Enhanced Antimicrobial Activity of N-Terminal Derivatives of a Novel Brevinin-1 Peptide from The Skin Secretion of Odorrana schmackeri

 ,
,  , , , and
, , , and
Abstract
:1. Introduction
2. Results
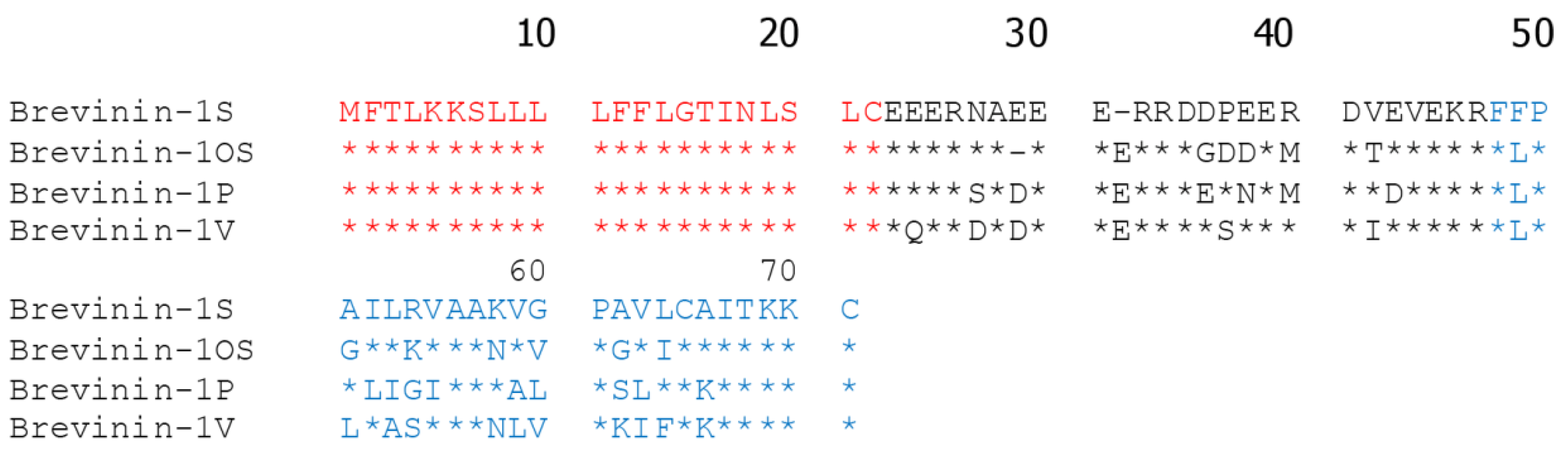
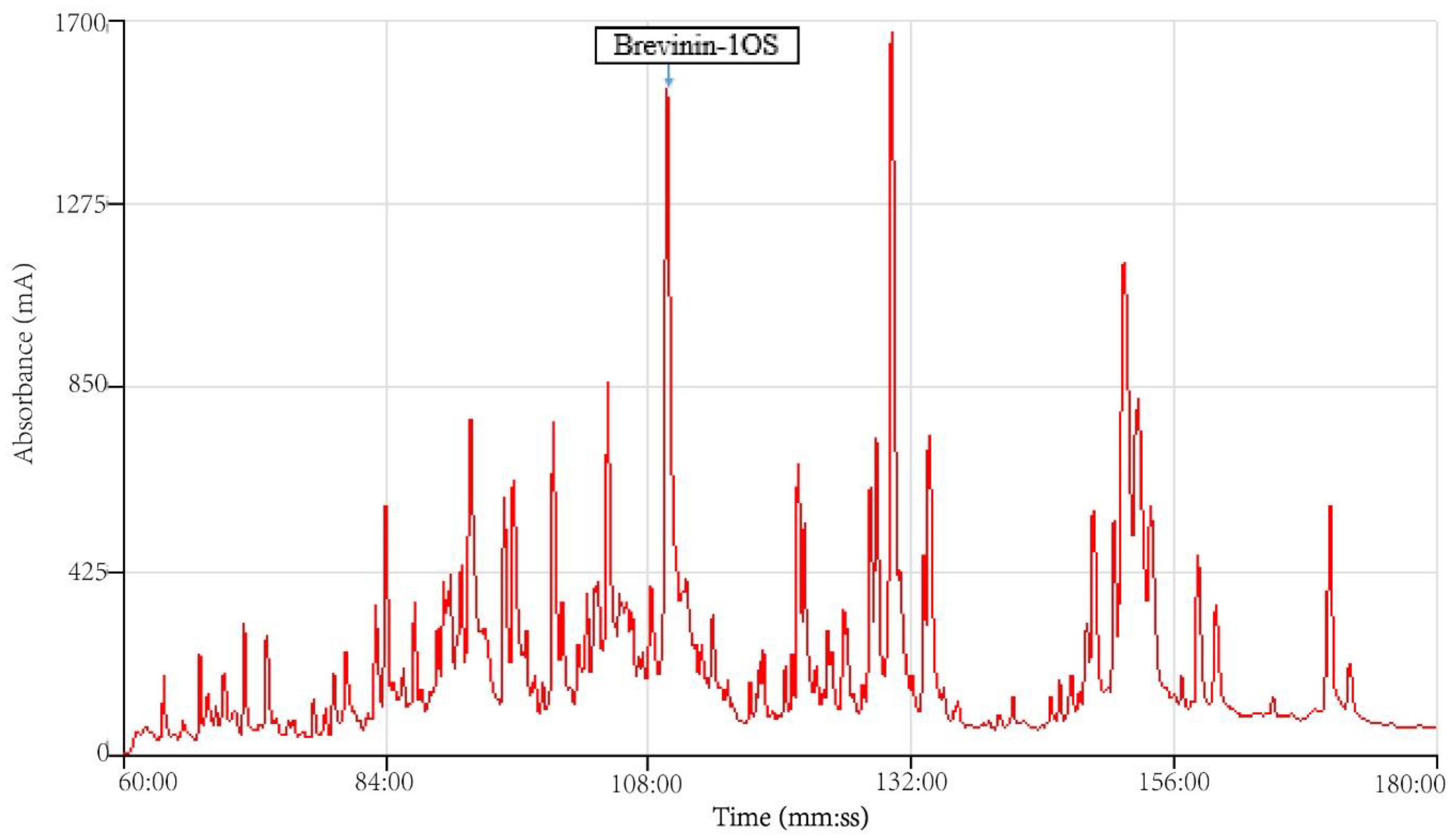
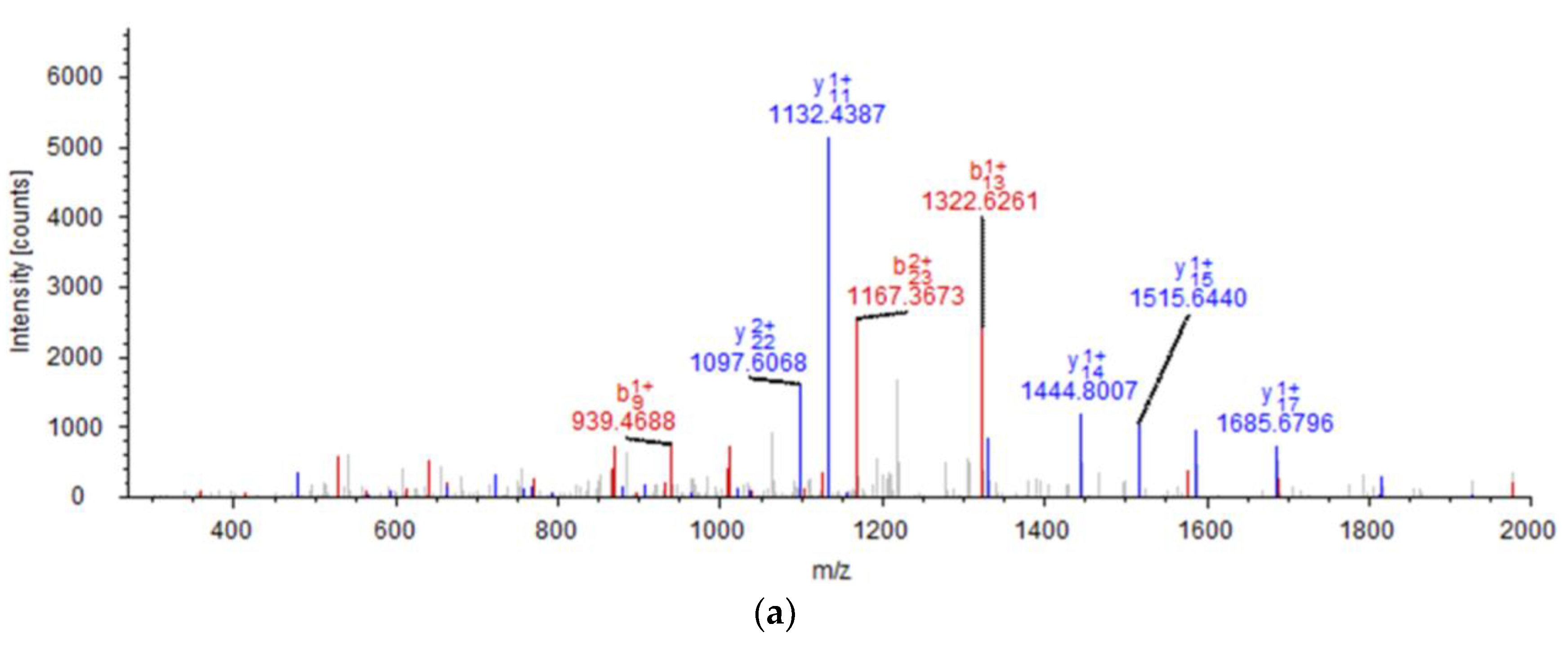
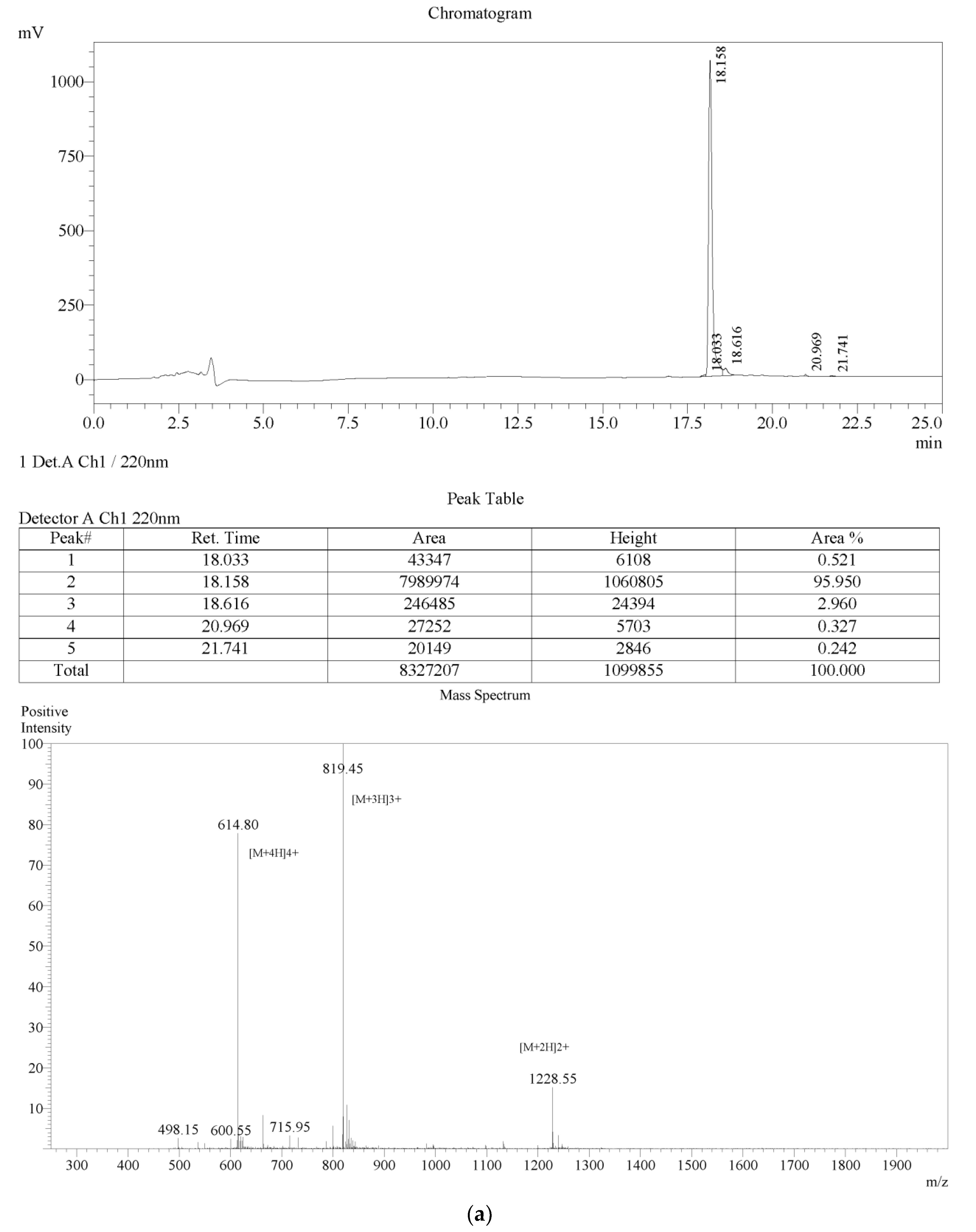
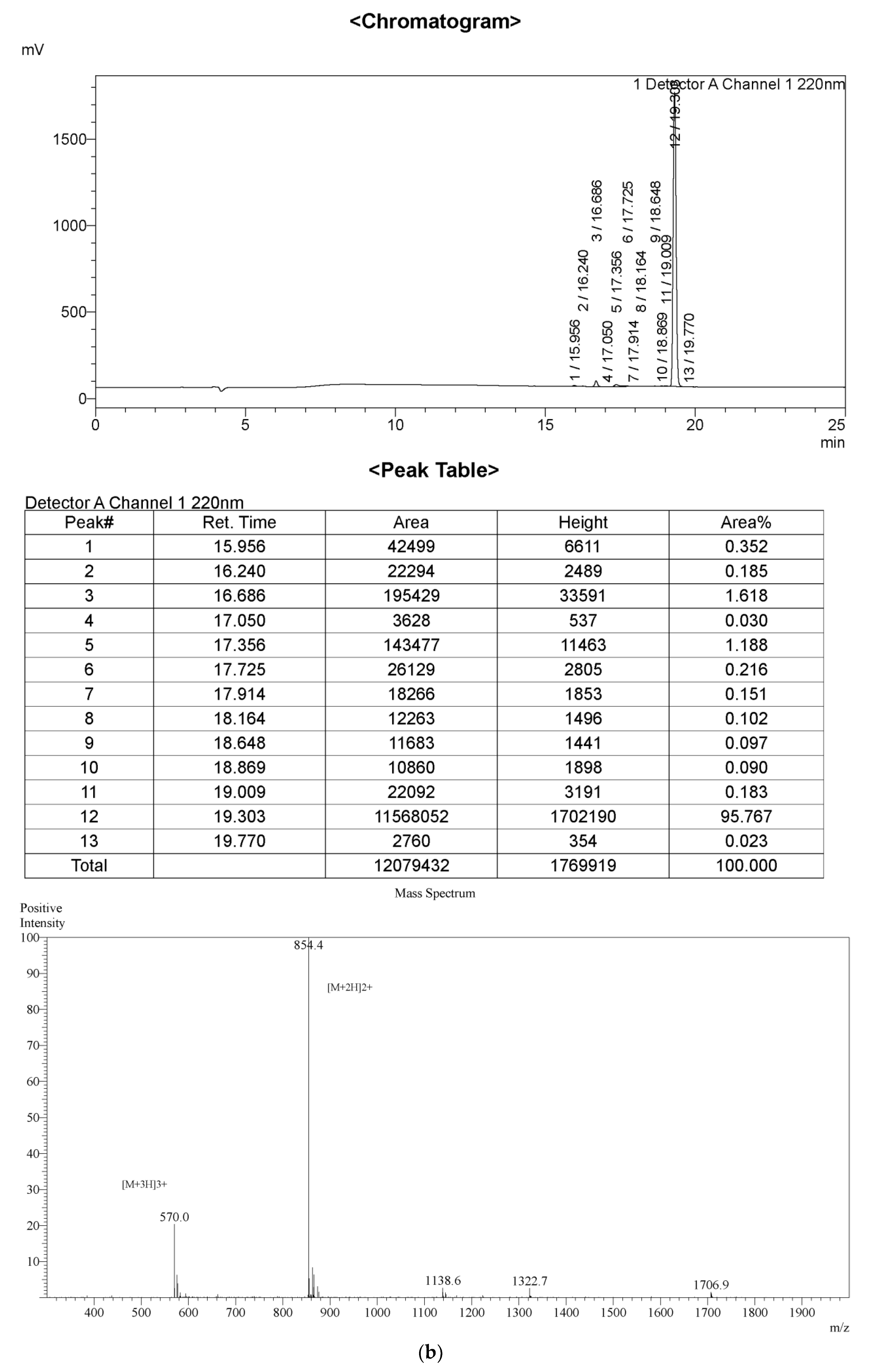
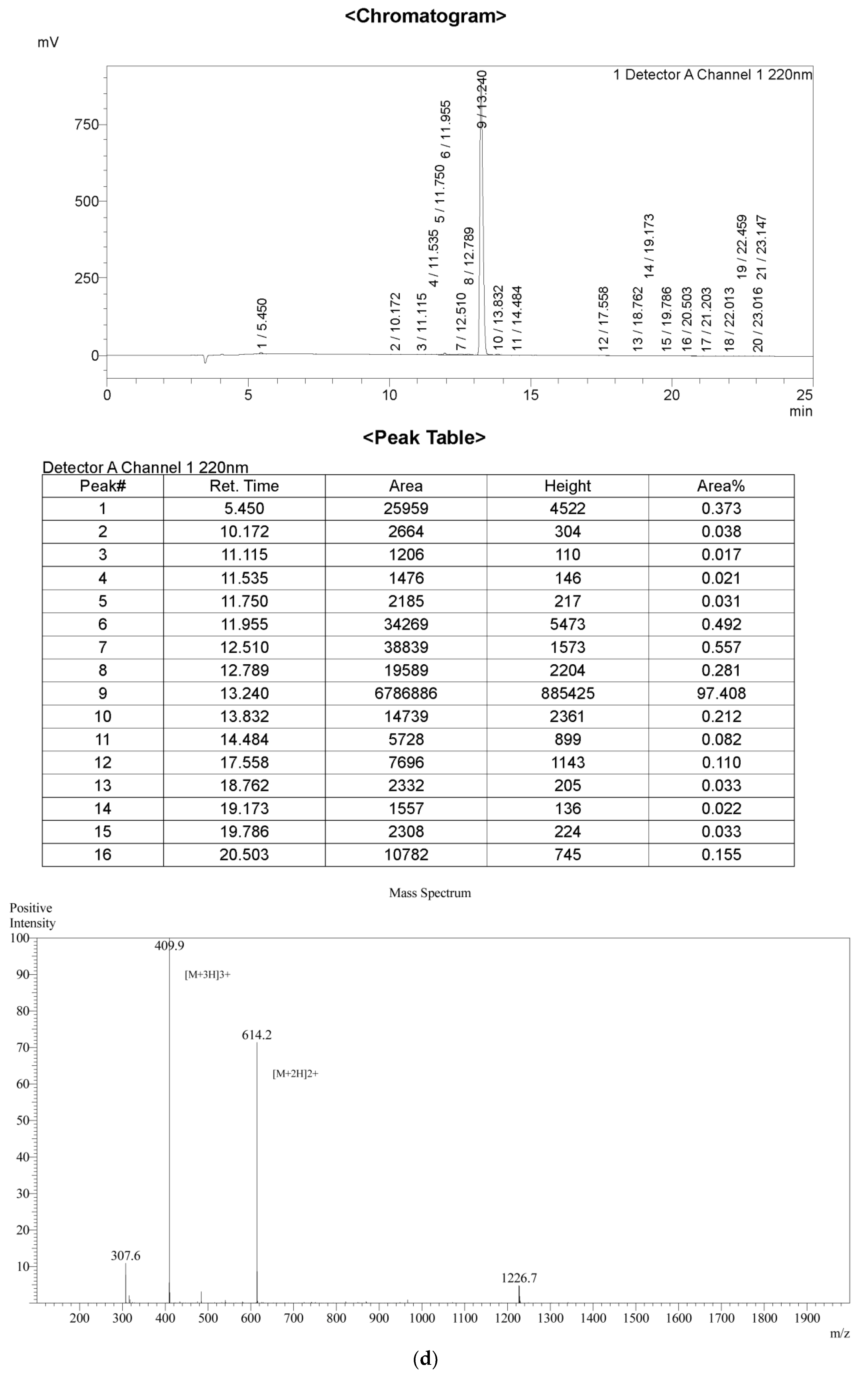
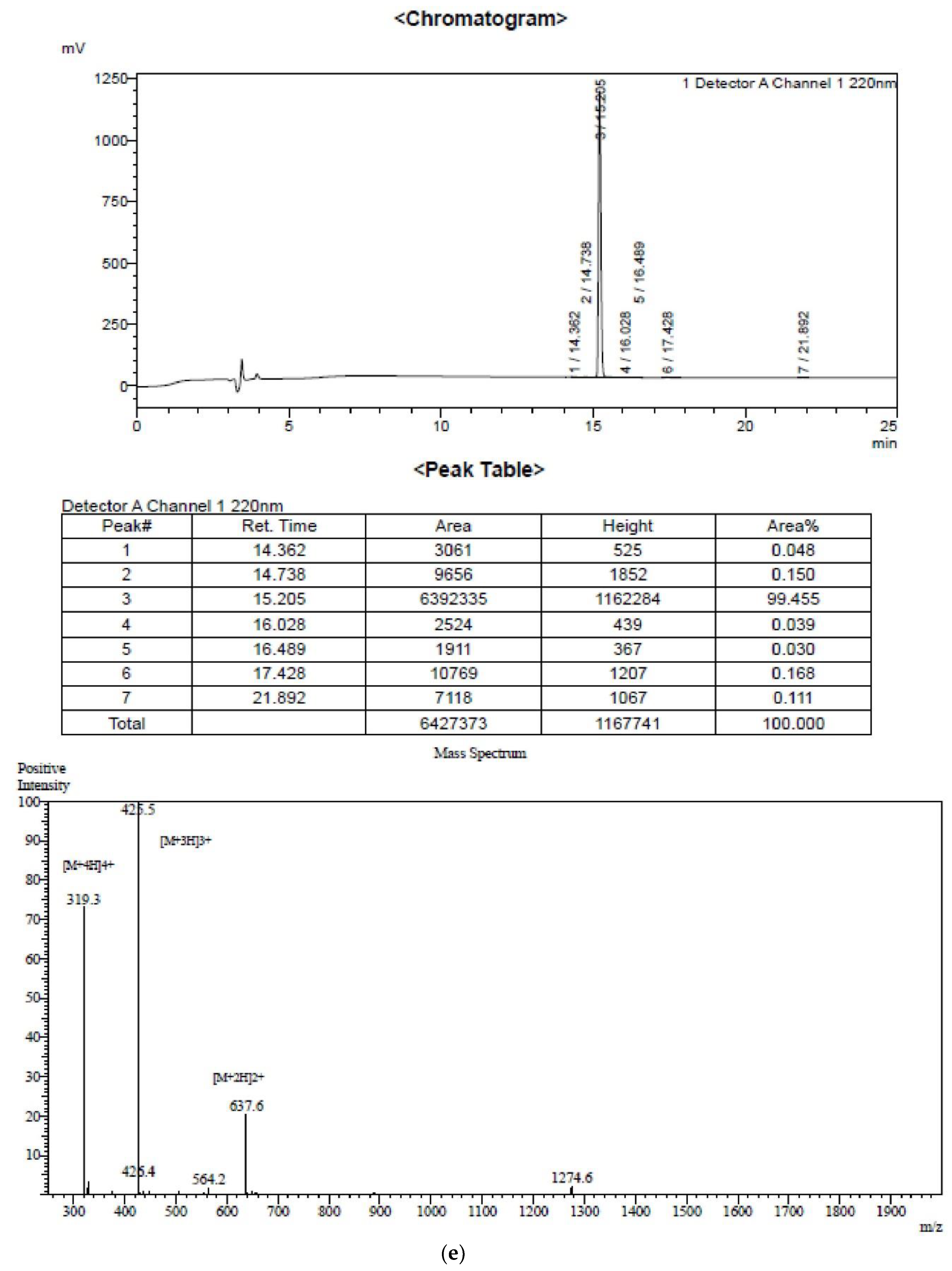
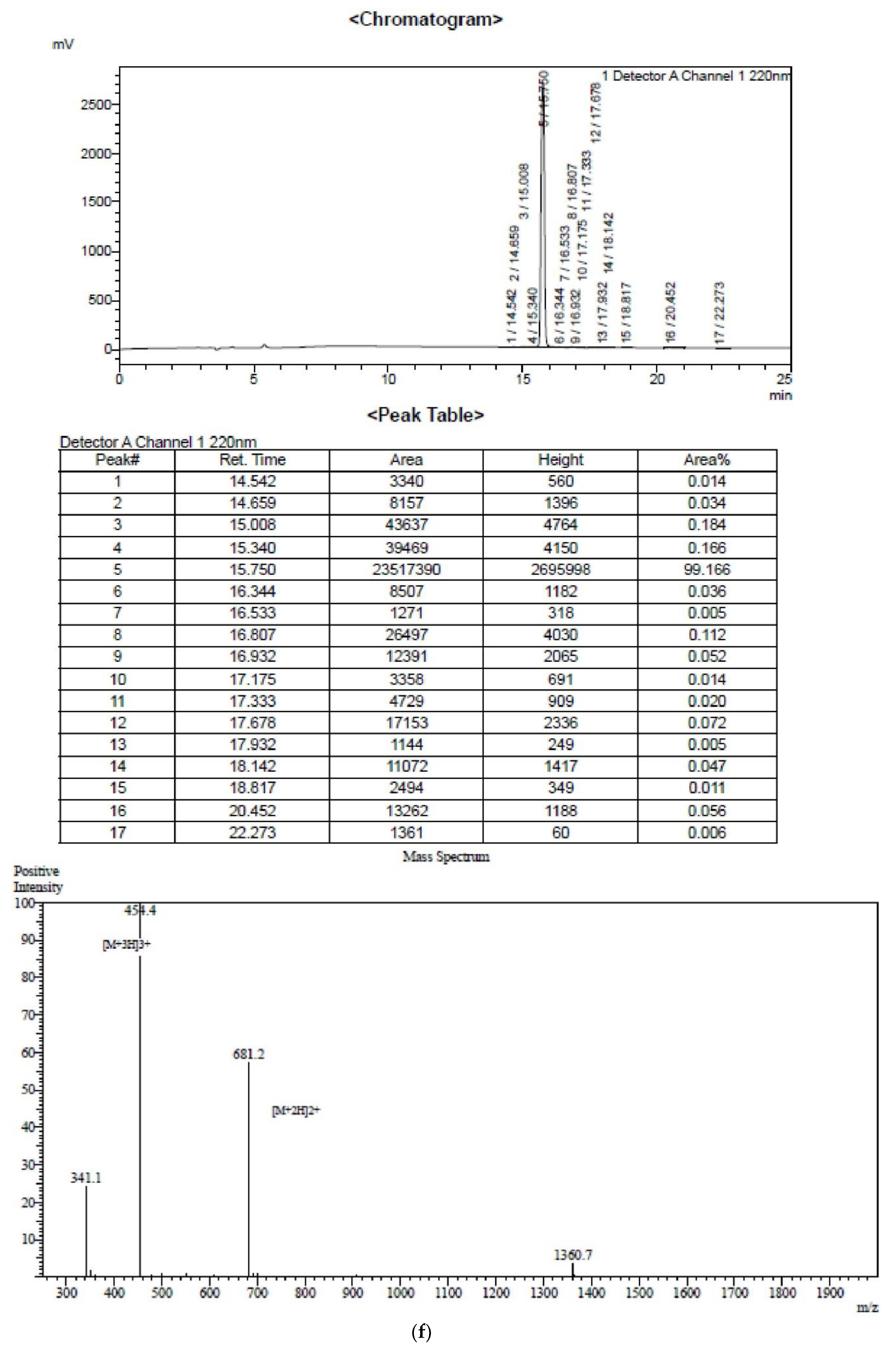
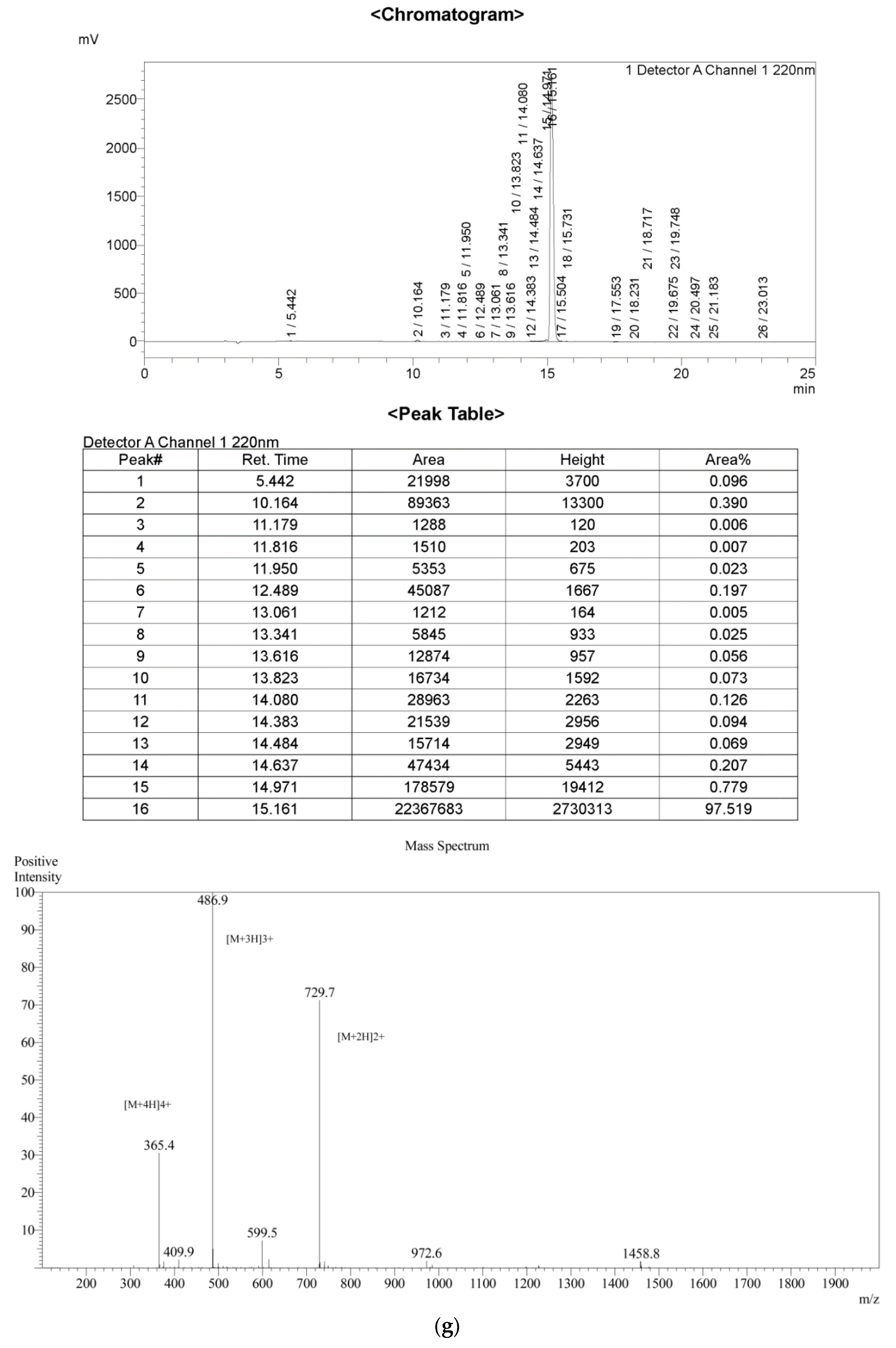
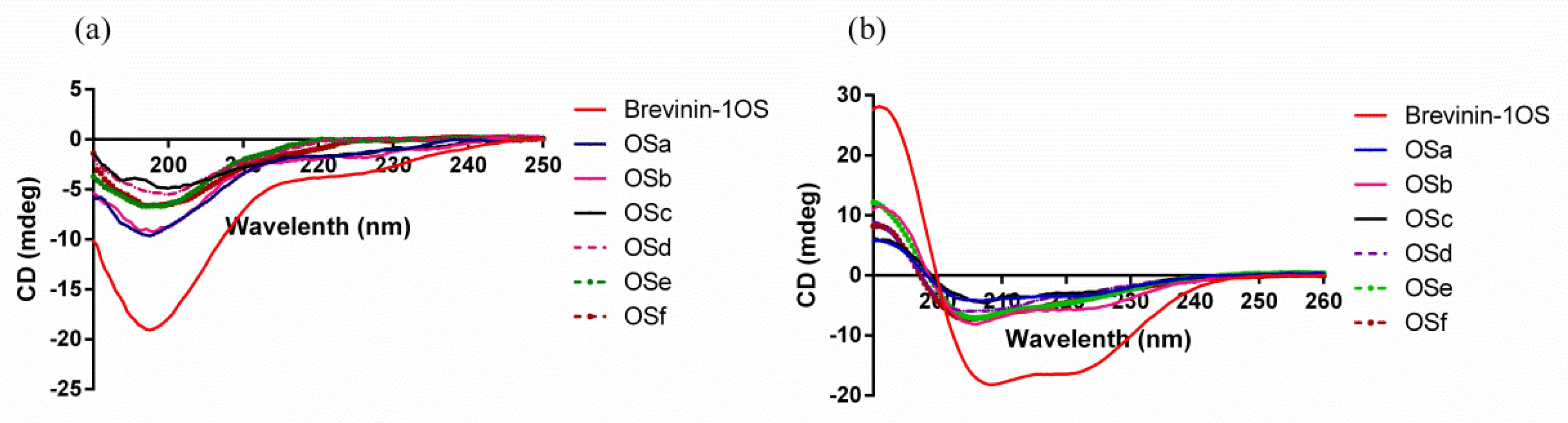
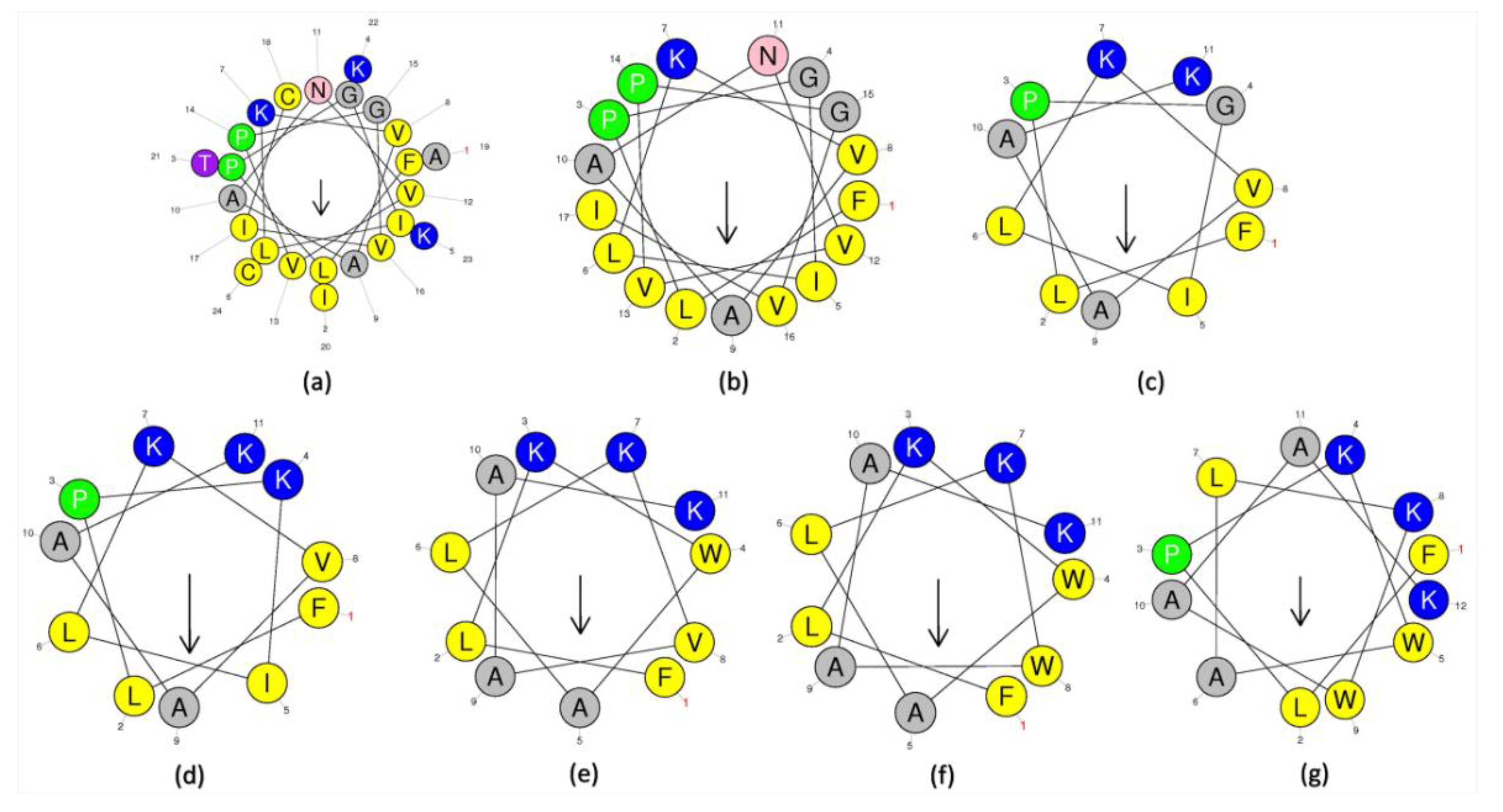
2.1. Peptide Identification, Design and Conformational Studies
2.2. Antimicrobial, Antibiofilm and Haemolytic Activity of Brevinin-1OS and Its Analogues
2.3. Antimicrobial Activity in Different Environmental Conditions
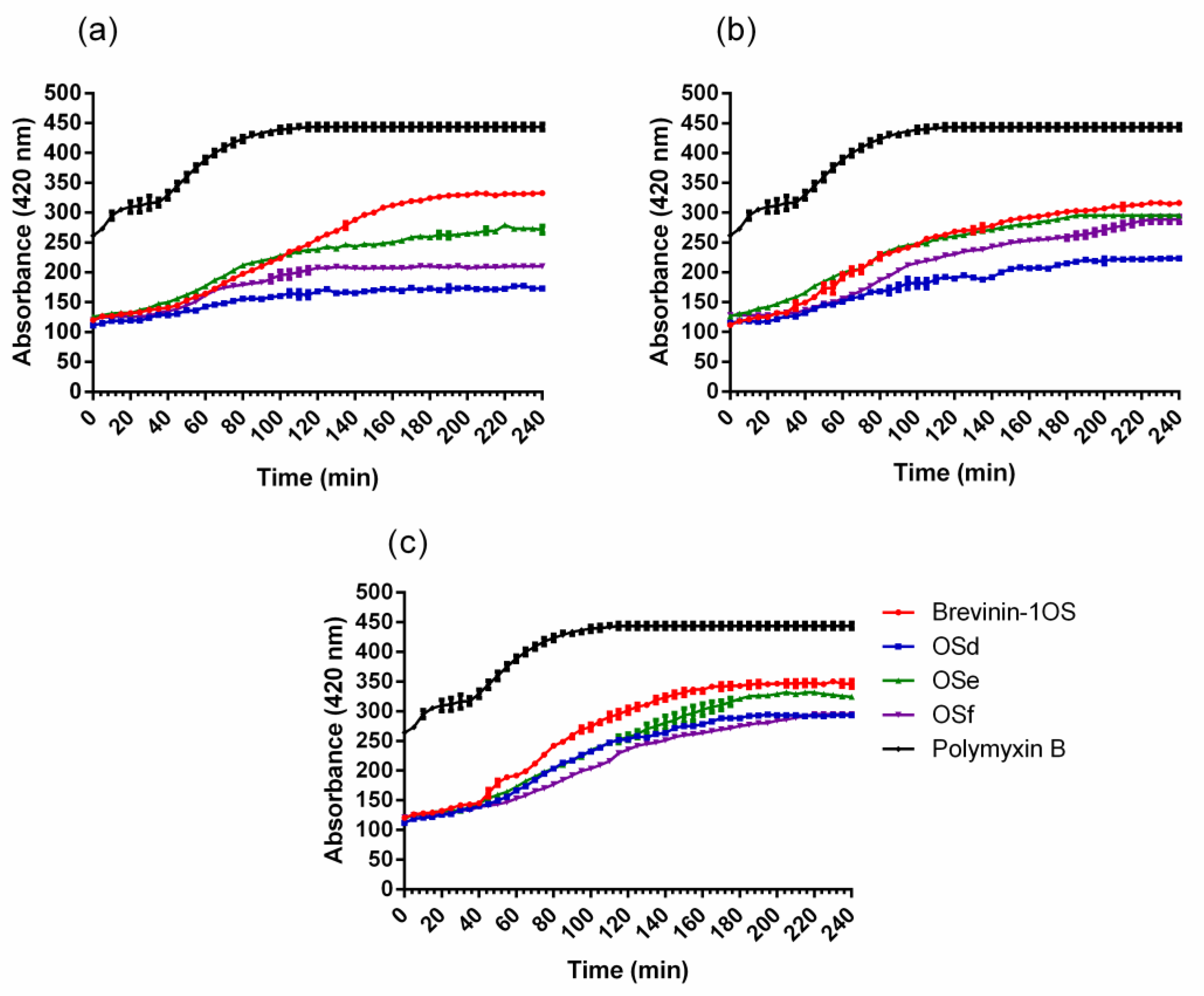
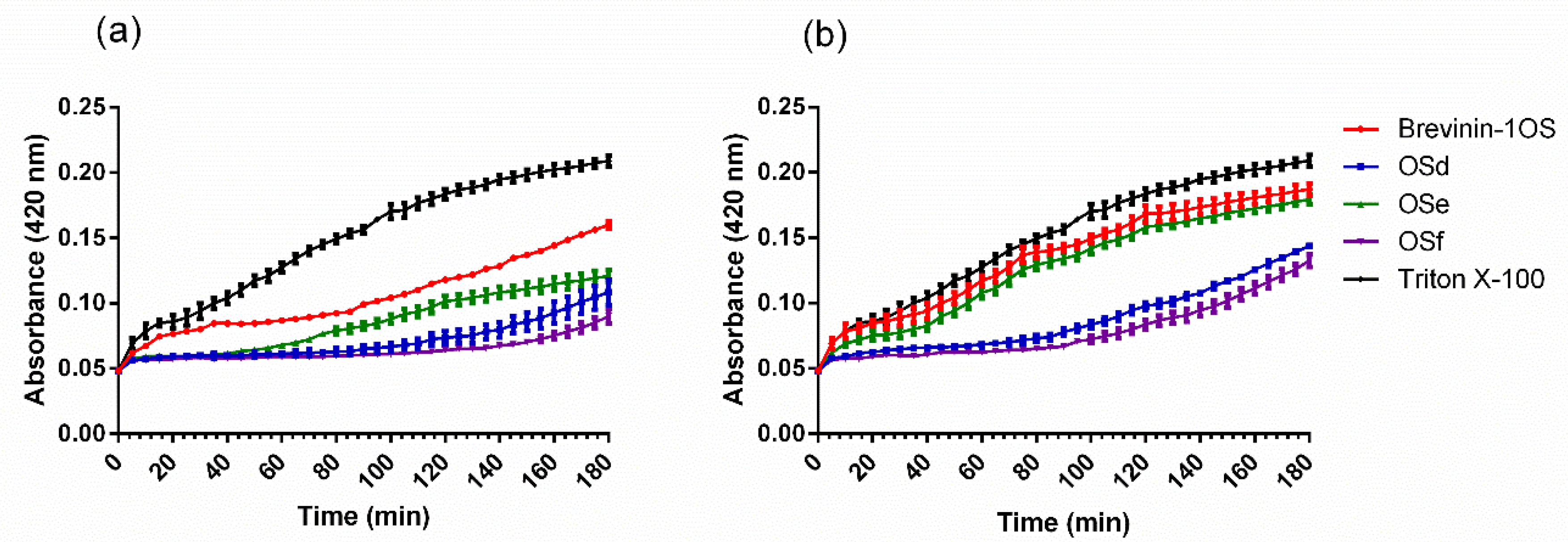
2.4. Outer Membrane (OM) and Inner Membrane (IM) Permeability Assays
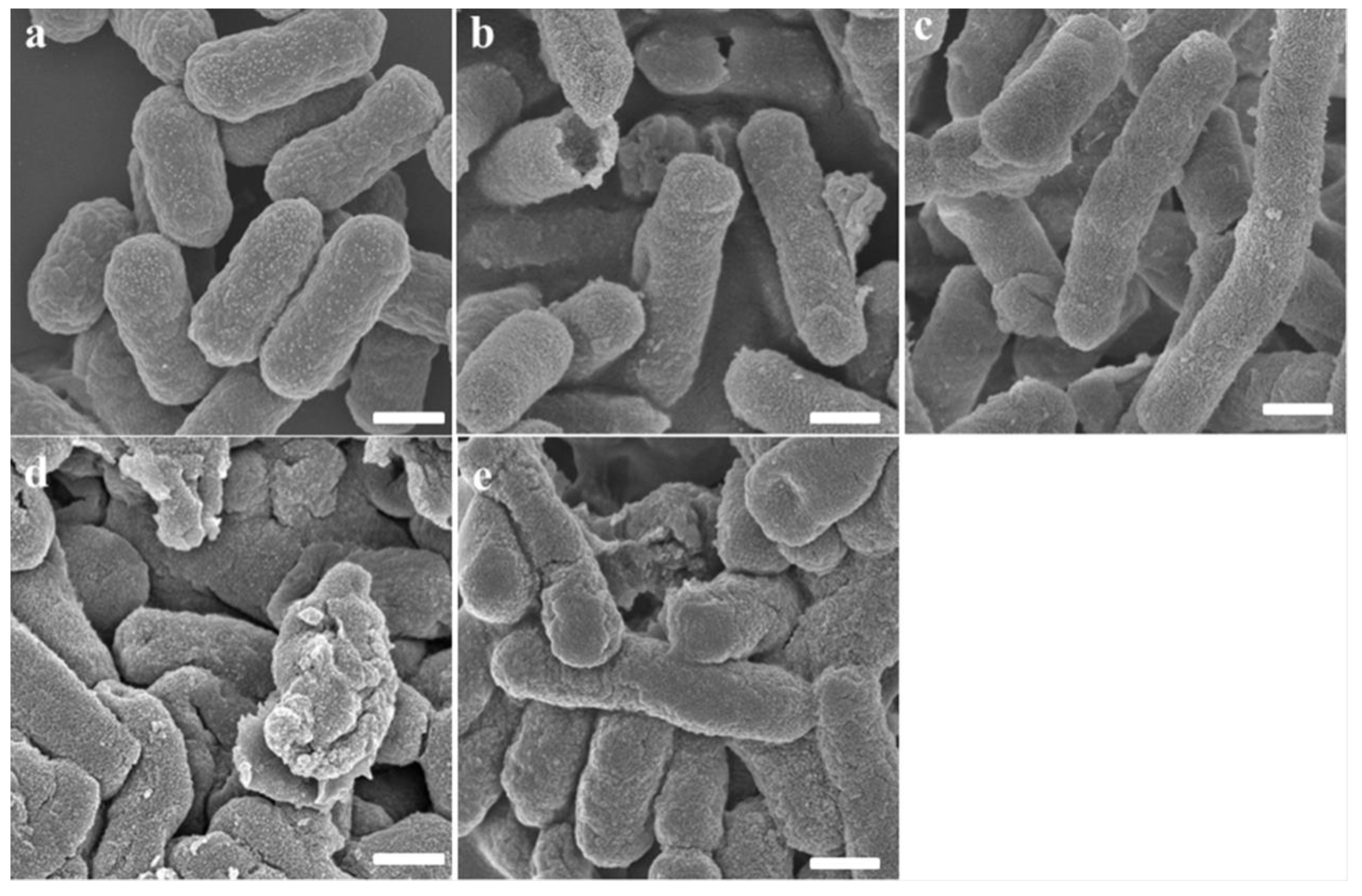
2.5. Scanning Electron Microscopy (SEM)
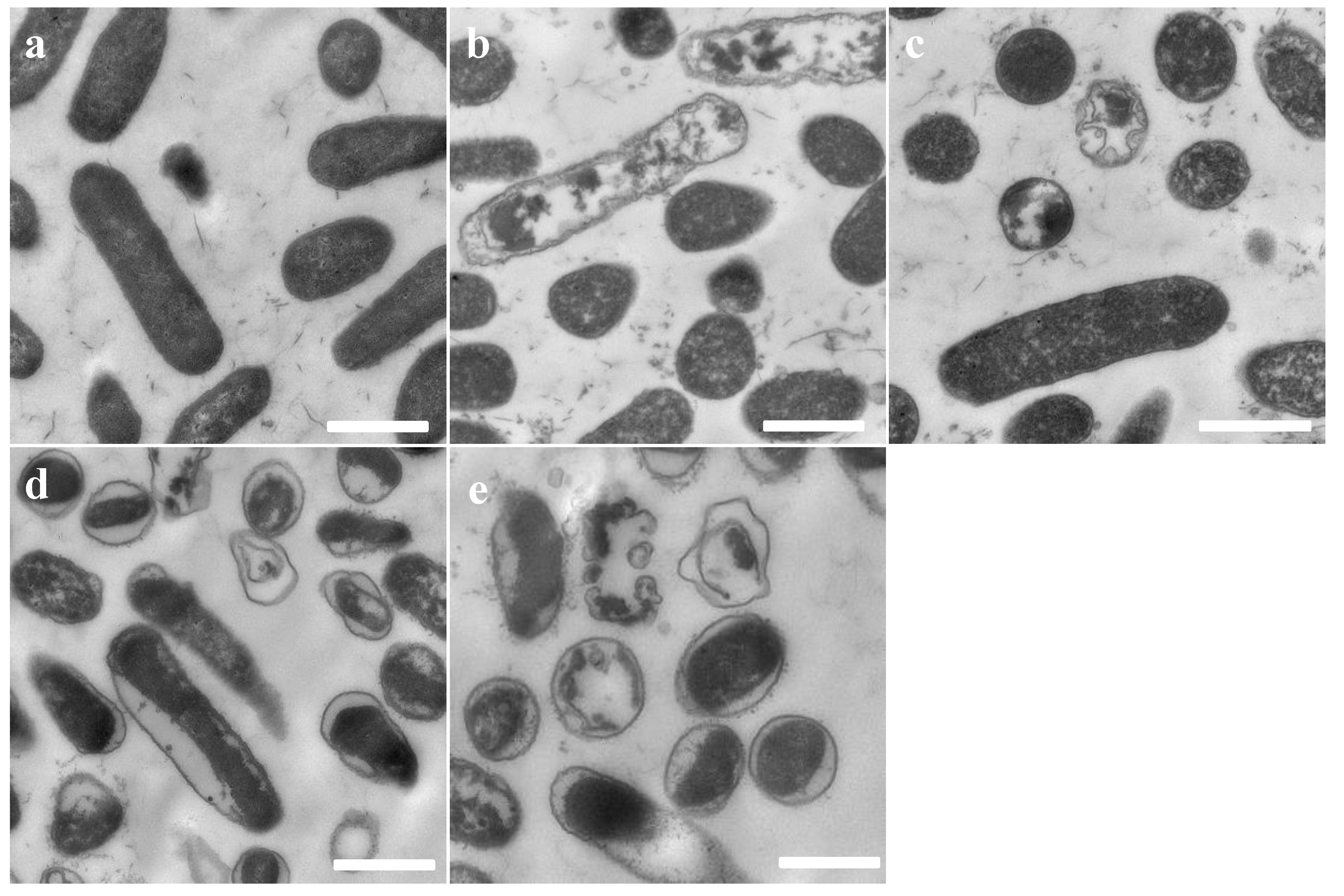
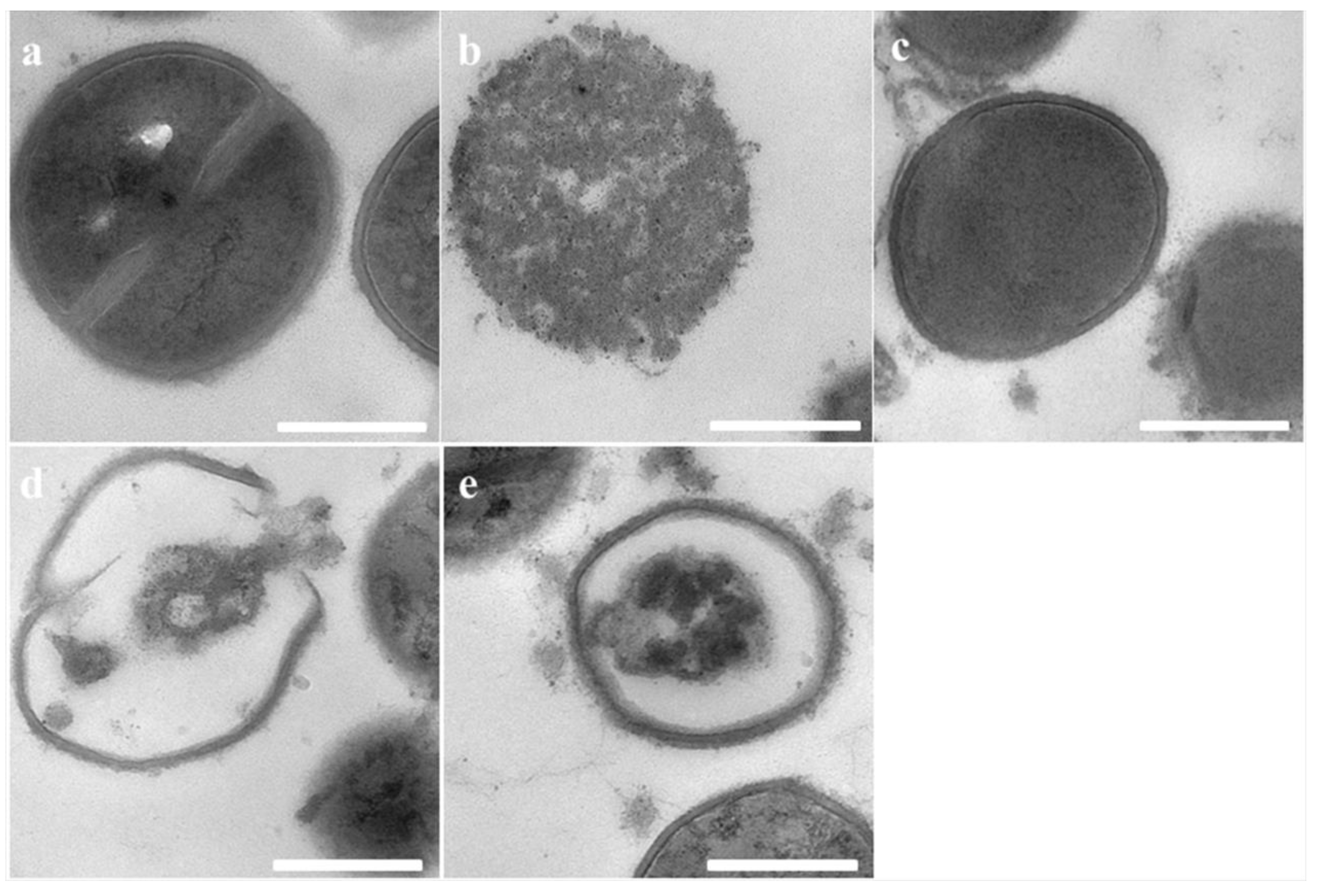
2.6. Transmission Electron Microscopy (TEM)
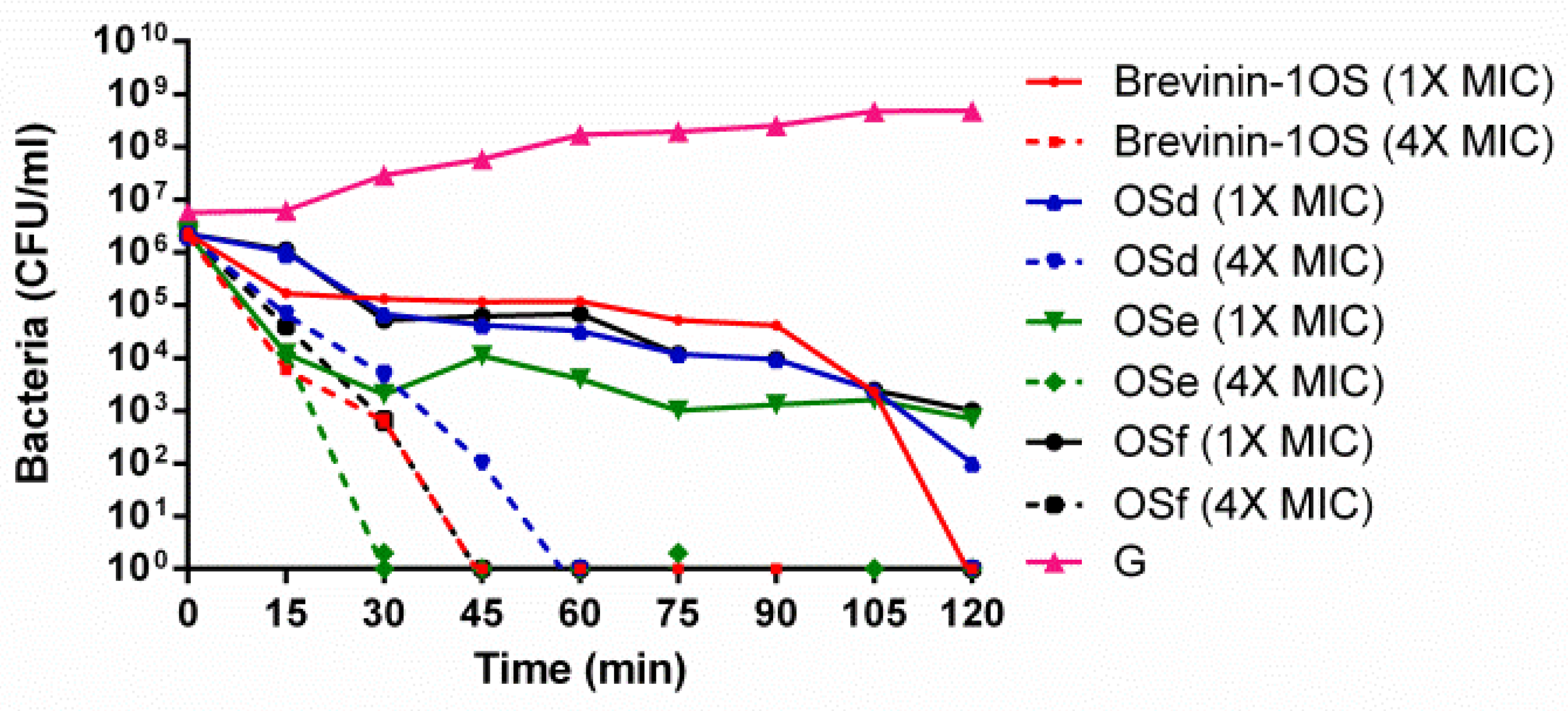
2.7. Time-Killing Assay Against MRSA
2.8. Skin Disinfection Test of Peptides
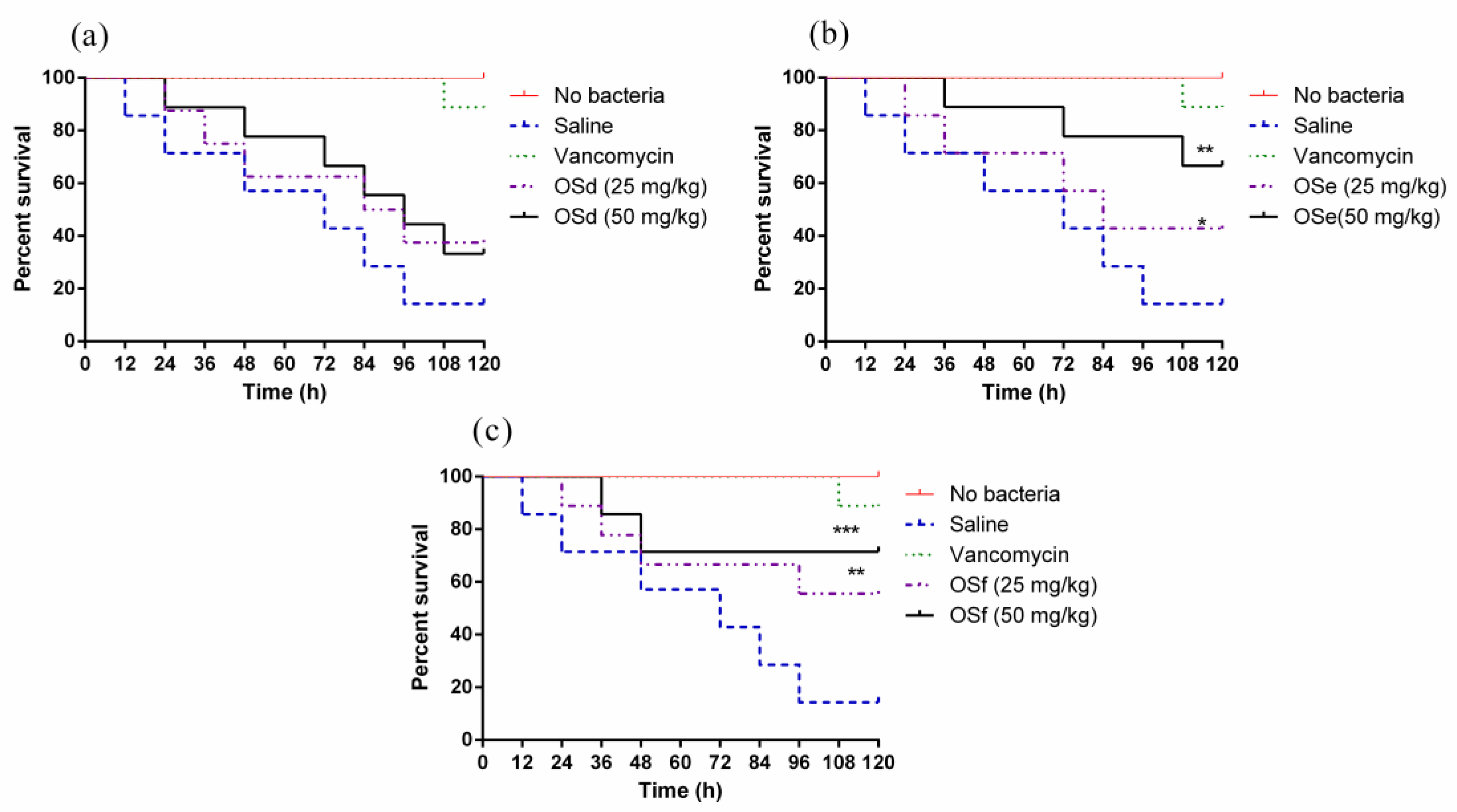
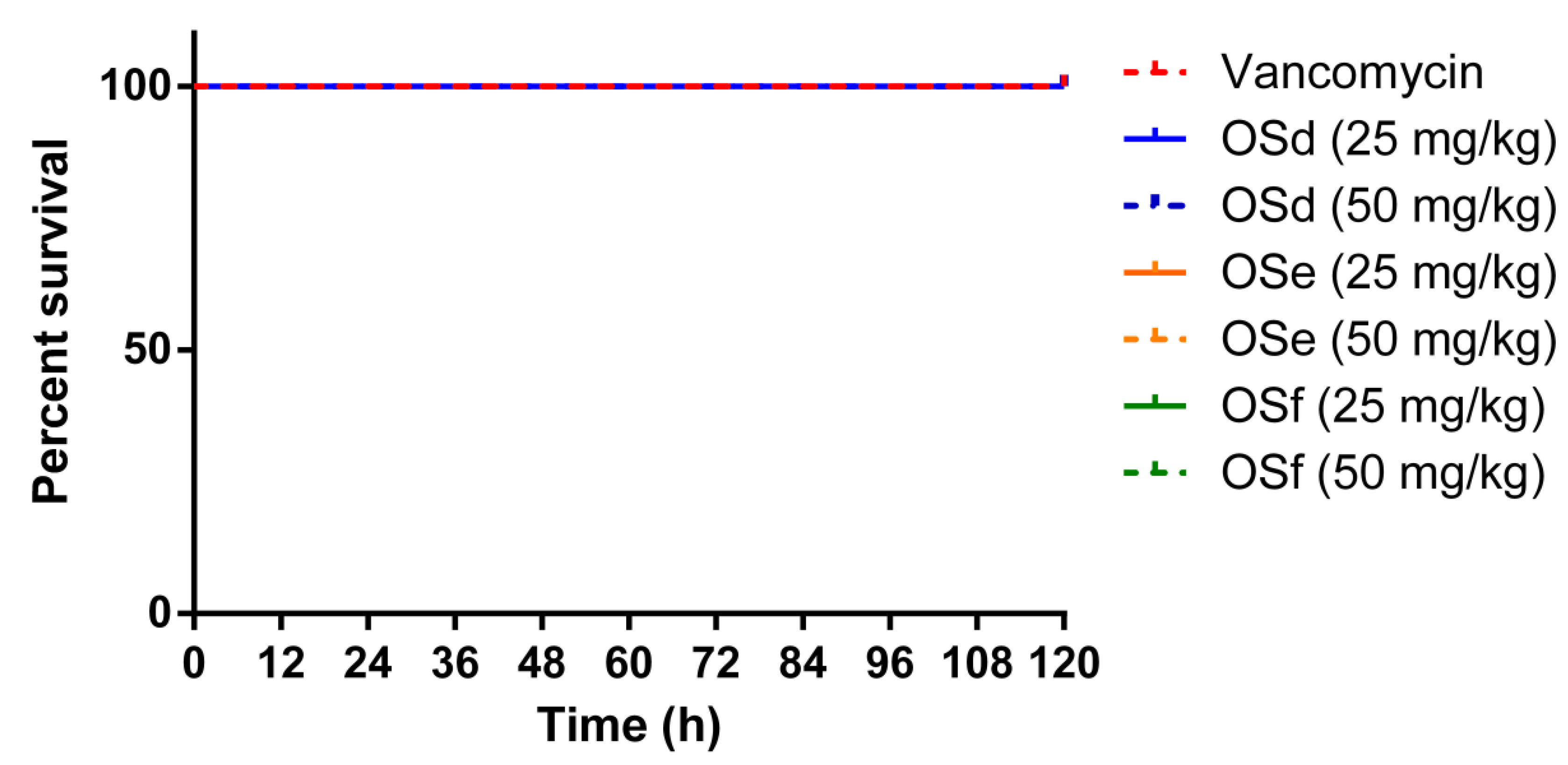
2.9. Treatment of MRSA-Infected Galleria Mellonella Larvae with Peptides
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Specimen Preparation and Secretion Harvesting
5.2. “Shotgun” Cloning of Biosynthetic Precursor and Identification of the Mature Peptide from the Skin Secretion of Odorrana Schmackeri
5.3. Peptide Design and Synthesis
5.4. Analysis of the Secondary Structure of Synthetic Peptides
5.5. Antimicrobial Activity Assays
5.6. Anti-Biofilm Assays
5.7. Haemolysis Assays
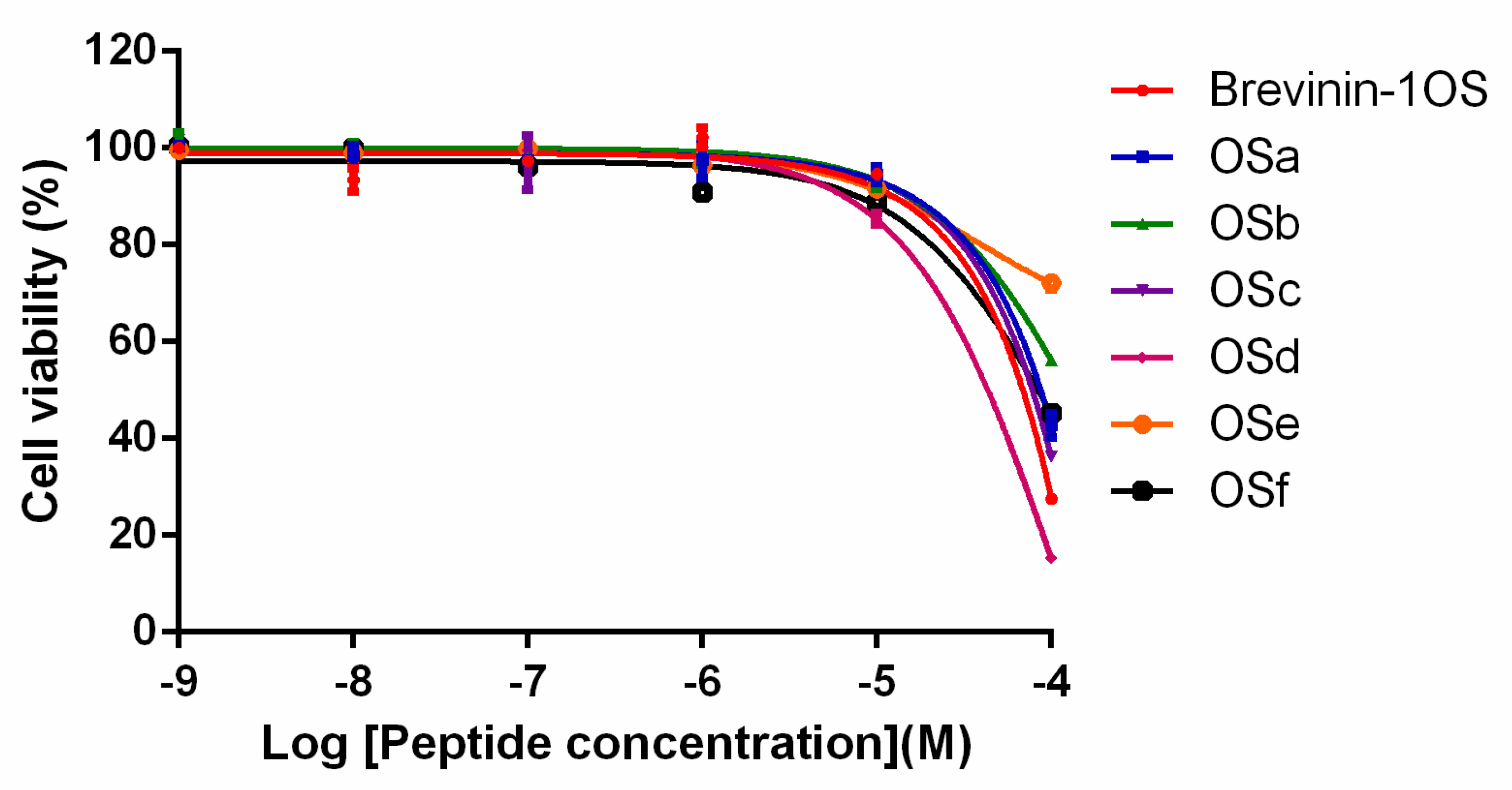
5.8. Measurement of Cytotoxicity
5.9. The Stability Assay of Peptides
5.10. Outer Membrane and Inner Membrane Permeability Assays
5.11. The Observation of Bacteria Morphology by Scanning Electron Microscopy (SEM) and Transmission Electron Microscopy (TEM)
5.12. Time-Killing Assays
5.13. Skin Disinfection Tests
5.14. Evaluation of Anti-MRSA Effects of the Peptides Using the Larvae of Galleria Mellonella
Author Contributions
Funding
Conflicts of Interest
Appendix A
















References
- Huang, L.; Chen, D.; Wang, L.; Lin, C.; Ma, C.; Xi, X.; Chen, T.; Shaw, C.; Zhou, M. Dermaseptin-PH: A Novel Peptide with Antimicrobial and Anticancer Activities from the Skin Secretion of the South American Orange-Legged Leaf Frog, Pithecopus (Phyllomedusa) hypochondrialis. Molecules 2017, 22, 1805. [Google Scholar] [CrossRef] [PubMed]
- Raja, Z.; André, S.; Abbassi, F.; Humblot, V.; Lequin, O.; Bouceba, T.; Correia, I.; Casale, S.; Foulon, T.; Sereno, D. Insight into the mechanism of action of temporin-SHa, a new broad-spectrum antiparasitic and antibacterial agent. PLoS ONE 2017, 12, e0174024. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, H.; Zhou, J.; Zhang, K.; Chu, H.; Liu, D.; Poon, V.K.-M.; Chan, C.C.-S.; Leung, H.-C.; Fai, N.; Lin, Y.-P. A novel peptide with potent and broad-spectrum antiviral activities against multiple respiratory viruses. Sci. Rep. 2016, 6, 22008. [Google Scholar] [CrossRef] [PubMed]
- Seo, M.-D.; Won, H.-S.; Kim, J.-H.; Mishig-Ochir, T.; Lee, B.-J. Antimicrobial peptides for therapeutic applications: A review. Molecules 2012, 17, 12276–12286. [Google Scholar] [CrossRef] [Green Version]
- Zhang, M.-Z.; Chen, Q.; Yang, G.-F. A review on recent developments of indole-containing antiviral agents. Eur. J. Med. Chem. 2015, 89, 421–441. [Google Scholar] [CrossRef] [PubMed]
- Kosikowska, P.; Lesner, A. Antimicrobial peptides (AMPs) as drug candidates: A patent review (2003–2015). Expert Opin. Ther. Pat. 2016, 26, 689–702. [Google Scholar] [CrossRef] [PubMed]
- Mahlapuu, M.; Håkansson, J.; Ringstad, L.; Björn, C. Antimicrobial Peptides: An Emerging Category of Therapeutic Agents. Front. Cell. Infect. Microbiol. 2016, 6, 194. [Google Scholar] [CrossRef] [Green Version]
- Amblard, M.; Fehrentz, J.-A.; Martinez, J.; Subra, G. Methods and protocols of modern solid phase peptide synthesis. Mol. Biotechnol. 2006, 33, 239–254. [Google Scholar] [CrossRef]
- Raibaut, L.; El Mahdi, O.; Melnyk, O. Solid phase protein chemical synthesis. In Protein Ligation and Total Synthesis II; Springer: Cham, Switzerland, 2014; pp. 103–154. [Google Scholar]
- Björn, C.; Noppa, L.; Salomonsson, E.N.; Johansson, A.-L.; Nilsson, E.; Mahlapuu, M.; Håkansson, J. Efficacy and safety profile of the novel antimicrobial peptide PXL150 in a mouse model of infected burn wounds. Int. J. Antimicrob. Agents 2015, 45, 519–524. [Google Scholar] [CrossRef]
- Fox, J.L. Antimicrobial peptides stage a comeback. Nat. Biotechnol. 2013, 31, 379. [Google Scholar] [CrossRef]
- Zhu, X.; Dong, N.; Wang, Z.; Ma, Z.; Zhang, L.; Ma, Q.; Shan, A. Design of imperfectly amphipathic α-helical antimicrobial peptides with enhanced cell selectivity. Acta Biomater. 2014, 10, 244–257. [Google Scholar] [CrossRef] [PubMed]
- Ong, Z.Y.; Wiradharma, N.; Yang, Y.Y. Strategies employed in the design and optimization of synthetic antimicrobial peptide amphiphiles with enhanced therapeutic potentials. Adv. Drug Deliv. Rev. 2014, 78, 28–45. [Google Scholar] [CrossRef]
- Morikawa, N.; Hagiwara, K.i.; Nakajima, T. Brevinin-1 and-2, unique antimicrobial peptides from the skin of the frog, Rana brevipoda porsa. Biochem. Biophys. Res. Commun. 1992, 189, 184–190. [Google Scholar] [CrossRef]
- Clark, D.P.; Durell, S.; Maloy, W.L.; Zasloff, M. Ranalexin: A novel antimicrobial peptide from bullfrog (Rana catesbeiana) skin, structurally related to the bacterial antibiotic, polymyxin. J. Biol. Chem. 1994, 269, 10849–10855. [Google Scholar] [PubMed]
- Wang, H.; Lu, Y.; Zhang, X.; Hu, Y.; Yu, H.; Liu, J.; Sun, J. The novel antimicrobial peptides from skin of Chinese broad-folded frog, Hylarana latouchii (Anura: Ranidae). Peptides 2009, 30, 273–282. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Yan, X.; Yu, H.; Hu, Y.; Yu, Z.; Zheng, H.; Chen, Z.; Zhang, Z.; Liu, J. Isolation, characterization and molecular cloning of new antimicrobial peptides belonging to the brevinin-1 and temporin families from the skin of Hylarana latouchii (Anura: Ranidae). Biochimie 2009, 91, 540–547. [Google Scholar] [CrossRef]
- Conlon, J.M.; Kolodziejek, J.; Nowotny, N.; Leprince, J.; Vaudry, H.; Coquet, L.; Jouenne, T.; King, J.D. Characterization of antimicrobial peptides from the skin secretions of the Malaysian frogs, Odorrana hosii and Hylarana picturata (Anura:Ranidae). Toxicon Off. J. Int. Soc. Toxinology 2008, 52, 465–473. [Google Scholar] [CrossRef]
- Reshmy, V.; Preeji, V.; Parvin, A.; Santhoshkumar, K.; George, S. Three novel antimicrobial peptides from the skin of the Indian bronzed frog Hylarana temporalis (Anura: Ranidae). J. Pept. Sci. 2011, 17, 342–347. [Google Scholar] [CrossRef]
- Chen, Q.; Cheng, P.; Ma, C.; Xi, X.; Wang, L.; Zhou, M.; Bian, H.; Chen, T. Evaluating the Bioactivity of a Novel Broad-Spectrum Antimicrobial Peptide Brevinin-1GHa from the Frog Skin Secretion of Hylarana guentheri and Its Analogues. Toxins 2018, 10, 413. [Google Scholar] [CrossRef] [Green Version]
- Park, S.-H.; Kim, H.-E.; Kim, C.-M.; Yun, H.-J.; Choi, E.-C.; Lee, B.-J. Role of proline, cysteine and a disulphide bridge in the structure and activity of the anti-microbial peptide gaegurin 5. Biochem. J. 2002, 368 Pt 1, 171. [Google Scholar] [CrossRef] [Green Version]
- Kumari, V.; Nagaraj, R. Structure-function studies on the amphibian peptide brevinin 1E: Translocating the cationic segment from the C-terminal end to a central position favors selective antibacterial activity. J. Pept. Res. 2001, 58, 433–441. [Google Scholar] [CrossRef]
- Abraham, P.; Sundaram, A.; Asha, R.; Reshmy, V.; George, S.; Kumar, K.S. Structure-activity relationship and mode of action of a frog secreted antibacterial peptide B1CTcu5 using synthetically and modularly modified or deleted (SMMD) peptides. PLoS ONE 2015, 10, e0124210. [Google Scholar] [CrossRef]
- Turner, J.; Cho, Y.; Dinh, N.-N.; Waring, A.J.; Lehrer, R.I. Activities of LL-37, a cathelin-associated antimicrobial peptide of human neutrophils. Antimicrob. Agents Chemother. 1998, 42, 2206–2214. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chu, H.-L.; Yu, H.-Y.; Yip, B.-S.; Chih, Y.-H.; Liang, C.-W.; Cheng, H.-T.; Cheng, J.-W. Boosting salt resistance of short antimicrobial peptides. Antimicrob. Agents Chemother. 2013, 57, 4050–4052. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Amiche, M.; Seon, A.A.; Pierre, T.N.; Nicolas, P. The dermaseptin precursors: A protein family with a common preproregion and a variable C-terminal antimicrobial domain. FEBS Lett. 1999, 456, 352–356. [Google Scholar] [CrossRef] [Green Version]
- Conlon, J.M.; Kolodziejek, J.; Nowotny, N. Antimicrobial peptides from ranid frogs: Taxonomic and phylogenetic markers and a potential source of new therapeutic agents. Biochim. et Biophys. Acta (BBA) Proteins Proteom. 2004, 1696, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Suh, J.-Y.; Lee, K.-H.; Chi, S.-W.; Hong, S.-Y.; Choi, B.-W.; Moon, H.-M.; Choi, B.-S. Unusually stable helical kink in the antimicrobial peptide—A derivative of gaegurin. FEBS Lett. 1996, 392, 309–312. [Google Scholar] [CrossRef] [Green Version]
- Won, H.-S.; Kang, S.-J.; Lee, B.-J. Action mechanism and structural requirements of the antimicrobial peptides, gaegurins. Biochim. et Biophys. Acta (BBA) Biomembr. 2009, 1788, 1620–1629. [Google Scholar] [CrossRef] [Green Version]
- Jiang, Z.; Vasil, A.I.; Hale, J.D.; Hancock, R.E.; Vasil, M.L.; Hodges, R.S. Effects of net charge and the number of positively charged residues on the biological activity of amphipathic alpha-helical cationic antimicrobial peptides. Biopolymers 2008, 90, 369–383. [Google Scholar] [CrossRef]
- Biedermannova, L.; Riley, K.E.; Berka, K.; Hobza, P.; Vondrasek, J. Another role of proline: Stabilization interactions in proteins and protein complexes concerning proline and tryptophane. Phys. Chem. Chem. Phys. 2008, 10, 6350–6359. [Google Scholar] [CrossRef]
- Won, H.S.; Park, S.H.; Kim, H.E.; Hyun, B.; Kim, M.; Lee, B.J.; Lee, B.J. Effects of a tryptophanyl substitution on the structure and antimicrobial activity of C-terminally truncated gaegurin 4. FEBS J. 2002, 269, 4367–4374. [Google Scholar] [CrossRef] [PubMed]
- De Planque, M.R.; Bonev, B.B.; Demmers, J.A.; Greathouse, D.V.; Koeppe, R.E.; Separovic, F.; Watts, A.; Killian, J.A. Interfacial anchor properties of tryptophan residues in transmembrane peptides can dominate over hydrophobic matching effects in peptide-lipid interactions. Biochemistry 2003, 42, 5341–5348. [Google Scholar] [CrossRef] [PubMed]
- Ma, Z.; Wei, D.; Yan, P.; Zhu, X.; Shan, A.; Bi, Z. Characterization of cell selectivity, physiological stability and endotoxin neutralization capabilities of α-helix-based peptide amphiphiles. Biomaterials 2015, 52, 517–530. [Google Scholar] [CrossRef]
- Mattei, B.; Miranda, A.; Perez, K.R.; Riske, K.A. Structure–activity relationship of the antimicrobial peptide gomesin: The role of peptide hydrophobicity in its interaction with model membranes. Langmuir 2014, 30, 3513–3521. [Google Scholar] [CrossRef]
- Chen, Y.; Guarnieri, M.T.; Vasil, A.I.; Vasil, M.L.; Mant, C.T.; Hodges, R.S. Role of peptide hydrophobicity in the mechanism of action of α-helical antimicrobial peptides. Antimicrob. Agents Chemother. 2007, 51, 1398–1406. [Google Scholar] [CrossRef] [Green Version]
- Mohamed, M.F.; Brezden, A.; Mohammad, H.; Chmielewski, J.; Seleem, M.N. A short D-enantiomeric antimicrobial peptide with potent immunomodulatory and antibiofilm activity against multidrug-resistant Pseudomonas aeruginosa and Acinetobacter baumannii. Sci. Rep. 2017, 7, 6953. [Google Scholar] [CrossRef]
- Mingeot-Leclercq, M.-P.; Décout, J.-L. Bacterial lipid membranes as promising targets to fight antimicrobial resistance, molecular foundations and illustration through the renewal of aminoglycoside antibiotics and emergence of amphiphilic aminoglycosides. MedChemComm 2016, 7, 586–611. [Google Scholar] [CrossRef]
- Teixeira, V.; Feio, M.J.; Bastos, M. Role of lipids in the interaction of antimicrobial peptides with membranes. Prog. Lipid Res. 2012, 51, 149–177. [Google Scholar] [CrossRef]
- Zhu, X.; Zhang, L.; Wang, J.; Ma, Z.; Xu, W.; Li, J.; Shan, A. Characterization of antimicrobial activity and mechanisms of low amphipathic peptides with different α-helical propensity. Acta Biomater. 2015, 18, 155–167. [Google Scholar] [CrossRef]
- Ahmed, T.A.; Hammami, R. Recent insights into structure–function relationships of antimicrobial peptides. J. Food Biochem. 2018, 43, e12546. [Google Scholar] [CrossRef] [Green Version]
- Hall, C.W.; Mah, T.-F. Molecular mechanisms of biofilm-based antibiotic resistance and tolerance in pathogenic bacteria. FEMS Microbiol. Rev. 2017, 41, 276–301. [Google Scholar] [CrossRef] [PubMed]
- Woo, S.-G.; Lee, S.-Y.; Lee, S.-M.; Lim, K.-H.; Ha, E.-J.; Eom, Y.-B. Activity of novel inhibitors of Staphylococcus aureus biofilms. Folia Microbiol. 2017, 62, 157–167. [Google Scholar] [CrossRef] [PubMed]
- McAuliffe, L.; Ayling, R.D.; Ellis, R.J.; Nicholas, R.A. Biofilm-grown Mycoplasma mycoides subsp. mycoides SC exhibit both phenotypic and genotypic variation compared with planktonic cells. Vet. Microbiol. 2008, 129, 315–324. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, S.-C.; Park, Y.; Hahm, K.-S. The role of antimicrobial peptides in preventing multidrug-resistant bacterial infections and biofilm formation. Int. J. Mol. Sci. 2011, 12, 5971–5992. [Google Scholar] [CrossRef] [Green Version]
- Chung, P.Y.; Khanum, R. Antimicrobial peptides as potential anti-biofilm agents against multidrug-resistant bacteria. J. Microbiol. Immunol. Infect. 2017, 50, 405–410. [Google Scholar] [CrossRef]
- Zhou, X.; Ma, C.; Zhou, M.; Zhang, Y.; Xi, X.; Zhong, R.; Chen, T.; Shaw, C.; Wang, L. Pharmacological Effects of Two Novel Bombesin-Like Peptides from the Skin Secretions of Chinese Piebald Odorous Frog (Odorrana schmackeri) and European Edible Frog (Pelophylax kl. esculentus) on Smooth Muscle. Molecules 2017, 22, 1798. [Google Scholar] [CrossRef] [Green Version]
- Nair, S.; Desai, S.; Poonacha, N.; Vipra, A.; Sharma, U. Antibiofilm Activity and Synergistic Inhibition of Staphylococcus aureus Biofilms by Bactericidal Protein P128 in Combination with Antibiotics. Antimicrob Agents Chemother. 2016, 60, 7280–7289. [Google Scholar]
- Wang, B.; Navath, R.S.; Menjoge, A.R.; Balakrishnan, B.; Bellair, R.; Dai, H.; Romero, R.; Kannan, S.; Kannan, R.M. Inhibition of bacterial growth and intramniotic infection in a guinea pig model of chorioamnionitis using PAMAM dendrimers. Int. J. Pharm. 2010, 395, 298–308. [Google Scholar] [CrossRef] [Green Version]
- Yan, J.; Wang, K.; Dang, W.; Chen, R.; Xie, J.; Zhang, B.; Song, J.; Wang, R. Two hits are better than one: Membrane-active and DNA binding-related double-action mechanism of NK-18, a novel antimicrobial peptide derived from mammalian NK-lysin. Antimicrob. Agents Chemother. 2013, 57, 220–228. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pag, U.; Oedenkoven, M.; Papo, N.; Oren, Z.; Shai, Y.; Sahl, H.-G. In vitro activity and mode of action of diastereomeric antimicrobial peptides against bacterial clinical isolates. J. Antimicrob. Chemother. 2004, 53, 230–239. [Google Scholar] [CrossRef] [Green Version]
- Fan, X.; Yang, F.; Nie, C.; Yang, Y.; Ji, H.; He, C.; Cheng, C.; Zhao, C. Mussel-Inspired Synthesis of NIR-Responsive and Biocompatible Ag–Graphene 2D Nanoagents for Versatile Bacterial Disinfections. ACS Appl. Mater. Interfaces 2018, 10, 296–307. [Google Scholar] [CrossRef] [PubMed]
- Desbois, A.P.; Coote, P.J. Wax moth larva (Galleria mellonella): An in vivo model for assessing the efficacy of antistaphylococcal agents. J. Antimicrob. Chemother. 2011, 66, 1785–1790. [Google Scholar] [CrossRef] [PubMed] [Green Version]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Peptide | Sequence | H | µH | Net Charge | α-Helicity (%) |
|---|---|---|---|---|---|
| Brevinin-1OS | FLPGILKVAANVVPGVICAITKKC | 0.734 | 0.386 | +3 | 94.61 |
| OSa | FLPGILKVAANVVPGVI | 0.832 | 0.457 | +1 | 29.6 |
| OSb | FLPGILKVAAK-NH2 | 0.688 | 0.527 | +3 | 27.5 |
| OSc | FLPKILKVAAK-NH2 | 0.598 | 0.587 | +4 | 26.9 |
| OSd | FLKWALKVAAK-NH2 | 0.602 | 0.439 | +4 | 51.66 |
| OSe | FLKWALKWAAK-NH2 | 0.695 | 0.491 | +4 | 39.09 |
| OSf | FLPKWALKWAAK-NH2 | 0.697 | 0.370 | +4 | 34.8 |
| Microorganisms | MICs 1/MBCs 2 (µM) | |||||||
|---|---|---|---|---|---|---|---|---|
| Brevinin-1OS | OSa | OSb | OSc | OSd | OSe | OSf | ||
| Gram-positive | S. aureus | 4/16 | >512/>512 | 256/>512 | 256/>512 | 16/32 | 4/4 | 8/16 |
| MRSA | 32/32 | >512/>512 | >512/>512 | >512/>512 | 32/32 | 8/8 | 16/64 | |
| E. faecalis | 8/16 | >512/>512 | >512/>512 | >512/>512 | 32/64 | 16/16 | 32/64 | |
| Yeast | C. albicans | 8/64 | >512/>512 | 512/>512 | 256/>512 | 16/32 | 4/4 | 8/16 |
| Gam-negative | E. coli | 128/>512 | >512/>512 | >512/>512 | >512/>512 | 32/32 | 16/16 | 8/16 |
| P. aeruginosa | 256/>512 | >512/>512 | >512/>512 | >512/>512 | 64/64 | 16/16 | 64/64 | |
| HC50 3 | 145.8 | >512 | 381.4 | >512 | >512 | 64.03 | 328.3 | |
| TI 4 (Overall) | 5.74 | 1 | 0.53 | 1.59 | 35.92 | 7.13 | 20.52 | |
| TI (Gram-positive and yeast) | 15.33 | 1 | 0.63 | 2 | 45.24 | 9.51 | 21.61 | |
| Microorganisms | MBIC50 1/MBEC50 2 (µM) | MBIC90 3/MBEC90 4 (µM) | ||||||
|---|---|---|---|---|---|---|---|---|
| Brevinin-1OS | OSd | OSe | OSf | Brevinin-1OS | OSd | OSe | OSf | |
| S. aureus | 4/32 | 8/32 | 4/32 | 8/32 | 4/64 | 32/128 | 8/32 | 32/128 |
| MRSA | 8/64 | 32/32 | 8/32 | 16/32 | 32/256 | 64/256 | 16/32 | 64/128 |
| E. faecalis | 32/256 | 64/128 | 64/128 | 32/128 | 64/>512 | 128/256 | 64/128 | 64/256 |
| E. coli | 128/>512 | 64/>512 | 16/>512 | 32/>512 | 128/>512 | 128/>512 | 32/>512 | 128/>512 |
| P. aeruginosa | 256/512 | 128/>512 | 64/>512 | 64/>512 | 256/512 | 256/>512 | 128/>512 | 256/>512 |
| Peptides | Microorganisms | MIC (µM) | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| NaCl (mM) | MgCl2 (mM) | pH | Temperature (°C) | Serum (10%) | ||||||||||
| 50 | 100 | 150 | 0.5 | 1 | 2 | 6 | 7 | 8 | 100 | 75 | 50 | |||
| Brevinin-1OS | S. aureus | 8 | 8 | 8 | 4 | 8 | 8 | 8 | 4 | 4 | 4 | 4 | 4 | 4 |
| MRSA | 8 | 16 | 32 | 16 | 16 | 16 | 64 | 8 | 4 | 16 | 16 | 16 | 8 | |
| E. faecalis | 32 | 32 | 64 | 32 | 32 | 32 | 128 | 32 | 16 | 16 | 16 | 16 | 16 | |
| C. albicans | 32 | 32 | 32 | 16 | 16 | 16 | 64 | 16 | 16 | 16 | 16 | 16 | 8 | |
| E. coli | 64 | 64 | 64 | 128 | 128 | 128 | 32 | 32 | 128 | 64 | 64 | 64 | 256 | |
| P. aeruginosa | >512 | >512 | >512 | >512 | >512 | >512 | 128 | 256 | 256 | 256 | 256 | 256 | >512 | |
| OSd | S. aureus | 128 | 256 | 256 | 128 | 128 | 256 | 64 | 32 | 32 | 32 | 32 | 32 | 128 |
| MRSA | 512 | 512 | 512 | 512 | 512 | 512 | 256 | 128 | 64 | 128 | 64 | 64 | 128 | |
| E. faecalis | 512 | 512 | 512 | 512 | 512 | 512 | 256 | 128 | 128 | 256 | 256 | 256 | >512 | |
| C. albicans | 128 | 128 | 256 | 64 | 64 | 128 | 64 | 32 | 32 | 128 | 32 | 32 | 64 | |
| E. coli | 256 | 256 | 512 | 128 | 256 | 512 | 64 | 64 | 64 | 256 | 64 | 64 | 128 | |
| P. aeruginosa | 256 | 512 | 512 | 256 | 256 | 256 | 128 | 128 | 64 | 128 | 64 | 64 | 256 | |
| OSe | S. aureus | 16 | 16 | 16 | 16 | 16 | 16 | 16 | 8 | 4 | 8 | 8 | 8 | 32 |
| MRSA | 16 | 16 | 32 | 16 | 16 | 32 | 64 | 16 | 8 | 16 | 16 | 16 | 64 | |
| E. faecalis | 64 | 64 | 64 | 64 | 64 | 64 | 32 | 64 | 32 | 64 | 64 | 64 | 128 | |
| C. albicans | 16 | 16 | 16 | 8 | 8 | 16 | 16 | 8 | 4 | 8 | 8 | 8 | 16 | |
| E. coli | 32 | 32 | 32 | 16 | 64 | 64 | 32 | 16 | 16 | 16 | 16 | 16 | 64 | |
| P. aeruginosa | 64 | 64 | 128 | 64 | 64 | 128 | 32 | 64 | 32 | 32 | 32 | 32 | 128 | |
| OSf | S. aureus | 64 | 64 | 64 | 64 | 64 | 64 | 128 | 8 | 16 | 8 | 8 | 8 | 32 |
| MRSA | 128 | 256 | 256 | 128 | 128 | 256 | 512 | 16 | 32 | 32 | 16 | 16 | 64 | |
| E. faecalis | 256 | 256 | 512 | 256 | 256 | 256 | 512 | 32 | 128 | 32 | 32 | 32 | 128 | |
| C. albicans | 64 | 64 | 128 | 32 | 64 | 64 | 128 | 8 | 16 | 16 | 8 | 8 | 32 | |
| E. coli | 64 | 128 | 128 | 128 | 128 | 256 | 128 | 8 | 32 | 16 | 8 | 8 | 32 | |
| P. aeruginosa | 128 | 256 | 256 | 128 | 256 | 256 | 128 | 256 | 64 | 64 | 64 | 64 | 64 | |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhou, X.; Liu, Y.; Gao, Y.; Wang, Y.; Xia, Q.; Zhong, R.; Ma, C.; Zhou, M.; Xi, X.; Shaw, C.; et al. Enhanced Antimicrobial Activity of N-Terminal Derivatives of a Novel Brevinin-1 Peptide from The Skin Secretion of Odorrana schmackeri. Toxins 2020, 12, 484. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins12080484
Zhou X, Liu Y, Gao Y, Wang Y, Xia Q, Zhong R, Ma C, Zhou M, Xi X, Shaw C, et al. Enhanced Antimicrobial Activity of N-Terminal Derivatives of a Novel Brevinin-1 Peptide from The Skin Secretion of Odorrana schmackeri. Toxins. 2020; 12(8):484. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins12080484
Chicago/Turabian StyleZhou, Xiaowei, Yue Liu, Yitian Gao, Yuanxing Wang, Qiang Xia, Ruimin Zhong, Chengbang Ma, Mei Zhou, Xinping Xi, Chris Shaw, and et al. 2020. "Enhanced Antimicrobial Activity of N-Terminal Derivatives of a Novel Brevinin-1 Peptide from The Skin Secretion of Odorrana schmackeri" Toxins 12, no. 8: 484. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins12080484