Purification, Toxicity and Functional Characterization of a New Proteinaceous Mussel Biotoxin from Bizerte Lagoon

 ,
,  , and
, and 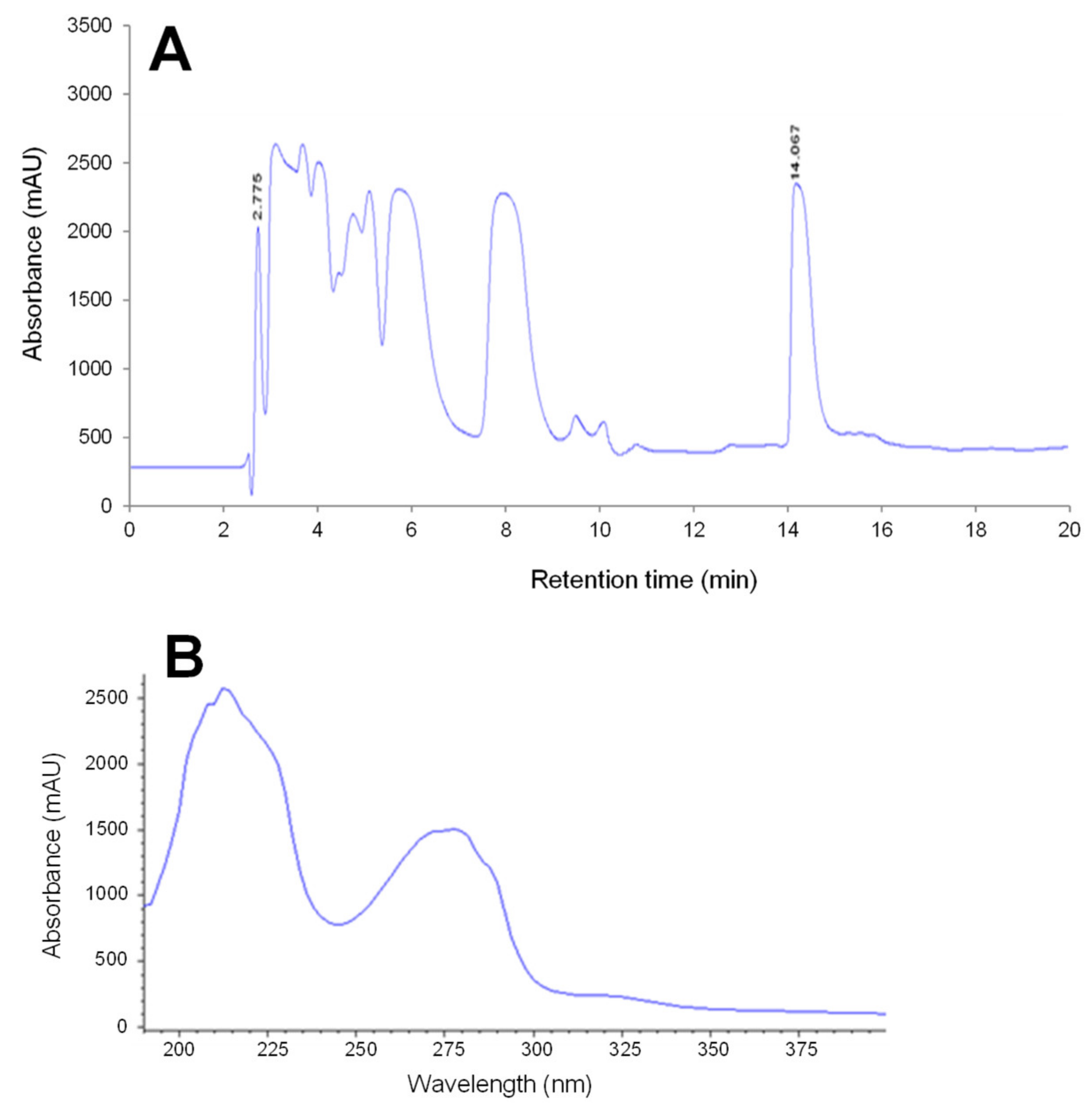{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Screening of Typical Toxins
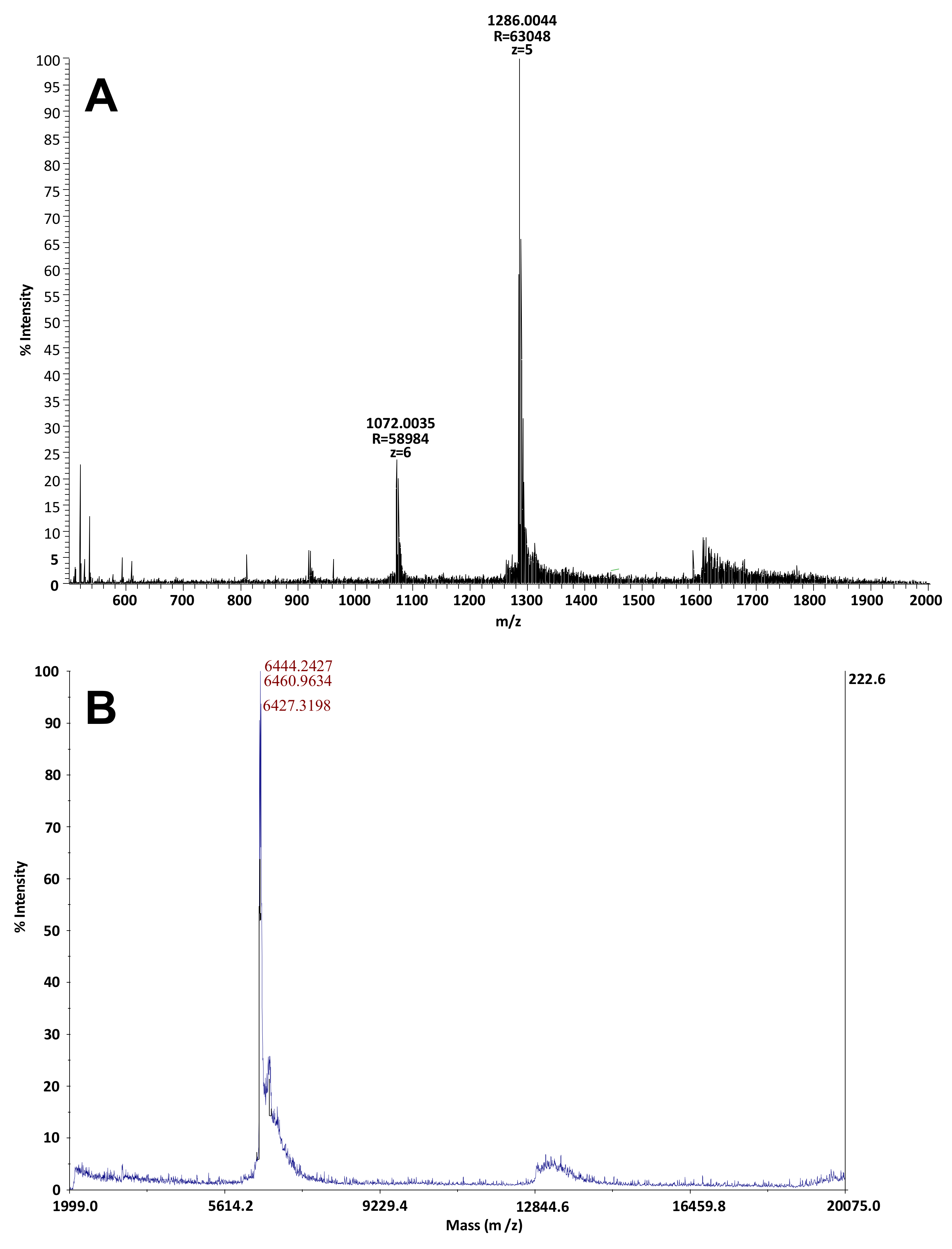
2.2. Identification and Characterization of the Toxic Fractions Responsible for Atypical Toxicity
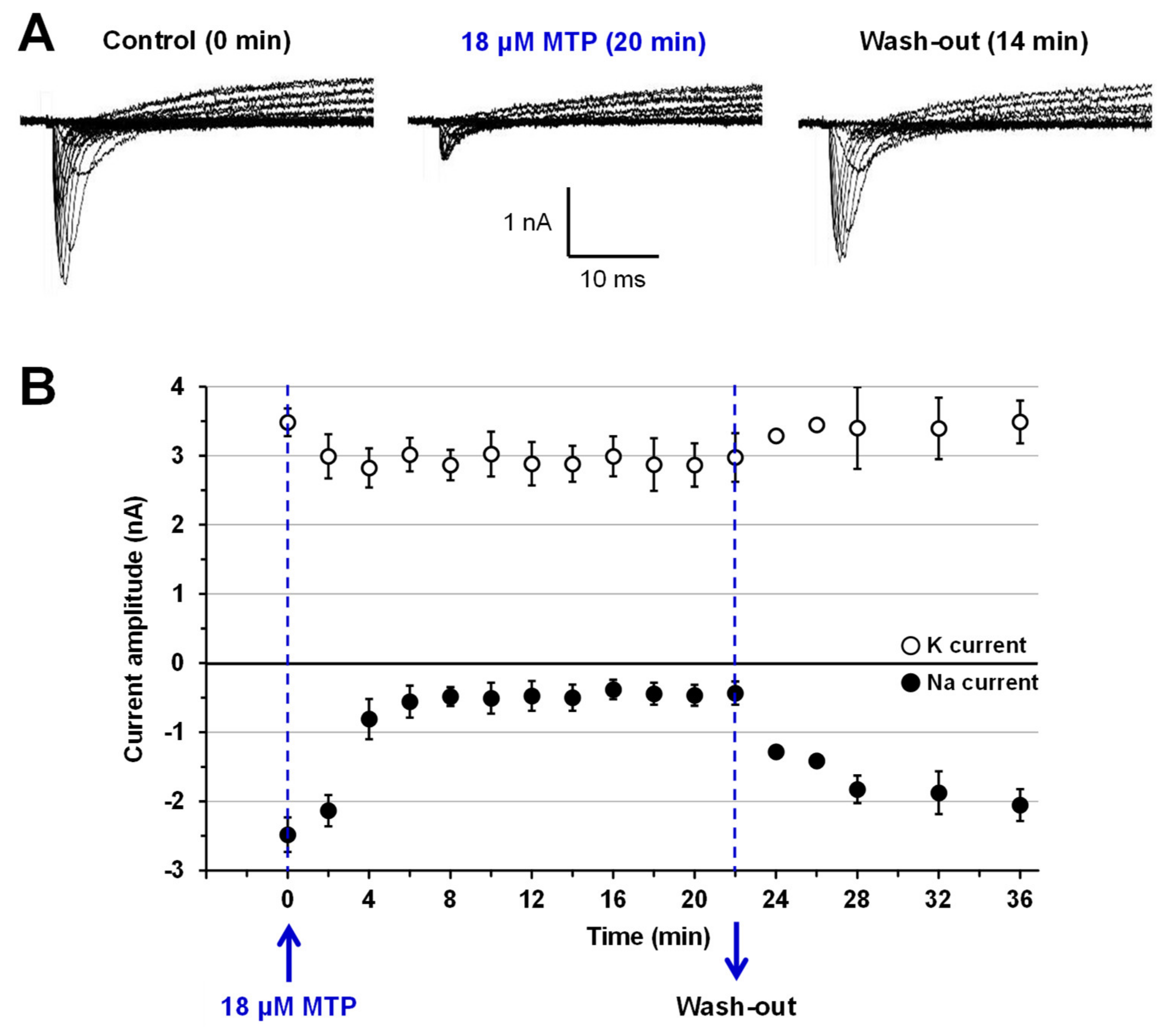
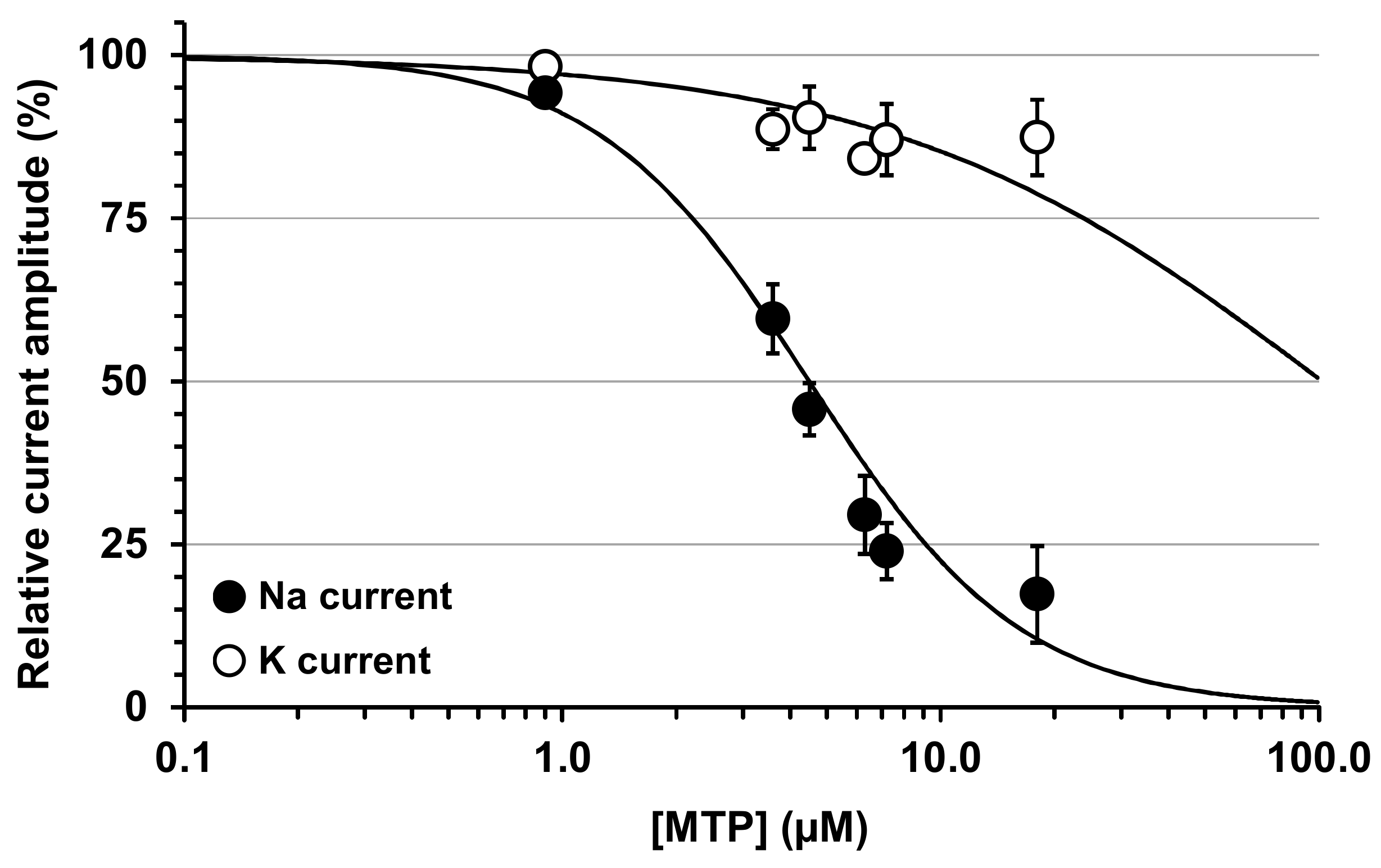
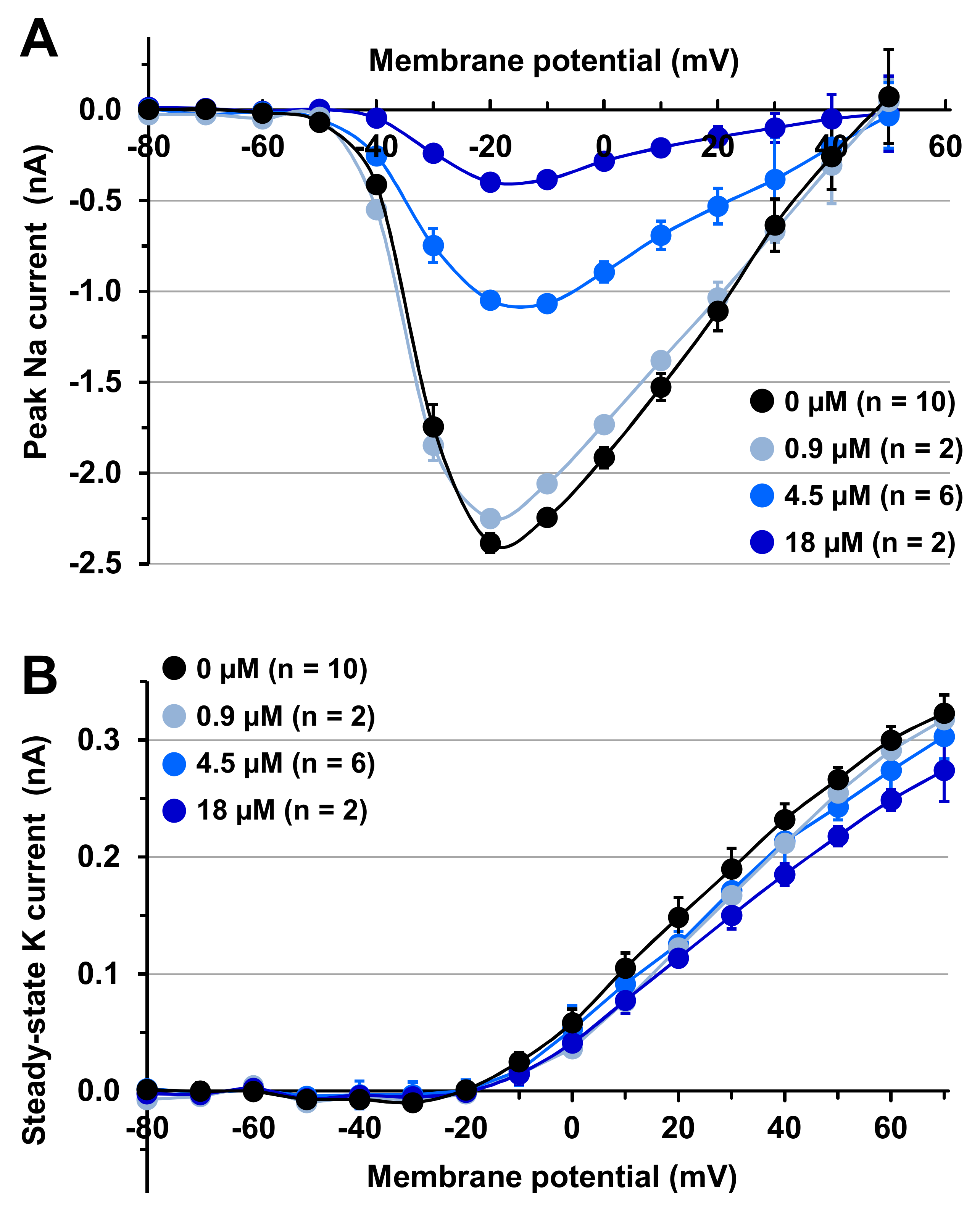
2.3. Effects of MTP on Mouse N18 Neuroblastoma Cells
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Ethical Approval
5.2. Mussels
5.3. Products
5.4. Toxicity Screening
5.5. Toxic Samples Extraction and Solvent Partition
5.6. Toxicity Assays
5.7. Purification Procedure and Bioguided Identification of Toxic Fractions
5.8. Characterization of the Toxic Fraction
Author Contributions
Funding
Conflicts of Interest
References
- Chateau-Degat, M.L. Les toxines marines: Problèmes de santé en émergence. VertigO-la revue électronique en sciences de l’environnemen 2003, 4, 1. [Google Scholar] [CrossRef]
- FAO. La Situation Mondiale des pêches et de l’aquaculture; FAO: Rome, Italy, 2012; p. 241. [Google Scholar]
- Hallegraeff, G.M. A review of harmful algal blooms and their apparent global increase. Phycologia 1993, 32, 79–99. [Google Scholar] [CrossRef] [Green Version]
- Murk, T.J.; Nicholas, J.; Smulders, F.J.M.; Bürk, C.; Gerssen, A. Marine biotoxins: Types of poisoning, underlying mechanisms of action and risk management programs. In Food Safety Assurance and Veterinary Public Health; Chemical hazards in foods of animal origin; Smulders, F.J.M., Rietjens, I.M.C.M., Rose, M.D., Eds.; Wageningen Academic Publishers: Wageningen, The Netherlands, 2019; Volume 7, pp. 207–223. [Google Scholar]
- Stivala, C.E.; Benoit, E.; Aráoz, R.; Servent, D.; Novikov, A.; Molgó, J.; Zakarian, A. Synthesis and biology of cyclic imine toxins, an emerging class of potent, globally distributed marine toxins. Nat. Prod. Rep. 2015, 32, 411–435. [Google Scholar] [CrossRef] [PubMed]
- Moreiras, G.; Leão, J.M.; Gago-Martínez, A. Analysis of Cyclic Imines in Mussels (Mytilus galloprovincialis) from Galicia (NW Spain) by LC-MS/MS. Int. J. Environ. Res. Public Health 2020, 17, 281. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kohlmeyer, J.; Volkmann-Kohlmeyer, B. Illustrated key of the filamentous higher marine fungi. Bot. Mar. 1991, 34, 1–61. [Google Scholar] [CrossRef]
- Abd-Elaah, G.A. The occurrence of fungi along the Red Sea coast and variability among isolates of Fusarium as revealed by isozyme analysis. J. Basic Microbiol. 1998, 38, 303–311. [Google Scholar] [CrossRef]
- Raheriniaina, C.E.; Maherizo, G.T.F.; Rabary, A.J.M.; Ramanantsoa, H.J.D.; Fitahia, E.M.; Solondrainy, T.J.; Rafenomanajara, T.H. Des souches des microfonges marines identifiées dans quelques sites des côtes sud ouest de Madagascar. Madarevues Actes Symp. Biomad 2009, V0, 9. [Google Scholar]
- Hallegraeff, G.; Coman, F.; Davies, C.; Hayashi, A.; McLeod, D.; Slotwinski, A.; Whittock, L.; Richardson, A.J. Australian Dust Storm Associated with Extensive Aspergillus sydowii Fungal “Bloom” in Coastal Waters. Appl. Environ. Microbiol. 2014, 80, 3315–3320. [Google Scholar] [CrossRef] [Green Version]
- Smith, G.W.; Ives, L.D.; Nagelkerken, I.A.; Richie, K.B. Caribbean sea-fan mortalities. Nature 1996, 383, 487. [Google Scholar] [CrossRef]
- Poirier, L.; Quiniou, F.; Ruiz, N.; Montagu, M.; Amiard, J.C.; Pouchus, Y.F. Toxicity assessment of Peptaïbols and contaminated sediments on Crassostrea gigas embryos. Aquat. Toxicol. 2007, 83, 254–262. [Google Scholar] [CrossRef] [Green Version]
- Ruiz, N.; Petit, K.E.; Vansteelandt, M.; Kerzaon, I.; Baudet, J.; Amzil, Z.; Biard, J.F.; Grovel, O.; Pouchus, Y.F. Enhancement of domoic acid neurotoxicity on diptera larvae bioassay by marine fungal metabolites. Toxicon 2010, 55, 805–810. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sarmiento-Ramírez, J.M.; Abella, E.; Martín, M.P.; Tellería, M.T.; López-Jurado, L.F.; Marco, A.; Diéguez-Uribeondo, J. Fusarium solani is responsible for mass mortalities in nests of loggerhead sea turtle, Caretta caretta, in Boavista, Cape Verde. FEMS Microbiol. Lett. 2010, 312, 192–200. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sallenave, C.; Pouchus, Y.F.; Robiou du Pont, T.; Lassus, P.; Verbist, J.F. Toxigenic saprophytic fungi in marine shellfish farming areas. Mycopathologia 2000, 149, 21–25. [Google Scholar] [CrossRef] [PubMed]
- Sallenave, C.; Pouchus, Y.F.; Bardouil, M.; Lassus, P.; Roquebert, M.F.; Verbist, J.F. Bioaccumulation of mycotoxins by shellfish: Contamination of mussels by metabolites of a Trichoderma koningii strain isolated in the marine environment. Toxicon 1999, 37, 77–83. [Google Scholar] [CrossRef]
- Grovel, O.; Pouchus, Y.F.; Verbist, J.F. Accumulation of gliotoxin, a cytotoxic mycotoxin from Aspergillus fumigatus, in blue mussel (Mytilus edulis). Toxicon 2003, 42, 297–300. [Google Scholar] [CrossRef]
- Hayashi, A.; José Dorantes-Aranda, J.; Bowman, J.P.; Hallegraeff, G. Combined Cytotoxicity of the Phycotoxin Okadaic Acid and Mycotoxins on Intestinal and Neuroblastoma Human Cell Models. Toxins 2018, 10, 526. [Google Scholar] [CrossRef] [Green Version]
- Matallah-Boutiba, A.; Ruiz, N.; Sallenave-Namont, C.; Grovel, O.; Amiard, J.C.C.; Pouchus, Y.F.; Boutiba, Z. Screening for toxigenic marine-derived fungi in Algerian mussels and their immediate environment. Aquaculture 2012, 342–343, 75–79. [Google Scholar] [CrossRef]
- Greco, G.; Cecchi, G.; Di Piazza, S.; Cutroneo, L.; Capello, M.; Zotti, M. Fungal characterisation of a contaminated marine environment: The case of the Port of Genoa (North-Western Italy). Webbia 2018, 7792, 1–10. [Google Scholar] [CrossRef]
- Marrouchi, R.; Benoit, E.; Le Caer, J.P.; Belayouni, N.; Belghith, H.; Molgó, J.; Kharrat, R. Toxic C17-Sphinganine Analogue Mycotoxin, contaminating Tunisian mussels, causes flaccid paralysis in rodents. Mar. Drugs. 2013, 11, 4724–4740. [Google Scholar] [CrossRef]
- AOAC. Paralytic Shellfish Poison: Biological method. Final action. In Official Method of Analysis, 15th ed.; Hellrich, K., Ed.; Association of Official Analytical Chemists (AOAC): Arlington, VA, USA, 1990; pp. 881–882. [Google Scholar]
- Yasumoto, T.; Oshima, Y.; Yamaguchi, M. Occurrence of a new type shellfish poisoning in the Tohoku district. Bull. Jpn. Soc. Sci. Fish. 1978, 44, 1249–1255. [Google Scholar] [CrossRef]
- Hille, B. Ion Channels of Excitable Membranes, 3rd ed.; Sinauer Associates Inc.: Sunderland, UK, 2001. [Google Scholar]
- Romdhane, M.S.; Eilersten, H.C.; Kefi-Dali Yahia, O.; Daly Yahia, M.N. Toxic dinoflagellate blooms in Tunisia lagoons: Causes and consequences for aquaculture. In Harmful algae; Reguera, B., Blanco, J., Fernández, M.L., Wyatt, T., Eds.; Xunta de Galicia and Intergovernmental Oceanographic Commission of UNESCO: Vigo, Spain, 1998; pp. 80–83. [Google Scholar]
- Biré, R.; Krys, S.; Fremy, J.M.; Dragacci, S.; Stirling, D.; Kharrat, R. First evidence on occurrence of gymnodimine in clams from Tunisia. J. Nat. Toxins 2002, 11, 269–275. [Google Scholar] [PubMed]
- Drira, Z.; Hamza, A.; Belhassen, M.; Ayadi, H.; Bouaïn, A.; Aleya, L. Dynamics of dinoflagellates and environmental factors during the summer in the Gulf of Gabes (Tunisia, Eastern Mediterranean Sea). Sci. Mar. 2008, 72, 59–71. [Google Scholar]
- Kharrat, R.; Servent, D.; Girard, E.; Ouanounou, G.; Amar, M.; Marrouchi, R.; Benoit, E.; Molgó, J. The marine phycotoxin gymnodimine targets muscular and neuronal nicotinic acetylcholine receptor subtypes with high affinity. J. Neurochem. 2008, 21, 952–963. [Google Scholar] [CrossRef] [PubMed]
- Marrouchi, R.; Rome, G.; Kharrat, R.; Molgó, J.; Benoit, E. Analysis of the action of gymnodimine-A and 13-desmethyl spirolide C on the mouse neuromuscular system in vivo. Toxicon 2013, 75, 27–34. [Google Scholar] [CrossRef] [PubMed]
- Sedmak, J.J.; Grossberg, S.E. A rapid, sensitive, and versatile assay for protein using Coomassie brilliant blue G250. Anal. Biochem. 1977, 79, 544–552. [Google Scholar] [CrossRef]
- Farabegoli, F.; Blanco, L.; Rodríguez, L.P.; Vieites, J.M.; Cabado, A.G. Phycotoxins in marine shellfish: Origin, occurrence and effects on humans. Mar. Drugs 2018, 16, 188. [Google Scholar] [CrossRef] [Green Version]
- Chugh, J.K.; Wallace, B.A. Peptaibols: Models for ion channels. Biochem. Soc. Trans. 2001, 29, 565–570. [Google Scholar] [CrossRef]
- Degenkolb, T.; Berg, A.; Gams, W.; Schlegel, B.; Gräfe, U. The occurrence of peptaibols and structurally related peptaibiotics in fungi and their mass spectrometric identification via diagnostic fragment ions. J. Pept. Sci. 2003, 9, 666–678. [Google Scholar] [CrossRef]
- Whitmore, L.; Wallace, B.A. The peptaibol database: A database for sequences and structures of naturally occurring peptaibols. Nucleic Acids Res. 2004, 32, 593–594. [Google Scholar] [CrossRef] [Green Version]
- Hammami, J.; Brahim, M.; Gueddari, M. Essai d’évaluation de la qualité des eaux de ruissellement du bassin versant de la lagune de BIZERTE. Bull. Inst. Nat. Sci. Technol. Mer Salammbô 2005, 32, 69–77. [Google Scholar]
- Cranford, P.; Anderson, R.; Archambault, P.; Balch, T.; Bates, S.; Bugden, G.; Callier, M.D.; Carver, C.; Comeau, L.; Hargrave, B.; et al. Indicators and thresholds for use in assessing shellfish aquaculture impacts on fish habitat. DFO Can. Sci. Advis. Secr. Res. Doc. 2006, 034, 166. [Google Scholar]
- Harvell, C.D.; Kim, K.; Burkholder, J.M.; Colwell, R.R.; Epstein, P.R.; Grimes, D.J.; Hofmann, E.E.; Lipp, E.K.; Osterhaus, A.D.; Overstreet, R.M.; et al. Emerging marine diseases—Climate links and anthropogenic factors. Science 1999, 285, 1505–1510. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kodama, M. Paralytic shellfish poisoning Toxins; biochemistry and origin. Aqua-Biosci. Monogr. 2010, 3, 1–38. [Google Scholar] [CrossRef] [Green Version]
- Lowry, O.H.; Rosebrough, N.J.; Farr, A.L.; Randall, R.J. Protein measurements with Folin-phenol reagent. J. Biol. Chem. 1951, 193, 265–275. [Google Scholar]
- Gonçalves, T.C.; Boukaiba, R.; Molgó, J.; Amar, M.; Partiseti, M.; Servent, D. Direct evidence for high affinity blockade of NaV1.6 channel subtype by huwentoxin-IV spider peptide, using multiscale functional approaches. Neuropharmacology 2018, 133, 404–414. [Google Scholar] [CrossRef]






© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Marrouchi, R.; Benoit, E.; Schlumberger, S.; Marzougui, Z.; Le Caer, J.-P.; Molgó, J.; Kharrat, R. Purification, Toxicity and Functional Characterization of a New Proteinaceous Mussel Biotoxin from Bizerte Lagoon. Toxins 2020, 12, 487. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins12080487
Marrouchi R, Benoit E, Schlumberger S, Marzougui Z, Le Caer J-P, Molgó J, Kharrat R. Purification, Toxicity and Functional Characterization of a New Proteinaceous Mussel Biotoxin from Bizerte Lagoon. Toxins. 2020; 12(8):487. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins12080487
Chicago/Turabian StyleMarrouchi, Riadh, Evelyne Benoit, Sébastien Schlumberger, Zeineb Marzougui, Jean-Pierre Le Caer, Jordi Molgó, and Riadh Kharrat. 2020. "Purification, Toxicity and Functional Characterization of a New Proteinaceous Mussel Biotoxin from Bizerte Lagoon" Toxins 12, no. 8: 487. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins12080487