Bacillus cereus contamination in infants and young children foods and ready-to-use foods in Yunnan from 2012 to 2016. J. Food Saf. Qual. 2017, 8, 3785–3789. [Google Scholar]
- Xie, C.; Wei, M.; Wang, H.; Jiang, Y.; Yang, X.; Hou, M. Investigation on hygiene and safety of school periphery food and infants food in Kunming in 2016. J. Food Saf. Qual. 2017, 8, 3903–3906. [Google Scholar]
- Sun, J.; Hu, X.; Lan, G.; Dong, K. Investigation on microbial contamination of infant formula powder during production process. Chin. J. Food Hyg. 2017, 29, 474–477. [Google Scholar]
- Li, Y.; Pei, X.; Yang, D.; Li, N. Occurrence of Bacillus cereus in infants and young children foods in 8 provinces in China. J. Food Res. 2014, 43, 435–438. [Google Scholar]
- Song, X.; Huang, X.; Xu, H.; Zhang, C.; Chen, S.; Liu, F.; Guan, S.; Zhang, S.; Zhu, K.; Wu, C. The prevalence of pathogens causing bovine mastitis and their associated risk factors in 15 large dairy farms in China: An observational study. Vet. Microbiol. 2020, 247. [Google Scholar] [CrossRef]
- Cui, Y.; Martlbauer, E.; Dietrich, R.; Luo, H.; Ding, S.; Zhu, K. Multifaceted toxin profile, an approach toward a better understanding of probiotic Bacillus cereus. Crit. Rev. Toxicol. 2019, 49, 342–356. [Google Scholar] [CrossRef]
- Lyu, Y.; Ye, L.; Xu, J.; Yang, X.; Chen, W.; Yu, H. Recent research progress with phospholipase C from Bacillus cereus. Biotechnol. Lett. 2016, 38, 23–31. [Google Scholar] [CrossRef] [PubMed]
- Gohar, M.; Faegri, K.; Perchat, S.; Ravnum, S.; Okstad, O.A.; Gominet, M.; Kolsto, A.B.; Lereclus, D. The PlcR virulence regulon of Bacillus cereus. PLoS ONE 2008, 3, e2793. [Google Scholar] [CrossRef]
- Lucking, G.; Dommel, M.K.; Scherer, S.; Fouet, A.; Ehling-Schulz, M. Cereulide synthesis in emetic Bacillus cereus is controlled by the transition state regulator AbrB, but not by the virulence regulator PlcR. Microbiology 2009, 155, 922–931. [Google Scholar] [CrossRef] [Green Version]
- Naka, T.; Takaki, Y.; Hattori, Y.; Takenaka, H.; Ohta, Y.; Kirihata, M.; Tanimori, S. Chemical structure of hydrolysates of cereulide and their time course profile. Bioorg. Med. Chem. Lett. 2020, 30. [Google Scholar] [CrossRef] [PubMed]
- Ehling-Schulz, M.; Vukov, N.; Schulz, A.; Shaheen, R.; Andersson, M.; Martlbauer, E.; Scherer, S. Identification and partial characterization of the nonribosomal peptide synthetase gene responsible for cereulide production in emetic Bacillus cereus. Appl. Environ. Microbiol. 2005, 71, 105–113. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- ISO 7932:2004. Microbiology of Food and Animal Feeding Stuffs—Horizontal Method for the Enumeration of Presumptive Bacillus Cereus—Colony-Count Technique at 30 Degrees C; ISO: Geneva, Switzerland, 2004. [Google Scholar]
- Yossa, N.; Arce, G.; Huang, M.J.; Yin, L.; Brown, E.; Hammack, T. Factors of detection of Bacillus cereus strains in eye cream. Int. J. Cosmet. Sci. 2017, 39, 179–187. [Google Scholar] [CrossRef]
- Magnusson, M.; Christiansson, A.; Svensson, B. Bacillus cereus spores during housing of dairy cows: factors affecting contamination of raw milk. J. Dairy Sci. 2007, 90, 2745–2754. [Google Scholar] [CrossRef]
- Shaheen, R.; Svensson, B.; Andersson, M.A.; Christiansson, A.; Salkinoja-Salonen, M. Persistence strategies of Bacillus cereus spores isolated from dairy silo tanks. Food Microbiol. 2010, 27, 347–355. [Google Scholar] [CrossRef]
- Ouertani, A.; Chaabouni, I.; Mosbah, A.; Long, J.; Barakat, M.; Mansuelle, P.; Mghirbi, O.; Najjari, A.; Ouzari, H.I.; Masmoudi, A.S.; et al. Two new secreted proteases generate a casein-derived antimicrobial peptide in Bacillus cereus food born isolate leading to bacterial competition in milk. Front. Microbiol. 2018, 9, 1148. [Google Scholar] [CrossRef]
- Lakshmi, B.K.M.; Muni Kumar, D.; Hemalatha, K.P.J. Purification and characterization of alkaline protease with novel properties from Bacillus cereus strain S8. J. Genet. Eng. Biotechnol. 2018, 16, 295–304. [Google Scholar] [CrossRef]
- Hakovirta, J.R.; Prezioso, S.; Hodge, D.; Pillai, S.P.; Weigel, L.M. Identification and analysis of informative single nucleotide polymorphisms in 16S rRNA gene sequences of the Bacillus cereus group. J. Clin. Microbiol. 2016, 54, 2749–2756. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Y.; Du, J.; Lai, Q.; Zeng, R.; Ye, D.; Xu, J.; Shao, Z. Proposal of nine novel species of the Bacillus cereus group. Int. J. Syst. Evol. Microbiol. 2017, 67, 2499–2508. [Google Scholar] [CrossRef] [PubMed]
- Wei, S.; Chelliah, R.; Park, B.J.; Park, J.H.; Forghani, F.; Park, Y.S.; Cho, M.S.; Park, D.S.; Oh, D.H. Molecular discrimination of Bacillus cereus group species in foods (lettuce, spinach, and kimbap) using quantitative real-time PCR targeting groEL and gyrB. Microb. Pathog. 2018, 115, 312–320. [Google Scholar] [CrossRef] [PubMed]
- Heini, N.; Stephan, R.; Ehling-Schulz, M.; Johler, S. Characterization of Bacillus cereus group isolates from powdered food products. Int. J. Food Microbiol. 2018, 283, 59–64. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kindle, P.; Etter, D.; Stephan, R.; Johler, S. Population structure and toxin gene profiles of Bacillus cereus sensu lato isolated from flour products. FEMS Microbiol. Lett. 2019, 366. [Google Scholar] [CrossRef] [PubMed]
- Antolinos, V.; Fernández, P.S.; Ros-Chumillas, M.; Periago, P.M.; Weiss, J. Development of a high-resolution melting-based approach for efficient differentiation among Bacillus cereus group isolates. Foodborne Pathog. Dis. 2012, 9, 777–785. [Google Scholar] [CrossRef]
- Zhang, J.; Di, B.; Shan, H.; Liu, J.; Zhou, Y.; Chen, H.; Hu, L.; Wu, X.; Bai, Z. Rapid detection of Bacillus cereus using cross-priming amplification. J. Food Prot. 2019, 82, 1744–1750. [Google Scholar] [CrossRef]
- Wehrle, E.; Moravek, M.; Dietrich, R.; Burk, C.; Didier, A.; Martlbauer, E. Comparison of multiplex PCR, enzyme immunoassay and cell culture methods for the detection of enterotoxinogenic Bacillus cereus. J. Microbiol. Methods 2009, 78, 265–270. [Google Scholar] [CrossRef]
- Oltuszak-Walczak, E.; Walczak, P. PCR detection of cytK gene in Bacillus cereus group strains isolated from food samples. J. Microbiol. Methods 2013, 95, 295–301. [Google Scholar] [CrossRef]
- Fagerlund, A.; Ween, O.; Lund, T.; Hardy, S.P.; Granum, P.E. Genetic and functional analysis of the cytK family of genes in Bacillus cereus. Microbiology 2004, 150, 2689–2697. [Google Scholar] [CrossRef] [Green Version]
- Yu, P.; Yu, S.; Wang, J.; Guo, H.; Zhang, Y.; Liao, X.; Zhang, J.; Wu, S.; Gu, Q.; Xue, L.; et al. Bacillus cereus isolated from vegetables in China: incidence, genetic diversity, virulence genes, and antimicrobial resistance. Front. Microbiol. 2019, 10, 948. [Google Scholar]
- Yu, S.; Yan, L.; Wu, X.; Li, F.; Wang, D.; Xu, H. Multiplex PCR coupled with propidium monoazide for the detection of viable Cronobacter sakazakii, Bacillus cereus, and Salmonella spp. in milk and milk products. J. Dairy Sci. 2017, 100, 7874–7882. [Google Scholar] [CrossRef] [Green Version]
- Zhou, P.; Xie, G.; Liang, T.; Yu, B.; Aguilar, Z.; Xu, H. Rapid and quantitative detection of viable emetic Bacillus cereus by PMA-qPCR assay in milk. Mol. Cell. Probes 2019, 47, 101437. [Google Scholar] [CrossRef] [PubMed]
- Baida, G.; Budarina, Z.I.; Kuzmin, N.P.; Solonin, A.S. Complete nucleotide sequence and molecular characterization of hemolysin II gene from Bacillus cereus. FEMS Microbiol. Lett. 1999, 180, 7–14. [Google Scholar] [CrossRef] [PubMed]
- Ueda, S.; Yamaguchi, M.; Eguchi, K.; Iwase, M. Identification of cereulide-producing Bacillus cereus by nucleic acid chromatography and reverse transcription real-time PCR. Biocontrol. Sci. 2016, 21, 45–50. [Google Scholar] [CrossRef] [Green Version]
- Dietrich, R.; Fella, C.; Strich, S.; Martlbauer, E. Production and characterization of monoclonal antibodies against the hemolysin BL enterotoxin complex produced by Bacillus cereus. Appl. Environ. Microbiol. 1999, 65, 4470–4474. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dietrich, R.; Moravek, M.; Burk, C.; Granum, P.E.; Martlbauer, E. Production and characterization of antibodies against each of the three subunits of the Bacillus cereus nonhemolytic enterotoxin complex. Appl. Environ. Microbiol. 2005, 71, 8214–8220. [Google Scholar] [CrossRef] [Green Version]
- Didier, A.; Dietrich, R.; Gruber, S.; Bock, S.; Moravek, M.; Nakamura, T.; Lindback, T.; Granum, P.E.; Martlbauer, E. Monoclonal antibodies neutralize Bacillus cereus Nhe enterotoxin by inhibiting ordered binding of its three exoprotein components. Infect. Immun. 2012, 80, 832–838. [Google Scholar] [CrossRef] [Green Version]
- Takahashi, N.; Nagai, S.; Fujita, A.; Ido, Y.; Kato, K.; Saito, A.; Moriya, Y.; Tomimatsu, Y.; Kaneta, N.; Tsujimoto, Y.; et al. Discrimination of psychrotolerant Bacillus cereus group based on MALDI-TOF MS analysis of ribosomal subunit proteins. Food Microbiol. 2020, 91, 103542. [Google Scholar] [CrossRef]
- Ducrest, P.J.; Pfammatter, S.; Stephan, D.; Vogel, G.; Thibault, P.; Schnyder, B. Rapid detection of Bacillus ionophore cereulide in food products. Sci. Rep. 2019, 9, 5814. [Google Scholar] [CrossRef] [Green Version]
- Ueda, S.; Nakajima, H.; Iwase, M.; Shinagawa, K.; Kuwabara, Y. LC-MS analysis of the emetic toxin, cereulide, produced by Bacillus cereus. Biocontrol. Sci. 2012, 17, 191–195. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamaguchi, M.; Kawai, T.; Kitagawa, M.; Kumeda, Y. A new method for rapid and quantitative detection of the Bacillus cereus emetic toxin cereulide in food products by liquid chromatography-tandem mass spectrometry analysis. Food Microbiol. 2013, 34, 29–37. [Google Scholar] [CrossRef]
- Saxena, R.; Singh, R. MALDI-TOF MS and CD spectral analysis for identification and structure prediction of a purified, novel, organic solvent stable, fibrinolytic metalloprotease from Bacillus cereus B80. Biomed. Res. Int. 2015, 2015, 527015. [Google Scholar] [CrossRef] [PubMed]
- Stark, T.; Marxen, S.; Rutschle, A.; Lucking, G.; Scherer, S.; Ehling-Schulz, M.; Hofmann, T. Mass spectrometric profiling of Bacillus cereus strains and quantitation of the emetic toxin cereulide by means of stable isotope dilution analysis and HEp-2 bioassay. Anal. Bioanal. Chem. 2013, 405, 191–201. [Google Scholar] [CrossRef] [PubMed]
- Shinagawa, K.; Ueno, Y.; Hu, D.; Ueda, S.; Sugii, S. Mouse lethal activity of a HEp-2 vacuolation factor, cereulide, produced by Bacillus cereus isolated from vomiting-type food poisoning. J. Vet. Med. Sci. 1996, 58, 1027–1029. [Google Scholar] [CrossRef] [Green Version]
- Jeßberger, N.; Dietrich, R.; Bock, S.; Didier, A.; Märtlbauer, E. Bacillus cereus enterotoxins act as major virulence factors and exhibit distinct cytotoxicity to different human cell lines. Toxicon 2014, 77, 49–57. [Google Scholar] [CrossRef]
- Tran, S.L.; Ramarao, N. Bacillus cereus immune escape: A journey within macrophages. FEMS Microbiol. Lett. 2013, 347, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Zhang, H.; Zhang, E.; He, J.; Li, W.; Wei, J. Distribution of virulence genes of Bacillus cereus in several specimens. Chin. J. Zoonoses 2019, 35, 683–687. [Google Scholar]
- Lin, Y.; Ren, F.; Zhao, L.; Guo, H. Genotypes and the persistence survival phenotypes of Bacillus cereus isolated from UHT milk processing lines. Food Control. 2017, 82, 48–56. [Google Scholar] [CrossRef]
- Sánchez Chica, J.; Correa, M.M.; Aceves-Diez, A.E.; Rasschaert, G.; Heyndrickx, M.; Castañeda-Sandoval, L.M. Genomic and toxigenic heterogeneity of Bacillus cereus sensu lato isolated from ready-to-eat foods and powdered milk in day care centers in Colombia. Foodborne Pathog. Dis. 2020, 17, 340–347. [Google Scholar] [CrossRef] [Green Version]
- Saleh-Lakha, S.; Leon-Velarde, C.G.; Chen, S.; Lee, S.; Shannon, K.; Fabri, M.; Downing, G.; Keown, B. A study to assess the numbers and prevalence of Bacillus cereus and its toxins in pasteurized fluid milk. J. Food Prot. 2017, 80, 1085–1089. [Google Scholar] [CrossRef] [PubMed]
- Glasset, B.; Herbin, S.; Guillier, L.; Cadel-Six, S.; Vignaud, M.L.; Grout, J.; Pairaud, S.; Michel, V.; Hennekinne, J.A.; Ramarao, N.; et al. Bacillus cereus-induced food-borne outbreaks in France, 2007 to 2014: epidemiology and genetic characterisation. Euro Surveill. 2016, 21. [Google Scholar] [CrossRef] [PubMed]
- Yibar, A.; Çetinkaya, F.; Soyutemiz, E.; Yaman, G. Prevalence, enterotoxin production and antibiotic resistance of Bacillus cereus isolated from milk and cheese Kafkas. Univ. Vet. Fak. Derg. 2017, 23, 635–642. [Google Scholar]
- Ramarao, N.; Tran, S.L.; Marin, M.; Vidic, J. Advanced methods for detection of Bacillus cereus and its pathogenic factors. Sensors 2020, 20, 2667. [Google Scholar] [CrossRef]
- Gao, S.; Ni, C.; Huang, W.; Hao, H.; Jiang, H.; Lv, Q.; Zheng, Y.; Liu, P.; Kong, D.; Jiang, Y. The interaction between flagellin and the glycosphingolipid Gb3 on host cells contributes to Bacillus cereus acute infection. Virulence 2020, 11, 769–780. [Google Scholar] [CrossRef] [PubMed]
- Ramm, F.; Dondapati, S.K.; Thoring, L.; Zemella, A.; Wustenhagen, D.A.; Frentzel, H.; Stech, M.; Kubick, S. Mammalian cell-free protein expression promotes the functional characterization of the tripartite non-hemolytic enterotoxin from Bacillus cereus. Sci. Rep. 2020, 10, 2887. [Google Scholar] [CrossRef] [Green Version]
- Lee, N.K.; Kim, W.S.; Paik, H.D. Bacillus strains as human probiotics: characterization, safety, microbiome, and probiotic carrier. Food Sci. Biotechnol. 2019, 28, 1297–1305. [Google Scholar] [CrossRef]
- Duc le, H.; Hong, H.A.; Barbosa, T.M.; Henriques, A.O.; Cutting, S.M. Characterization of Bacillus probiotics available for human use. Appl. Environ. Microbiol. 2004, 70, 2161–2171. [Google Scholar] [CrossRef] [Green Version]
- Green, D.H.; Wakeley, P.R.; Page, A.; Barnes, A.; Baccigalupi, L.; Ricca, E.; Cutting, S.M. Characterization of two Bacillus probiotics. Appl. Environ. Microbiol. 1999, 65, 4288–4291. [Google Scholar] [CrossRef] [Green Version]
- Hong, H.A.; Duc le, H.; Cutting, S.M. The use of bacterial spore formers as probiotics. FEMS Microbiol. Rev. 2005, 29, 813–835. [Google Scholar] [CrossRef] [Green Version]
- Colombo, M.; Todorov, S.D.; Eller, M.; Nero, L.A. The potential use of probiotic and beneficial bacteria in the Brazilian dairy industry. J. Dairy Res. 2018, 85, 487–496. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harms, A.; Maisonneuve, E.; Gerdes, K. Mechanisms of bacterial persistence during stress and antibiotic exposure. Science 2016, 354. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Cui, Y.; Peng, W.; Huang, X.; Ma, L.; Zheng, M.; Ding, S.; Zhu, K. Prevalence of pathogens harboring mobile antimicrobial resistance genes and virulence factors in retail beef and mutton. FEMS Microbiol. Lett. 2020. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Modes of action of the toxins in B. cereus. (a) Non-hemolytic enterotoxin (Nhe) and hemolysin BL (Hbl) perforate the cell membrane. Nhe promotes the NLRP3 inflammasome and induces caspase-8 dependent apoptosis. (b) Cereulide induces destroyed mitochondrial membrane potential (MMP) and leads to hepatocyte damage. (c) CytK and Hemolysins otherwise do harm to the target membrane, leading to cell lysis and cell apoptosis in macrophages.
Figure 1.
Modes of action of the toxins in B. cereus. (a) Non-hemolytic enterotoxin (Nhe) and hemolysin BL (Hbl) perforate the cell membrane. Nhe promotes the NLRP3 inflammasome and induces caspase-8 dependent apoptosis. (b) Cereulide induces destroyed mitochondrial membrane potential (MMP) and leads to hepatocyte damage. (c) CytK and Hemolysins otherwise do harm to the target membrane, leading to cell lysis and cell apoptosis in macrophages.

Figure 2.
Development of the dairy industry in China and the world. (a) The production, consumption, import and export of milk and milk products in China from 2012–2019. Data from National Bureau of Statistics of China. (b) The consumption of liquid milk, milk powder and cheese in China, America, European Union, Japan, New Zealand and Russia. (c) Microbial outbreaks in raw milk and dairy products during the past century all over the world [64]. Red marked isolates were B. cereus spp. strains.
Figure 2.
Development of the dairy industry in China and the world. (a) The production, consumption, import and export of milk and milk products in China from 2012–2019. Data from National Bureau of Statistics of China. (b) The consumption of liquid milk, milk powder and cheese in China, America, European Union, Japan, New Zealand and Russia. (c) Microbial outbreaks in raw milk and dairy products during the past century all over the world [64]. Red marked isolates were B. cereus spp. strains.

Figure 3.
The risk assessment of Bacillus cereus in milk and milk products. (a) B. cereus and spores circularly contaminate human markets, dairy farm and dairy products. (b) The red column diagram (lower left corner) represents the positive rate of Bacillus cereus in seven groups of milk and milk products. (c) The China map shows the regional specificity of B. cereus contamination. Blue indicates no reliable data are found in these provinces. There are four different levels of contamination rate and the darker the color is, the rate of B. cereus is higher. In particular, the reddish, orange, yellow and pale pink color signify the rate of 50–40%, 40–30%, 30–20% and 20–5%, respectively.
Figure 3.
The risk assessment of Bacillus cereus in milk and milk products. (a) B. cereus and spores circularly contaminate human markets, dairy farm and dairy products. (b) The red column diagram (lower left corner) represents the positive rate of Bacillus cereus in seven groups of milk and milk products. (c) The China map shows the regional specificity of B. cereus contamination. Blue indicates no reliable data are found in these provinces. There are four different levels of contamination rate and the darker the color is, the rate of B. cereus is higher. In particular, the reddish, orange, yellow and pale pink color signify the rate of 50–40%, 40–30%, 30–20% and 20–5%, respectively.

Figure 4.
Detection techniques of B. cereus and its virulence factors. (a) The percentages of articles using each detection technique for B. cereus and its virulence factors. These techniques include cytotoxicity tests, EIA, LC-MS/MALDI-TOF and PCR. (b) The positive rate of toxin genes in Bacillus samples from pasteurized milk or milk powder in China.
Figure 4.
Detection techniques of B. cereus and its virulence factors. (a) The percentages of articles using each detection technique for B. cereus and its virulence factors. These techniques include cytotoxicity tests, EIA, LC-MS/MALDI-TOF and PCR. (b) The positive rate of toxin genes in Bacillus samples from pasteurized milk or milk powder in China.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
The prevalence of Bacillus cereus and its virulence factors from dairy products in China.
| Source | Region | Year | No. of B. cereus Isolates/ No. of Samples | Detection of Toxin Genes (%) | Reference | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| nheA | nheB | nheC | hblA | hblC | hblD | cesB | cytK | HlyⅡ | |||||
| Raw milk | Beijing | 2013–2014 | 92/306 | 100 | 100 | 100 | 79 | 79 | 79 | ND | ND | ND | [13] |
| Raw milk | Northeast China | 2017–2018 | 56/350 | ND | ND | ND | ND | ND | ND | ND | ND | ND | [77] |
| Pasteurized milk | Major cities in China, including Beijing, Nanchang, Chengdu, Hefei, Wuhan, Shanghai, et al. | 2011–2016 | 70/258 | 99 | 99 | 94 | 47 | 68 | 68 | 5 | 73 | 54 | [14] |
| Pasteurized milk | Wuhan | 2006 | 26/54 | 71.7 | 62 | 71.7 | 37 | 66.3 | 71.7 | ND | ND | ND | [80] |
| Ice cream | Wuhan | 2006 | 24/40 | ND | ND | ND | ND | ND | ND | ND | ND | ND | [66] |
| Milk powder | Wenzhou | 2015–2016 | 76/400 | 75 | 100 | 90.8 | 35.5 | 29.0 | 21.1 | ND | 44.7 | ND | [81] |
| Infant formula | Chinese markets | 2012–2013 | 74/513 | ND | ND | ND | ND | ND | ND | ND | ND | ND | [82] |
| Infant formula | Chinese markets | 2015 | 57/135 | 87.7 | 87.7 | 49.1 | 24.6 | 22.8 | 17.5 | 3.5 | 22.8 | ND | [83] |
| Infant formula | Liaoning | 2016 | 22/176 | 90.9 | 72.7 | 100 | 0 | 59.1 | 54.5 | ND | 68.2 | ND | [84] |
| Infant formula | Chinese markets | 2013–2015 | 33/401 | ND | ND | ND | ND | ND | ND | ND | ND | ND | [7] |
| Infant formula | Liaoning | 2016–2017 | 70/166 | ND | ND | ND | ND | ND | ND | ND | ND | ND | [76] |
| Infant formula | Yunnan | 2012–2016 | 71/605 | ND | ND | ND | ND | ND | ND | ND | ND | ND | [85] |
| Infant formula | Kunming | 2016 | 5/126 | ND | ND | ND | ND | ND | ND | ND | ND | ND | [86] |
| Infant formula and processing facility | Gansu | 2013–2014 | 31/183 | ND | ND | ND | ND | ND | ND | ND | ND | ND | [87] |
| Infant formula | Heilongjiang, Hebei, Henan, Hubei, Hunan, Jiangsu, Jiangxi, Guangdong | 2012 | 115/817 | ND | ND | ND | ND | ND | ND | ND | ND | ND | [88] |
| Dairy products | Heilongjiang, Jilin, Hebei, Henan, Guizhou | 2018–2019 | 54/500 | 94.4 | 94.4 | 100 | 57.4 | 68.5 | 16.7 | 11.1 | 75.9 | 53.7 | [12] |
Table 2.
The distribution of Bacillus toxins in dairy products in China and other countries.
| Toxin Genes (%) | Origin | Source | Year | Reference | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| nheA | nheB | nheC | hblA | hblC | hblD | cesB | cytK | hlyⅡ | ||||
| nhe 100 | hbl 78.3 | 1.1 | -- | -- | Beijing, China | Dairy farms | 2013–2014 | [13] | ||||
| 90.9 | 72.7 | 100 | 0 | 59.1 | 54.5 | -- | 68.2 | -- | Liaoning, China | Milk powder | 2016 | [84] |
| 87.2 | 81.6 | 86.4 | 36 | 38.4 | 38.4 | 3.2 | 36.8 | -- | Hebei, Hainan, Yunnan province, et al., China | Milk powder | 2019 | [129] |
| 74.1 | 88.9 | 100 | 55.6 | 77.8 | 0 | 48.2 | 33.3 | -- | China | UHT milk processing line | 2014–2015 | [130] |
| 84.1 | 89.9 | 84.1 | 59.4 | 44.9 | 53.6 | 2.9 | 53.6 | -- | Brazil | Dairy production chain | 2016 | [62] |
| nhe 100 | hbl 29.5 | 0 | 24.1 | -- | Colombia | Ready-to-eat food and milk | 2013 | [131] | ||||
| 76.5 | -- | -- | -- | 41.2 | -- | 0 | 5.9 | -- | Canada | Pasteurized milk | 2014–2015 | [132] |
| 96 | 99 | 100 | 44 | 40 | 44 | -- | 42 | 23 | France | FBO | 2007–2014 | [133] |
| 6.3 | 2.1 | 4.2 | 11.5 | 10.4 | 16.7 | 9.4 | 75 | -- | Ghana | Dairy farm | 2015 | [73] |
| 60 | 60 | 60 | 13 | 13 | 113 | -- | 75 | -- | Turkey | Milk and cheese | 2013 | [134] |
Note: FBO indicates foodborne outbreak; “—” represents no analytical tests was performed to identify specific toxin genes.
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Liu, X.-Y.; Hu, Q.; Xu, F.; Ding, S.-Y.; Zhu, K. Characterization of Bacillus cereus in Dairy Products in China. Toxins 2020, 12, 454. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins12070454
AMA Style
Liu X-Y, Hu Q, Xu F, Ding S-Y, Zhu K. Characterization of Bacillus cereus in Dairy Products in China. Toxins. 2020; 12(7):454. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins12070454
Chicago/Turabian StyleLiu, Xiao-Ye, Qiao Hu, Fei Xu, Shuang-Yang Ding, and Kui Zhu. 2020. "Characterization of Bacillus cereus in Dairy Products in China" Toxins 12, no. 7: 454. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins12070454
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.