Mycotoxins and the Enteric Nervous System


1
Department of Clinical Physiology, Faculty of Veterinary Medicine, University of Warmia and Mazury in Olsztyn, Oczapowskiego 13, 10-957 Olsztyn, Poland
2
Department of Veterinary Prevention and Feed Hygiene, Faculty of Veterinary Medicine, University of Warmia and Mazury in Olsztyn, Oczapowskiego Str. 13, 10-718 Olsztyn, Poland
3
Department of Clinical Diagnostics, Faculty of Veterinary Medicine, University of Warmia and Mazury in Olsztyn, Oczapowskiego 14, 10-957 Olsztyn, Poland
*
Author to whom correspondence should be addressed.
Toxins 2020, 12(7), 461; https://0-doi-org.brum.beds.ac.uk/10.3390/toxins12070461
Submission received: 5 June 2020
/
Revised: 16 July 2020
/
Accepted: 17 July 2020
/
Published: 19 July 2020
(This article belongs to the Special Issue Mycotoxins Occurence in Feed and Their Influence on Animal Health)
Abstract
:Mycotoxins are secondary metabolites produced by various fungal species. They are commonly found in a wide range of agricultural products. Mycotoxins contained in food enter living organisms and may have harmful effects on many internal organs and systems. The gastrointestinal tract, which first comes into contact with mycotoxins present in food, is particularly vulnerable to the harmful effects of these toxins. One of the lesser-known aspects of the impact of mycotoxins on the gastrointestinal tract is the influence of these substances on gastrointestinal innervation. Therefore, the present study is the first review of current knowledge concerning the influence of mycotoxins on the enteric nervous system, which plays an important role, not only in almost all regulatory processes within the gastrointestinal tract, but also in adaptive and protective reactions in response to pathological and toxic factors in food.
Keywords:
mycotoxins; enteric nervous system; gastrointestinal tract; mammals; animal pathology; intestines; toxins; feedKey Contribution: Mycotoxins contained in food affect the living organism, especially the gastrointestinal tract and the enteric nervous system. This impact may be multidirectional and depends not only on the chemical structure of the mycotoxin and mammal species studied, but also on the type of the enteric plexuses and segment of the digestive tract.
1. Introduction
Mycotoxins are a group of several biochemicals synthesized as secondary metabolites by various species of fungi [1]. They are commonly found in a wide range of agricultural products, such as cereals (maize, wheat, rye), fresh and dried fruits, grape juice, spices, herbs and many others [2,3,4]. Moreover, the presence of mycotoxins has also been observed in food products of animal origin and water [3,4,5,6]. Previous studies have shown that mycotoxins show multidirectional harmful effects on human and animal health. It is known that mycotoxins may act on many internal organs and systems, including, among others, nervous, reproductive and immunological systems, metabolic processes and endocrine glands [7].
This widespread occurrence of mycotoxins and their adverse effects demonstrate that these substances are a serious health and economic problem of the contemporary world and therefore, mycotoxins are the most widely studied biological toxins [5,6]. However, many aspects of mycotoxin activity on eukaryotic organisms are unknown. One lesser-known issue is the influence of these substances on the enteric nervous system (ENS).
Since mycotoxins are present in food and drinking water, the gastrointestinal (GI) tract is the part of the body that first comes into contact with these toxic factors [8]. A relatively large number of studies have described mycotoxin-induced morphological and functional changes in the GI tract, whose character depends on the type of mycotoxin, mammal species studied, as well as the degree and length of exposure to mycotoxins [9,10,11,12,13,14,15,16,17,18,19]. The most common effects of mycotoxins on the GI tract include inflammatory and necrotic changes, disturbances in secretory activity and metabolism of the enterocytes, damage to the intestinal barrier and dysfunction in intestinal absorption [10,11,16,20]. Unfortunately, the impact of mycotoxins on the ENS has been neglected in toxicological studies for many years. There are a few recent studies published which describe this aspect of mycotoxin activity. These reports have indicated that the ENS plays a crucial role in the regulation of the majority of gastrointestinal functions, takes part in adaptive and protective processes and is one of the first barriers against pathological and toxic factors in food [15,16,17,21,22] and may also be compromised by the harmful effects of mycotoxins. Therefore, this work is an attempt to summarize the influence of mycotoxins on the ENS. To better understand this influence, a short description of the organization of the ENS is needed.
2. Anatomy of the Enteric Nervous System
The enteric nervous system is a specific part of the autonomic nervous system. It is situated in the wall of the gastrointestinal tract from the esophagus to the rectum and is responsible for the majority of gastrointestinal activities [23]. In terms of the number of nerve cells, the ENS is the second largest (after the brain, and before the spinal cord) nervous structure in mammals, which may contain an estimated 200–500 million neurons [24,25,26]. For this reason, as well as due to the complicated structure and high autonomy, the ENS is often called the intestinal brain [24].
Millions of neurons comprising the ENS are grouped in the neuronal ganglia, which are interconnected with a dense network of nerve fibers and form ganglionated plexuses. The localization and number of these plexuses depend on the mammal species and the segment of the GI tract. In rodents, the ENS in the esophagus and stomach is built of two types of intramural ganglia. The first type, the myenteric ganglion, is located between longitudinal and circular muscle layers. Myenteric ganglia are interconnected with a dense network of nerve fibers and form the myenteric plexus [27,28,29,30]. The second type of intramural ganglia, the submucous ganglion, is located in the submucous layer, near the muscularis mucosae of the mucosal layer. Contrary to muscular ganglia, the nerves interconnected with the submucous ganglia are rather sparse. Therefore, submucous ganglia in the esophagus and stomach do not form plexus [31], although some authors have described submucous plexus in rodent esophagus and stomach [32,33]. However, the situation is different in the small and large intestines in rodents. Both types of enteric ganglia (myenteric and submucous) located in the same places as in the esophagus and stomach are interconnected with a dense network of nerves. Therefore, two kinds of plexuses (the myenteric plexus and submucous plexus) are described in the rodent intestine [34,35,36,37].
In large mammals, the organization of the ENS in the esophagus and stomach is similar to rodents [38,39,40], although some authors have described three types of the enteric plexuses (such as in the intestine—see below) in the porcine stomach [41]. The only exception are ruminants, in which only one type of the enteric ganglia (myenteric ganglia) has been described in the forestomach. These ganglia are located between longitudinal and circular muscular layers, interconnected with the dense nerve fibers and form myenteric plexus [42,43].
In turn, there are three types of the enteric ganglia, which form intramural plexuses in the small and large intestine of large mammal species (for example, in the pig) (Figure 1) [44,45,46]. The first of them is the myenteric plexus located (similarly to rodents) between the longitudinal and circular muscle layer [45,46]. Moreover, two types of submucous plexuses located in the submucous layer of the intestinal wall have been observed: outer submucous plexus—located in close association with the adjacent circular muscle layer (on its inner side) and the inner submucous plexus—positioned closer to the intestinal lumen, near the muscularis mucosae [47,48,49]. These plexuses are also often named after their discoverers. The myenteric plexus often called Auerbach’s plexus, the outer submucous plexus—Schabadash’s plexus, and the inner submucous plexus (in rodents—the submucous plexus)—Meissner’s plexus [50,51].
As regards the organization of the human ENS, the distribution of the nervous structures in the esophagus and stomach is similar to rodents and large mammal species [23,31,52,53]. In the human small and large intestines, the organization of the ENS is not quite clear. Previous publications have described four types of enteric plexuses. In addition to the above-mentioned myenteric, outer submucous and inner submucous plexuses, the presence of an intermediate submucous plexus (IMSP)—a ganglionated plexus located in the submucous layer between the outer and inner submucous plexus has been reported [54]. However, at present, three kinds of plexuses located similarly to the porcine intestine are described in the human small and large intestine. In addition to the myenteric plexus located between the longitudinal and circular muscle layers, they include the plexus submucosus externus (PSE) near the circular muscle layers (on its inner side) and plexus submucosus internus (PSI) located closer to the intestinal lumen [23,31,55,56,57,58,59]. Contrary to the porcine inner submucous plexus, PSI in the human intestine is multi-layered, which means the particular ganglia within this plexus are located at a different depth of the submucous layer [56]. Other publications have shown that submucosal ganglia in the human colon are disseminated throughout the submucosal layer with significant inter-individual differences [60].
In addition to the above-mentioned main types of enteric ganglia, previous studies conducted in various mammal species have also reported the presence of small scattered neuronal ganglia in the mucosal layer (mucosal ganglia) and between the longitudinal muscular layer and serosa (subserosal ganglia), as well as a ganglionated plexus within the muscularis mucosae [31,61].
Enteric neurons are characterized by a high degree of differentiation in terms of morphological, functional and electrophysiological properties [25]. Moreover, the enteric neurons are also highly diverse with regard to their ability to synthesize neuronal active substances. Apart from acetylcholine (the main neuromediator in the ENS), a wide range of other neuronal factors have been described in enteric nervous structures [25,62,63,64,65]. The most important neuronal factors include vasoactive intestinal polypeptide (VIP), substance P (SP), galanin (GAL), nitric oxide and calcitonin gene-related peptide (CGRP). These substances may act as neuromediators and/or neuromodulators and participate in many regulatory processes including, among others, intestinal motility, secretion in the GI tract, immunological processes, blood flow, sensory stimuli conduction, intestinal digestion and absorption [16,23,25,62,63,64,65]. It should be noted that several active substances have been noted in the enteric neurons. Their exact roles are often still not quite clear. It is also known that the roles of the particular neuronal factors in the regulation of the stomach and intestine activity may depend on the segment of the GI tract and animal species studied. Such a substance is GAL and its participation in the control of the intestinal motility. Previous studies have shown that GAL induces the contraction of the ileal smooth muscles in the rat, guinea-pig rabbit and pig [66], while in the canine ileum and stomach it shows relaxant effects [67]. A similar situation is observed in the case of SP, which strongly stimulates the contraction of intestinal muscles in the rat and dog, while in humans such activity is rather limited [68,69,70]. Moreover, one substance very often appears to be involved in various GI tract activities. For example, CGRP (which is known as a key factor in sensory and pain stimuli conduction within the GI tract [71,72]) may also participate in the regulation of intestinal motility, mesenteric and intramural blood flow, gastric secretion, absorption of the nutrients in the intestine and protective reactions [73,74,75,76,77]. A detailed discussion of the exact functions played by all neuronal factors located in the enteric neurons is almost impossible because new active substances and their roles in various species are still being discovered. However, the main functions connected with the GI tract of selected neuronal substances occurring in the ENS are presented in Table 1.
In addition to neurons, the ENS also includes numerous glial cells, which are called enteric glial cells (EGC) [141,142,143]. Glial cells in the gastrointestinal tract are generally characterized by small size, irregular or stellate shape and numerous processes which are in direct contact with neuronal cell bodies and nerve fibers. Based on previous studies, it is known that ECG may be divided into four major types, and classification of the EGC is similar to that used in the case of glial cells in the central nervous system [144,145]. The first type is “protoplasmic” glial cells (type I glial cells), which are located between neuronal cells in the enteric ganglia and their appearance resembles protoplasmic astrocytes in the brain. The second type of glial cells (type II glial cells) are “fibrous” glial cells, whose processes accompany the nerves connecting the enteric ganglia with each other. These cells are similar to fibrous glial cells located in the central nervous system. Moreover, mucosal glial cells (type III glial cells) located near nerve fibers in the mucosal layer and intramuscular glial cells (type IV glial cells) accompanying the nerve fibers in the muscular layer have been described in the gastrointestinal tract [144,145].
It was initially thought that glial cells are only structural support to neurons, but it is now known that EGC play multidirectional functions in the regulation of various aspects of the ENS and all gastrointestinal tract activities [146,147]. Primarily, they take part in processes connected with the development, protection and nutrition of the enteric neuronal cells [148,149]. They regulate growth, maturation and differentiation of the enteric neurons, and affect synthesis and release of neuromediators and/or neuromodulators, thus constituting a key factor in maintaining intraneuronal homeostasis [145,149,150,151].
Moreover, EGC (especially mucosal glia) are involved in activities of the intestinal barrier integrity and functions [152]. It is known that EGC synthesize a wide range of several substances, such as glial-derived neurotrophic factor, transforming growth factor-β1 and neurotrophins, and act on the intestinal epithelial cells through paracrine mechanism [152,153]. Experimental studies have also shown that in animals with genetic ablation of EGC, the intestinal epithelial layer loses its integrity and disturbances in vascularization appear and lead to severe inflammatory processes [154].
Enteric glial cells also have important functions during intestinal pathological states. They participate in immune cell modulation in a wide range of the intestinal diseases, including ulcerative colitis, Crohn’s disease and colorectal cancer [142]. During inflammatory processes, proliferation of EGC occurs [155]. Glial cells participate in the immune recognition of pathological stimuli and may act as antigen-presenting immune cells [156]. Moreover, an increase in the production of some cytokines, including, among others, interleukins (IL-1β and IL-6) [157,158,159], as well as nerve growth factor (NGF) [160], glial fibrillary acidic protein (GFAP) [161] and nitric oxide (NO) [162] in glial cells has been noted during inflammatory processes.
It is also known that enteric glial cells play important roles in the pathogenesis of neurodegenerative diseases, including Parkinson’s, Alzheimer’s and Creutzfeldt-Jakob diseases. They are considered to be a possible trigger point for neurodegenerative processes, which through the gut–brain axis may efficiently affect neurodegenerative processes in the central nervous system [143,144,145,146,147,148,149,150,151,152,153,154,155,156,157,158,159,160,161,162,163].
An important feature of enteric neurons is the ability to change their morphological, physiological and neurochemical properties under the impact of physiological and pathological factors [15,65,164]. Changes in the ENS have been described during growth and aging, diet changes, as well as various intestinal pathological processes, systemic diseases and the impact of toxic substances [15,65,164,165]. Changes in enteric neurons are a sign of the adaptive and protective reactions and contribute to homeostasis maintenance in the GI tract [164,165]. Moreover, such changes appearing under the impact of disease or toxic substances may be the first signs of subclinical pathological processes or toxicity [166]. Some studies have indicated that mycotoxins may affect the morphology and neurochemical character of the enteric neurons. The following is a short characterization of several mycotoxin-induced changes in the enteric nervous system.
3. Mycotoxins Affecting the Enteric Neurons
3.1. Deoxynivalenol
Deoxynivalenol (DON—molecular weight 296.31 g/mol), belongs to the trichothecene family and is a substance produced by Fusarium spp. [9]. It is commonly found in barley, oat, rye, corn and rice [167,168]. The signs of toxicity depend on the dose, mammal species and duration of exposure. The most frequent symptoms of toxicity with DON include loss of appetite, decreased body weight gain, neuroendocrine disorders, vomiting and diarrhea [169].
In the GI tract, toxicity with DON results in a wide range of histopathological changes, such as inflammatory infiltration, necrotic changes in the intestinal villi, edema of lamina propria, a decrease in the number of goblet cells in the jejunum and the ileum, intensification of apoptosis and degeneration of lymphoid cells in the GI tract [10,170]. These changes, together with DON-induced disturbances in the synthesis of many active substances produced by the gastrointestinal mucosa lead to the injury of the intestinal barrier and abnormal nutrient absorption [11].
Within the nervous system, DON-induced changes include abnormal synthesis of neuronal neurotransmitters and/or neuromodulators and in disturbances in neuronal activity [171,172]. Moreover, in neuronal cells, DON induces apoptosis, affects the cerebral lipid peroxidation and influences neuronal calcium homeostasis, and these disturbances in the neuronal cells may lead to anorexic actions [172,173].
During a study performed on male Wistar rats (Rattus novergicus) aged 21 days, the influence of relatively low doses of DON on the ENS was described [14]. In that experiment, DON in various doses (from 0.2 mg/kg of chow to 2 mg/kg of chow) was given for 42 days, and the ENS was studied using immunohistochemistry and microscopic analysis. It was shown that this mycotoxin does not affect the myenteric ganglia organization in the jejunum [14]. Between control animals and rats receiving DON there were no differences in the density of glial cells located in the myenteric plexus, or the population density of myenteric neurons. Moreover, DON did not change the density of particular subpopulations of the myenteric neurons, i.e., cholinergic and nitrergic neurons [14]. However, all concentrations of DON studied in the above-mentioned experiment caused a decrease in the area of the general population of the myenteric neuronal cells, as well as cholinergic and nitrergic cell neurons. Moreover, DON also decreased the area of gliocytes located in the myenteric plexus [14] and decreased the myenteric ganglia area. It should be noted that during the cited study, besides changes in the ENS, the animals did not show any other symptoms of toxicity, including a decrease in body weight, diarrhea, loss of appetite or changes in the oxidative status [14]. This indicates that changes in the ENS are the first symptoms of toxicity with low doses of DON.
3.2. T2 Toxin
T2 toxin (molecular weight 466.5 g/mol), similar to DON, belongs to the trichothecene family of toxins. It is mainly synthesized by Fusarium sporotrichioides, F. langsethiae, F. acuminatum and F. poae and is recognized as the most acutely toxic trichothecene [174]. The impact of T2 toxin on the GI system manifests itself by (among others) histopathological changes in the intestinal mucosal layer (even with low doses), disturbances in the intestinal barrier functionality, influence on the enzymatic activity of enteric cells and inhibition of mucin production [175,176,177,178].
T2 toxin also shows neurotoxic activity and exposure to this substance results in a wide range of neurological symptoms, such as ataxia, muscular weakness, anorexia, as well as pathological lesions in the brain with disturbances in the functioning of this organ [179,180,181]. The main mechanisms underpinning the neurotoxic properties of T2 toxin are connected with reactive oxygen species and oxidative stress, as well as with mitochondrial dysfunction (consisting of the inhibition of the mitochondrial membrane potential and intensification of apoptosis) [182].
The ENS was studied using immunofluorescence in an experiment in vivo performed on juvenile (8-week-old) female domestic pigs of the White Large Polish Breed subjected to oral administration of T2 toxin at the level of 12 µg/kg body weight/day for 42 days [15]. Significant changes in the neurochemical character of the enteric neurons and nerve fibers located in the GI tract wall were described in this study. The character of changes depended on the type of the enteric plexus and the intestinal segment. It was reported that the administration of T-2 toxin increases the number of enteric neurons containing VIP in the porcine stomach and duodenum. These changes concern both myenteric and submucous plexuses and they are more visible in the duodenum, especially in the myenteric and outer submucous plexuses [15]. The same study showed that T-2 toxin also increases the number of nerve fibers containing VIP located in the muscular and mucosal layers of the porcine stomach and duodenum [15]. As previously indicated (Table 1), VIP in one of the potent inhibitory factors in the ENS and causes the hyperpolarization and relaxation of the gastrointestinal muscles and sphincters [132,133]. Moreover, VIP (as a vasodilator) increases blood flow in the wall of the GI tract and mesentery [132,134]. This substance may also affect the secretory activity of the GI tract, and the character of this activity depends on the GI tract segment [138,139,140]. It is known that VIP inhibits the gastric acid secretion in the stomach, but stimulates the secretion of the intestinal juice. VIP also has neuroprotective properties and increases the survivability of the enteric neurons [131]. Moreover, it is involved in immunological processes and shows anti-inflammatory properties. VIP also inhibits macrophages and inhibits the secretion of pro-inflammatory factors [135,136,137]. It is assumed that the increase in the number of VIP-positive enteric nervous structures under the impact of T2 toxin is connected with the protective and anti-inflammatory properties of VIP.
The influence of T2 toxin on the number of the enteric neurons containing cocaine and amphetamine-regulated transcript (CART) has been also reported [183]. In this study, T2 toxin was orally administrated to juvenile sows of the Large White Polish breed in a dose of 200 µg/kg of feed (the suggested permissible level of this toxin in the feed for pigs) for 42 days and the immunoreactivity in the ENS was evaluated using immunofluorescence. After the administration of T2 toxin, an increase in the number of CART-positive enteric neurons in all types of enteric plexuses as well as the number of nerve fibers containing CART in the mucosal and muscular layers in the stomach, duodenum and descending colon were described. The most visible changes were noted in the submucous plexus in the stomach and inner submucous plexus in the descending colon, where the number of CART-positive nerves under the impact of T2 toxin more than doubled [183]. It should be underlined that the exact functions of CART in the ENS are not clear [184]. A few studies concerning this issue have shown that CART inhibits the secretion of hydrochloric acid in the stomach and influences colonic motility [91,92]. This activity is probably done via the gut–brain axis because the direct impact of CART on isolated intestinal muscles does not cause changes in intestinal muscle contractility. The regulation of intestinal activity through the gut–brain axis is more likely since CART is known as an important factor regulating the feeding behavior in the central nervous system [185]. Moreover, numerous studies in which an increase in CART levels in the ENS has been observed strongly suggest that this peptide also takes part in protective and adaptive reactions in response to pathological, toxicological and physiological factors [46,166,184].
Another study (also performed with the immunofluorescence technique) concerning the impact of T2 toxin on the ENS was also performed on juvenile female pigs of the Large White Polish breed, which were treated with given T2 toxin orally in the dose of 12 µg/kg body weight/day for 42 days [16]. In this study, it was shown that T2 toxin affects the population of neurons containing calcitonin gene-related peptide located in the enteric plexuses in the porcine descending colon [16]. The administration of T2 toxin caused an increase in the number of CGRP-positive neurons in the myenteric, outer submucous and inner submucous plexuses, as well as an increase in the density of intramucosal nerves immunoreactive to these neuronal factors, without changes in the number of CGRP-positive nerve fibers in the muscular layer [16]. Moreover, it was shown that T2 toxin changes the neurochemical character of CGRP-positive neuronal cells, which were expressed by fluctuations in the degree of co-localization of CGRP with other neuronal factors (including substance P, nitric oxide synthase, galanin, CART peptide and vesicular acetylcholine transporter) in the same enteric nervous structures [16].
CGRP is a substance which primarily occurs in sensory neurons and is involved in sensory and pain stimuli conduction [71,72,93,94,95]. Moreover, CGRP in the GI tract takes part in the regulation of intestinal motility and increases blood flow in the mesenteric vessels [94,96,97,98,99]. It is also known that CGRP inhibits gastric acid secretion in the stomach with simultaneous induction of somatostatin release and regulates the absorption of nutrients from the intestine [186]. Previous studies have also shown that CGRP takes part in inflammatory processes in the intestine [99,100,101]. The multidirectional functions of CGRP in the ENS appear to be confirmed by a wide range of other neuronal substances present in CGRP-positive enteric neurons (such as substance P, nitric oxide CART peptide and galanin) which also play various roles in the GI tract (Table 1).
These reports of the influence of T2 toxin on the expression of a wide range of neuronal factors responsible for various regulatory processes in the ENS [15,16,183], strongly suggest that even relatively low doses of this mycotoxin may influence various intestinal activities, such as motility, secretion, conduction of sensory stimuli and regulation of the blood flow in the intestinal wall [15,16,183].
3.3. Zearalenon
Zearalenon (ZEN—molecular weight 318.364 g/mol) is synthesized mainly by Fusarium graminearum, culmorum, crookwellense and roseum and is found in barley, oat, wheat and bread [187]. The toxicity of ZEN is connected with its chemical structure, which allows it to act on the estrogen receptors, which are present in many internal organs [8]. ZEN can cross the blood-brain barrier and may influence neurons in the central nervous system [188,189]. It has been shown that exposure to ZEN leads to the abnormal synthesis of neuronal factors and enzymes in the brain neurons, induces apoptosis of the neuronal cells, increases oxidative stress reactions, influences the development of the nervous system, may cause behavioral aberrations and affects glial cell functions [189,190,191,192]. In turn, in the GI system, ZEN (among others) disturbs intestinal homeostasis, changes intestinal microbiome, causes inflammatory cell proliferation and inflammation in the intestinal mucosal layer [11,193,194,195].
Although the impact of ZEN on the GI tract is relatively well known, studies concerning the influence of this mycotoxin on intestinal innervation are limited to two studies performed on the pigs of the Large White Polish breed (approximately 8 weeks old), in which the nervous structure was evaluated with the immunofluorescence technique [19,21].
These studies have shown that the administration of relatively low doses of ZEN—10 μg/kg body weight/day [19] or 0.1 mg/kg of chow/day [21], administered for 42 days affect the neurochemical coding of nerve fibers in the mucosal and muscular layers of the ileum. For the intramuscular nerves, these changes involved an increase in the number of fibers immunoreactive to CART, substance P, nitric oxide synthase, VIP and pituitary adenylate cyclase-activating peptide and a decrease in the number of fibers containing galanin [19]. In the mucosal layer, ZEN not only caused an increase in the number of nerve fibers containing SP and/or VIP, but also changed the morphology of these nerves [21]. In animals treated with ZEN, nerves immunoreactive to SP and/or VIP become thicker and more visible than in the control animals [21]. It should be underlined that all the above-mentioned neuronal factors play important multidirectional roles in the regulation of the intestinal activity both in physiological conditions as well as during pathological processes, the most important of which are listed in Table 1.
The impact of ZEN on the ENS in the porcine descending colon has also been reported. A study concerning this issue was performed on juvenile (8-week-old) female pigs of the Large White Polish breed, which were treated with a dose of ZEN at the level of 6 µg/kg b.w./day given orally for 42 days [16]. In this study, the ENS evaluation was conducted with the immunofluorescence technique. The impact of ZEN was similar to the influence of T2 toxin. ZEN increased the number of neurons containing CGRP (whose functions in the ENS are described in the subchapter concerning T2-toxin and presented in Table 1) in all types of the enteric plexuses located in the descending colon [16]. Moreover, ZEN-induced changes in the neurochemical character of CGRP-positive enteric neurons were also reported [16]. These changes consisted of an increase in the degree of co-localization of CGRP with other neuronal factors (including substance P, galanin, CART and nitric oxide synthase, which was used as a marker of neuron synthesized nitric oxide) in neurons within all types of the enteric plexuses and intramural nerve fibers [16]. The functions of the above-mentioned neuronal active substances are presented in Table 1.
3.4. Patulin
Patulin (PAT-molecular weight 154.12 g/mol) is produced by various species belonging to Penicillium, Aspergillus, Paecilomyces and Byssochlamys [196,197] and is present in fruits (especially in apples) and vegetables [196,197]. Previous studies have shown that exposure to patulin causes damage to the intestinal barrier and inflammatory processes in the GI tract and influences the gut microbiota and the production of the mucus by enterocytes [198,199]. The neurotoxic activity of PAT is also known. It causes damage to the DNA in brain neuronal cells, mitochondrial and lysosomal dysfunction, a reduction of ATP levels and intensification of oxidative stress reactions [200,201].
The influence of patulin on the enteric neurons has been the subject of only one study. This study was performed on the cell culture of the enteric neurons prepared from 2–3-month-old C57B6/J OlaHsd mice and included various methods, such as growth and viability testing, a cytotoxicity test, evaluation of calcium signaling, measurement of glucose content, neurite outgrowth measurement and a reactive oxygen species (ROS) test [202]. The enteric neurons were treated with P coprobium extract, which decreased their viability with a half-maximal effective concentration (EC50) of 1 ng/µL This study also showed that patulin affects excitability and glucose consumption of the enteric neurons, which results in a patulin-induced reduction of ATP levels and glucose concentration in the enteric neurons. It has been also reported that patulin causes disorders in calcium signaling in the enteric neurons and affects neuronal morphology, which results in a reduction of neurite outgrowth and total neurite mass [202].
3.5. Fumonisins
Fumonisins are synthesized by Fusarium proliferatum and Fusarium verticillioides and characterized by a high degree of toxicity [203]. Numerous types of these mycotoxins have been described, but the most toxicologically important are fumonisin B1 (molecular weight 721.838 g/mol), fumonisin B2 (molecular weight 705.83 g/mol) and fumonisin B3 (molecular weight 705.8 g/mol), due to their high levels in cereal grains and crop products [12,204]. Among the numerous internal organs and systems which may be affected by fumonisins, the nervous system is one of the most susceptible to the adverse effects of these mycotoxins. It is known that fumonisins may enhance neurodegenerative reactions and impair the developmental processes in neurons located in the central nervous system, and some studies have reported connections between exposure to these mycotoxins and the risk of neurodegenerative diseases, such as multiple sclerosis, Alzheimer’s disease and Parkinson’s disease [205,206]. Exposure to fumonisins also results in changes in the GI tract, which manifest as disturbances in intestinal absorption, changes in the enterocytes and abnormalities in the intestinal immunological processes leading to increased susceptibility to infections [20].
However, knowledge of the influence of fumonisins on the ENS is extremely limited. One study concerning this issue was performed using the immunohistochemistry method on male Wistar rats (Rattus novergicus), which were 21 days old [12]. This study showed that a mixture of fumonisin B1 and B2 added to food in doses of 1 and 3 mg/kg of body weight (i.e., in doses which may be present in “natural” conditions in the food of humans and animals) given for 63 days does not affect the organization of the myenteric plexus in the rat jejunum [12]. Such doses of fumonisins do not result in changes in the general number of myenteric plexus and the number of myenteric neurons causing nitric oxide synthase, which is a marker of structures synthesizing nitric oxide. However, some changes in the myenteric neurons were observed under the impact of the mentioned doses of fumonisins. These changes consisted of a reduction in the size (without changes in their morphology) of neurons located in the myenteric plexus and included both neurons immunoreactive to pan-neuronal marker HuCD and nitric oxide synthase. Suoza et al. (2014) [12] reported that fumonisins not only affect the development and growth of neurons in the central nervous system but may also influence these processes in the ENS.
The influence of fumonisins on the ENS in the rat duodenum and jejunum of adolescent (5-weeks-old) male Wistar rats was also studied by Rudyk et al. (2020), using the immunohistochemistry method and histomorphometric analysis [13]. A mixture of fumonisins B1 and B2 were administered in a dose of 90 mg/kg of body weight for 21 days. That study demonstrated that fumonisins influence the following parameters within myenteric and submucous plexuses: area, perimeter, mean Feret diameter, mean diameter and sphericity [13]. It was also found that the impact of fumonisins on the ENS depends on the segment of the GI tract and the type of the enteric plexus. Fumonisin-induced changes in the duodenum were less visible, concerned only the submucous plexus and consisted of a reduction of area and mean diameter of ganglia, while the other parameters in the submucous plexus and all parameters studied in the myenteric plexus were not subjected to change. In the jejunum, changes were noted in the myenteric and submucous plexuses and consisted of an increase in the sphericity of ganglia and a reduction of other parameters in both types of plexuses. Moreover, the most visible changes were noted in the myenteric plexus.
The mechanisms of the impact of fumonisins on the ENS are unknown, but they probably inhibit ceramide synthase—an enzyme contributing to sphingolipid synthesis [207].
4. Mycotoxin Consumption and Human Gastrointestinal Diseases
The multidirectional adverse effects of mycotoxins on the GI tract (Table 2) cause that exposure to these substances may result in various disturbances of the GI activity in humans. However, the common prevalence of mycotoxins in the human environment and food indicates that participation of these chemicals in the development of intestinal diseases in humans may be an important public health problem all over the world [208].
The impact of mycotoxins on the intestinal barrier functions, intestinal immunity, secretory activity and gut microflora, as well as their genotoxic/mutagenic and carcinogenic effects are mainly known from experimental studies (Table 2). Such studies do not always fully reflect the conditions of natural exposure to mycotoxins. The first problem is the dose of mycotoxins, which is very difficult to determine in the human diet [254,255]. The second, more important, problem is the fact that food may contain several or even a dozen mycotoxins at the same time. These mycotoxins may chemically interact with each other, which leads to changes in their toxic properties and bio-availability. In this case, synergistic interactions between mycotoxins is particularly dangerous [255,256]. For example, previous studies have shown that mixtures of ZEN and DON or DON, T2 and ZEN show higher toxicity than these individual mycotoxins [175,257]. Moreover, it is known that human food may also contain other active substances and contaminations, such as bacterial products, pesticides, phytotoxins, chemical contaminations and preservatives, which not only affect mycotoxin activity but may contribute to various disorders in the GI tract [258]. That is why it is so difficult to determine the effective participation of mycotoxins in the development of human gastrointestinal diseases.
A comparison of histopathological changes occurring in the GI tract during human gastrointestinal diseases and changes in the intestine caused by mycotoxins has shown that the negative development in the GI tract in both cases are similar [255]. This may suggest a correlation between a degree of exposure to mycotoxins and the risk of human gastrointestinal diseases, as well as the participation of mycotoxins in the development of various diseases, including inflammatory bowel disease, Crohn’s disease, coeliac disease and colorectal cancer [255]. However, only comprehensive epidemiological studies on the relationships between mycotoxin levels in food, blood and urine and the occurrence of particular diseases conducted on a large human population would explain the connection between exposure to mycotoxins and the risk of human gastrointestinal diseases. Unfortunately, such studies are fragmentary and relatively few. These studies have reported that aflatoxins (especially aflatoxin B1) may pose a carcinogenic risk and exposure to these chemicals may increase the risk of gastric and colorectal cancer [259,260]. Other studies suggest a correlation between the exposure to ZEN and colorectal cancer [261], as well as relationships between exposure to aflatoxins and Crohn’s Disease, coeliac disease and ulcerative colitis [262]. Despite this, differences in concentration of patulin and citrinin in plasma and urine between healthy people and patients suffering from colorectal cancer have not been observed, which may suggest that these mycotoxins are not key factors leading to this disease [263].
5. Conclusions
Based on previous studies, it is known that mycotoxins affect the enteric nervous system (Table 3). This impact may be multidirectional and depends not only on the chemical structure of the mycotoxin and mammal species studied, but also on the type of the enteric plexuses and segment of the digestive tract. Mycotoxins may act on the size and morphological properties of intestinal nervous structures and the neurochemical character of the enteric neurons. These changes are probably a result of adaptive and protective reactions, which affect homeostasis maintenance. Moreover, mycotoxin-induced changes in the ENS are often the first sign of exposure to low doses of mycotoxins. Understanding the exact mechanisms connected with the influence of mycotoxins on the intestinal innervation may be very important in determining mycotoxin dose limits, which are safe and neutral for the living organism. Unfortunately, the current information about the influence of mycotoxins on the ENS is relatively limited and elucidation of all aspects connected with this issue requires further research.
Author Contributions
Conceptualization, S.G., M.G. and K.M.; supervision, S.G.; writing—original draft, S.G., M.G. and K.M.; writing—review and editing, S.G. All authors have read and agreed to the published version of the manuscript.
Funding
Project financially supported by Minister of Science and Higher Education in the range of the program entitled “Regional Initiative of Excellence” for the years 2019–2022: Project No. 010/RID/2018/19, amount of funding 12.000.000 PLN.
Conflicts of Interest
The authors declare that they have no conflict of interest.
References
- Cimbalo, A.; Alonso-Garrido, M.; Font, G.; Manyes, L. Toxicity of mycotoxins in vivo on vertebrate organisms: A review. Food Chem. Toxicol. 2020, 137, 111161. [Google Scholar] [CrossRef] [PubMed]
- De Ruyck, K.; De Boevre, M.; Huybrechts, I.; De Saeger, S. Dietary mycotoxins, co-exposure, and carcinogenesis in humans: Short review. Mutat. Res. 2015, 766, 32–41. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- González, N.; Marquès, M.; Nadal, M.; Domingo, J.L. Occurrence of environmental pollutants in foodstuffs: A review of organic vs. conventional food. Food Chem. Toxicol. 2019, 125, 370–375. [Google Scholar] [CrossRef] [PubMed]
- Gonkowski, S.; Obremski, K.; Makowska, K.; Rytel, L.; Mwaanga, E.S. Levels of Zearalenone and its metabolites in sun-dried kapenta fish and water of Lake Kariba in Zambi—A preliminary study. Sci. Total Environ. 2018, 637–638, 1046–1050. [Google Scholar] [CrossRef] [PubMed]
- Milićević, D.R.; Skrinjar, M.; Baltić, T. Real and perceived risks for mycotoxin contamination in foods and feeds: Challenges for food safety control. Toxins 2010, 2, 572–592. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alshannaq, A.; Yu, J.H. Occurrence, toxicity, and analysis of major mycotoxins in food. Int. J. Environ. Res. Public Health 2017, 14, 632. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rykaczewska, A.; Gajęcka, M.; Onyszek, E.; Cieplińska, K.; Dąbrowski, M.; Lisieska-Żołnierczyk, S.; Bulińska, M.; Babuchowski, A.; Gajęcki, M.T.; Zielonka, Ł. Imbalance in the blood concentrations of selected steroids in prepubertal gilts depending on the time of exposure to low doses of zearalenone. Toxins 2019, 11, 561. [Google Scholar] [CrossRef] [Green Version]
- Gajęcka, M.; Zielonka, Ł.; Gajęcki, M. Activity of zearalenone in the porcine intestinal tract. Molecules 2017, 22, 18. [Google Scholar] [CrossRef] [Green Version]
- Khoshal, A.K.; Novak, B.; Martin, P.G.P.; Jenkins, T.; Neves, M.; Schatzmayr, G.; Oswald, I.P.; Pinton, P. Co-Occurrence of DON and emerging mycotoxins in worldwide finished pig feed and their combined toxicity in intestinal cells. Toxins 2019, 11, 727. [Google Scholar] [CrossRef] [Green Version]
- Przybylska-Gornowicz, B.; Tarasiuk, M.; Lewczuk, B.; Prusik, M.; Ziółkowska, N.; Zielonka, Ł.; Gajęcki, M.; Gajęcka, M. The effects of low doses of two Fusarium toxins, zearalenone and deoxynivalenol, on the pig jejunum. A light and electron microscopic study. Toxins 2015, 7, 4684–4705. [Google Scholar] [CrossRef] [Green Version]
- Przybylska-Gornowicz, B.; Lewczuk, B.; Prusik, M.; Hanuszewska, M.; Petrusewicz-Kosińska, M.; Gajęcka, M.; Zielonka, Ł.; Gajęcki, M. The effects of deoxynivalenol and zearalenone on the pig large intestine. A light and electron microscopic study. Toxins 2018, 10, 148. [Google Scholar] [CrossRef] [Green Version]
- Sousa, F.C.; Schamber, C.R.; Amorin, S.S.; Natali, M.R. Effect of fumonisin-containing diet on the myenteric plexus of the jejunum in rats. Auton. Neurosci. 2014, 185, 93–99. [Google Scholar] [CrossRef] [Green Version]
- Rudyk, H.; Tomaszewska, E.; Arciszewski, M.B.; Muszyński, S.; Tomczyk-Warunek, A.; Dobrowolski, P.; Donaldson, J.; Brezvyn, O.; Kotsyumbas, I. Histomorphometrical changes in intestine structure and innervation following experimental fumonisins intoxication in male Wistar rats. Pol. J. Vet. Sci. 2020, 23, 77–88. [Google Scholar] [CrossRef] [PubMed]
- Rissato, D.F.; de Santi Rampazzo, A.P.; Borges, S.C.; Sousa, F.C.; Busso, C.; Buttow, N.C.; Natali, M.R.M. Chronic ingestion of deoxynivalenol-contaminated diet dose-dependently decreases the area of myenteric neurons and gliocytes of rats. Neurogastroenterol. Motil. 2020, 32, e13770. [Google Scholar] [CrossRef] [PubMed]
- Makowska, K.; Obremski, K.; Gonkowski, S. The impact of T-2 toxin on vasoactive intestinal polypeptide-like immunoreactive (VIP-LI) nerve structures in the wall of the porcine stomach and duodenum. Toxins 2018, 10, 138. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Makowska, K.; Obremski, K.; Zielonka, L.; Gonkowski, S. The influence of low doses of zearalenone and T-2 toxin on calcitonin gene related peptide-like immunoreactive (CGRP-LI) neurons in the ENS of the porcine descending colon. Toxins 2017, 9, 98. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alassane-Kpembi, I.; Pinton, P.; Oswald, I.P. Effects of mycotoxins on the intestine. Toxins 2019, 11, 159. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liew, W.P.; Mohd-Redzwan, S. Mycotoxin: Its impact on gut health and microbiota. Front. Cell Infect. Microbiol. 2018, 8, 60. [Google Scholar] [CrossRef] [Green Version]
- Gonkowski, S.; Obremski, K.; Calka, J. The influence of low doses of zearalenone on distribution of selected active substances in nerve fibers within the circular muscle layer of porcine ileum. J. Mol. Neurosci. 2015, 56, 878–886. [Google Scholar] [CrossRef] [Green Version]
- Bouhet, S.; Oswald, I. The intestine as a possible target for fumonisin toxicity. Mol. Nutr. Food Res. 2007, 51, 925–931. [Google Scholar] [CrossRef]
- Obremski, K.; Gonkowski, S.; Wojtacha, P. Zearalenone-induced changes in the lymphoid tissue and mucosal nerve fibers in the porcine ileum. Pol. J. Vet. Sci. 2015, 18, 357–365. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pinton, P.; Oswald, I.P. Effect of deoxynivalenol and other Type B trichothecenes on the intestine: A review. Toxins 2014, 6, 1615–1643. [Google Scholar] [CrossRef] [PubMed]
- Furness, J.B.; Callaghan, B.P.; Rivera, L.R.; Cho, H.J. The enteric nervous system and gastrointestinal innervation: Integrated local and central control. Adv. Exp. Med. Biol. 2014, 817, 39–71. [Google Scholar] [CrossRef] [PubMed]
- Gershon, M.D. The enteric nervous system: A second brain. Hosp. Pract. 1999, 34, 31–52. [Google Scholar] [CrossRef]
- Furness, J.B. Extrinsic and intrinsic sources of calcitonin gene-related peptide immunoreactivity in the lamb ileum: A morphometric and neurochemical investigation. Cell Tissue Res. 2006, 323, 183–196. [Google Scholar]
- Schneider, S.; Wright, C.M.; Heuckeroth, R.O. Unexpected roles for the second brain: Enteric nervous system as master regulator of bowel function. Annu. Rev. Physiol. 2019, 81, 235–259. [Google Scholar] [CrossRef]
- Morikawa, S.; Komuro, T. Distribution of myenteric NO neurons along the guinea-pig esophagus. J. Auton. Nerv. Syst. 1998, 74, 91–99. [Google Scholar] [CrossRef]
- Reiche, D.; Michel, K.; Pfannkuche, H.; Schemann, M. Projections and neurochemistry of interneurones in the myenteric plexus of the guinea-pig gastric corpus. Neurosci. Lett. 2000, 295, 109–112. [Google Scholar] [CrossRef]
- Zhang, G.Q.; Yang, S.; Li, X.S.; Zhou, D.S. Expression and possible role of IGF-IR in the mouse gastric myenteric plexus and smooth muscles. Acta Histochem. 2014, 116, 788–794. [Google Scholar] [CrossRef]
- Zimmermann, J.; Neuhuber, W.L.; Raab, M. Homer1 (VesL-1) in the rat esophagus: Focus on myenteric plexus and neuromuscular junction. Histochem. Cell Biol. 2017, 148, 189–206. [Google Scholar] [CrossRef]
- Furness, J.B. The Enteric Nervous System; Blackwell Publishing: Oxford, UK, 2006; pp. 1–274. [Google Scholar]
- Kamikawa, Y.; Shimo, Y. Pharmacological characterization of the opioid receptor in the submucous plexus of the guinea-pig oesophagus. Br. J. Pharmacol. 1983, 78, 693–699. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kunisawa, Y.; Komuro, T. Interstitial cells of Cajal associated with the submucosal plexus of the Guinea-pig stomach. Neurosci. Lett. 2008, 434, 273–276. [Google Scholar] [CrossRef] [PubMed]
- Heinicke, E.A.; Kiernan, J.A. An immunohistochemical study of the myenteric plexus of the colon in the rat and mouse. J. Anat. 1990, 170, 51–62. [Google Scholar] [PubMed]
- Sayegh, A.I.; Ritter, R.C. Morphology and distribution of nitric oxide synthase-, neurokinin-1 receptor-, calretinin-, calbindin-, and neurofilament-M-immunoreactive neurons in the myenteric and submucosal plexuses of the rat small intestine. Anat. Rec. A Discov. Mol. Cell. Evol. Biol. 2003, 271, 209–216. [Google Scholar] [CrossRef] [PubMed]
- Monro, R.L.; Bornstein, J.C.; Bertrand, P.P. Synaptic transmission from the submucosal plexus to the myenteric plexus in Guinea-pig ileum. Neurogastroenterol. Motil. 2008, 20, 1165–1173. [Google Scholar] [CrossRef]
- Li, J.P.; Zhang, T.; Gao, C.J.; Kou, Z.Z.; Jiao, X.W.; Zhang, L.X.; Wu, Z.Y.; He, Z.Y.; Li, Y.Q. Neurochemical features of endomorphin-2-containing neurons in the submucosal plexus of the rat colon. World J. Gastroenterol. 2015, 21, 9936–9944. [Google Scholar] [CrossRef]
- Rekawek, W.; Sobiech, P.; Gonkowski, S.; Żarczyńska, K.; Snarska, A.; Waśniewski, T.; Wojtkiewicz, J. Distribution and chemical coding patterns of cocaine- and amphetamine-regulated transcript-like immunoreactive (CART-LI) neurons in the enteric nervous system of the porcine stomach cardia. Pol. J. Vet. Sci. 2015, 18, 515–522. [Google Scholar] [CrossRef]
- Bulc, M.; Palus, K.; Całka, J.; Zielonka, Ł. Changes in immunoreactivity of sensory substances within the enteric nervous system of the porcine stomach during experimentally induced diabetes. J. Diabetes Res. 2018, 2018, 4735659. [Google Scholar] [CrossRef]
- Makowska, K.; Rytel, L.; Lech, P.; Osowski, A.; Kruminis-Kaszkiel, E.; Gonkowski, S. Cocaine- and amphetamine-regulated transcript (CART) peptide in the enteric nervous system of the porcine esophagus. Comptes Rendus Biol. 2018, 341, 325–333. [Google Scholar] [CrossRef]
- Kaleczyc, J.; Klimczuk, M.; Franke-Radowiecka, A.; Sienkiewicz, W.; Majewski, M.; Łakomy, M. The distribution and chemical coding of intramural neurons supplying the porcine stomach—The study on normal pigs and on animals suffering from swine dysentery. Anat. Histol. Embryol. 2007, 36, 186–193. [Google Scholar] [CrossRef]
- Teixeira, A.F.; Wedel, T.; Krammer, H.J.; Kühnel, W. Structural differences of the enteric nervous system in the cattle forestomach revealed by whole mount immunohistochemistry. Ann. Anat. 1998, 180, 393–400. [Google Scholar] [CrossRef]
- Arciszewski, M.B.; Barabasz, S.; Skobowiat, C.; Maksymowicz, W.; Majewski, M. Immunodetection of cocaine- and amphetamine-regulated transcript in the rumen, reticulum, omasum and abomasum of the sheep. Anat. Histol. Embryol. 2009, 38, 62–67. [Google Scholar] [CrossRef] [PubMed]
- Timmermans, J.P.; Barbiers, M.; Scheuermann, D.W.; Stach, W.; Adriaensen, D.; Mayer, B.; De Groodt-Lasseel, M.H. Distribution pattern, neurochemical features and projections of nitrergic neurons in the pig small intestine. Ann. Anat. 1994, 176, 515–525. [Google Scholar] [CrossRef]
- Makowska, K. Chemically induced inflammation and nerve damage affect the distribution of vasoactive intestinal polypeptide-like immunoreactive (VIP-LI) nervous structures in the descending colon of the domestic pig. Neurogastroenterol. Motil. 2018, 30, e13439. [Google Scholar] [CrossRef]
- Makowska, K.; Gonkowski, S. Age and sex-dependent differences in the neurochemical characterization of calcitonin gene-related peptide-like immunoreactive (CGRP-LI) nervous structures in the porcine descending colon. Int. J. Mol. Sci. 2019, 20, 1024. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kapp, S.; Schrödl, F.; Neuhuber, W.; Brehmer, A. Chemical coding of submucosal type V neurons in porcine ileum. Cells Tissues Organs 2006, 184, 31–41. [Google Scholar] [CrossRef]
- Gonkowski, S.; Całka, J. Changes in the somatostatin (SOM)-like immunoreactivity within nervous structures of the porcine descending colon under various pathological factors. Exp. Mol. Pathol. 2010, 88, 416–423. [Google Scholar] [CrossRef]
- Gonkowski, S. Substance P as a neuronal factor in the enteric nervous system of the porcine descending colon in physiological conditions and during selected pathogenic processes. Biofactors 2013, 39, 542–551. [Google Scholar] [CrossRef]
- Scheuermann, D.W.; Stach, W. Fluorescence microscopic study of the architecture and structure of an adrenergic network in the plexus myentericus (Auerbach), plexus submucosus externus (Schabadasch) and plexus submucosus internus (Meissner) of the porcine small intestine. Acta Anat. 1984, 119, 49–59. [Google Scholar] [CrossRef]
- Wakabayashi, K.; Takahashi, H.; Ohama, E.; Ikuta, F. Tyrosine hydroxylase-immunoreactive intrinsic neurons in the Auerbach’s and Meissner’s plexuses of humans. Neurosci. Lett. 1989, 96, 259–263. [Google Scholar] [CrossRef]
- Hwang, S.E.; Hieda, K.; Kim, J.H.; Murakami, G.; Abe, S.; Matsubara, A.; Cho, B.H. Region-specific differences in the human myenteric plexus: An immunohistochemical study using donated elderly cadavers. Int. J. Colorectal Dis. 2014, 29, 783–791. [Google Scholar] [CrossRef] [PubMed]
- Mandić, P.; Filipović, T.; Gasić, M.; Djukić-Macut, N.; Filipović, M.; Bogosavljević, I. Quantitative morphometric analysis of the myenteric nervous plexus ganglion structures along the human digestive tract. Vojnosanit. Pregl. 2016, 73, 559–565. [Google Scholar] [CrossRef] [PubMed]
- Ibba-Manneschi, L.; Martini, M.; Zecchi-Orlandini, S.; Faussone-Pellegrini, M.S. Structural organization of enteric nervous system in human colon. Histol. Histopathol. 1995, 10, 17–25. [Google Scholar] [PubMed]
- Wedel, T.; Roblick, U.; Gleiss, J.; Schiedeck, T.; Bruch, H.P.; Kühnel, W.; Krammer, H.J. Organization of the enteric nervous system in the human colon demonstrated by wholemount immunohistochemistry with special reference to the submucous plexus. Ann. Anat. 1999, 181, 327–337. [Google Scholar] [CrossRef]
- Brehmer, A.; Rupprecht, H.; Neuhuber, W. Two submucosal nerve plexus in human intestines. Histochem. Cell Biol. 2010, 133, 149–161. [Google Scholar] [CrossRef]
- Jabari, S.; de Oliveira, E.C.; Brehmer, A.; da Silveira, A.B. Chagasic megacolon: Enteric neurons and related structures. Histochem. Cell Biol. 2014, 142, 235–244. [Google Scholar] [CrossRef] [Green Version]
- Zetzmann, K.; Strehl, J.; Geppert, C.; Kuerten, S.; Jabari, S.; Brehmer, A. Calbindin D28k-immunoreactivity in human enteric neurons. Int. J. Mol. Sci. 2018, 19, 194. [Google Scholar] [CrossRef] [Green Version]
- Oponowicz, A.; Kozłowska, A.; Gonkowski, S.; Godlewski, J.; Majewski, M. Changes in the distribution of cocaine- and amphetamine-regulated transcript-containing neural structures in the human colon affected by the neoplastic process. Int. J. Mol. Sci. 2018, 19, 414. [Google Scholar] [CrossRef] [Green Version]
- Graham, K.D.; López, S.H.; Sengupta, R.; Shenoy, A.; Schneider, S.; Wright, C.M.; Feldman, M.; Furth, E.; Valdivieso, F.; Lemke, A.; et al. Robust, 3-Dimensional visualization of human colon enteric nervous system without tissue sectioning. Gastroenterology 2020, 158, 2221–2235.e5. [Google Scholar] [CrossRef]
- Crowe, R.; Burnstock, G. The subserosal ganglia of the human taenia. Neurosci. Lett. 1990, 119, 203–206. [Google Scholar] [CrossRef]
- Timmermans, J.P.; Scheuermann, D.W.; Stach, W.; Adriaensen, D.; De Groodt Lesseal, M.H.A. Functional morphology of the enteric nervous system with special reference to large mammals. Eur. J. Morphol. 1992, 30, 113–122. [Google Scholar]
- Timmermans, J.P.; Adriaensen, D.; Cornelissen, W.; Scheuermann, D.W. Structural organization and neuropeptide distribution in the mammalian enteric nervous system, with special attention to those components involved in mucosal reflexes. Comp. Biochem. Physiol. 1997, 118, 331–340. [Google Scholar] [CrossRef]
- Arciszewski, M.B.; Barabasz, S.; Całka, J. Expression of substance P, vasoactive intestinal peptide and galanin in cultured myenteric neurons from the ovine abomasum. Vet. Med. 2009, 3, 118–124. [Google Scholar] [CrossRef] [Green Version]
- Gonkowski, S.; Burliński, P.; Skobowiat, C.; Majewski, M.; Całka, J. Inflammation- and axotomy-induced changes in galanin-like immunoreactive (GAL-LI) nerve structures in the porcine descending colon. Acta Vet. Hung. 2010, 58, 91–103. [Google Scholar] [CrossRef] [PubMed]
- Botella, A.; Delvaux, M.; Frexinos, J.; Bueno, L. Comparative effects of galanin on isolated smooth muscle cells from ileum in five mammalian species. Life Sci. 1992, 50, 1253–1261. [Google Scholar] [CrossRef]
- Fox-Threlkeld, J.E.T.; McDonald, T.J.; Cipris, S.; Woskowska, Z.; Daniel, E.E. Galanin inhibition of vasoactive intestinal polypeptide release and circular muscle motility in the isolated perfused canine ileum. Gastroenterology 1991, 101, 1471–1476. [Google Scholar] [CrossRef]
- Lördal, M.; Johansson, C.; Hellström, P.M. Myoelectric pattern and effects on small bowel transit induced by the tachykinins neurokinin A, neurokinin B, substance P and neuropedtide K in the rat. Neurogastroenterol. Motil. 1993, 5, 33–40. [Google Scholar] [CrossRef]
- Lördal, M.; Theodorsson, E.; Hellström, P.M. Tachykinins influence interdigestive rhythm and contractile strength of human small intestine. Dig. Dis. Sci. 1997, 42, 1940–1949. [Google Scholar] [CrossRef]
- Thor, P.J.; Sendur, R.; Konturek, S.J. Influence of substance P on myoelectric activity of the small bowel. Am. J. Physiol. 1982, 243, G493–G496. [Google Scholar] [CrossRef]
- Roza, C.; Reeh, P.W.; Substance, P. calcitonin gene related peptide and PGE2 co-released from the mouse colon: A new model to study nociceptive and inflammatory responses in viscera, in vitro. Pain 2001, 93, 213–219. [Google Scholar] [CrossRef]
- Wolf, M.; Schrödl, F.; Neuhuber, W.; Brehmer, A. Calcitonin gene-related peptide: A marker for putative primary afferent neurons in the pig small intestinal myenteric plexus? Anat. Rec. 2007, 290, 1273–1279. [Google Scholar] [CrossRef] [PubMed]
- Lambrecht, N.; Burchert, M.; Respondek, M.; Muller, K.M.; Peskar, B.M. Role of calcitonin gene-related peptide and nitric oxide in the gastroprotective effect of capsaicin in the rat. Gastroenterology 1993, 104, 1371–1380. [Google Scholar] [CrossRef]
- Barada, K.A.; Saade, N.E.; Atweh, S.F.; Khoury, C.I.; Nassar, C.F. Calcitonin gene-related peptide regulates amino acid absorption across rat jejunum. Regul. Pept. 2000, 90, 39–45. [Google Scholar] [CrossRef]
- Leung, F.W.; Iwata, F.; Seno, K.; Leung, J.W. Acid-induced mesenteric hyperemia in rats: Role of CGRP, substance P, prostaglandin, adenosine, and histamine. Dig. Dis. Sci. 2003, 48, 523–532. [Google Scholar] [CrossRef]
- De Fontgalland, D.; Wattchow, D.A.; Costa, M.; Brookes, S.J.H. Immunohistochemical characterization of the innervation of human colonic mesenteric and submucosal blood vessels. Neurogastroenterol. Motil. 2008, 20, 1212–1226. [Google Scholar] [CrossRef]
- Kaiser, E.A.; Rea, B.J.; Kuburas, A.; Kovacevich, B.R.; Garcia-Martinez, L.F.; Recober, A.; Russo, A.F. Anti-CGRP antibodies block CGRP-induced diarrhea in mice. Neuropeptides 2017, 64, 95–99. [Google Scholar] [CrossRef] [PubMed]
- Delvalle, N.M.; Fried, D.E.; Rivera-Lopez, G.; Gaudette, L.; Gulbransen, B.D. Cholinergic activation of enteric glia is a physiological mechanism that contributes to the regulation of gastrointestinal motility. Am. J. Physiol. Gastrointest. Liver Physiol. 2018, 315, G473–G483. [Google Scholar] [CrossRef] [PubMed]
- Scheurer, U.; Drack, E.; Halter, F. Cyclooxygenase inhibitors affect Met-enkephalin- and acetylcholine-stimulated motility of the isolated rat colon. J. Pharmacol. Exp. Ther. 1985, 234, 742–746. [Google Scholar] [PubMed]
- Johnson, C.D.; Barlow-Anacker, A.J.; Pierre, J.F.; Touw, K.; Erickson, C.S.; Furness, J.B.; Epstein, M.L.; Gosain, A. Deletion of choline acetyltransferase in enteric neurons results in postnatal intestinal dysmotility and dysbiosis. FASEB J. 2018, 32, 4744–4752. [Google Scholar] [CrossRef] [Green Version]
- Aikawa, N.; Kishibayashi, N.; Karasawa, A.; Ohmori, K. The effect of zaldaride maleate, an antidiarrheal compound, on acetylcholine-induced intestinal electrolyte secretion. Biol. Pharm. Bull. 2000, 23, 1377–1378. [Google Scholar] [CrossRef] [Green Version]
- Ogata, H.; Podolsky, D.K. Trefoil peptide expression and secretion is regulated by neuropeptides and acetylcholine. Am. J. Physiol. 1997, 273, G348–G354. [Google Scholar] [CrossRef]
- Specian, R.D.; Neutra, M.R. Mechanism of rapid mucus secretion in goblet cells stimulated by acetylcholine. J. Cell Biol. 1980, 85, 626–640. [Google Scholar] [CrossRef] [PubMed]
- Hansen, L.; Lampert, S.; Mineo, H.; Holst, J.J. Neural regulation of glucagon-like peptide-1 secretion in pigs. Am. J. Physiol. Endocrinol. Metab. 2004, 287, E939–E947. [Google Scholar] [CrossRef] [Green Version]
- Al-Barazie, R.M.; Bashir, G.H.; Qureshi, M.M.; Mohamed, Y.A.; Al-Sbiei, A.; Tariq, S.; Lammers, W.J.; Al-Ramadi, B.K.; Fernandez-Cabezudo, M.J. Cholinergic activation enhances resistance to oral Salmonella infection by modulating innate immune defense mechanisms at the intestinal barrier. Front. Immunol. 2018, 9, 551. [Google Scholar] [CrossRef] [PubMed]
- Dhawan, S.; Hiemstra, I.H.; Verseijden, C.; Hilbers, F.W.; Te Velde, A.A.; Willemsen, L.E.; Stap, J.; den Haan, J.M.; de Jonge, W.J. Cholinergic receptor activation on epithelia protects against cytokine-induced barrier dysfunction. Acta Physiol. 2015, 213, 846–859. [Google Scholar] [CrossRef]
- Matteoli, G.; Boeckxstaens, G.E. The vagal innervation of the gut and immune homeostasis. Gut 2013, 62, 1214–1222. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- van der Zanden, E.P.; Boeckxstaens, G.E.; de Jonge, W.J. The vagus nerve as a modulator of intestinal inflammation. Neurogastroenterol. Motil. 2009, 21, 6–17. [Google Scholar] [CrossRef] [PubMed]
- Nijhuis, L.E.; Olivier, B.J.; de Jonge, W.J. Neurogenic regulation of dendritic cells in the intestine. Biochem. Pharmacol. 2010, 80, 2002–2008. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.S.; Fox-Threlkeld, J.E.; Furness, J.B. Innervation of intestinal arteries by axons with immunoreactivity for the vesicular acetylcholine transporter (VAChT). J. Anat. 1998, 192, 107–117. [Google Scholar] [CrossRef]
- Okumura, T.; Yamada, H.; Motomura, W.; Kohgo, Y. Cocaine- amphetamine-regulated transcript (CART) acts in the central nervous system to inhibit gastric acid secretion via brain corticotrophin-releasing factor system. Endocrinology 2000, 141, 2854–2860. [Google Scholar] [CrossRef]
- Tebbe, J.J.; Ortmann, E.; Schumacher, K.; Mönnikes, H.; Kobelt, P.; Arnold, R.; Schäffer, M.K.H. Cocaine- and amphetamine-regulated transcript stimulates colonic motility via central CRF receptor activation and peripheral cholinergic pathways in fed conscious rats. Neurogastroenterol. Motil. 2004, 16, 489–496. [Google Scholar] [CrossRef] [PubMed]
- Brehmer, A.; Croner, R.; Dimmler, A.; Papadopoulos, T.; Schrödl, F.; Neuhuber, W. Immunohistochemical characterization of putative primary afferent (sensory) myenteric neurons in human small intestine. Auton. Neurosci. 2004, 112, 49–59. [Google Scholar] [CrossRef] [PubMed]
- Grider, J.R. CGRP as a transmitter in the sensory pathway mediating peristaltic reflex. Am. J. Physiol. 1994, 266, G1139–G1145. [Google Scholar] [CrossRef] [PubMed]
- Kuramoto, H.; Kadowaki, M. Enhancement of CGRP sensory afferent innervation in the gut during the development of food allergy in an experimental murine model. Biochem. Biophys. Res. Commun. 2013, 430, 895–900. [Google Scholar]
- Holzer, P.; Guth, P.H. Neuropeptide control of rat gastric mucosal blood flow. Increase by calcitonin gene-related peptide and vasoactive intestinal polypeptide, but not substance P and neurokinin A. Circ. Res. 1991, 68, 100–105. [Google Scholar] [CrossRef] [Green Version]
- Bulut, K.; Felderbauer, P.; Deters, S.; Hoeck, K.; Schmidt-Choudhury, A.; Schmidt, W.E.; Hoffmann, P. Sensory neuropeptides and epithelial cell restitution: The relevance of SP- and CGRP-stimulated mast cells. Int. J. Colorectal Dis. 2008, 23, 535–541. [Google Scholar] [CrossRef]
- Tam, C.; Brain, S.D. The assessment of vasoactive properties of CGRP and adrenomedullin in the microvasculature: A study using in vivo and in vitro assays in the mouse. J. Mol. Neurosci. 2004, 22, 117–124. [Google Scholar] [CrossRef]
- Pawlik, W.W.; Obuchowicz, R.; Biernat, J.; Sendur, R.; Jaworek, J. Role of calcitonin gene related peptide in the modulation of intestinal circulatory, metabolic, and myoelectric activity during ischemia/reperfusion. J. Physiol. Pharmacol. 2000, 51, 933–942. [Google Scholar]
- Holzer, P. Role of visceral afferent neurons in mucosal inflammation and defense. Curr. Opin. Pharmacol. 2007, 7, 563–569. [Google Scholar] [CrossRef] [Green Version]
- Reinshagen, M.; Flämig, G.; Ernst, S.; Geerling, I.; Wong, H.; Walsh, J.H.; Eysselein, V.E.; Adler, G.J. Calcitonin gene-related peptide mediates the protective effect of sensory nerves in a model of colonic injury. Pharmacol. Exp. Ther. 1998, 286, 657–661. [Google Scholar]
- Bálint, A.; Fehér, E.; Kisfalvi, I.J.; Máté, M.; Zelles, T.; Vizi, E.S.; Varga, G. Functional and immunocytochemical evidence that galanin is a physiological regulator of human jejunal motility. J. Physiol. Paris 2001, 95, 129–135. [Google Scholar] [CrossRef]
- Matkowskyj, K.A.; Nathaniel, R.; Prasad, R.; Weihrauch, D.; Rao, M.; Benya, R.V. Galanin contributes to the excess colonic fluid secretion observed in dextran sulfate sodium murine colitis. Inflamm. Bowel Dis. 2004, 10, 408–416. [Google Scholar] [CrossRef]
- Piqueras, L.; Taché, Y.; Martinez, V. Galanin inhibits gastric acid secretion through a somatostatin-independent mechanism in mice. Peptides 2004, 25, 1287–1295. [Google Scholar] [CrossRef]
- Matkowskyj, K.; Royan, S.V.; Blunier, A.; Hecht, G.; Rao, M.; Benya, R.V. Age-dependent differences in galanin-dependent colonic fluid secretion after infection with Salmonella typhimurium. Gut 2009, 58, 1201–1206. [Google Scholar] [CrossRef] [PubMed]
- Daniel, E.E.; Haugh, C.; Woskowska, Z.; Cipris, S.; Jury, J.; Fox-Threlkeld, J.E.T. Role of nitric oxide—Related inhibition in intestinal function: Relation to vasoactive intestinal polypeptide. Am. J. Physiol. 1994, 266, 31–39. [Google Scholar] [CrossRef] [PubMed]
- Grider, J.R. Interplay of VIP and nitric oxide in regulation of the descending relaxation phase of peristalsis. Am. J. Physiol. 1993, 264, G334–G340. [Google Scholar] [CrossRef]
- Groneberg, D.; Voussen, B.; Friebe, A. Integrative control of gastrointestinal motility by nitric oxide. Curr. Med. Chem. 2016, 23, 2715–2735. [Google Scholar] [CrossRef]
- Walker, M.Y.; Pratap, S.; Southerland, J.H.; Farmer-Dixon, C.M.; Lakshmyya, K.; Gangula, P.R. Role of oral and gut microbiome in nitric oxide-mediated colon motility. Nitric Oxide 2018, 73, 81–88. [Google Scholar] [CrossRef]
- Mourad, F.H.; O’Donnell, L.J.; Andre, E.A.; Bearcroft, C.P.; Owen, R.A.; Clark, M.L.; Farthing, M.J. L-Arginine, nitric oxide, and intestinal secretion: Studies in rat jejunum in vivo. Gut 1996, 39, 539–544. [Google Scholar] [CrossRef] [Green Version]
- Izzo, A.A.; Mascolo, N.; Capasso, F. Nitric oxide as a modulator of intestinal water and electrolyte transport. Dig. Dis. Sci. 1998, 43, 1605–1620. [Google Scholar] [CrossRef]
- Mourad, F.H.; Barada, K.A.; Abdel-Malak, N.; Bou Rached, N.A.; Khoury, C.I.; Saade, N.E.; Nassar, C.F. Interplay between nitric oxide and vasoactive intestinal polypeptide in inducing fluid secretion in rat jejunum. J. Physiol. 2003, 550, 863–871. [Google Scholar] [CrossRef] [PubMed]
- Barry, M.K.; Alois, J.D.; Pickering, S.P.; Yeo, C.J. Nitric oxide modulates water and electrolyte transport in the ileum. Ann. Surg. 1994, 219, 382–388. [Google Scholar] [CrossRef]
- Fan, W.Q.; Smolich, J.J.; Wild, J.; Yu, V.Y.; Walker, A.M. Nitric oxide modulates regional blood flow differences in the fetal gastrointestinal tract. Am. J. Physiol. 1996, 271, G598–G604. [Google Scholar] [CrossRef]
- Jansson, L.; Carlsson, P.O.; Bodin, B.; Andersson, A.; Källskog, O. Neuronal nitric oxide synthase and splanchnic blood flow in anaesthetized rats. Acta. Physiol. Scand. 2005, 183, 257–262. [Google Scholar] [CrossRef] [PubMed]
- Roediger, W.E. Nitric oxide damage to colonocytes in colitis-by-association: Remote transfer of nitric oxide to the colon. Digestion 2002, 65, 191–195. [Google Scholar] [CrossRef]
- Varga, S.; Juhász, L.; Gál, P.; Bogáts, G.; Boro, M.; Palásthy, Z.; Szabó, A.; Kaszaki, J. Neuronal nitric oxide mediates the anti-inflammatory effects of intestinal ischemic preconditioning. J. Surg. Res. 2019, 244, 241–250. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Katsoulis, S.; Schmidt, W.E. Role of PACAP in the regulation of gastrointestinal motility. Ann. N. Y. Acad. Sci. 1996, 805, 364–378. [Google Scholar] [CrossRef] [PubMed]
- Ozawa, M.; Aono, M.; Moriga, M. Central effects of pituitary adenylate cyclase activating polypeptide (PACAP) on gastric motility and emptying in rats. Dig. Dis. Sci. 1999, 44, 735–743. [Google Scholar] [CrossRef] [PubMed]
- Felley, C.P.; Qian, J.M.; Mantey, S.; Pradhan, T.; Jensen, R.T. Chief cells possess a receptor with high affinity for PACAP and VIP that stimulates pepsinogen release. Am. J. Physiol. 1992, 263, G901–G907. [Google Scholar] [CrossRef]
- Läuff, J.M.; Modlin, I.M.; Tang, L.H. Biological relevance of pituitary adenylate cyclase-activating polypeptide (PACAP) in the gastrointestinal tract. Regul. Pept. 1999, 84, 1–12. [Google Scholar] [CrossRef]
- Kuwahara, A.; Kuwahara, Y.; Mochizuki, T.; Yanaihara, N. Action of pituitary adenylate cyclase-activating polypeptide on ion transport in guinea pig distal colon. Am. J. Physiol. 1993, 264, G433–G441. [Google Scholar] [CrossRef] [PubMed]
- Fuchs, M.; Adermann, K.; Raab, H.R.; Forssmann, W.G.; Kuhn, M. Pituitary adenylate cyclase-activating polypeptide: A potent activator of human intestinal ion transport. Ann. N. Y. Acad. Sci. 1996, 805, 640–647. [Google Scholar] [CrossRef] [PubMed]
- Wei, M.X.; Hu, P.; Wang, P.; Naruse, S.; Nokihara, K.; Wray, V.; Ozaki, T. Possible key residues that determine left gastric artery blood flow response to PACAP in dogs. World. J. Gastroenterol. 2010, 16, 4865–4870. [Google Scholar] [CrossRef] [PubMed]
- Shimizu, Y.; Matsuyama, H.; Shiina, T.; Takewaki, T.; Furness, J.B. Tachykinins and their functions in the gastrointestinal tract. Cell. Mol. Life Sci. 2008, 65, 295–311. [Google Scholar] [CrossRef]
- Shibata, C.; Sasaki, I.; Naito, H.; Ohtani, N.; Matsuno, S.; Mizumoto, A.; Iwanaga, Y.; Itoh, Z.; Tohoku, J. Effects of substance P on gastric motility differ depending on the sites and vagal innervation in conscious dogs. Exp. Med. 1994, 174, 119–128. [Google Scholar] [CrossRef] [Green Version]
- Donnerer, J.; Barthó, L.; Holzer, P.; Lembeck, F. Intestinal peristalsis associated with release of immunoreactive substance P. Neuroscience 1984, 11, 913–918. [Google Scholar] [CrossRef]
- Greenwood, B.; Doolittle, T.; See, N.A.; Koch, T.R.; Dodds, W.J.; Davison, J.S. Effects of substance P and vasoactive intestinal polypeptide on contractile activity and epithelial transport in the ferret jejunum. Gastroenterology 1990, 98, 1509–1517. [Google Scholar] [CrossRef]
- Perdue, M.H.; Galbraith, R.; Davison, J.S. Evidence for substance P as a functional neurotransmitter inguinea pig small intestinal mucosa. Regul. Pept. 1987, 18, 63–74. [Google Scholar] [CrossRef]
- Pothoulakis, C.; Castagliuolo, I.; LaMont, J.T.; Jaffer, A.; OKeane, J.C.; Snider, R.M.; Leeman, S.E. CP-96,345, a substance P antagonist, inhibits rat intestinal responses to Clostridium difficile toxin A but not choleratoxin. Proc. Natl. Acad. Sci. USA 1994, 91, 947–951. [Google Scholar] [CrossRef] [Green Version]
- Arciszewski, M.B.; Ekblad, E. Effects of vasoactive intestinal peptide and galanin on survival of cultured porcine myenteric neurons. Regul. Pept. 2005, 125, 185–192. [Google Scholar] [CrossRef]
- Eklund, S.; Jodal, M.; Lundgren, O.; Sjöqvist, A. Effects of vasoactive intestinal polypeptide on blood flow, motility and fluid transport in the gastrointestinal tract of the cat. Acta Physiol. Scand. 1979, 105, 461–468. [Google Scholar] [CrossRef] [PubMed]
- Krantis, A.; Mattar, K.; Glasgow, I. Rat gastroduodenal motility in vivo: Interaction of GABA and VIP in control of spontaneous relaxations. Am. J. Physiol. 1998, 275, G897–G903. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iwasaki, M.; Akiba, Y.; Kaunitz, J.D. Recent advances in vasoactive intestinal peptide physiology and pathophysiology: Focus on the gastrointestinal system. F1000Resrarch 2019, 8, F1000. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Talbot, J.; Hahn, P.; Kroehling, L.; Nguyen, H.; Li, D.; Littman, D.R. Feeding-dependent VIP neuron-ILC3 circuit regulates the intestinal barrier. Nature 2020, 579, 575–580. [Google Scholar] [CrossRef]
- Kovsca Janjatovic, A.; Valpotic, H.; Kezic, D.; Lacković, G.; Gregorovic, G.; Sladoljev, S.; Mršić, G.; Popovic, M.; Valpotic, I. Secretion of immunomodulating neuropeptides (VIP, SP) and nitric oxide synthase in porcine small intestine during postnatal development. Eur. J. Histochem. 2012, 56, e30. [Google Scholar] [CrossRef] [Green Version]
- Ottaway, C.A. Neuroimmunomodulation in the intestinal mucosa. Gastroenterol. Clin. N. Am. 1991, 20, 511–529. [Google Scholar]
- Burleigh, D.E.; Banks, M.R. Stimulation of intestinal secretion by vasoactive intestinal peptide and cholera toxin. Auton. Neurosci. Basic Clin. 2007, 133, 64–75. [Google Scholar] [CrossRef]
- Nassar, C.F.; Abdallah, L.E.; Barada, K.A.; Atweh, S.F.; Saadé, N.F. Effects of intravenous vasoactive intestinal peptide injection on jejunal alanine absorption and gastric acid secretion in rats. Regul. Pept. 1995, 55, 261–267. [Google Scholar] [CrossRef]
- Mourad, F.H.; Nassar, C.F. Effect of vasoactive intestinal polypeptide (VIP) antagonism on rat jejunal fluid and electrolyte secretion induced by cholera and Escherichia coli enterotoxins. Gut 2000, 47, 382–386. [Google Scholar] [CrossRef] [Green Version]
- Jessen, K.R.; Mirsky, R. Glial cells in the enteric nervous system contain glial fibrillary acidic protein. Nature 1980, 286, 736–737. [Google Scholar] [CrossRef]
- Pochard, C.; Coquenlorge, S.; Freyssinet, M.; Naveilhan, P.; Bourreille, A.; Neunlist, M.; Rolli-Derkinderen, M. The multiple faces of inflammatory enteric glial cells: Is Crohn’s disease a gliopathy? Am. J. Physiol. Gastrointest. Liver Physiol. 2018, 315, G1–G11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seguella, L.; Capuano, R.; Sarnelli, G.; Esposito, G. Play in advance against neurodegeneration: Exploring enteric glial cells in gut-brain axis during neurodegenerative diseases. Expert Rev. Clin. Pharmacol. 2019, 12, 555–564. [Google Scholar] [CrossRef] [PubMed]
- Gulbransen, B.D.; Sharkey, K.A. Novel functional roles for enteric glia in the gastrointestinal tract. Nat. Rev. Gastroenterol. Hepatol. 2012, 9, 625–632. [Google Scholar] [CrossRef]
- de Mattos Coelho-Aguiar, J.; Bon-Frauches, A.C.; Gomes, A.L.; Veríssimo, C.P.; Aguiar, D.P.; Matias, D.; Thomasi, B.B.; Gomes, A.S.; Brito, G.A.; Moura-Neto, V. The enteric glia: Identity and functions. Glia 2015, 63, 921–935. [Google Scholar] [CrossRef] [PubMed]
- Jessen, K.R. Glial cells. Int. J. Biochem. Cell Biol. 2004, 36, 1861–1867. [Google Scholar] [CrossRef] [PubMed]
- Grubišić, V.; Gulbransen, B.D. Enteric glia: The most alimentary of all glia. J. Physiol. 2017, 595, 557–570. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abdo, H.; Derkinderen, P.; Gomes, P.; Chevalier, J.; Aubert, P.; Masson, D.; Galmiche, J.P.; Vanden Berghe, P.; Neunlist, M.; Lardeux, B. Enteric glial cells protect neurons from oxidative stress in part via reduced glutathione. FASEB J. 2010, 24, 1082–1094. [Google Scholar] [CrossRef]
- Neunlist, M.; Rolli-Derkinderen, M.; Latorre, R.; Van Landeghem, L.; Coron, E.; Derkinderen, P.; De Giorgio, R. Enteric glial cells: Recent developments and future directions. Gastroenterology 2014, 147, 1230–1237. [Google Scholar] [CrossRef]
- Ruhl, A. Glial cells in the gut. Neurogastroenterol. Motil. 2005, 17, 777–790. [Google Scholar] [CrossRef]
- Boesmans, W.; Cirill, C.; Van den Abbeel, V.; Van den Haute, C.; Depoortere, I.; Tack, J.; Vanden Berghe, P. Neuro-transmitters involved in fast excitatory neurotransmission directly activate enteric glial cells. Neurogastroenterol. Motil. 2013, 25, e151–e160. [Google Scholar] [CrossRef] [Green Version]
- Vergnolle, N.; Cirillo, C. Neurons and glia in the enteric nervous system and epithelial barrier function. Physiology 2018, 33, 269–280. [Google Scholar] [CrossRef] [PubMed]
- Bauman, B.D.; Meng, J.; Zhang, L.; Louiselle, A.; Zheng, E.; Banerjee, S.; Roy, S.; Segura, B.J. Enteric glial-mediated enhancement of intestinal barrier integrity is compromised by morphine. J. Surg. Res. 2017, 219, 214–221. [Google Scholar] [CrossRef] [PubMed]
- Bush, T.G.; Savidge, T.C.; Freeman, T.C.; Cox, H.J.; Campbell, E.A.; Mucke, L.; Johnson, M.H.; Sofroniew, M.V. Fulminant jejuno-ileitis following ablation of enteric glia in adult transgenic mice. Cell 1998, 93, 189–201. [Google Scholar] [CrossRef] [Green Version]
- Joseph, N.M.; He, S.; Quintana, E.; Kim, Y.G.; Núñez, G.; Morrison, S.J. Enteric glia are multipotent in culture but primarily form glia in the adult rodent gut. J. Clin. Investig. 2011, 121, 3398–3411. [Google Scholar] [CrossRef]
- Da Silveira, A.B.; de Oliveira, E.C.; Neto, S.G.; Luquetti, A.O.; Fujiwara, R.T.; Oliveira, T.C.; Brehmer, A. Enteroglial cells act as antigen-presenting cells in chagasic megacolon. Hum. Pathol. 2011, 42, 522–553. [Google Scholar] [CrossRef]
- Rühl, A.; Franzke, S.; Collins, S.M.; Stremmel, W. Interleukin-6 expression and regulation in rat enteric glial cells. Am. J. Physiol. Gastrointest. Liver Physiol. 2001, 280, G1163–G1171. [Google Scholar] [CrossRef] [Green Version]
- Murakami, M.; Ohta, T.; Ito, S. Lipopolysaccharides enhance the action of bradykinin in enteric neurons via secretion of interleukin-1beta from enteric glial cells. J. Neurosci. Res. 2009, 87, 2095–2104. [Google Scholar] [CrossRef]
- Kermarrec, L.; Durand, T.; Gonzales, J.; Pabois, J.; Hulin, P.; Neunlist, M.; Neveu, I.; Naveilhan, P. Rat enteric glial cells express novel isoforms of Interleukine-7 regulated during inflammation. Neurogastroenterol. Motil. 2019, 31, e13467. [Google Scholar] [CrossRef] [Green Version]
- von Boyen, G.B.; Steinkamp, M.; Reinshagen, M.; Schäfer, K.H.; Adler, G.; Kirsch, J. Nerve growth factor secretion in cultured enteric glia cells is modulated by proinflammatory cytokines. J. Neuroendocrinol. 2006, 18, 820–825. [Google Scholar] [CrossRef]
- Rosenbaum, C.; Schick, M.A.; Wollborn, J.; Heider, A.; Scholz, C.S.; Cecil, A.; Niesler, B.; Hirrlinger, J.; Walles, H.; Metzger, M. Activation of myenteric glia during acute inflammation in vitro and in vivo. PLoS ONE 2016, 11, e0151335. [Google Scholar] [CrossRef]
- Cirillo, C.; Sarnelli, G.; Esposito, G.; Grosso, M.; Petruzzelli, R.; Izzo, G.; D’Armiento, C.F.P.; Rocco, A.; Nardone, G.; Iuvone, T.; et al. Increased mucosal nitric oxide production in ulcerative colitis is mediated in part by the enteroglial-derived S100B protein. Neurogastroenterol. Motil. 2009, 21, 1209-e112. [Google Scholar] [CrossRef]
- Caputi, V.; Giron, M.C. Microbiome-gut-brain axis and toll-like receptors in Parkinson’s disease. Int. J. Mol. Sci. 2018, 19, 1689. [Google Scholar] [CrossRef] [Green Version]
- Vasina, V.; Barbara, G.; Talamonti, L.; Stanghellini, V.; Corinaldesi, R.; Tonini, M.; De Ponti, F.; De Georgio, R. Enteric neuroplasticy evoked by inflammation. Auton. Neurosci. 2006, 126–127, 264–272. [Google Scholar] [CrossRef] [PubMed]
- Obata, Y.; Pachnis, V. The Effect of microbiota and the immune system on the development and organization of the enteric nervous system. Gastroenterology 2016, 151, 836–844. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Szymanska, K.; Gonkowski, S. Neurochemical characterization of the enteric neurons within the porcine jejunum in physiological conditions and under the influence of bisphenol A (BPA). Neurogastroenterol. Motil. 2019, 31, e13580. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Shao, H.; Luo, X.; Wang, R.; Li, Y.; Li, Y.; Luo, Y.; Chen, Z. Effect of ozone treatment on deoxynivalenol and wheat quality. PLoS ONE 2016, 11, e0147613. [Google Scholar] [CrossRef] [PubMed]
- Habrowska-Górczyńska, D.E.; Kowalska, K.; Urbanek, K.A.; Domińska, K.; Sakowicz, A.; Piastowska-Ciesielska, A.W. Deoxynivalenol modulates the viability, ROS production and apoptosis in prostate cancer cells. Toxins 2019, 11, 265. [Google Scholar] [CrossRef] [Green Version]
- Payros, D.; Alassane-Kpembi, I.; Pierron, A.; Loiseau, N.; Pinton, P.; Oswald, I.P. Toxicology of deoxynivalenol and its acetylated and modified forms. Arch. Toxicol. 2016, 90, 2931–2957. [Google Scholar] [CrossRef] [PubMed]
- Matejova, I.; Modra, H.; Blahova, J.; Franc, A.; Fictum, P.; Sevcikova, M.; Svobodova, Z. The effect of mycotoxin deoxynivalenol on haematological and biochemical indicators and histopathological changes in rainbow trout (Oncorhynchus Mykiss). BioMed Res. Int. 2014, 2014, 310680. [Google Scholar] [CrossRef] [Green Version]
- Faeste, C.K.; Pierre, F.; Ivanova, L.; Sayyari, A.; Massotte, D. Behavioural and metabolomic changes from chronic dietary exposure to low-level deoxynivalenol reveal impact on mouse well-being. Arch. Toxicol. 2019, 93, 2087–2102. [Google Scholar] [CrossRef]
- Girardet, C.; Bonnet, M.S.; Jdir, R.; Sadoud, M.; Thirion, S.; Tardivel, C.; Roux, J.; Lebrun, B.; Mounien, L.; Trouslard, J.; et al. Central inflammation and sickness-like behavior induced by the food contaminant deoxynivalenol: A PGE2-independent mechanism. Toxicol. Sci. 2011, 124, 179–191. [Google Scholar] [CrossRef] [PubMed]
- Tominaga, M.; Momonaka, Y.; Yokose, C.; Tadaishi, M.; Shimizu, M.; Yamane, T.; Oishi, Y.; Kobayashi-Hattori, K. Anorexic action of deoxynivalenol in hypothalamus and intestine. Toxicon 2016, 118, 54–60. [Google Scholar] [CrossRef]
- Hussein, H.S.; Brasel, J.M. Toxicity, metabolism, and impact of mycotoxins on humans and animals. Toxicology 2001, 167, 101–134. [Google Scholar] [CrossRef]
- Obremski, K.; Zielonka, L.; Gajecka, M.; Jakimiuk, E.; Bakuła, T.; Baranowski, M.; Gajecki, M. Histological estimation of the small intestine wall after administration of feed containing deoxynivalenol, T-2 toxin and zearalenone in the pig. Pol. J. Vet. Sci. 2008, 11, 339–345. [Google Scholar]
- Goossens, J.; Pasmans, F.; Verbrugghe, E.; Vandenbroucke, V.; De Baere, S.; Meyer, E.; Haesebrouck, F.; De Backer, P.; Croubels, S. Porcine intestinal epithelial barrier disruption by the Fusarium mycotoxins deoxynivalenol and T-2 toxin promotes transepithelial passage of doxycycline and paromomycin. BMC Vet. Res. 2012, 8, 245. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Osselaere, A.; Li, S.J.; De Bock, L.; Devreese, M.; Goossens, J.; Vandenbroucke, V.; Van Bocxlaer, J.; Boussery, K.; Pasmans, F.; Martel, A.; et al. Toxic effects of dietary exposure to T-2 toxin on intestinal and hepatic biotransformation enzymes and drug transporter systems in broiler chickens. Food Chem. Toxicol. 2013, 55, 150–155. [Google Scholar] [CrossRef]
- Lin, R.; Sun, Y.; Ye, W.; Zheng, T.; Wen, J.; Deng, Y. T-2 toxin inhibits the production of mucin via activating the IRE1/XBP1 pathway. Toxicology 2019, 424, 152230. [Google Scholar] [CrossRef]
- Sheng, K.; Lu, X.; Yue, J.; Gu, W.; Gu, C.; Zhang, H.; Wu, W. Role of neurotransmitters 5-hydroxy-tryptamine and substance P in anorexia induction following oral exposure to the trichothecene T-2 toxin. Food Chem. Toxicol. 2019, 123, 1–8. [Google Scholar] [CrossRef]
- Guo, P.; Liu, A.; Huang, D.; Wu, Q.; Fatima, Z.; Tao, Y.; Cheng, G.; Wang, X.; Yuan, Z. Brain damage and neurological symptoms induced by T-2 toxin in rat brain. Toxicol. Lett. 2018, 286, 96–107. [Google Scholar] [CrossRef]
- Chaudhary, M.; Rao, P.V. Brain oxidative stress after dermal and subcutaneous exposure of T-2 toxin in mice. Food Chem. Toxicol. 2010, 48, 3436–3442. [Google Scholar] [CrossRef]
- Dai, C.; Xiao, X.; Sun, F.; Zhang, Y.; Hoyer, D.; Shen, J.; Tang, S.; Velkov, T. T-2 toxin neurotoxicity: Role of oxidative stress and mitochondrial dysfunction. Arch. Toxicol. 2019, 93, 3041–3056. [Google Scholar] [CrossRef]
- Makowska, K.; Gonkowski, S.; Zielonka, L.; Dabrowski, M.; Calka, J. T2 toxin-induced changes in cocaine- and amphetamine-regulated transcript (CART)-like immunoreactivity in the enteric nervous system within selected fragments of the porcine digestive tract. Neurotox. Res. 2017, 31, 136–147. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Makowska, K.; Gonkowski, S. Cocaine- and amphetamine-regulated transcript (CART) peptide in mammals gastrointestinal system—A review. Ann. Anim. Sci. 2017, 1, 2–21. [Google Scholar] [CrossRef] [Green Version]
- Stanley, S.A.; Small, C.J.; Murphy, K.G.; Rayes, E.; Abbott, C.R.; Seal, L.J.; Morgan, D.G.; Sunter, D.; Dakin, C.L.; Kim, M.S.; et al. Actions of cocaine- and amphetamine-regulated transcript (CART) peptide on regulation of appetite and hypothalamo-pituitary axes in vitro and in vivo in male rats. Brain Res. 2001, 893, 186–194. [Google Scholar] [CrossRef]
- Helton, W.S.; Mulholland, M.M.; Bunnett, N.W.; Debas, H.T. Inhibition of gastric and pancreatic secretion in dogs by CGRP: Role of somatostatin. Am. J. Physiol. 1989, 256, G715–G720. [Google Scholar] [CrossRef] [PubMed]
- Knutsen, H.-K.; Alexander, J.; Barregård, L.; Bignami, M.; Brüschweiler, B.; Ceccatelli, S.; Cottrill, B.; Dinovi, M.; Edler, L.; Grasl-Kraupp, B.; et al. Risks for animal health related to the presence of zearalenone and its modified forms in feed. EFSA J. 2017, 15, 4851. [Google Scholar]
- Lephart, E.D.; Thompson, J.M.; Setchell, K.D.; Adlercreutz, H.; Weber, K.S. Phytoestrogens decrease brain calcium-binding proteins but do not alter hypothalamic androgen metabolizing enzymes in adult male rats. Brain Res. 2000, 859, 123–131. [Google Scholar] [CrossRef]
- Venkataramana, M.; Chandra Nayaka, S.; Anand, T.; Rajesh, R.; Aiyaz, M.; Divakara, S.T.; Murali, H.S.; Prakash, H.S.; Lakshmana Rao, P.V. Zearalenone induced toxicity in SHSY-5Y cells: The role of oxidative stress evidenced by N-acetyl cysteine. Food Chem. Toxicol. 2014, 65, 335–342. [Google Scholar] [CrossRef] [PubMed]
- Ren, Z.H.; Deng, H.D.; Deng, Y.T.; Deng, J.L.; Zuo, Z.C.; Yu, S.M.; Hu, Y.C. Effect of the Fusarium toxins, zearalenone and deoxynivalenol, on the mouse brain. Environ. Toxicol. Pharmacol. 2016, 46, 62–70. [Google Scholar] [CrossRef] [PubMed]
- Kiss, D.S.; Ioja, E.; Toth, I.; Barany, Z.; Jocsak, G.; Bartha, T.; Horvath, T.L.; Zsarnovszky, A. Comparative analysis of zearalenone effects on thyroid receptor alpha (TRα) and beta (TRβ) expression in rat primary cerebellar cell cultures. Int. J. Mol. Sci. 2018, 19, 1440. [Google Scholar] [CrossRef] [Green Version]
- Khezri, A.; Herranz-Jusdado, J.G.; Ropstad, E.; Fraser, T.W. Mycotoxins induce developmental toxicity and behavioural aberrations in zebrafish larvae. Environ. Pollut. 2018, 242, 500–506. [Google Scholar] [CrossRef] [PubMed]
- Gajęcka, M.; Stopa, E.; Tarasiuk, M.; Zielonka, Ł.; Gajęcki, M. The expression of type-1 and type-2 nitric oxide synthase in selected tissues of the gastrointestinal tract during mixed mycotoxicosis. Toxins 2013, 5, 2281–2292. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lahjouji, T.; Bertaccini, A.; Neves, M.; Puel, S.; Oswald, I.P.; Soler, L. Acute exposure to zearalenone disturbs intestinal homeostasis by modulating the Wnt/β-Catenin signaling pathway. Toxins 2020, 12, 113. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cieplińska, K.; Gajęcka, M.; Dąbrowski, M.; Rykaczewska, A.; Lisieska-Żołnierczyk, S.; Bulińska, M.; Zielonka, Ł.; Gajęcki, M.T. Time-dependent changes in the intestinal microbiome of gilts exposed to low zearalenone doses. Toxins 2019, 11, 296. [Google Scholar] [CrossRef] [Green Version]
- Artigot, M.P.; Loiseau, N.; Laffitte, J.; Mas-Reguieg, L.; Tadrist, S.; Oswald, I.P.; Puel, O. Molecular cloning and functional characterization of two CYP619 cytochrome P450s involved in biosynthesis of patulin in Aspergillus clavatus. Microbiology 2009, 155, 1738–1747. [Google Scholar] [CrossRef] [Green Version]
- Biango-Daniels, M.N.; Hodge, K.T. Paecilomyces rot: A New apple disease. Plant Dis. 2018, 102, 1581–1587. [Google Scholar] [CrossRef]
- Assunção, R.; Pinhão, M.; Loureiro, S.; Alvito, P.; João Silva, M. A Multi-endpoint approach to the combined toxic effects of patulin and ochratoxin a in human intestinal cells. Toxicol. Lett. 2019, 313, 120–129. [Google Scholar] [CrossRef]
- Mohan, H.M.; Collins, D.; Maher, S.; Walsh, E.G.; Winter, D.C.; O’Brien, P.J.; Brayden, D.J.; Baird, A.W. The mycotoxin patulin increases colonic epithelial permeability in vitro. Food. Chem. Toxicol. 2012, 50, 4097–4102. [Google Scholar] [CrossRef]
- Malekinejad, H.; Aghazadeh-Attari, J.; Rezabakhsh, A.; Sattari, M.; Ghasemsoltani-Momtaz, B. Neurotoxicity of mycotoxins produced in vitro by Penicillium roqueforti isolated from maize and grass silage. Hum. Exp. Toxicol. 2015, 34, 997–1005. [Google Scholar] [CrossRef]
- Vidal, A.; Ouhibi, S.; Gali, R.; Hedhili, A.; De Saeger, S.; De Boevre, M. The mycotoxin patulin: An updated short review on occurrence, toxicity and analytical challenges. Food. Chem. Toxicol. 2019, 129, 249–256. [Google Scholar] [CrossRef]
- Brand, B.; Stoye, N.M.; Guilherme, M.D.S.; Nguyen, V.T.T.; Baumgaertner, J.C.; Schüffler, A.; Thines, E.; Endres, K. Identification of patulin from Penicillium coprobium as a toxin for enteric neurons. Molecules 2019, 24, 2776. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peltomaa, R.; Vaghini, S.; Patiño, B.; Benito-Peña, E.; Moreno-Bondi, M.C. Species-specific optical genosensors for the detection of mycotoxigenic Fusarium fungi in food samples. Anal. Chim. Acta 2016, 935, 231–238. [Google Scholar] [CrossRef] [PubMed]
- Palencia, E.R.; Mitchell, T.R.; Snook, M.E.; Glenn, A.E.; Gold, S.; Hinton, D.M.; Riley, R.T.; Bacon, C.W. Analyses of black Aspergillus species of peanut and maize for ochratoxins and fumonisins. J. Food Prot. 2014, 77, 805–813. [Google Scholar] [CrossRef]
- Waes, J.G.; Starr, L.; Maddox, J.; Aleman, F.; Voss, K.A.; Wilberding, J.; Riley, R.T. Maternal fumonisin exposure and risk for neural tube defects: Mechanisms in an in vivo mouse model. Birth Defects Res. A Clin. Mol. Teratol. 2005, 73, 487–497. [Google Scholar] [CrossRef] [PubMed]
- Purzycki, C.B.; Shain, D.H. Fungal toxins and multiple sclerosis: A compelling connection. Brain Res. Bull. 2010, 82, 4–6. [Google Scholar] [CrossRef] [PubMed]
- Jenkins, G.R.; Tolleson, W.H.; Newkirk, D.K.; Roberts, D.W.; Rowland, K.L.; Saheki, T.; Kobayashi, K.; Howard, P.C.; Melchior, W.B., Jr. Identification of fumonisin B1 as an inhibitor of argininosuccinate synthetase using fumonisin affinity chromatography and in vitro kinetic studies. J. Biochem. Mol. Toxicol. 2000, 14, 320–328. [Google Scholar] [CrossRef]
- Kolf-Clauw, M.; Castellote, J.; Joly, B.; Bourges-Abella, N.; Raymond-Letron, I.; Pinton, P.; Oswald, I.P. Development of a pig jejunal explant culture for studying the gastrointestinal toxicity of the mycotoxin deoxynivalenol: Histopathological analysis. Toxicol. In Vitro 2009, 23, 1580–1584. [Google Scholar] [CrossRef]
- Pestka, J.J. Deoxynivalenol: Mechanisms of action, human exposure, and toxicological relevance. Arch. Toxicol. 2010, 84, 663–679. [Google Scholar] [CrossRef]
- Pestka, J.J.; Smolinski, A.T. Deoxynivalenol: Toxicology and potential effects on humans. J. Toxicol. Environ. Health B Crit. Rev. 2005, 8, 39–69. [Google Scholar] [CrossRef]
- Pestka, J.J.; Lin, W.S.; Miller, E.R. Emetic activity of the trichothecene 15-acetyldeoxynivalenol in swine. Food Chem. Toxicol. 1987, 25, 855–858. [Google Scholar] [CrossRef]
- Young, L.G.; McGirr, L.; Valli, V.E.; Lumsden, J.H.; Lun, A. Vomitoxin in corn fed to young pigs. J. Anim. Sci. 1983, 57, 655–664. [Google Scholar] [CrossRef] [PubMed]
- Prelusky, D.B.; Trenholm, H.L. The efficacy of various classes of anti-emetics in preventing deoxynivalenol-induced vomiting in swine. Nat. Toxins 1983, 1, 296–302. [Google Scholar] [CrossRef]
- Lee, H.J.; Ryu, D. Worldwide occurrence of mycotoxins in cereals and cereal-derived food products: Public health perspectives of their co-occurrence. J. Agric. Food Chem. 2017, 65, 7034–7051. [Google Scholar] [CrossRef]
- Pierron, A.; Mimoun, S.; Murate, L.S.; Loiseau, N.; Lippi, Y.; Bracarense, A.P.; Liaubet, L.; Schatzmayr, G.; Berthiller, F.; Moll, W.D.; et al. Intestinal toxicity of the masked mycotoxin deoxynivalenol-3-β-D-glucoside. Arch. Toxicol. 2016, 90, 2037–2046. [Google Scholar] [CrossRef] [PubMed]
- Alizadeh, A.; Braber, S.; Akbari, P.; Garssen, J.; Fink-Gremmels, J. Deoxynivalenol impairs weight gain and affects markers of gut health after low-dose, short-term exposure of growing pigs. Toxins 2015, 7, 2071–2095. [Google Scholar] [CrossRef]
- Bracarense, A.P.; Lucioli, J.; Grenier, B.; Pacheco, G.D.; Moll, W.D.; Schatzmayr, G.; Oswald, I.P. Chronic ingestion of deoxynivalenol and fumonisin, alone or in interaction, induces morphological and immunological changes in the intestine of piglets. Br. J. Nutr. 2012, 107, 1776–1786. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gerez, J.R.; Pinton, P.; Callu, P.; Grosjean, F.; Oswald, I.P.; Bracarense, A.P. Deoxynivalenol alone or in combination with nivalenol and zearalenone induce systemic histological changes in pigs. Exp. Toxicol. Pathol. 2015, 67, 89–98. [Google Scholar] [CrossRef]
- Kasuga, F.; Hara-Kudo, Y.; Saito, N.; Kumagai, S.; Sugita-Konishi, Y. In vitro effect of deoxynivalenol on the differentiation of human colonic cell lines Caco-2 and T84. Mycopathologia 1998, 142, 161–167. [Google Scholar] [CrossRef]
- Bensassi, F.; El Golli-Bennour, E.; Abid-Essefi, S.; Bouaziz, C.; Hajlaoui, M.R.; Bacha, H. Pathway of deoxynivalenol-induced apoptosis in human colon carcinoma cells. Toxicology 2009, 264, 104–109. [Google Scholar] [CrossRef] [PubMed]
- Kouadio, J.H.; Mobio, T.A.; Baudrimont, I.; Moukha, S.; Dano, S.D.; Creppy, E. Comparative study of cytotoxicity and oxidative stress induced by deoxynivalenol, zearalenone or fumonisin B1 in human intestinal cell line Caco-2. Toxicology 2005, 213, 56–65. [Google Scholar] [CrossRef]
- Awad, W.A.; Böhm, J.; Razzazi-Fazeli, E.; Zentek, J. Effects of feeding deoxynivalenol contaminated wheat on growth performance, organ weights and histological parameters of the intestine of broiler chickens. J. Anim. Physiol. Anim. Nutr. 2006, 90, 32–37. [Google Scholar] [CrossRef]
- Girish, C.K.; Smith, T.K. Effects of feeding blends of grains naturally contaminated with Fusarium mycotoxins on small intestinal morphology of turkeys. Poult. Sci. 2008, 87, 1075–1082. [Google Scholar] [CrossRef] [PubMed]
- Tremel, H.; Strugala, G.; Forth, W.; Fichtl, B. Dexamethasone decreases lethality of rats in acute poisoning with T-2 toxin. Arch. Toxicol. 1985, 57, 74–75. [Google Scholar] [CrossRef]
- Chi, M.S.; Robison, T.S.; Mirocha, C.J.; Reddy, K.R. Acute toxicity of 12,13-epoxytrichothecenes in one-day-old broiler chicks. Appl. Environ. Microbiol. 1978, 35, 636–640. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Richard, J.L. Some major mycotoxins and their mycotoxicoses—An overview. Int. J. Food Microbiol. 2007, 119, 3–10. [Google Scholar] [CrossRef]
- Pang, V.F.; Lorenzana, R.M.; Beasley, V.R.; Buck, W.B.; Haschek, W.M. Experimental T-2 toxicosis in swine. III. Morphologic changes following intravascular administration of T-2 toxin. Fundam. Appl. Toxicol. 1987, 8, 298–309. [Google Scholar] [CrossRef]
- Beasley, V.R.; Lundeen, G.R.; Poppenga, R.H.; Buck, W.B. Distribution of blood flow to the gastrointestinal tract of swine during T-2 toxin-induced shock. Fundam. Appl. Toxicol. 1987, 9, 588–594. [Google Scholar] [CrossRef]
- Ványi, A.; Glávits, R.; Gajdács, E.; Sándor, G.; Kovács, F. Changes induced in newborn piglets by the trichothecene toxin T-2. Acta. Vet. Hung. 1991, 39, 29–37. [Google Scholar] [PubMed]
- Bratich, P.M.; Buck, W.B.; Haschek, W.M. Prevention of T-2 toxin-induced morphologic effects in the rat by highly activated charcoal. Arch. Toxicol. 1990, 64, 251–253. [Google Scholar] [CrossRef] [PubMed]
- Kumagai, S.; Shimizu, T. Effects of fusarenon-X and T-2 toxin on intestinal absorption of monosaccharide in rats. Arch. Toxicol. 1988, 61, 489–495. [Google Scholar] [CrossRef] [PubMed]
- Thompson, W.L.; Wannemacher, R.W., Jr. In vivo effects of T-2 mycotoxin on synthesis of proteins and DNA in rat tissues. Toxicol. Appl. Pharmacol. 1990, 105, 483–491. [Google Scholar] [CrossRef]
- Gao, X.; Sun, L.; Zhang, N.; Li, C.; Zhang, J.; Xiao, Z.; Qi, D. Gestational zearalenone exposure causes reproductive and developmental toxicity in pregnant rats and female offspring. Toxins 2017, 9, 21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Obremski, K.; Gajęcka, M.; Zielonka, Ł.; Jakimiuk, E.; Gajęcki, M. Morphology and ultrastructure of small intestine mucosa in gilts with zearalenone mycotoxicosis. Pol. J. Vet. Sci. 2005, 8, 301–307. [Google Scholar] [PubMed]
- Obremski, K.; Poniatowska-Broniek, G. Zearalenone induces apoptosis and inhibits proliferation in porcine ileal Peyer’s patch lymphocytes. Pol. J. Vet. Sci. 2015, 18, 153–161. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Taranu, I.; Braicu, C.; Marin, D.E.; Pistol, G.C.; Motiu, M.; Balacescu, L.; Beridan Neagoe, I.; Burlacu, R. Exposure to zearalenone mycotoxin alters in vitro porcine intestinal epithelial cells by differential gene expression. Toxicol. Lett. 2015, 232, 310–325. [Google Scholar] [CrossRef] [PubMed]
- Girgis, G.N.; Barta, J.R.; Brash, M.; Smith, T.K. Morphologic changes in the intestine of broiler breeder pullets fed diets naturally contaminated with Fusarium mycotoxins with or without coccidial challenge. Avian Dis. 2010, 54, 67–73. [Google Scholar] [CrossRef]
- McKinley, E.R.; Carlton, W.W.; Boon, G.D. Patulin mycotoxicosis in the rat: Toxicology, pathology and clinical pathology. Food Chem. Toxicol. 1982, 20, 289–300. [Google Scholar] [CrossRef]
- Escoula, L.; More, J.; Baradat, C. The toxins by Byssochlamys nivea Westling. I. Acute toxicity of patulin in adult rats and mice. Ann. Rech. Vet. 1977, 8, 41–49. [Google Scholar]
- Speijers, G.J.; Franken, M.A.; van Leeuwen, F.X. Subacute toxicity study of patulin in the rat: Effects on the kidney and the gastro-intestinal tract. Food Chem. Toxicol. 1988, 26, 23–30. [Google Scholar] [CrossRef]
- McKinley, E.R.; Carlton, W.W. Patulin mycotoxicosis in Swiss ICR mice. Food Cosmet. Toxicol. 1980, 18, 181–187. [Google Scholar] [CrossRef]
- McKinley, E.R.; Carlton, W.W. Patulin mycotoxicosis in the Syrian hamster. Food Cosmet. Toxicol. 1980, 18, 173–179. [Google Scholar] [CrossRef]
- Dailey, R.E.; Brouwer, E.; Blaschka, A.M.; Reynaldo, E.F.; Green, S.; Monlux, W.S.; Ruggles, D.I. Intermediate-duration toxicity study of patulin in rats. J. Toxicol. Environ. Health 1977, 2, 713–725. [Google Scholar] [CrossRef] [PubMed]
- McLaughlin, J.; Lambert, D.; Padfield, P.J.; Burt, J.P.; O’Neill, C.A. The mycotoxin patulin, modulates tight junctions in caco-2 cells. Toxicol. In Vitro 2009, 23, 83–89. [Google Scholar] [CrossRef] [PubMed]
- Maidana, L.; Gerez, J.R.; El Khoury, R.; Pinho, F.; Puel, O.; Oswald, I.P.; Bracarense, A.P. Effects of patulin and ascladiol on porcine intestinal mucosa: An ex vivo approach. Food Chem. Toxicol. 2016, 98, 189–194. [Google Scholar] [CrossRef] [Green Version]
- Rotter, B.A.; Thompson, B.K.; Prelusky, D.B.; Trenholm, H.L.; Stewart, B.; Miller, J.D.; Savard, M.E. Response of growing swine to dietary exposure to pure fumonisin B1 during an eight-week period: Growth and clinical parameters. Nat. Toxins 1996, 4, 42–50. [Google Scholar] [CrossRef]
- Dilkin, P.; Zorzete, P.; Mallmann, C.A.; Gomes, J.D.; Utiyama, C.E.; Oetting, L.L.; Corrêa, B. Toxicological effects of chronic low doses of aflatoxin B(1) and fumonisin B(1)-containing Fusarium moniliforme culture material in weaned piglets. Food Chem. Toxicol. 2003, 41, 1345–1353. [Google Scholar] [CrossRef]
- Ledoux, D.R.; Brown, T.P.; Weibking, T.S.; Rottinghaus, G.E. Fumonisin toxicity in broiler chicks. J. Vet. Diagn. Investig. 1992, 4, 330–333. [Google Scholar] [CrossRef] [Green Version]
- Bhat, R.V.; Shetty, P.H.; Rao, P.A.; Rao, V.S. A foodborne disease outbreak due to the consumption of moldy sorghum and maize containing fumonisin mycotoxins. J. Toxicol. Clin. Toxicol. 1997, 35, 249–255. [Google Scholar] [CrossRef]
- Schmelz, E.M.; Dombrink-Kurtzman, M.A.; Roberts, P.C.; Kozutsumi, Y.; Kawasaki, T.; Merrill, A.H., Jr. Induction of apoptosis by fumonisin B1in HT29 cells is mediated by the accumulation of endogenous free sphingoid bases. Toxicol. Appl. Pharmacol. 1998, 148, 252–260. [Google Scholar] [CrossRef] [Green Version]
- Theumer, M.G.; Lopez, A.G.; Masih, D.T.; Chulze, S.N.; Rubinstein, H.R. Immunobiological effects of fumonisin B1in experimental subchronic mycotoxicoses in rats. Clin. Diagn. Lab. Immunol. 2002, 9, 149–155. [Google Scholar]
- Casado, J.M.; Theumer, M.; Masih, D.T.; Chulze, S.; Rubinstein, H.R. Experimental subchronic mycotoxicoses in mice: Individual and combined effects of dietary exposure to fumonisins and aflatoxin B1. Food Chem. Toxicol. 2001, 39, 579–586. [Google Scholar] [CrossRef]
- Bouhet, S.; Hourcade, E.; Loiseau, N.; Fikry, A.; Martinez, S.; Roselli, M.; Galtier, P.; Mengheri, E.; Oswald, I.P. The mycotoxin, fumonisin B1alters the proliferation and the barrier function of porcine intestinal epithelial cells. Toxicol. Sci. 2004, 77, 165–171. [Google Scholar] [CrossRef] [Green Version]
- Carballo, D.; Tolosa, J.; Ferrer, E.; Berrada, H. Dietary exposure assessment to mycotoxins through total diet studies. A review. Food Chem. Toxicol. 2019, 128, 8–20. [Google Scholar] [CrossRef] [PubMed]
- Maresca, M.; Fantini, J. Some food-associated mycotoxins as potential risk factors in humans predisposed to chronic intestinal inflammatory diseases. Toxicon 2010, 56, 282–294. [Google Scholar] [CrossRef] [PubMed]
- Capriotti, A.L.; Caruso, G.; Cavaliere, C.; Foglia, P.; Samperi, R.; Laganà, A. Multiclass mycotoxin analysis in food, environmental and biological matrices with chromatography/mass spectrometry. Mass Spectrom. Rev. 2012, 31, 466–503. [Google Scholar] [CrossRef] [PubMed]
- Chen, F.; Ma, Y.; Xue, C.; Ma, J.; Xie, Q.; Wang, G.; Bi, Y.; Cao, Y. The combination of deoxynivalenol and zearalenone at permitted feed concentrations causes serious physiological effects in young pigs. J. Vet. Sci. 2008, 9, 39–44. [Google Scholar] [CrossRef]
- Sergent, T.; Ribonnet, L.; Kolosova, A.; Garsou, S.; Schaut, A.; De Saeger, S.; Van Peteghem, C.; Larondelle, Y.; Pussemier, L.; Schneider, Y.J. Molecular and cellular effects of food contaminants and secondary plant components and their plausible interactions at the intestinal level. Food Chem. Toxicol. 2008, 46, 813–841. [Google Scholar] [CrossRef]
- Harrison, J.C.; Carvajal, M.; Garner, R.C. Does aflatoxin exposure in the United Kingdom constitute a cancer risk? Environ. Health Perspect. 1993, 99, 99–105. [Google Scholar] [CrossRef]
- Eom, S.Y.; Yim, D.H.; Zhang, Y.; Yun, J.K.; Moon, S.I.; Yun, H.Y.; Song, Y.J.; Youn, S.J.; Hyun, T.; Park, J.S.; et al. Dietary aflatoxin B1 intake, genetic polymorphisms of CYP1A2, CYP2E1, EPHX1, GSTM1, and GSTT1, and gastric cancer risk in Korean. Cancer Causes Control 2013, 24, 1963–1972. [Google Scholar] [CrossRef]
- Przybyłowicz, K.E.; Arłukowicz, T.; Danielewicz, A.; Morze, J.; Gajęcka, M.; Zielonka, Ł.; Fotschki, B.; Sawicki, T. Association between mycotoxin exposure and dietary habits in colorectal cancer development among a Polish population: A study protocol. Int. J. Environ. Res. Public Health 2020, 17, 698. [Google Scholar] [CrossRef] [Green Version]
- Roy, R.N.; Russell, R.I. Crohn’s Disease and Aflatoxins. J. R. Soc. Health 1992, 112, 277–279. [Google Scholar] [CrossRef] [PubMed]
- Ouhibi, S.; Vidal, A.; Martins, C.; Gali, R.; Hedhili, A.; De Saeger, S.; De Boevre, M. LC-MS/MS methodology for simultaneous determination of patulin and citrinin in urine and plasma applied to a pilot study in colorectal cancer patients. Food Chem. Toxicol. 2020, 136, 110994. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Organization of the enteric nervous system in the intestine of the domestic pig: MP—myenteric plexus, OSP—outer submucous plexus, ISP—inner submucous plexus, LML—longitudinal muscular layer, CML—circular muscular layer, SML—submucosal layer, ML—mucosal—layer.
Figure 1.
Organization of the enteric nervous system in the intestine of the domestic pig: MP—myenteric plexus, OSP—outer submucous plexus, ISP—inner submucous plexus, LML—longitudinal muscular layer, CML—circular muscular layer, SML—submucosal layer, ML—mucosal—layer.
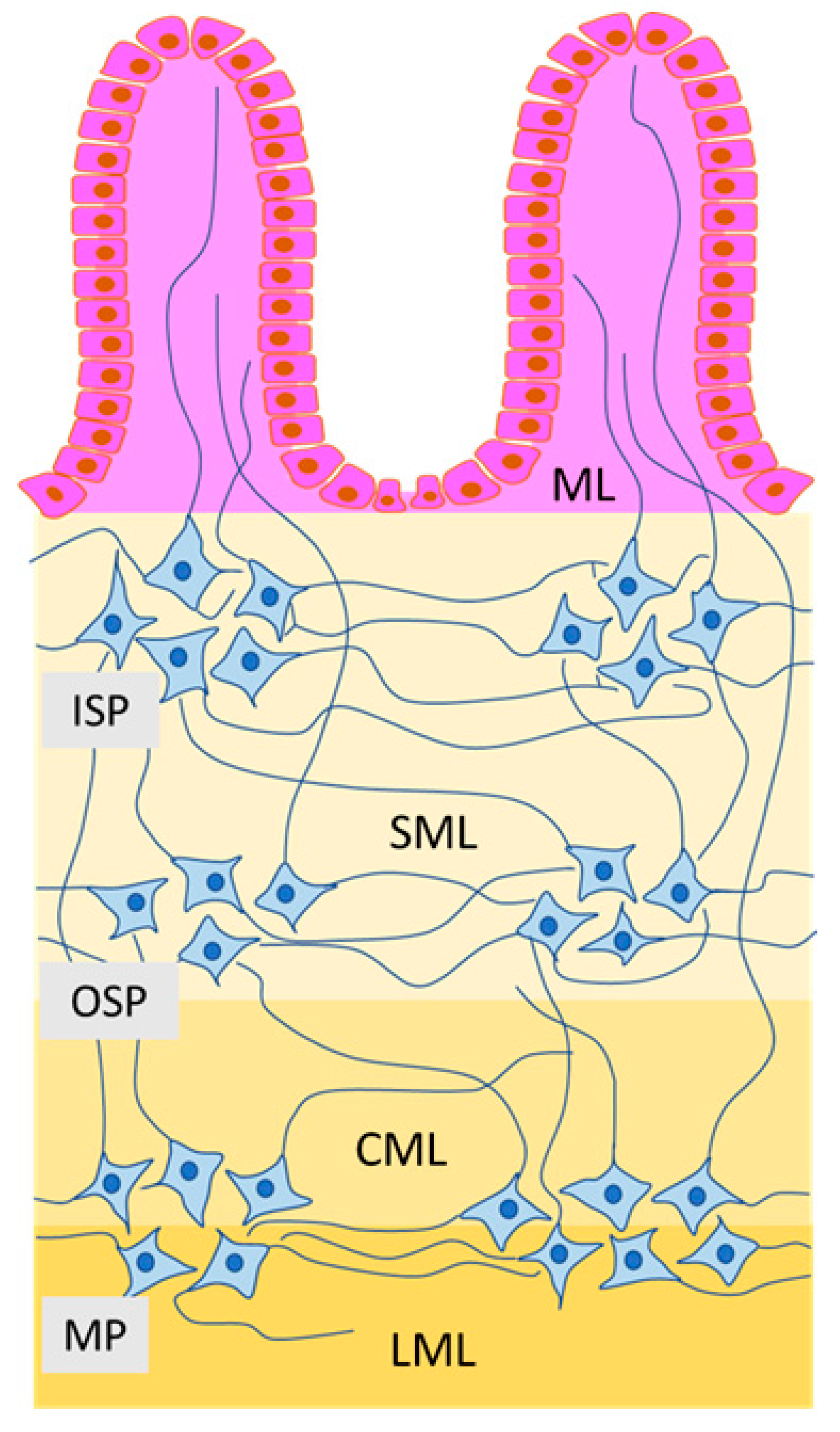

{kind=link}
Table 1.
Functions of selected active substances in the enteric nervous system.
| Active Neuronal Substance in the ENS (Alphabetical Order) | Selected Functions | References |
|---|---|---|
| Acetylcholine (Ach) | Stimulation of the intestinal motility | [78,79,80] |
| Stimulation of electrolyte, water, enzymes and hormones secretion | [81,82,83,84] | |
| Participation in protective mechanisms | [82,85,86] | |
| Ant-inflammatory and immunostymulatory effects | [87,88,89] | |
| Blood flow regulation | [90] | |
| Cocaine and Amphetamine Regulated Transcript (CART) | Inhibition of gastric acid secretion | [91] |
| Regulation of the intestinal motility | [92] | |
| Calcitonin Gene-Related Peptide (CGRP) | Participation in sensory and pain stimuli conduction | [71,72,93,94,95] |
| Regulation of the intestinal motility | [94] | |
| Blood flow regulation | [96,97,98,99] | |
| Protective roles | [73,99,100,101] | |
| The influence on intestinal absorption | [74] | |
| Galanin (GAL) | Intestinal motility regulation | [66,67,68,69,70,102] |
| Influence on secretory activity | [103,104,105] | |
| Participation in inflammatory processes | [103,105,106] | |
| Nitric Oxide (NO) | Inhibition of the intestinal motility | [106,107,108] |
| Participation in inflammatory processes | [109] | |
| Regulation of intestinal secretion, water and electrolyte transport | [110,111,112,113] | |
| Regulation of blood flow | [114,115] | |
| Participation in inflammatory processes | [116,117] | |
| Pituitary Adenylate Cyclase-Activating Polypeptide (PACAP) | Inhibition of the intestinal motility | [118,119] |
| Stimulation of gastric secretory activity | [120,121] | |
| Regulation of ion transport and Luminal fluid regulation in the large intestine | [121,122,123] | |
| Regulation of blood flow | [124] | |
| Substance P (SP) | Protective roles | [100] |
| Sensory stimuli conduction | [93,125] | |
| Regulation of the intestinal motility | [125,126,127] | |
| Regulation of water and electrolytes secretion | [125,128,129] | |
| Participation in inflammatory processes | [125,130] | |
| Vasoactive Intestinal Polypeptide (VIP) | Neuroprotective functions | [131] |
| Regulation of the intestinal motility | [132,133] | |
| Vasodialtory activity | [132,134] | |
| Participation in intestinal immunomodulation | [135,136,137] | |
| Influences on intestinal secretion | [138,139,140] |
Table 2.
Gastrointestinal signs and effects of mycotoxins on the gastrointestinal tract.
| Mycotoxin | Gastrointestinal Signs of Toxicity | References | Influence on the Digestive Tract | References |
|---|---|---|---|---|
| Doxynivalenol (DON) | Abdominal pain, increased salivation, diarrhea, vomiting, anorexia, decrease body weight gain | [169,209,210,211,212,213,214] | IPEC-J2 cell line from porcine jejunal epithelium: cytotoxicity, decrease in transepithelial electrical resistance, disruption of epithelial integrity | [176] |
| Porcine jejunal explant samples: shortened and coalescent villi, lysis of enterocytes, edema, upregulation of proinflammatory cytokines expression | [215,216] | |||
| Pigs of White Large Polish Breed: increase in the mucosal thickness and the intestinal crypt depth, atrophy of the villi, changes in the number of goblet cells, inflammatory infiltration, intensification of apoptosis, changes in ultrastructure of intestinal cells | [10,11,175,214,217,218] | |||
| Human Colonic Cell Lines Caco-2, T84, HT-29: decrease in cell proliferation, changes in permeability, genotoxicity, intensification of apoptosis, increase in the expression of proinflammatory cytokines, influence on DNA synthesis | [215,219,220,221] | |||
| Poultry: decrease in the high of villi | [222,223] | |||
| T2 Toxin | Gastrointestinal bleeding, diarrhea, vomiting, decreased feed consumption and weight gain | [224,225,226] | IPEC-J2 cell line from porcine jejunal epithelium: cytotoxic effects, disruption of intestinal barrier integrity | [176] |
| human intestinal Caco-2 cells disturbances in intestinal barrier, enzymatic activity of enteric cells, inhibition of mucin production | [178] | |||
| Pigs of White Large Polish Breed or crossbred pigs: congestion and hemorrhage of the gastrointestinal mucosal layer, inflammatory infiltration, in high doses—necrotic changes | [175,227,228,229] | |||
| Sprague-Daw-ley rats: inflammatory and necrotic changes in, lymphocytic necrosis in intestinal Peyer’s patches, influence on nutrients absorption, influence on DNA synthesis | [230,231,232] | |||
| Zearalenone (ZEN) | Gastrointestinal symptoms are not typical for ZEN toxicity. Decrease in feed intake and body weight, changes in intestinal microbiome | [195,233] | Pigs of various breeds: increase in the mucosal thickness, increase in the number of goblet cells, increase in lymphocyte number in epithelium, intensification of apoptosis, influence on enzymatic activity of mucosal cells, changes in intestinal microbiome | [10,11,175,193,194,195,234,235] |
| Intestinal porcine epithelial cell line (IPEC-1): influence on cell activity by changes in gene expression | [236] | |||
| Poultry: changes in the high of intestinal villi | [237] | |||
| Patulin (PAT) | Anorexia, salivation, distended abdomen loss of body weight, bleeding from the digestive tract and diarrhea | [238,239,240,241,242,243] | Human intestinal Caco-2 cells: the influence on permeability and ion transport in the mucosa, epithelial desquamation and sub mucosal swelling, genotoxicity effects, modulation of tight junctions | [198,199,244] |
| Rodents: mucosal layer injury, ulceration, fibrosis in the sub mucosa, necrosis | [238,239,240,241,242] | |||
| Porcine jejunal explant samples: villi atrophy and necrosis, decrease in the number of goblet cells, increase in cell apoptosis | [245] | |||
| Fumonisins (FUM) | reduction of feed consumption and body weight, abdominal pain, diarrhea | [246,247,248,249] | Human Colonic Cell Lines Caco-2, HT-29: growth inhibition and apoptosis induction, impact on mitochondrial metabolism, necrosis | [221,250] |
| Rodents: inflammatory infiltration increase in the number of mitotic figures in the intestinal crypts, necrotic changes | [251,252] | |||
| Intestinal porcine epithelial cell line (IPEC-1): inhibition of cell proliferation, intestinal barrier dysfunction | [253] |
Table 3.
Influence of mycotoxins on the enteric nervous system.
| Mycotoxin | Dose Examined | Animal Species or Kind of Tissues | Experimental Method Used in the Study | Character of Changes in the ENS | References |
|---|---|---|---|---|---|
| Doxynivalenol | from 0.2 mg/kg of chow to 2 mg/kg of chow | Wistar rats (Rattus novergicus) | immunohistochemistry and microscopic analysis | Reduction of the area of general population of the myenteric neurons, glial cells in the myenteric plexus and whole myenteric ganglia. | [14] |
| T2 Toxin | 12 µg/kg body weight/day | domestic pig of the White Large Polish Breed | Immunofluorescence method and microscopic analysis | Increase in the number of VIP-positive enteric neurons and intramucosal and intramuscular nerve fibers containing VIP in the stomach and duodenum. | [15] |
| 200 µg/kg of feed | domestic pig of the White Large Polish Breed | Immunofluorescence method and microscopic analysis | Increase in the number of CART-positive enteric neurons and intramucosal and intramuscular nerve fibers containing CART in the stomach, duodenum and descending colon. | [55] | |
| 12 µg/kg body weight/day | domestic pig of the White Large Polish Breed | Immunofluorescence method and microscopic analysis | Increase in the number and changes in neurochemical character of CGRP-positive enteric neurons in the descending colon. | [16] | |
| Zearalenon | 10 μg/kg body weight/day | domestic pig of the White Large Polish Breed | Immunofluorescence method and microscopic analysis | Increase in the number of nerve fibers immunoreactive to CART, SP, NOS, VIP, PACAP and decrease in the number of GAL-positive nerve fibers in the muscular layer of the ileum. | [19] |
| 0.1 mg/kg of chow/day | domestic pig of the White Large Polish Breed | Immunofluorescence method and microscopic analysis | Increase in the number of nerve fibers immunoreactive to SP and VIP with changes in their morphology | [21] | |
| 12 µg/kg body weight/day | domestic pig of the White Large Polish Breed | Immunofluorescence method and microscopic analysis | Increase in the number and changes in neurochemical character of neurons immunoreactive to CGRP in the descending colon. | [16] | |
| Patulin | EC50 = 1 ng/µL | culture of the enteric neurons from C57B6/J OlaHsd mice | Growth and viability testing, cytotoxicity test, evaluation of calcium signaling, measurement of glucose content, neurite outgrowth measurement and reactive oxygen species (ROS) test | Reduction of ATP levels and glucose concentration, disorders in calcium signaling in the enteric neurons, changes in their morphology. | [71] |
| Fumonisins | 1 and 3 mg/kg body weight | Wistar rats (Rattus novergicus) | immunohistochemistry method | Reduction of the size of neurons in the enteric ganglia. | [12] |
| 90 mg/kg body weight | Wistar rats (Rattus novergicus) | immunohistochemistry method and histomorphometrical analysis | Reduction of area and mean diameter of the submucous plexuses in duodenum. Reduction of area and mean diameter of myenteric and submucous plexuses in the jejunum, increase of sphericity of the enteric ganglia. | [13] |
VIP—vasoactive intestinal polypeptide; CART—cocaine- and amphetamine-regulated transcript; CGRP—calcitonin gene related peptide; SP—substance P; NOS—nitric oxide synthase; PACAP—pituitary adenylate cyclase activating peptide; GAL—galanin.
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Gonkowski, S.; Gajęcka, M.; Makowska, K. Mycotoxins and the Enteric Nervous System. Toxins 2020, 12, 461. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins12070461
AMA Style
Gonkowski S, Gajęcka M, Makowska K. Mycotoxins and the Enteric Nervous System. Toxins. 2020; 12(7):461. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins12070461
Chicago/Turabian StyleGonkowski, Sławomir, Magdalena Gajęcka, and Krystyna Makowska. 2020. "Mycotoxins and the Enteric Nervous System" Toxins 12, no. 7: 461. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins12070461
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.